Back to Journals » Drug Design, Development and Therapy » Volume 19
Luteolin’s Potential in Managing Osteoporosis and Bone Metabolism Disorders: Preclinical Insights
Authors Du J, Qin W ![]() , Wen F, Zhao D, Yin X, Guo Z, Feng Q, Gu E, Pan Z, Wang L
, Wen F, Zhao D, Yin X, Guo Z, Feng Q, Gu E, Pan Z, Wang L
Received 14 June 2025
Accepted for publication 11 October 2025
Published 29 October 2025 Volume 2025:19 Pages 9715—9732
DOI https://doi.org/10.2147/DDDT.S547141
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Muzammal Hussain
Jianqiang Du,1,2,* Wenxiu Qin,2,* Fayan Wen,3,* Dan Zhao,3 Xuesong Yin,3 Ziyu Guo,2 Qijing Feng,1 Enpeng Gu,1 Zhicheng Pan,4 Linjue Wang1
1Department of Orthopedics, The Second Affiliated Hospital of Tianjin University of Traditional Chinese Medicine, Tianjin, People’s Republic of China; 2Graduate School, Tianjin University of Traditional Chinese Medicine, Tianjin, People’s Republic of China; 3Graduate School, Gansu University of Traditional Chinese Medicine, Lanzhou, People’s Republic of China; 4Department of Orthopedics, Binhai New Area Hospital of Traditional Chinese Medicine and the Fourth Affiliated Hospital of Tianjin University of Traditional Chinese Medicine, Tianjin, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Linjue Wang, Email [email protected] Zhicheng Pan, Email [email protected]
Abstract: Bone homeostasis is a process of dynamic regulation between bone resorption and bone formation, and is the foundation for normal skeletal development in the human body. Once this balance is disrupted, it will lead to bone loss and decreased bone density, profoundly affecting the occurrence and development of bone metabolism-related diseases. For example, common clinical bone metabolic disorders such as osteoporosis, avascular necrosis of the femoral head, and alveolar bone resorption. However, current treatments for bone metabolism-related diseases often have certain limitations, which has driven the development of complementary medicine that helps maintain bone health. Luteolin is a natural flavonoid compound renowned for its exceptional medicinal value and is often used to treat various diseases. In vitro studies indicate that luteolin not only protects osteoblast activity and promotes their osteogenic differentiation, but also inhibits osteoclast activation and their bone resorption function. Simultaneously, it regulates the osteogenic-adipogenic differentiation capacity of bone marrow mesenchymal stem cells, thereby promoting bone formation. Furthermore, Furthermore, in vivo studies indicate that luteolin significantly enhances parameters such as bone mass, trabecular thickness, and density in animal models of bone metabolic disorders, thereby promoting bone remodeling. In terms of mechanisms, luteolin may regulate signaling pathways related to bone metabolism, such as the PI3K/Akt, Wnt, RANKL/RANK/OPG, Nrf2, ERK and JAK/STAT pathways, to inhibit oxidative stress, reduce inflammatory responses, regulate cell death, and modulate the immune microenvironment. Additionally, we explored the pharmacokinetics of luteolin and the current status of drug delivery systems, including luteolin-loaded nanoparticles, biomimetic hydrogels, and porous scaffolds. These systems have effectively achieved controlled release of luteolin, providing drug delivery solutions with clinical translation potential for bone defect repair. The above evidence provides a solid foundation for future applications in bone metabolic homeostasis management and treatment based on luteolin.
Keywords: luteolin, bone formation, bone resorption, cell model, animal model, molecular mechanism, pharmacokinetics
Introduction
Bone is a highly dynamic and metabolically active tissue, one of the largest organ systems in the human body, serving functions such as mechanical support, muscle attachment, and calcium and phosphorus storage.1,2 Bone metabolism is a dynamic process of continuous renewal and repair within the skeletal system. At its core lies the coordinated interaction between bone resorption and bone formation. The dynamic equilibrium maintained between these two processes constitutes bone homeostasis, which is crucial for ensuring stable bone mass, structural integrity, and normal physiological function of the skeleton. Bone resorption is primarily driven by osteoclasts.3 After attaching to the bone surface, osteoclasts secrete acidic substances to dissolve bone minerals and enzymes to break down the bone matrix, thereby removing old bone tissue. Bone formation is carried out by osteoblasts,4 which first synthesize and secrete substances like collagen to construct the bone matrix. Subsequently, this matrix undergoes mineralization, gradually forming new bone tissue. Maintaining homeostasis relies on multiple factors, including hormones, vitamin D, mechanical stimuli, and cytokines. Excessive resorption or insufficient formation disrupts this balance, leading to bone loss. Thus, skeletal health depends on the precise regulation of metabolic processes and equilibrium between internal and external environments.5
Imbalance in bone metabolism homeostasis is one of the fundamental causes of osteoporosis (OP) and osteonecrosis of the femoral head (ONFH) and other metabolic bone diseases. OP, as a classic metabolic bone disease, is characterized by low bone mass and destruction of bone microstructure, resulting in significantly increased bone fragility and a high risk of fractures.6 It has been reported that the prevalence of OP worldwide is approximately one-fifth of the population.7 Drug therapy plays a key role in the prevention and treatment of OP. Currently, commonly used drug options include bone nutrition supplements, bone resorption inhibitors, and bone formation promoters (Table 1). However, some of these drugs may cause serious side effects during long-term use, often limiting their clinical application. ONFH is a refractory orthopedic disease characterized by microscopic structural damage to the femoral head, collapse of the femoral head, and resulting hip joint pain.8 Currently, approximately 150,000 to 200,000 new cases of ONFH are diagnosed annually in China.9 In the late stages of ONFH, surgery has become the primary treatment method, such as total hip arthroplasty. However, artificial joints have a limited lifespan, and a second replacement may be necessary, and the trauma caused by the surgery itself is unavoidable. In addition, alveolar bone defects are a chronic disease process that can be caused by periodontal disease, oral and maxillofacial trauma, and other factors, leading to deformities, oral functional impairments, and significantly impacting patients’ quality of life.10 For severe alveolar bone defects, autologous bone transplantation is a commonly used treatment method, but it has limitations such as limited supply, potential deformities at the donor site, and risks of infection.11 Overall, the treatment of bone metabolic disorders currently faces certain challenges, making the development of effective and safe new therapeutic strategies urgently needed.
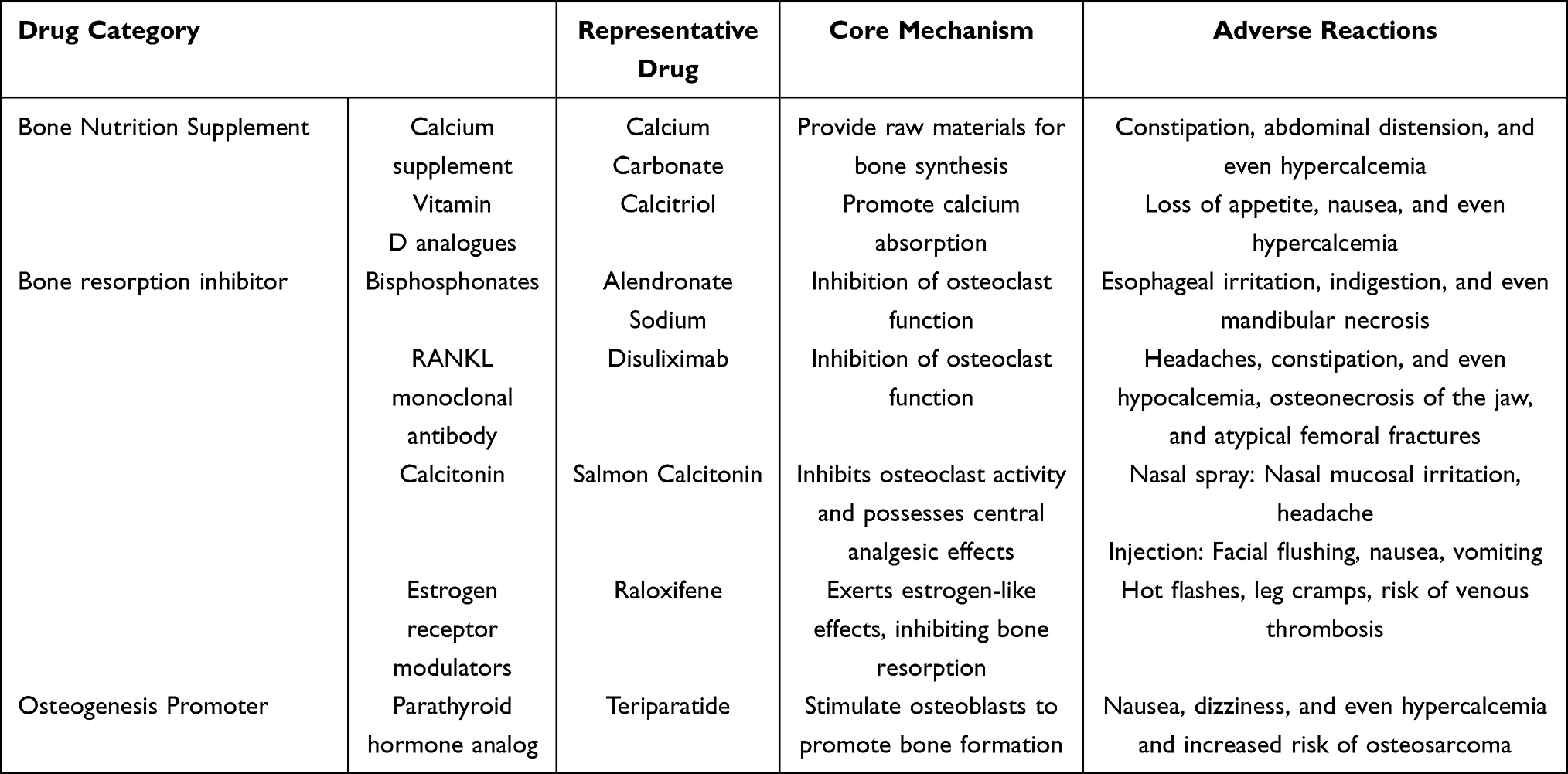 |
Table 1 Commonly Used Drugs for the Treatment of Osteoporosis |
Traditional herbal medicines have a long history of use in the prevention and treatment of diseases. In recent years, natural active compounds derived from medicinal plants have attracted significant attention from scholars due to their wide range of pharmacological properties.12,13 Luteolin is a natural flavonoid compound found in various medicinal plants and vegetables. It has a variety of pharmacological effects, including antitumor, antioxidant, anti-inflammatory, and neuroprotective properties,14–17 and is a widely occurring compound with medicinal value. Notably, as research on luteolin deepens, some of its medicinal properties are directly related to its protective effects on bone metabolic homeostasis. Specifically, it can regulate important signaling pathways, act on osteoclasts, osteoblasts, and bone marrow mesenchymal stem cells, and influence bone formation and bone resorption, which is of great significance for the bone remodeling process. In addition, as a readily available natural compound, luteolin has the characteristics of low cost and few adverse reactions, making it more suitable for long-term use than synthetic drugs. It is expected to become a potential treatment option for bone metabolic diseases in the future. However, existing cellular models struggle to replicate the complex microenvironment of the human body, and animal models exhibit differences from human pathological states. This discrepancy may create a gap between the preclinical findings of luteolin and its actual clinical application, necessitating further refinement of models to bridge this gap.
To comprehensively and systematically review existing preclinical evidence on luteolin’s regulation of bone metabolism, this review employed a multi-database search strategy encompassing PubMed, Web of Science, Google Scholar, China National Knowledge Infrastructure (CNKI), and Wanfang Data Knowledge Service Platform. Keywords included “luteolin”, “bone”, “bone metabolism”, “bone formation”, “bone loss”, “osteoblast”, “osteoclast”, and “bone marrow mesenchymal stem cells”. The search period was set from the inception of each database to May 2025. The literature type was focused on original research (cell experiments, animal studies), excluding review articles, clinical studies, conference abstracts, and literature with incomplete data or unavailable full texts.
Physicochemical Properties of Luteolin
Luteolin is widely distributed and found in traditional Chinese medicines such as Lonicera japonica, Chrysanthemum indicum, and Xiakucao, as well as in foods like celery, Chinese cabbage, cauliflower, and onions.18 It is a yellow crystalline substance with the chemical formula C15H10O6, a molecular weight of 286.24 g/mol, and a density of 1.652 g/cm3. Additionally, the melting point of luteolin is 330°C. Its chemical structure is shown in Figure 1. Its structural features include two aromatic rings (A and B) connected by a heterocyclic ring (C), with four hydroxyl groups at positions C5, C7, C3′, and C4′. The hydroxyl groups and the double bond between C2 and C3 may be closely related to the biological activity of luteolin.19 Structure-activity relationship studies have revealed that the potent antioxidant properties of luteolin are attributed to the hydroxyl groups at the C5, C7, C3′, and C4′ sites. Additionally, the carbonyl oxygen at the C4 site is responsible for the antibacterial properties of the compound. Similarly, bactericidal activity has been attributed to the double bond between C2 and C3.20 It is relatively stable at room temperature, slightly soluble in hot water, poorly soluble in cold water, weakly acidic, soluble in alkaline solutions, and highly soluble in organic solvents such as ethanol, methanol, dimethyl sulfoxide, and ether.21
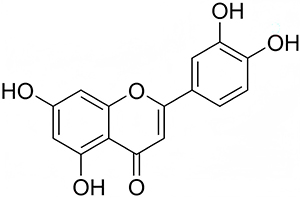 |
Figure 1 Structural formula of luteolin. |
In vitro Study of Luteolin Regulation of Bone Metabolism
Bone metabolism is closely related to the two processes of bone formation and bone resorption, which involve osteoblasts, osteoclasts, and bone marrow mesenchymal stem cells. The regulatory effect of luteolin on the above cells is its basic mechanism for maintaining bone metabolic homeostasis.22,23 Here, we summarize the available data on the intervention of luteolin in bone metabolism in vitro studies (Table 2).
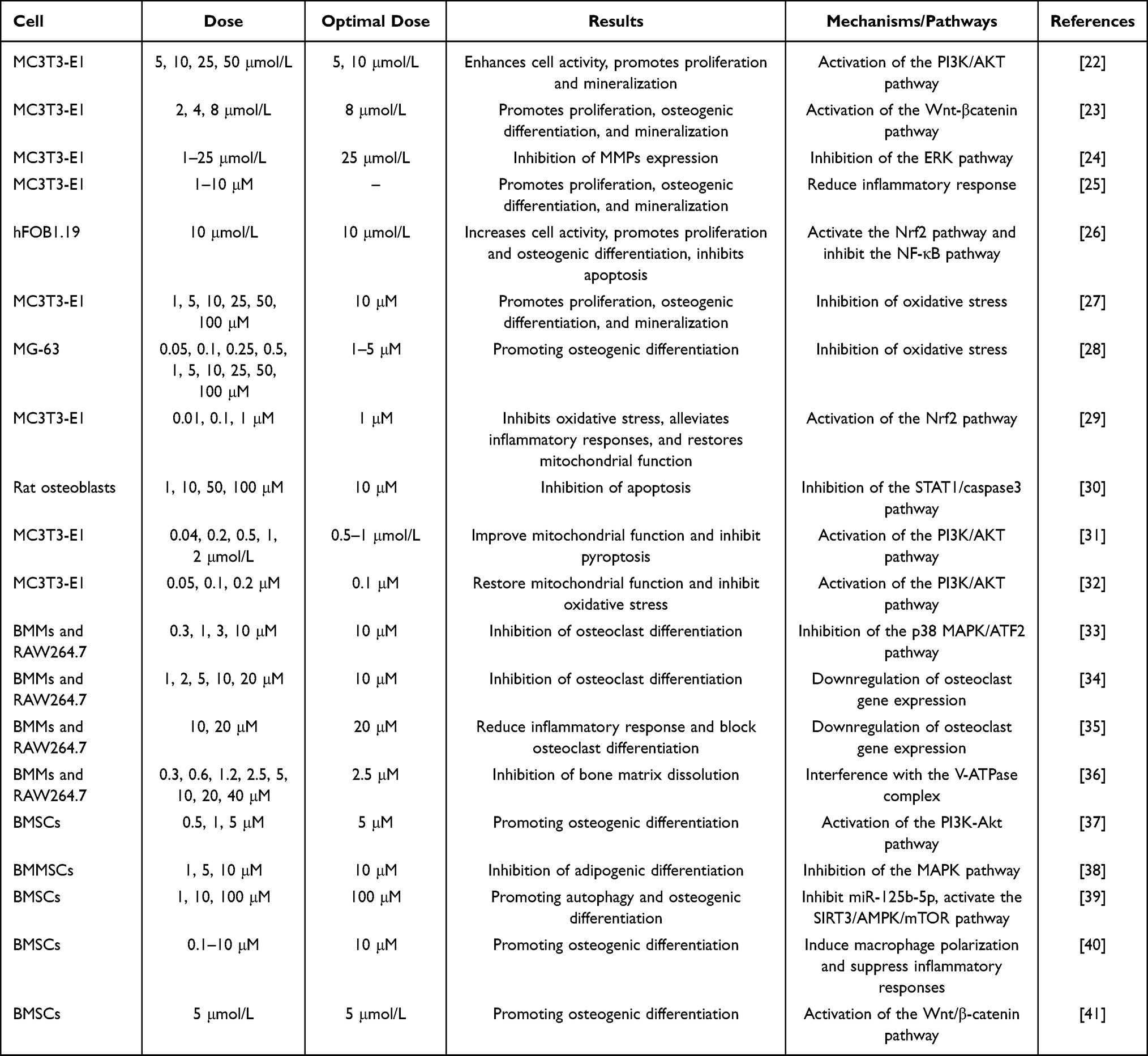 |
Table 2 Bone Metabolism Regulation of Luteolin in vitro Experiments |
Regulation of Osteoblasts
As the core functional cells of bone metabolism, osteoblasts play an indispensable role in maintaining the dynamic balance of bone homeostasis. With the continuous advancement of research, it has been confirmed that luteolin can promote bone formation by regulating the life activities of osteoblasts.
Previous studies have shown that inflammation is closely related to bone metabolic homeostasis. Cytokines such as IL-1β and TNF-α can induce osteoblasts to enhance the expression of matrix metalloproteinases (MMPs), which ultimately lead to bone destruction by regulating bone matrix degradation.42 Yang et al24 found that luteolin significantly inhibits the expression of MMP-9 and −13 in osteoblasts induced by IL-1β, thereby protecting against bone loss, and this effect is positively correlated with dose. Additionally, inflammatory mediators can induce the expression of inducible nitric oxide synthase (iNOS) in osteoblasts, and NO can lead to restricted bone formation.43 However, luteolin inhibits the expression of NO, prostaglandin E2 (PGE2), TNF-α, and IL-6 induced by 3-morpholino-N-nitrosourea (SIN-1) in osteoblasts, while significantly enhancing the alkaline phosphatase (ALP) activity and osteocalcin (OC) secretion in MC3T3-E1 osteoblasts.25 Notably, these effects can be blocked by the anti-estrogen drug tamoxifen, suggesting that luteolin may exert its effects through estrogen-mediated mechanisms, potentially positioning it as a natural selective estrogen receptor modulator. In fact, inflammation may also lead to increased levels of oxidative stress at the site of the lesion.44 Reactive oxygen species (ROS) can enhance the expression of pro-inflammatory genes by stimulating intracellular signaling cascades. This vicious cycle is often one of the main mechanisms promoting disease development.45 In Peng et al’s study,26 it was shown that luteolin not only inhibits the expression of IL-6, IL-8, and TNF-α but also reduces ROS levels and increases superoxide dismutase (SOD) activity, thereby protecting human osteoblasts (hFOB1.19) from oxidative damage and inflammatory stimulation. Subsequent scholars have also confirmed this finding.27 In summary, luteolin can regulate osteoblast function stimulated by inflammatory mediators, providing a reference for its application in the treatment of bone metabolic diseases.
In addition, diabetes is a major contributing factor to impaired bone formation or bone loss.46 One reason for this is that high blood sugar levels lead to a significant increase in ROS levels in the body, which in turn inhibits the expression of key transcription factors related to osteoblast differentiation.47 In a study28 investigating the regulation of bone formation by luteolin under high-glucose stress conditions, luteolin exhibited a clear biphasic response in MG-63 osteoblasts: low concentrations of luteolin protected human MG-63 osteoblast-like cells from hyperglycemic stress by inhibiting ROS and restoring ALP activity; in contrast, high concentrations of luteolin completely reversed these effects. Notably, the protective concentration of luteolin in this study was approximately 20–26 times lower than the toxic concentration, making it a safe bone metabolism regulator. Methylglyoxal (MG) is a highly reactive diketone metabolite produced during glucose metabolism and is the main precursor of advanced glycation end products (AGEs),48 which are involved in the pathogenesis of diabetes and inflammation. Many diseases are thought to be closely related to MG-derived AGEs, including diabetic bone metabolism disorders.49 Suh et al29 pointed out that luteolin can directly clear ROS and inhibit oxidative damage by promoting the nuclear translocation of nuclear factor erythroid 2-related factor 2 (Nrf2) and enhancing the expression of downstream antioxidant enzymes; simultaneously, it upregulates the level of peroxisome proliferator-activated receptor gamma coactivator 1α (PGC-1α) to maintain cellular mitochondrial homeostasis; additionally, luteolin reduces TNF-α secretion and sRAGE expression levels, blocking the AGE-RAGE-mediated inflammatory cascade, thereby alleviating MG-induced toxicity in osteoblasts MC3T3-E1 through multiple pathways. These findings provide experimental evidence for the potential application of luteolin in diabetic bone disease, but further preclinical validation is required.
An increasing number of studies have shown that long-term use of glucocorticoids can induce osteoblast apoptosis and increase osteoclast activation.50,51 It has been reported that dexamethasone (Dex) can induce apoptosis in mouse osteoblasts.52 However, luteolin can effectively inhibit Dex-induced apoptosis in rat osteoblasts by inhibiting the phosphorylation of transcription activator 1 (STAT1) and reducing the cleavage of caspase-9 and caspase-3, thereby downregulating the expression of B-cell lymphoma-2 (Bcl-2) and Bcl-2-associated X (Bax).30 STAT1 is a member of the STAT protein family and has pro-apoptotic effects.53 This suggests that luteolin may be a potential drug for treating glucocorticoid-induced bone metabolic bone disease. In addition, pyroptosis is closely related to bone metabolism. Under the influence of certain pathological factors, pyroptosis becomes overly activated, leading to the death of bone metabolism-related cells and mediating the occurrence of various bone metabolism-related diseases.54 A recent study31 showed that, in vitro, osteoblasts treated with luteolin can alleviate GSDME-mediated pyroptosis and maintain bone formation by improving mitochondrial dysfunction.
Mitochondrial dysfunction plays a key role in bone metabolism.55 cAMP response element-binding protein (CREB) is a transcription factor that is partially localized in mitochondria. CREB activation can be mediated by kinases such as Akt; however, mitochondrial dysfunction inhibits Akt activity, resulting in reduced CREB phosphorylation levels.56,57 According to Choi and his team’s report,32 luteolin can simultaneously enhance PI3K and Akt activity and CREB phosphorylation levels, restore mitochondrial function, and inhibit oxidative stress reactions, thereby protecting MC3T3-E1 osteoblasts from damage induced by the mitochondrial electron transport chain inhibitor Antimycin A (AMA). In addition, Kwon58 and his team found that luteolin stimulates osteoblast differentiation and simultaneously inhibits adipocyte differentiation by inducing heat shock protein (Hsp) 40 (Dnajb1), which is a potential strategy for preventing bone metabolic diseases. However, the underlying mechanism still needs to be further clarified.
Regulation of Osteoclasts
Osteoclasts are multinucleated cells formed by the fusion of differentiated mononuclear-macrophage cells, and they are the primary cells responsible for bone resorption in maintaining bone metabolic homeostasis within the body.59,60 Numerous bioactive factors, including RANKL, TNF-αand IL-1β can stimulate osteoclasts and induce their activation.61 Recent studies have shown that luteolin can selectively inhibit osteoclast activation and is used in metabolic bone diseases characterized by bone resorption.
In an earlier study, Lee et al33 observed that luteolin could dose-dependently inhibit the differentiation of bone marrow-derived macrophages (BMMs) and RAW264.7 cells stimulated by RANKL into osteoclasts. Mechanistically, luteolin blocks the osteoclast differentiation signaling pathway by inhibiting the phosphorylation of ATF2 downstream of p38 MAPK and the expression of NFATc1 protein, while disrupting the actin ring of mature osteoclasts to inhibit the formation of bone resorption pits. This dual mechanism provides a theoretical basis for luteolin’s intervention in osteoclast generation and activation.
According to Kim and his team’s report,34 luteolin reduces the number and activity of TRAP-positive multinucleated cells in a dose-dependent manner, while significantly downregulating the expression of osteoclast differentiation-related genes such as NFATc1, c-Src, DC-STAMP, and MMP-9, thereby inhibiting the differentiation of BMMs and Raw264.7 cells into osteoclasts. Shin et al35 observed in vitro experiments that flavonoids significantly inhibit the release of inflammatory factors induced by titanium particles in BMMs, and dose-dependently reduce the number of TRAP-positive multinucleated cells while downregulating the expression of osteoclast-related genes such as NFATc1 and c-fos, thereby blocking osteoclast differentiation and bone resorption activity. This is consistent with the results of Kim et al, confirming that flavonoids have a clear inhibitory effect on bone loss.
Additionally, studies have shown that bone resorption is partially achieved by recruiting V-ATPase to the plasma membrane of polarized, active osteoclasts.62 V-ATPase is composed of multiple subunits, among which the a3 and d2 subunits are highly specifically expressed in osteoclasts. Defects in the a3 or d2 genes can lead to impaired bone resorption.63,64 Therefore, interfering with the interaction between the a3 and d2 subunits may serve as a therapeutic target for inhibiting bone resorption. Recent experimental results indicate that luteolin can significantly reduce osteoclast bone resorption activity by specifically inhibiting the interaction between the V-ATPase a3 and d2 subunits, without affecting cell differentiation or survival.36 This provides a potential new target for developing luteolin-based anti-bone resorption therapies.
Regulation of Bone Marrow Mesenchymal Stem Cells
Bone marrow mesenchymal stem cells (BMSCs) have the ability to self-renew and differentiate into various cell types, such as osteoblasts and adipocytes.65 The differentiation process of BMSCs into osteoblasts and adipocytes is not independent but interrelated; an imbalance in osteogenic-adipogenic differentiation can disrupt bone metabolic homeostasis. Luteolin has been shown to increase the expression of type I collagen (Col I), osteopontin (OPN), and Runt-related transcription factor 2 (Runx2) in BMSCs, thereby promoting their osteogenic differentiation.37
Peroxisome proliferator-activated receptor (PPAR) gamma is one of the PPAR nuclear receptors and is a major regulator of fat production.66 PPARγ can inhibit the transcription and expression of Runx2, thereby hindering its osteogenic differentiation, while promoting massive differentiation into adipocytes, ultimately leading to bone metabolic diseases. Wu et al38 found that luteolin can dose-dependently downregulate the mRNA and protein expression of adipogenesis markers such as PPARγ, C/EBPα, and SREBP1c, thereby inhibiting the adipogenic differentiation of human bone marrow mesenchymal stem cells (hBM-MSCs). In addition, they also noted that the derivative luteolin-4′-sulfate sodium had a significantly weaker inhibitory effect on the above process than luteolin. One possible reason is that the replacement of 4′-hydroxy with sulfate sodium weakened the anti-adipogenic activity, indicating that the anti-adipogenic effect of luteolin is closely related to the 4′-hydroxy in its chemical structure. Overall, this study suggests that luteolin is a potential therapeutic option for bone metabolic diseases and provides a theoretical basis for its structural optimization.
In addition, targeting regulators of autophagy flux is very important in bone remodeling.67 Mir-125b is a key microRNA that significantly affects osteoblast differentiation by regulating different pathways, thereby promoting OP progression.68 Sirtuin 3 (SIRT3) is one of the most important targets of miRNA and exerts its effects by regulating the expression of autophagy-related genes, such as autophagy-related protein 5 (ATG5) and microtubule-associated protein 1A/1B light chain 3 (LC3), through the upstream mediators mTOR and protein kinase (AMPK) of the autophagy pathway.69,70 It has been reported that luteolin can activate autophagy and reverse the inhibitory effects of DEX on bone formation markers Runx2, OPN, and OCN by inhibiting miR-125b-5p, upregulating SIRT3 and AMPK, and downregulating mTOR, thereby promoting the osteogenic differentiation of BMSCs.39
Macrophages play multiple roles in the construction of the immune microenvironment, and their polarization from pro-inflammatory (M1) to anti-inflammatory/repairing (M2) is crucial for bone metabolism.71 Hu et al40 confirmed that luteolin can induce macrophages to polarize from M1 to M2 type, inhibit the expression of proinflammatory factors, and promote the secretion of anti-inflammatory factors, thereby promoting the osteogenic differentiation of BMSCs. Similarly, two recent studies have reached similar conclusions, demonstrating that luteolin can induce macrophage polarization, inhibit inflammatory responses, and promote osteogenic differentiation of BMSCs.72,73 These studies provide an innovative strategy for bone regeneration that combines osteogenic and bone immune regulatory functions.
In addition, previous studies have reported that BMSCs differentiated into osteoblasts can be used for alveolar bone regeneration.74 Luteolin may promote osteogenic differentiation of rat BMSCs by regulating β-catenin and Cyclin D1 expression, thereby increasing ALP activity.41 Thus, It is a potential inducer in the osteogenic differentiation process of BMSCs and a potential therapeutic agent for alveolar bone loss. According to reports, Cyclin D1 is an important molecule in the Wnt/β-catenin pathway. Activation of the Wnt/β-catenin pathway can induce Cyclin D1 expression, affecting osteocyte differentiation and bone remodeling.75
In vivo Study of the Regulatory Effect of Luteolin on Bone Metabolism
In the field of medical research, animal models play an important role in the study of disease prevention and treatment. Their significance lies in providing a critical experimental platform and bridge for understanding the mechanisms of human diseases and evaluating the safety and efficacy of potential therapeutic interventions. We have summarized the available data on the use of luteolin in animal models of bone metabolic diseases (Table 3).
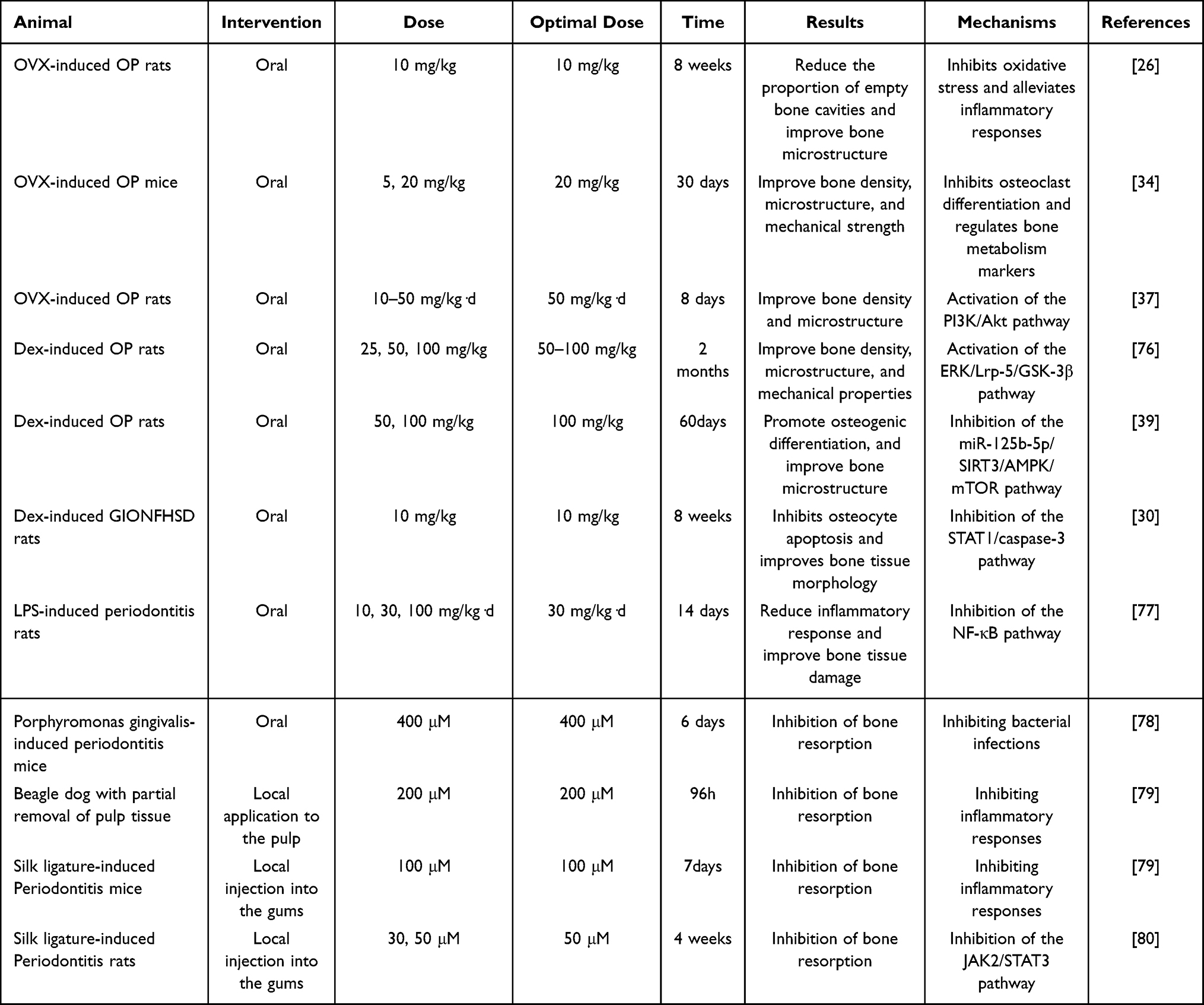 |
Table 3 Bone Metabolism Regulation by Luteolin in in vivo Experiments |
Bone Metabolism Regulation of Luteolin in OP Model
Postmenopausal osteoporosis (PMOP) is a metabolic bone disease characterized by bone loss, decreased bone density, and increased risk of fracture, caused by a decrease in estrogen levels secreted by the ovaries after menopause in women.81 The castration modeling method is a modeling method that simulates PMOP by reducing estrogen levels in mice through ovarian removal surgery (OVX).82 Chai et al26 established a PMOP mouse model via OVX surgery and administered continuous intervention with luteolin for 12 weeks. The results showed that luteolin could increase bone mineral density (BMD), trabecular number (Tb.N), and bone volume fraction (BV/TV) in mice, while reducing trabecular separation (Tb.Sp). Additionally, serum analysis indicated that luteolin could reduce malondialdehyde (MDA) levels and enhance SOD and glutathione peroxidase (GSH-Px) activity, thereby reversing OVX-induced oxidative stress. These findings have also been confirmed in other studies.83 In addition, Kim34 and his team observed that the maximum load value of the femur in OVX mice was significantly lower than that in the sham surgery group, but luteolin could reverse this change (P<0.5). Furthermore, they examined the levels of osteogenic activity markers OC and bone resorption markers type I collagen carboxy-terminal peptide (CTX). The results showed that luteolin could increase serum OC levels and reduce CTX levels in mice, thereby preventing bone loss. Liang et al37 found that luteolin significantly enhanced the osteogenic differentiation potential of BMSCs in OVX mice. Notably, high doses of luteolin resulted in a greater increase in Runx2 levels compared to the positive drug group (alendronate sodium and vitamin D3 tablets), but further validation in clinical trials is necessary. In summary, these findings suggest that luteolin, as a natural compound, can effectively promote bone formation and inhibit bone loss, potentially serving as an alternative therapy for PMOP, particularly for individuals contraindicated for estrogen therapy.
The use of glucocorticoids (GC) is an important triggering factor for OP. In experimental studies, the administration of sustained-release glucocorticoid drugs via gavage, injection, or subcutaneous implantation is a commonly used method for modeling OP.84 Zheng et al76 established an OP rat model via intramuscular injection of DXM and administered luteolin via oral gavage for treatment. After two months, the results showed that luteolin significantly reversed phenomena such as reduced trabecular bone number, thinner trabeculae, and decreased bone parameters (BV/TV, TMC, Tb.Th.) in OP rats. Additionally, they observed that luteolin at doses of 25, 50, and 100 mg/kg significantly increased the maximum load, maximum bending stress, stiffness coefficient, and elastic modulus of the femur in a dose-dependent manner, thereby promoting bone strength in OP rats. These findings are consistent with the effects of luteolin treatment in an OVX mouse model. In a recent experiment, Tang et al39 also confirmed that luteolin effectively restored DEX-induced bone metabolic homeostasis imbalance. Further mechanistic studies revealed that this was primarily associated with luteolin’s ability to inhibit miR-125b-5p expression, activate the SIRT3/AMPK/mTOR axis, and promote autophagy. In summary, existing studies indicate that luteolin exhibits significant bone metabolic regulatory effects in OVX and GC-induced OP models. Additionally, the efficacy of luteolin in other types of OP models warrants further investigation.
Bone Metabolism Regulation of Luteolin in a Femoral Head Necrosis Model
Steroid-induced avascular necrosis of the femoral head is a condition caused by long-term, high-dose use of GC, characterized by vascular embolism, impaired blood supply, ischemic necrosis of bone tissue, and fracture of trabecular bone.85 Yan et al30 successfully established a glucocorticoid-induced osteonecrosis of the femoral head (GIONFH) model by intramuscularly injecting 10 mg/kg Dex into SD rats twice a week for 8 weeks, and then intervened with luteolin. The results showed that the incidence of GIONFH was significantly reduced in the Lut group compared with the model group. Compared with the control group, the model group showed a significantly increased proportion of cavities on HE staining, whereas a lower proportion of cavities was observed in the luteolin group. Micro-CT results of femoral head bone tissue showed that in the model group, cartilage was severely damaged, with significant decreases in Tb.N and BV/TV, and a significant increase in Tb.Sp. However, luteolin significantly reversed these changes, protecting against Dex-induced bone loss. In addition, they found that, mechanistically, luteolin may exert its protective effect against bone loss in GIONFH model rats by inhibiting Dex-induced apoptosis activation through the STAT1/caspase3 signaling pathway.
Bone Metabolism Regulation of Luteolin in Alveolar Bone Loss Models
Periodontal disease-induced alveolar bone loss is a common form of bone pathology and a major cause of tooth loss in adults.86 Modulating bone remodeling can greatly influence the occurrence and severity of alveolar bone loss. Research indicates that lipopolysaccharide (LPS) is one of the most important molecules involved in the development of periapical inflammation and alveolar bone resorption. An increase in its concentration leads to the release of various pro-inflammatory mediators, which activate multiple pathways to cause inflammation in periodontal tissues.87 Casili et al77 used intragingival injection of LPS to induce a periodontitis model in SD rats and then administered luteolin intervention. The results showed that luteolin could reduce LPS-induced alveolar bone loss, with the best effects observed at medium and high doses (30 and 100 mg/kg), while the low dose (10 mg/kg) had no significant improvement. Kariu et al78 found that luteolin can alleviate the extent of alveolar bone resorption around the maxillary molars by inhibiting the growth of Porphyromonas gingivalis in the oral cavity. Kawakami et al79 established Beagle and mouse models of alveolar bone resorption and intervened with luteolin. The results showed that luteolin significantly reduced bone resorption in both animal models, making it a promising candidate drug for the treatment of pulpitis and alveolar bone resorption. In a recent study, Ma et al80 induced periodontitis in SD rats using ligation. Local injection of luteolin significantly restored trabecular bone parameters (Tb.Th, BV/TV) while reducing the expression of TNF-α, IL-6, IL-1β, and IL-10, alleviating alveolar bone loss, and thereby improving periodontal inflammation and bone destruction. The above evidence indicates that luteolin, as a natural compound, significantly improves inflammation around the teeth and progressive bone destruction, and has the potential to become a strategy for oral health management.
Mechanism of Luteolin in Regulating Bone Metabolism
Bone metabolism represents a dynamic equilibrium between bone formation and resorption processes, potentially involving key signaling pathways that influence bone metabolism, including PI3K/Akt, Wnt, RANKL/RANK/OPG, Nrf2, and ERK pathways. We note that luteolin exerts beneficial regulatory effects on mitochondrial dysfunction, programmed cell death, oxidative stress, and the immune microenvironment through these pathways (Figure 2), making it a significant target for preventing and treating bone metabolism-related diseases.
 |
Figure 2 Mechanistic pathways by which luteolin influences bone metabolism. |
PI3K/Akt Signaling Pathway
PI3K is a lipid kinase that, upon stimulation by extracellular first messengers such as growth factors and cytokines, phosphorylates phosphatidylinositol bisphosphate (PIP2) to form phosphatidylinositol trisphosphate (PIP3). PIP3 acts as an important intracellular second messenger, responding to and transmitting extracellular signals.88 Akt is a serine/threonine protein kinase and one of the main downstream effectors of PI3K. PIP3 can activate Akt through phosphorylation of the Thr308 and Ser473 sites within Akt.89 Activated AKT activates or inhibits downstream target proteins such as glycogen synthase kinase (GSK) 3 and mTOR through phosphorylation, thereby regulating cellular life activities.90 The PI3K/Akt signaling pathway has a profound impact on the proliferation and differentiation of osteoblasts and is one of the important pathways regulating bone metabolism.91,92 Research indicates that luteolin can regulate bone metabolism by activating the PI3K/Akt pathway, thereby improving anti-mitochondrial function and suppressing oxidative stress, which prevents osteocyte apoptosis and pyroptosis.31,32 Additionally, by activating the PI3K-AKT signaling pathway, luteolin promotes the polarization of macrophages from the M1 phenotype to the M2 phenotype. This creates an anti-inflammatory immune state conducive to the osteogenic differentiation of BMSCs, thereby achieving immunoregulation of bone metabolism.72
Wnt Signaling Pathway
The Wnt signaling pathway is involved in the regulation of normal organ development and adult tissue function, including cell proliferation, differentiation, migration, polarity, and gene expression.93 The Wnt signaling pathway is divided into classical and non-classical Wnt signaling pathways based on whether they depend on β-catenin. The classic Wnt pathway is the β-catenin-dependent pathway, which is initiated when Wnt ligands bind to Frizzled receptors and low-density lipoprotein receptor-related protein 5/6 (LRP5/6), leading to the accumulation of intracellular β-catenin and its nuclear translocation. β-catenin then binds to the T-cell factor/lymphoid enhancer-binding factor (TCF/Lef) site, thereby activating the expression of downstream target genes.94 The Wnt signaling pathway is also regulated by inhibitors such as secreted frizzled-related proteins (SFRPs), Dickkopf (DKK), and sclerostin (SOST). Based on extensive in vitro and in vivo research, evidence suggests that the classic Wnt pathway plays an important role in the proliferation, differentiation, and function of osteoblasts, thereby regulating the formation and maintenance of bone tissue.95 Compared with the classical Wnt pathway that depends on β-catenin, the non-classical Wnt signaling pathway can activate signaling pathways independent of β-catenin and, in some cases, can also inhibit the classical Wnt pathway. The non-classical Wnt signaling pathway is further divided into the planar cell polarity (PCP) pathway and the Wnt/Ca2+ pathway. Recent studies have increasingly shown that non-classical Wnt signaling plays an important role in bone formation.96 Researchers have observed that luteolin promotes bone metabolism regulation by enhancing the mRNA expression of Lrp5 and β-catenin—key factors in the Wnt pathway—within MC3T3-E1 osteoblasts, thereby facilitating cell proliferation, differentiation, and mineralization.23
RANKL/RANK/OPG Signaling Pathway
The receptor activator of nuclear factor-kappa B ligand (RANKL)/receptor activator of nuclear factor-kappa B (RANK)/osteoprotegerin (OPG) signaling pathway is considered to be an important pathway regulating osteoclast function in bone metabolism.97 RANKL is a member of the tumor necrosis factor (TNF) ligand superfamily and is expressed by osteoblasts, osteocytes, and immune cells that make up bone tissue. Its expression is also regulated by various cytokines, glucocorticoids, and parathyroid hormone.98 RANK is a member of the TNF receptor superfamily and is expressed in various human tissues and cells, such as osteoclast progenitor cells, mature osteoclasts, and dendritic cells.99 RANKL binds to RANK on the surface of osteoclast progenitor cells, activates downstream intracellular signaling pathways through tumor necrosis factor receptor-associated factors (TNF receptor-associated factors, TRAFs), primarily including mitogen-activated protein kinase (MAPK) and nuclear factor-κB (NF-κB), stimulating osteoclast maturation and differentiation, promoting bone remodeling, and maintaining calcium homeostasis.100 OPG is a member of the TNF receptor superfamily, widely expressed in most human tissues, and regulated by various cytokines, peptides, hormones, and drugs.101 In bone tissue, OPG is secreted by osteoblasts and bone marrow stromal cells and acts as a soluble receptor for RANKL. OPG has a higher affinity for RANKL than RANK, and by competitively blocking the binding of RANKL to RANK, it prevents the transmission of signals from osteoblasts to osteoclasts, thereby inhibiting osteoclast differentiation and maturation.102 Research indicates that luteolin can inhibit osteoclast differentiation and reduce bone loss by regulating the RANKL/RANK/OPG signaling pathway.34
Nrf2 Signaling Pathway
Oxidative stress is a complex biological process characterized by excessive accumulation of ROS in cells or tissues, leading to an imbalance between the oxidative and antioxidant systems.103 Bone repair and bone remodeling are also redox-regulated processes, and physiological redox status is critical for the balance between osteoblast and osteoclast generation.104 Excessive production of ROS can increase osteoclast formation and activity, reduce osteoblast differentiation, and cause changes in bone structure.105 It is worth noting that Nrf2 plays a crucial role in maintaining cellular oxidative stress homeostasis. Nrf2 activates multiple downstream target genes by binding to upstream antioxidant response element (ARE) gene sequences, including heme oxygenase-1 (HO-1), SOD, GSH, and catalase (CAT).106,107 In Nrf2-deficient mouse models, significant bone loss can be observed, one of the reasons being that Nrf2 deficiency leads to an increase in osteoclasts and a decrease in osteoblast activity.108 Therefore, activating the Nrf2 signaling pathway and its downstream genes can ROS within osteoblasts, prevent oxidative stress-induced damage, and promote osteoblast differentiation and bone formation. A previous study26 confirmed that under LPS-induced inflammatory microenvironments and H2O2-induced oxidative stress, luteolin can protect osteoblasts from damage by upregulating Nrf2 expression in the human osteoblast cell line hFOB1.19. This action synergistically inhibits the NF-κB pathway, alleviates cellular oxidative stress and inflammatory responses, and thereby participates in bone metabolism regulation. Furthermore, luteolin activates the Nrf2 signaling pathway, upregulates HO-1 expression to scavenge ROS, inhibits M1 macrophage polarization, and promotes M2 macrophage polarization, thereby achieving regulation of bone metabolism immunity.73
ERK Signaling Pathway
The ERK signaling pathway is one of the major branches of the MAPK signaling pathway, extensively involved in cellular proliferation and differentiation processes.109 Activation of the ERK signaling pathway begins when growth factor receptors on the cell surface bind to growth factors and undergo autophosphorylation, subsequently activating the rat sarcoma viral oncogene homologue (RAS) protein attached to their interior. The RAS protein binds to guanosine triphosphate (GTP), which in turn promotes the activation of rapidly accelerated fibrosarcoma (RAF) kinase. The activated RAF protein then phosphorylates the downstream mitogen-activated protein kinase (MEK), which further phosphorylates the downstream ERK protein.110 Upon activation, ERK undergoes nuclear translocation and regulates downstream transcription factors, indirectly influencing the expression levels of key osteogenic factors such as Runx2, OSX, ALP, and type I collagen.111 Studies indicate that in glucocorticoid-induced OP models, luteolin promotes bone formation by activating the ERK signaling pathway. This activation regulates the downstream Lrp-5/GSK-3β/β-catenin pathway, thereby enhancing the expression of osteogenic markers, increasing ALP activity, promoting mineralized nodule formation, and mitigating bone loss.76
JAK/STAT Signaling Pathway
The core components of the Janus tyrosine kinase (JAK)/signal transducer and activator of transcription (STAT) signaling pathway include two classes of proteins: JAKs and STATs. In mammals, the JAK family comprises four members: JAK1, JAK2, JAK3, and TYK2. The STAT family consists of seven members: STAT1, STAT2, STAT3, STAT4, STAT5a, STAT5b, and STAT6.112 When signaling molecules bind to receptors, they induce conformational changes in the receptors and activate the associated JAKs. The activated JAK kinases transfer phosphate groups to STAT proteins through autophosphorylation, generating phosphorylated STAT proteins. This promotes conformational changes in the bound STATs, leading to the formation of homodimers.113 The formation of STAT dimers enables their nuclear translocation, where extracellular signals are ultimately transcribed into RNA via DNA, thereby altering gene expression and completing signal transduction. Reports indicate that JAK/STAT expression plays a crucial role in the progression of various diseases, with the JAK2/STAT3 pathway demonstrated to be pivotal in bone metabolic disorders.114 Research by Ma et al80 revealed that luteolin directly inhibits JAK2/STAT3 pathway activation, significantly reducing p-JAK2/JAK2 and p-STAT3/STAT3 ratios, and reduce p-STAT3 nuclear translocation. This further restores mitochondrial morphology and function while promoting macrophage polarization toward the M2 phenotype. Ultimately, it improves alveolar bone metabolism by mitigating inflammatory damage and inhibiting osteoclast activation.
Perspective
Pharmacokinetics
Luteolin has been used in the prevention and treatment of various diseases due to its wide range of pharmacological effects. However, its poor water solubility and liposolubility result in low bioavailability, which greatly limits its clinical use. Studies have shown that the oral absorption rate of 100 mg/kg luteolin in rats is only 10%, with a maximum plasma concentration of only 3.79 μg/mL. At the same time, luteolin is rapidly cleared from the plasma, and it is difficult to achieve effective plasma concentrations through either oral administration or intravenous injection.115 Tu et al116 found that within the range of 10–120 mg/kg, the oral absolute bioavailability of luteolin in rats was 3.87%–5.57%. Furthermore, due to factors such as solubility, membrane permeability, and transporters, the oral absorption of this compound may not increase proportionally with its dose (ie, it may not exhibit linear metabolism), thereby posing challenges for the formulation optimization of related oral preparations. In addition, they found that luteolin exhibited linear kinetic characteristics within the dose range of 10–80 mg/kg, with no significant changes observed when the dose was increased to 120 mg/kg. Therefore, they concluded that by limiting the luteolin dose to 80 mg/kg or below, its linear metabolic characteristics could be utilized to better evaluate the enhanced oral absorption performance of improved formulations, thereby providing important evidence for the development of related formulations.
Drug Delivery System
To advance the clinical application of luteolin, researchers have conducted in-depth studies on its formulation technology and new dosage forms. Wu et al117 developed a luteolin-loaded bio-composite material (LUPT40) containing 40 v% tantalum (Ta) and polyimide (PI), which significantly promotes osteoblast differentiation and inhibits osteoclast differentiation in vitro. In vivo experiments also confirmed its significant bone-forming effects. Importantly, LUPT40 achieves high bioavailability through the slow release of luteolin. Hu et al40 developed a 3D porous biodegradable scaffold (Poly(HEMA-co-3APBA)/LUT) that protects the activity of luteolin through borate ester bonds and prolongs its release, continuously inducing M2 polarization of macrophages and thereby promoting bone regeneration, providing an “immune-osteogenic” synergistic strategy for the treatment of critical-size bone defects (CSBDs). Yu72 and his team combined 3D bioprinting technology, metal-organic frameworks (MOFs), and hydrogels to develop a multifunctional biomimetic scaffold (LUT@ZIF-8/GelMA) with excellent physical properties and biocompatibility. This scaffold can slowly release luteolin and zinc ions, regulate the immune microenvironment, and promote the osteogenic differentiation of BMSCs. Similarly, subsequent scholars73 developed a manganese-luteolin (Mn-Lut) nanoflower biphasic hydrogel scaffold based on 3D printing technology, which has the functions of sustained release of manganese ions and luteolin, antagonizing oxidative stress, and regulating macrophage polarization to the M2 type, thereby promoting the osteogenic and chondrogenic differentiation of BMSCs, significantly promoting bone and cartilage regeneration in a rat bone-cartilage defect model. In addition, these studies indicate that at the experimental concentration of luteolin administered, no significant cellular toxicity was observed in the short term, and no abnormalities in liver or kidney function were detected in vivo. However, long-term safety assessments remain limited. In summary, the above design approach is beneficial for improving the bioavailability of luteolin and provides a valuable research foundation for subsequent studies. In addition, with the increasing number of new luteolin-based formulations, there is great potential for tissue engineering applications in regulating bone metabolism for bone repair.
Toxicity
Clarifying the toxicological properties of luteolin is essential for ensuring its safe application as a natural product in food and pharmaceutical fields. Luteolin is generally recognized as a low-toxicity bioactive natural compound. Research by Wen et al118 demonstrated extremely low toxicity of luteolin toward human umbilical vein endothelial cells (HUVEC). Furthermore, in Vero cells (African green monkey kidney cells), luteolin exhibits low toxicity and potent cytoprotective properties: concentrations of 80 μmol/L and above effectively inhibit Shiga toxin (Stx1, Stx2)-induced cytotoxicity, while remaining non-toxic to Vero cells at 400 μmol/L.119 In vivo studies indicate that intraperitoneal administration of 10 mg/kg luteolin to rats showed no drug-related adverse reactions or toxicity within 24 hours. The median lethal dose (LD50) for intraperitoneal luteolin in rats was 411 mg/kg, while the oral LD50 exceeded 5000 mg/kg.120 Furthermore, in zebrafish models, luteolin concentrations of 100 μM and 500 μM did not affect embryo survival and improved hatching delays and malformation rates in mutant embryos, demonstrating favorable safety profiles.121 Although these findings suggest luteolin is a relatively safe natural compound, the literature on luteolin toxicity studies remains limited. Furthermore, its safety for human use remains unclear and requires further evaluation in clinical trials.
Conclusions and Perspectives
In summary, we have found that luteolin promotes bone activity and inhibits bone resorption in both cell and animal models, thereby balancing bone metabolism and maintaining bone health. The specific mechanisms involve regulating multiple signaling pathways, including PI3K/Akt, Wnt, RANKL/RANK/OPG, MAPK and Nrf2, to inhibit oxidative stress, reduce inflammatory responses, regulate cell death, and modulate the immune microenvironment. These results may provide a basis for transferring the therapeutic use of woodruff testing to clinical practice. However, several critical issues remain to be explored in current research. On one hand, studies are still in the early stages of clinical translation, with a gap existing between experimental findings and practical clinical application. Therefore, human trials are necessary to validate its clinical safety and efficacy, thereby bridging this translational gap. On the other hand, luteolin exhibits suboptimal absorption and metabolic stability, necessitating further optimization of drug delivery routes and bioavailability. Furthermore, standardized drug dosages are essential for ensuring the reliability and reproducibility of experimental results. Overall, as a natural compound, luteolin has good bone metabolism regulation effects and shows promise as a potential alternative therapy for the treatment of bone metabolic diseases.
Acknowledgment
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was supported by Enpeng Gu Tianjin Famous Traditional Chinese Medicine Inheritance Studio construction project, Modern Traditional Chinese Medicine Haihe Laboratory Science and Technology Project (GEP20250102); The Tianjin Municipal Health Commission Integrated Chinese and Western Medicine Project (2023072), and the Tianjin Binhai New Area Hospital of Traditional Chinese Medicine Science and Technology Project (bhzy2024q02).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Henderson B, Nair SP. Hard labour: bacterial infection of the skeleton. Trends Microbiol. 2003;11(12):570–577. doi:10.1016/j.tim.2003.10.005
2. Zhou M, An YZ, Guo Q, Zhou HY, Luo XH. Energy homeostasis in the bone. Trends Endocrinol Metab. 2024;35(5):439–451. doi:10.1016/j.tem.2023.12.009
3. Fang F, Yang J, Wang J, et al. The role and applications of extracellular vesicles in osteoporosis. Bone Res. 2024;12(1):4. doi:10.1038/s41413-023-00313-5
4. Zhang L, Kwack KH, Thiyagarajan R, et al. Tristetraprolin regulates the skeletal phenotype and osteoclastogenic potential through monocytic myeloid-derived suppressor cells. FASEB J. 2024;38(1):e23338. doi:10.1096/fj.202301703R
5. Zhu G, Chen W, Tang CY, et al. Knockout and Double Knockout of Cathepsin K and Mmp9 reveals a novel function of Cathepsin K as a regulator of osteoclast gene expression and bone homeostasis. Int J Biol Sci. 2022;18(14):5522–5538. doi:10.7150/ijbs.72211
6. Black DM, Geiger EJ, Eastell R, et al. Atypical femur fracture risk versus fragility fracture prevention with bisphosphonates. New Engl J Med. 2020;383(8):743–753. doi:10.1056/NEJMoa1916525
7. Salari N, Ghasemi H, Mohammadi L, et al. The global prevalence of osteoporosis in the world: a comprehensive systematic review and meta-analysis. J Orthop Surg Res. 2021;16(1):609. doi:10.1186/s13018-021-02772-0
8. Solomon L. Idiopathic necrosis of the femoral head: pathogenesis and treatment. Can J Surg. 1981;24(6):573–578.
9. Liu F, Wang W, Yang L, et al. An epidemiological study of etiology and clinical characteristics in patients with nontraumatic osteonecrosis of the femoral head. J Res Med Sci. 2017;22:15. doi:10.4103/1735-1995.200273
10. Al MD, Ghosh YA, Xin H, et al. Hydrogel: a potential material for bone tissue engineering repairing the segmental mandibular defect. Polymers. 2022;14(19):4186. doi:10.3390/polym14194186
11. Holly D, Klein M, Mazreku M, et al. Stem cells and their derivatives-implications for alveolar bone regeneration: a comprehensive review. Int J Mol Sci. 2021;22(21):11746. doi:10.3390/ijms222111746
12. Qu Z, Zhao S, Zhang Y, Wang X, Yan L. Natural compounds for bone remodeling: targeting osteoblasts and relevant signaling pathways. Biomed Pharmacother. 2024;180:117490. doi:10.1016/j.biopha.2024.117490
13. Prabakaran S, Rohini K. Evaluation of osteogenic potential of Centella asiatica extract @ hydroxyapatite based composite coating on titanium implant by dip coating method for orthopedic applications. J Aust Ceram Soc. 2025;61(2):755–773. doi:10.1007/s41779-025-01150-x
14. Jang TY, Jung AY, Kyung TS, Kim DY, Hwang JH, Kim YH. Anti-allergic effect of luteolin in mice with allergic asthma and rhinitis. Cent Eur J Immunol. 2017;42(1):24–29. doi:10.5114/ceji.2017.67315
15. Theoharides TC. COVID-19, pulmonary mast cells, cytokine storms, and beneficial actions of luteolin. Biofactors. 2020;46(3):306–308. doi:10.1002/biof.1633
16. Zheng YZ, Chen DF, Deng G, Guo R, Fu ZM. The surrounding environments on the structure and antioxidative activity of luteolin. J Mol Model. 2018;24(7):149. doi:10.1007/s00894-018-3680-1
17. Gonzalez-Alamilla EN, Gonzalez-Cortazar M, Valladares-Carranza B, et al. Chemical constituents of Salix babylonica L. and their antibacterial activity against gram-positive and gram-negative animal bacteria. Molecules. 2019;24(16):2992. doi:10.3390/molecules24162992
18. Kou JJ, Shi JZ, He YY, et al. Luteolin alleviates cognitive impairment in Alzheimer’s disease mouse model via inhibiting endoplasmic reticulum stress-dependent neuroinflammation. Acta Pharmacol Sin. 2022;43(4):840–849. doi:10.1038/s41401-021-00702-8
19. Seelinger G, Merfort I, Wolfle U, Schempp CM. Anti-carcinogenic effects of the flavonoid luteolin. Molecules. 2008;13(10):2628–2651. doi:10.3390/molecules13102628
20. Ntalouka F, Tsirivakou A. Luteolin: a promising natural agent in management of pain in chronic conditions. Front Pain Res. 2023;4:1114428. doi:10.3389/fpain.2023.1114428
21. Tsai HY, Chen MY, Hsu C, et al. Luteolin phosphate derivatives generated by cultivating Bacillus subtilis var. Natto BCRC 80517 with luteolin. J Agr Food Chem. 2022;70(28):8738–8745. doi:10.1021/acs.jafc.2c03524
22. Zhao C, Sun H, Yan X, et al. Potential mechanism of luteolin activating the PI3K/AKT signaling pathway to promote proliferation and osteogenic differentiation in MC3T3-E1 cells. J Integr Med. 2024;19(11):2177–2186+2197. doi:10.13935/j.cnki.sjzx.241112
23. Shang D, Wang H, Han Y, Zhao C. Effects of luteolin on proliferation, differentiation, mineralization, and wnt signaling pathway in in vitro cultured mouse pre-osteoblasts MC3T3-E1. J Bengbu Med Coll. 2017;42(01):44–47. doi:10.13898/j.cnki.issn.1000-2200.2017.01.012
24. Yang H, Liu Q, Ahn JH, et al. Luteolin downregulates IL-1beta-induced MMP-9 and −13 expressions in osteoblasts via inhibition of ERK signalling pathway. J Enzyme Inhib Med Chem. 2012;27(2):261–266. doi:10.3109/14756366.2011.587415
25. Choi EM. Modulatory effects of luteolin on osteoblastic function and inflammatory mediators in osteoblastic MC3T3-E1 cells. Cell Biol Int. 2007;31(9):870–877. doi:10.1016/j.cellbi.2007.01.038
26. Peng Z, Zhang W, Hong H, Liu L. Effect of luteolin on oxidative stress and inflammation in the human osteoblast cell line hFOB1.19 in an inflammatory microenvironment. BMC Pharmacol Toxicol. 2024;25(1):40. doi:10.1186/s40360-024-00764-4
27. Fatokun AA, Tome M, Smith RA, Darlington LG, Stone TW. Protection by the flavonoids quercetin and luteolin against peroxide- or menadione-induced oxidative stress in MC3T3-E1 osteoblast cells. Nat Prod Res. 2015;29(12):1127–1132. doi:10.1080/14786419.2014.980252
28. Abbasi N, Khosravi A, Aidy A, Shafiei M. Biphasic response to luteolin in MG-63 osteoblast-like cells under high glucose-induced oxidative stress. Iran J Med Sci. 2016;41(2):118–125.
29. Suh KS, Chon S, Choi EM. Luteolin alleviates methylglyoxal-induced cytotoxicity in osteoblastic MC3T3-E1 cells. Cytotechnology. 2016;68(6):2539–2552. doi:10.1007/s10616-016-9977-y
30. Yan Z, Zhan J, Qi W, et al. The protective effect of luteolin in glucocorticoid-induced osteonecrosis of the femoral head. Front Pharmacol. 2020;11:1195. doi:10.3389/fphar.2020.01195
31. Chai S, Yang Y, Wei L, et al. Luteolin rescues postmenopausal osteoporosis elicited by OVX through alleviating osteoblast pyroptosis via activating PI3K-AKT signaling. Phytomedicine. 2024;128:155516. doi:10.1016/j.phymed.2024.155516
32. Choi EM. Luteolin protects osteoblastic MC3T3-E1 cells from antimycin A-induced cytotoxicity through the improved mitochondrial function and activation of PI3K/Akt/CREB. Toxicol In Vitro. 2011;25(8):1671–1679. doi:10.1016/j.tiv.2011.07.004
33. Lee JW, Ahn JY, Hasegawa S, et al. Inhibitory effect of luteolin on osteoclast differentiation and function. Cytotechnology. 2009;61(3):125–134. doi:10.1007/s10616-010-9253-5
34. Kim TH, Jung JW, Ha BG, et al. The effects of luteolin on osteoclast differentiation, function in vitro and ovariectomy-induced bone loss. J Nutr Biochem. 2011;22(1):8–15. doi:10.1016/j.jnutbio.2009.11.002
35. Shin DK, Kim MH, Lee SH, Kim TH, Kim SY. Inhibitory effects of luteolin on titanium particle-induced osteolysis in a mouse model. Acta Biomater. 2012;8(9):3524–3531. doi:10.1016/j.actbio.2012.05.002
36. Crasto GJ, Kartner N, Yao Y, et al. Luteolin inhibition of V-ATPase a3-d2 interaction decreases osteoclast resorptive activity. J Cell Biochem. 2013;114(4):929–941. doi:10.1002/jcb.24434
37. Liang G, Zhao J, Dou Y, et al. Mechanism and experimental verification of luteolin for the treatment of osteoporosis based on network pharmacology. Front Endocrinol. 2022;13:866641. doi:10.3389/fendo.2022.866641
38. Oh JH, Karadeniz F, Lee JI, Seo Y, Jang M, Kong C. Effect and comparison of luteolin and its derivative sodium luteolin-4′-sulfonate on adipogenic differentiation of human bone marrow-derived mesenchymal stem cells through AMPK-mediated PPAR γ signaling. Evid Based Complement Alternat Med. 2020;2020:8894910. doi:10.1155/2020/8894910
39. Tang L, Fan X, Xu Y, Zhang Y, Li G. Luteolin inhibits dexamethasone-induced osteoporosis by autophagy activation through miR-125b-5p/SIRT3/AMPK/mTOR axis, an in vitro and in vivo study. Food Sci Nutr. 2025;13(3):e70071. doi:10.1002/fsn3.70071
40. Hu Y, Tang L, Wang Z, et al. Inducing in situ M2 macrophage polarization to promote the repair of bone defects via scaffold-mediated sustained delivery of luteolin. J Control Release. 2024;365:889–904. doi:10.1016/j.jconrel.2023.11.015
41. Zhou Z, Ma X, Yao H, et al. Effects of combined naringin and luteolin on gene expression related to the Wnt/β-catenin pathway during osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. Adv Mod Biomed Sci. 2023;23(02):258–262. doi:10.13241/j.cnki.pmb.2023.02.010
42. Nagase H, Woessner JJ. Matrix metalloproteinases. J Biol Chem. 1999;274(31):21491–21494. doi:10.1074/jbc.274.31.21491
43. Moncada S, Palmer RM, Higgs EA. Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev. 1991;43(2):109–142.
44. Barnes PJ. Oxidative stress-based therapeutics in COPD. Redox Biol. 2020;33:101544. doi:10.1016/j.redox.2020.101544
45. Sul OJ, Ra SW. Quercetin prevents LPS-induced oxidative stress and inflammation by modulating NOX2/ROS/NF-kB in lung epithelial cells. Molecules. 2021;26(22):6949. doi:10.3390/molecules26226949
46. Lozano D, de Castro LF, Dapia S, et al. Role of parathyroid hormone-related protein in the decreased osteoblast function in diabetes-related osteopenia. Endocrinology. 2009;150(5):2027–2035. doi:10.1210/en.2008-1108
47. Bai XC, Lu D, Bai J, et al. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-kappaB. Biochem Biophys Res Commun. 2004;314(1):197–207. doi:10.1016/j.bbrc.2003.12.073
48. Wu CH, Huang SM, Lin JA, Yen GC. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011;2(5):224–234. doi:10.1039/c1fo10026b
49. Talukdar D, Chaudhuri BS, Ray M, Ray S. Critical evaluation of toxic versus beneficial effects of methylglyoxal. Biochemistry. 2009;74(10):1059–1069. doi:10.1134/s0006297909100010
50. Banuelos J, Shin S, Cao Y, et al. BCL-2 protects human and mouse Th17 cells from glucocorticoid-induced apoptosis. Allergy. 2016;71(5):640–650. doi:10.1111/all.12840
51. Wang T, Liu X, He C. Glucocorticoid-induced autophagy and apoptosis in bone. Apoptosis. 2020;25(3–4):157–168. doi:10.1007/s10495-020-01599-0
52. Gu Q, Chen M, Zhang Y, Huang Y, Yang H, Shi Q. Haem oxygenase-1 induction prevents glucocorticoid-induced osteoblast apoptosis through activation of extracellular signal-regulated kinase1/2 signalling pathway. J Orthop Translat. 2019;19:29–37. doi:10.1016/j.jot.2019.04.003
53. Butturini E, Cozzolino F, Boriero D, et al. S-glutathionylation exerts opposing roles in the regulation of STAT1 and STAT3 signaling in reactive microglia. Free Radic Biol Med. 2018;117:191–201. doi:10.1016/j.freeradbiomed.2018.02.005
54. Murakami T, Nakaminami Y, Takahata Y, Hata K, Nishimura R. Activation and function of NLRP3 inflammasome in bone and joint-related diseases. Int J Mol Sci. 2022;23(10):5365. doi:10.3390/ijms23105365
55. Chen L, Shi X, Xie J, et al. Apelin-13 induces mitophagy in bone marrow mesenchymal stem cells to suppress intracellular oxidative stress and ameliorate osteoporosis by activation of AMPK signaling pathway. Free Radic Biol Med. 2021;163:356–368. doi:10.1016/j.freeradbiomed.2020.12.235
56. Li XY, Zhan XR, Liu XM, Wang XC. CREB is a regulatory target for the protein kinase Akt/PKB in the differentiation of pancreatic ductal cells into islet beta-cells mediated by hepatocyte growth factor. Biochem Biophys Res Commun. 2011;404(2):711–716. doi:10.1016/j.bbrc.2010.12.048
57. Fan R, Li X, Gu X, Chan JC, Xu G. Exendin-4 protects pancreatic beta cells from human islet amyloid polypeptide-induced cell damage: potential involvement of AKT and mitochondria biogenesis. Diabetes Obes Metab. 2010;12(9):815–824. doi:10.1111/j.1463-1326.2010.01238.x
58. Kwon S, Kim S, Song N, et al. Antiadipogenic and proosteogenic effects of luteolin, a major dietary flavone, are mediated by the induction of DnaJ (Hsp40) Homolog, Subfamily B, Member 1. J Nutr Biochem. 2016;30:24–32. doi:10.1016/j.jnutbio.2015.11.013
59. Suda T, Takahashi N, Martin TJ. Modulation of osteoclast differentiation. Endocr Rev. 1992;13(1):66–80. doi:10.1210/edrv-13-1-66
60. Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and activation. Nature. 2003;423(6937):337–342. doi:10.1038/nature01658
61. Pacifici R. Cytokines, estrogen, and postmenopausal osteoporosis--the second decade. Endocrinology. 1998;139(6):2659–2661. doi:10.1210/endo.139.6.6087
62. Henriksen K, Bollerslev J, Everts V, Karsdal MA. Osteoclast activity and subtypes as a function of physiology and pathology--implications for future treatments of osteoporosis. Endocr Rev. 2011;32(1):31–63. doi:10.1210/er.2010-0006
63. Lee SH, Rho J, Jeong D, et al. v-ATPase V0 subunit d2-deficient mice exhibit impaired osteoclast fusion and increased bone formation. Nat Med. 2006;12(12):1403–1409. doi:10.1038/nm1514
64. Manolson MF, Yu H, Chen W, et al. The a3 isoform of the 100-kDa V-ATPase subunit is highly but differentially expressed in large (>or=10 nuclei) and small (<or= nuclei) osteoclasts. J Biol Chem. 2003;278(49):49271–49278. doi:10.1074/jbc.M309914200
65. Luo ZW, Li FX, Liu YW, et al. Aptamer-functionalized exosomes from bone marrow stromal cells target bone to promote bone regeneration. Nanoscale. 2019;11(43):20884–20892. doi:10.1039/c9nr02791b
66. Chen Q, Shou P, Zheng C, et al. Fate decision of mesenchymal stem cells: adipocytes or osteoblasts? Cell Death Differ. 2016;23(7):1128–1139. doi:10.1038/cdd.2015.168
67. Samare-Najaf M, Neisy A, Samareh A, et al. The constructive and destructive impact of autophagy on both genders’ reproducibility, a comprehensive review. Autophagy. 2023;19(12):3033–3061. doi:10.1080/15548627.2023.2238577
68. Ogasawara T, Ito S, Ogashira S, et al. The expression of MIR125B transcripts and bone phenotypes in Mir125b2-deficient mice. PLoS One. 2024;19(7):e0304074. doi:10.1371/journal.pone.0304074
69. Yang C, Fan H, Wu Y, et al. T-2 toxin exposure induces ovarian damage in sows: lncRNA CUFF.253988.1 promotes cell apoptosis by inhibiting the SIRT3/PGC1α pathway. Ecotox Environ Safe. 2024;283:116787. doi:10.1016/j.ecoenv.2024.116787
70. Chen FF, Liu JF, Zhou DM. SIRT3 enhances the protective effect of Xyloketal B on seizure-induced brain injury by regulating AMPK/mTOR signaling-mediated autophagy. Kaohsiung J Med Sci. 2024;40(1):74–85. doi:10.1002/kjm2.12765
71. Miron RJ, Bosshardt DD. OsteoMacs: key players around bone biomaterials. Biomaterials. 2016;82:1–19. doi:10.1016/j.biomaterials.2015.12.017
72. Yu SY, Wu T, Xu KH, et al. 3D bioprinted biomimetic MOF-functionalized hydrogel scaffolds for bone regeneration: synergistic osteogenesis and osteoimmunomodulation. Mater Today Bio. 2025;32:101740. doi:10.1016/j.mtbio.2025.101740
73. Yuan S, Li H, Xu Z, et al. Immune regulation and repair of osteochondral defects using manganese-luteolin hydrogel scaffold. J Control Release. 2025;384:113920. doi:10.1016/j.jconrel.2025.113920
74. Mohamed-Ahmed S, Fristad I, Lie SA, et al. Adipose-derived and bone marrow mesenchymal stem cells: a donor-matched comparison. Stem Cell Res Ther. 2018;9(1):168. doi:10.1186/s13287-018-0914-1
75. Wang X, Luo E, Bi R, Ye B, Hu J, Zou S. Wnt/beta-catenin signaling is required for distraction osteogenesis in rats. Connect Tissue Res. 2018;59(1):45–54. doi:10.1080/03008207.2017.1300154
76. Jing Z, Wang C, Yang Q, et al. Luteolin attenuates glucocorticoid-induced osteoporosis by regulating ERK/Lrp-5/GSK-3beta signaling pathway in vivo and in vitro. J Cell Physiol. 2019;234(4):4472–4490. doi:10.1002/jcp.27252
77. Casili G, Ardizzone A, Lanza M, et al. Treatment with luteolin improves lipopolysaccharide-induced periodontal diseases in rats. Biomedicines. 2020;8(10):442. doi:10.3390/biomedicines8100442
78. Kariu T, Hamada N, Lakshmyya K. Luteolin inhibits Porphyromonas gingivalis growth and alleviates alveolar bone destruction in experimental murine periodontitis. Biosci Biotechnol Biochem. 2023;88(1):37–43. doi:10.1093/bbb/zbad137
79. Kawakami K, Fukuda T, Toyoda M, et al. Luteolin is a potential immunomodulating natural compound against pulpal inflammation. Biomed Res Int. 2024;2024:8864513. doi:10.1155/2024/8864513
80. Ma S, He H, Ren X, et al. Luteolin ameliorates periodontitis by modulating mitochondrial dynamics and macrophage polarization via the JAK2/STAT3 pathway. Int Immunopharmacol. 2025;144:113612. doi:10.1016/j.intimp.2024.113612
81. Li J, Chen X, Lu L, Yu X. The relationship between bone marrow adipose tissue and bone metabolism in postmenopausal osteoporosis. Cytokine Growth Factor Rev. 2020;52:88–98. doi:10.1016/j.cytogfr.2020.02.003
82. Brent MB. Pharmaceutical treatment of bone loss: from animal models and drug development to future treatment strategies. Pharmacol Ther. 2023;244:108383. doi:10.1016/j.pharmthera.2023.108383
83. Xu Y, Xie X, Yan W, Xue B, Xiong Y. Mechanism of luteolin modulating PI3K/AKT pathway to improve bone loss in postmenopausal osteoporosis rat model. J Trop Med. 2022;22(05):639–643+688+744.
84. Baofeng L, Zhi Y, Bei C, Guolin M, Qingshui Y, Jian L. Characterization of a rabbit osteoporosis model induced by ovariectomy and glucocorticoid. Acta Orthop. 2010;81(3):396–401. doi:10.3109/17453674.2010.483986
85. Zhang F, Wei L, Wang L, et al. FAR591 promotes the pathogenesis and progression of SONFH by regulating Fos expression to mediate the apoptosis of bone microvascular endothelial cells. Bone Res. 2023;11(1):27. doi:10.1038/s41413-023-00259-8
86. Leone CW, Bokhadhoor H, Kuo D, et al. Immunization enhances inflammation and tissue destruction in response to Porphyromonas gingivalis. Infect Immun. 2006;74(4):2286–2292. doi:10.1128/IAI.74.4.2286-2292.2006
87. Barksby HE, Nile CJ, Jaedicke KM, Taylor JJ, Preshaw PM. Differential expression of immunoregulatory genes in monocytes in response to Porphyromonas gingivalis and Escherichia coli lipopolysaccharide. Clin Exp Immunol. 2009;156(3):479–487. doi:10.1111/j.1365-2249.2009.03920.x
88. Fontana F, Giannitti G, Marchesi S, Limonta P. The PI3K/Akt pathway and glucose metabolism: a dangerous liaison in cancer. Int J Biol Sci. 2024;20(8):3113–3125. doi:10.7150/ijbs.89942
89. Chamcheu JC, Roy T, Uddin MB, et al. Role and therapeutic targeting of the PI3K/Akt/mTOR signaling pathway in skin cancer: a review of current status and future trends on natural and synthetic agents therapy. Cells. 2019;8(8):803. doi:10.3390/cells8080803
90. Rokutanda S, Fujita T, Kanatani N, et al. Akt regulates skeletal development through GSK3, mTOR, and FoxOs. Dev Biol. 2009;328(1):78–93. doi:10.1016/j.ydbio.2009.01.009
91. Kang HY, Cho CL, Huang KL, et al. Nongenomic androgen activation of phosphatidylinositol 3-kinase/Akt signaling pathway in MC3T3-E1 osteoblasts. J Bone Miner Res. 2004;19(7):1181–1190. doi:10.1359/JBMR.040306
92. Gu YX, Du J, Si MS, Mo JJ, Qiao SC, Lai HC. The roles of PI3K/Akt signaling pathway in regulating MC3T3-E1 preosteoblast proliferation and differentiation on SLA and SLActive titanium surfaces. J Biomed Mater Res A. 2013;101(3):748–754. doi:10.1002/jbm.a.34377
93. Wang Y, Liu J, Zheng S, Cao L, Li Y, Sheng R. The deubiquitinase USP10 mediates crosstalk between the LKB1/AMPK axis and Wnt/beta-catenin signaling in cancer. FEBS Lett. 2023;597(24):3061–3071. doi:10.1002/1873-3468.14763
94. Du JH, Lin SX, Wu XL, et al. The function of wnt ligands on osteocyte and bone remodeling. J Dent Res. 2019;98(8):930–938. doi:10.1177/0022034519854704
95. Chae WJ, Bothwell A. Dickkopf1: an immunomodulatory ligand and Wnt antagonist in pathological inflammation. Differentiation. 2019;108:33–39. doi:10.1016/j.diff.2019.05.003
96. Takada I, Mihara M, Suzawa M, et al. A histone lysine methyltransferase activated by non-canonical Wnt signalling suppresses PPAR-gamma transactivation. Nat Cell Biol. 2007;9(11):1273–1285. doi:10.1038/ncb1647
97. Zhao Y, Wang HL, Li TT, Yang F, Tzeng CM. Baicalin ameliorates dexamethasone-induced osteoporosis by regulation of the RANK/RANKL/OPG signaling pathway. Drug Des Devel Ther. 2020;14:195–206. doi:10.2147/DDDT.S225516
98. Saidenberg KN, Bessis N, Cohen-Solal M, De Vernejoul MC, Boissier MC. Osteoprotegerin and inflammation. Eur Cytokine Netw. 2002;13(2):144–153.
99. Carrillo-Lopez N, Martinez-Arias L, Fernandez-Villabrille S, et al. Role of the RANK/RANKL/OPG and Wnt/beta-Catenin systems in CKD bone and cardiovascular disorders. Calcified Tissue Int. 2021;108(4):439–451. doi:10.1007/s00223-020-00803-2
100. Li L, Sapkota M, Gao M, Choi H, Soh Y. Macrolactin F inhibits RANKL-mediated osteoclastogenesis by suppressing Akt, MAPK and NFATc1 pathways and promotes osteoblastogenesis through a BMP-2/smad/Akt/Runx2 signaling pathway. Eur J Pharmacol. 2017;815:202–209. doi:10.1016/j.ejphar.2017.09.015
101. Sezer O, Heider U, Zavrski I, Kuhne CA, Hofbauer LC. RANK ligand and osteoprotegerin in myeloma bone disease. Blood. 2003;101(6):2094–2098. doi:10.1182/blood-2002-09-2684
102. Mbundi L, Meikle ST, Busquets R, Dowell NG, Cercignani M, Santin M. Gadolinium tagged osteoprotegerin-mimicking peptide: a novel magnetic resonance imaging biospecific contrast agent for the inhibition of osteoclastogenesis and osteoclast activity. Nanomaterials. 2018;8(6):399. doi:10.3390/nano8060399
103. Ebrahimi S, Alalikhan A, Aghaee-Bakhtiari SH, Hashemy SI. The redox modulatory effects of SP/NK1R system: implications for oxidative stress-associated disorders. Life Sci. 2022;296:120448. doi:10.1016/j.lfs.2022.120448
104. Hong SE, Lee J, Seo DH, et al. Euphorbia factor L1 inhibits osteoclastogenesis by regulating cellular redox status and induces Fas-mediated apoptosis in osteoclast. Free Radic Biol Med. 2017;112:191–199. doi:10.1016/j.freeradbiomed.2017.07.030
105. Kimball JS, Johnson JP, Carlson DA. Oxidative stress and osteoporosis. J Bone Joint Surg Am. 2021;103(15):1451–1461. doi:10.2106/JBJS.20.00989
106. Guo S, Zhang Q. Paeonol protects melanocytes against hydrogen peroxide-induced oxidative stress through activation of Nrf2 signaling pathway. Drug Dev Res. 2021;82(6):861–869. doi:10.1002/ddr.21793
107. Krajka-Kuzniak V, Paluszczak J, Baer-Dubowska W. The Nrf2-ARE signaling pathway: an update on its regulation and possible role in cancer prevention and treatment. Pharmacol Rep. 2017;69(3):393–402. doi:10.1016/j.pharep.2016.12.011
108. Han J, Yang K, An J, Jiang N, Fu S, Tang X. The role of NRF2 in bone metabolism - friend or foe? Front Endocrinol. 2022;13:813057. doi:10.3389/fendo.2022.813057
109. Yue J, Lopez JM. Understanding MAPK signaling pathways in apoptosis. Int J Mol Sci. 2020;21(7):2346. doi:10.3390/ijms21072346
110. Lavoie H, Gagnon J, Therrien M. ERK signalling: a master regulator of cell behaviour, life and fate. Nat Rev Mol Cell Bio. 2020;21(10):607–632. doi:10.1038/s41580-020-0255-7
111. Dou X, Wei X, Liu G, et al. Effect of porous tantalum on promoting the osteogenic differentiation of bone marrow mesenchymal stem cells in vitro through the MAPK/ERK signal pathway. J Orthop Transl. 2019;19:81–93. doi:10.1016/j.jot.2019.03.006
112. Philips RL, Wang Y, Cheon H, et al. The JAK-STAT pathway at 30: much learned, much more to do. Cell. 2022;185(21):3857–3876. doi:10.1016/j.cell.2022.09.023
113. Wang L, Zhao D, Wang H, Wang L, Liu X, Zhang H. FPS-ZM1 inhibits LPS-induced microglial inflammation by suppressing JAK/STAT signaling pathway. Int Immunopharmacol. 2021;100:108117. doi:10.1016/j.intimp.2021.108117
114. Zhu J, Tang Y, Wu Q, Ji YC, Feng ZF, Kang FW. HIF-1alpha facilitates osteocyte-mediated osteoclastogenesis by activating JAK2/STAT3 pathway in vitro. J Cell Physiol. 2019;234(11):21182–21192. doi:10.1002/jcp.28721
115. Lin LC, Pai YF, Tsai TH. Isolation of luteolin and luteolin-7-O-glucoside from Dendranthema morifolium Ramat Tzvel and their pharmacokinetics in rats. J Agr Food Chem. 2015;63(35):7700–7706. doi:10.1021/jf505848z
116. Tu L, Yang R, Liu J, Cheng M, Jin Y, Feng J. Comparative pharmacokinetics of different doses of luteolin in rats. Chin Pat Med. 2022;44(12):3753–3757.
117. Wu Z, Kaewmanee R, Yang Q, et al. Luteolin-loaded biocomposites containing tantalum and polyimide with antibacterial effects for facilitating osteogenic differentiation and bone bonding. J Mater Chem B. 2023;11(42):10218–10233. doi:10.1039/d3tb01546g
118. Wen L, Zhang W, Hu J, et al. Luteolin target HSPB1 regulates endothelial cell ferroptosis to protect against radiation vascular injury. PLoS One. 2024;19(10):e0311922. doi:10.1371/journal.pone.0311922
119. Yuan L, Nakamichi R, Hirata Y, et al. Mechanism for inhibition of cytotoxicity of Shiga toxin by luteolin. Toxicol In Vitro. 2023;87:105537. doi:10.1016/j.tiv.2022.105537
120. Kanai K, Nagata S, Hatta T, et al. Therapeutic anti-inflammatory effects of luteolin on endotoxin-induced uveitis in Lewis rats. J Vet Med Sci. 2016;78(8):1381–1384. doi:10.1292/jvms.16-0196
121. De Leo E, Elmonem MA, Berlingerio SP, et al. Cell-based phenotypic drug screening identifies luteolin as candidate therapeutic for nephropathic cystinosis. J Am Soc Nephrol. 2020;31(7):1522–1537. doi:10.1681/ASN.2019090956
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.