Back to Journals » OncoTargets and Therapy » Volume 9
Long noncoding RNAs in gastric cancer: functions and clinical applications
Authors Wang J ![]() , Sun J, Wang J, Song Y
, Sun J, Wang J, Song Y ![]() , Gao P, Shi J, Chen P, Wang Z
, Gao P, Shi J, Chen P, Wang Z
Received 29 August 2015
Accepted for publication 28 November 2015
Published 10 February 2016 Volume 2016:9 Pages 681—697
DOI https://doi.org/10.2147/OTT.S95412
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Faris Farassati
Jiajun Wang,* Jingxu Sun,* Jun Wang, Yongxi Song, Peng Gao, Jinxin Shi, Ping Chen, Zhenning Wang
Department of Surgical Oncology and General Surgery, First Hospital of China Medical University, Shenyang, People’s Republic of China
*These authors contributed equally to this work
Abstract: Over the last two decades, genome-wide studies have revealed that only a small fraction of the human genome encodes proteins; long noncoding RNAs (lncRNAs) account for 98% of the total genome. These RNA molecules, which are >200 nt in length, play important roles in diverse biological processes, including the immune response, stem cell pluripotency, cell proliferation, apoptosis, differentiation, invasion, and metastasis by regulating gene expression at the epigenetic, transcriptional, and posttranscriptional levels. However, the detailed molecular mechanisms underlying lncRNA function are only partially understood. Recent studies showed that many lncRNAs are aberrantly expressed in gastric cancer (GC) tissues, gastric juice, plasma, and cells, and these alterations are linked to the occurrence, progression, and outcome of GC. Here, we review the current knowledge of the biological functions and clinical aspects of lncRNAs in GC.
Keywords: long noncoding RNA, gastric cancer, biomarker, target therapy
Introduction
Gastric cancer (GC) is one of the most frequently diagnosed gastrointestinal neoplasms in East Asia, Eastern Europe, and parts of Central and South America, and the second most lethal malignancy worldwide.1 Owing to a lack of appropriate molecular biomarkers, GC patients are often underdiagnosed. Most cases are diagnosed at an advanced stage, at which point the prognosis is uncertain even with surgery, chemotherapy, and radiotherapy because of the risk of relapse, distant metastasis, and chemoresistance.2 A better understanding of the molecular mechanisms underlying the development of GC may help identify potential diagnostic and prognostic biomarkers and therapeutic targets.
The complete sequencing of the human genome showed that only 1.5%–2% of genes encode proteins and that the remaining genes are transcribed as noncoding RNAs (ncRNAs), which are now known to play important roles in a wide variety of biological processes in both normal development and in disease states.3–5 Based on their functions, ncRNAs are classified as housekeeping or regulatory ncRNAs. The former include ribosomal RNA, transfer RNA, small nuclear RNA, and small nucleolar RNA, and they are constitutively expressed; the latter include short interfering RNAs, piwi-interacting RNAs, microRNAs (miRNAs), and long noncoding RNAs (lncRNAs), and they are expressed in a spatially and temporally restricted manner. Regulatory ncRNAs are divided into two classes based on their length: short/small ncRNAs (<200 nt) and lncRNAs (>200 nt).6 In addition, lncRNAs can be categorized as sense, antisense, bidirectional, intronic, or intergenic depending on their proximity to the nearest protein-coding transcripts.7,8
Recent studies showed that aberrant lncRNA expression is associated with various biological processes, including proliferation, metastasis, migration, and epithelial-to-mesenchymal transition (EMT) in several cancers.9–11 In this review, we briefly summarize the current state of knowledge on the role of lncRNAs in GC. lncRNAs that have been linked to GC cell proliferation and apoptosis are listed in Table 1, and lncRNAs that have been linked to GC cell invasion and metastasis are listed in Table 2.
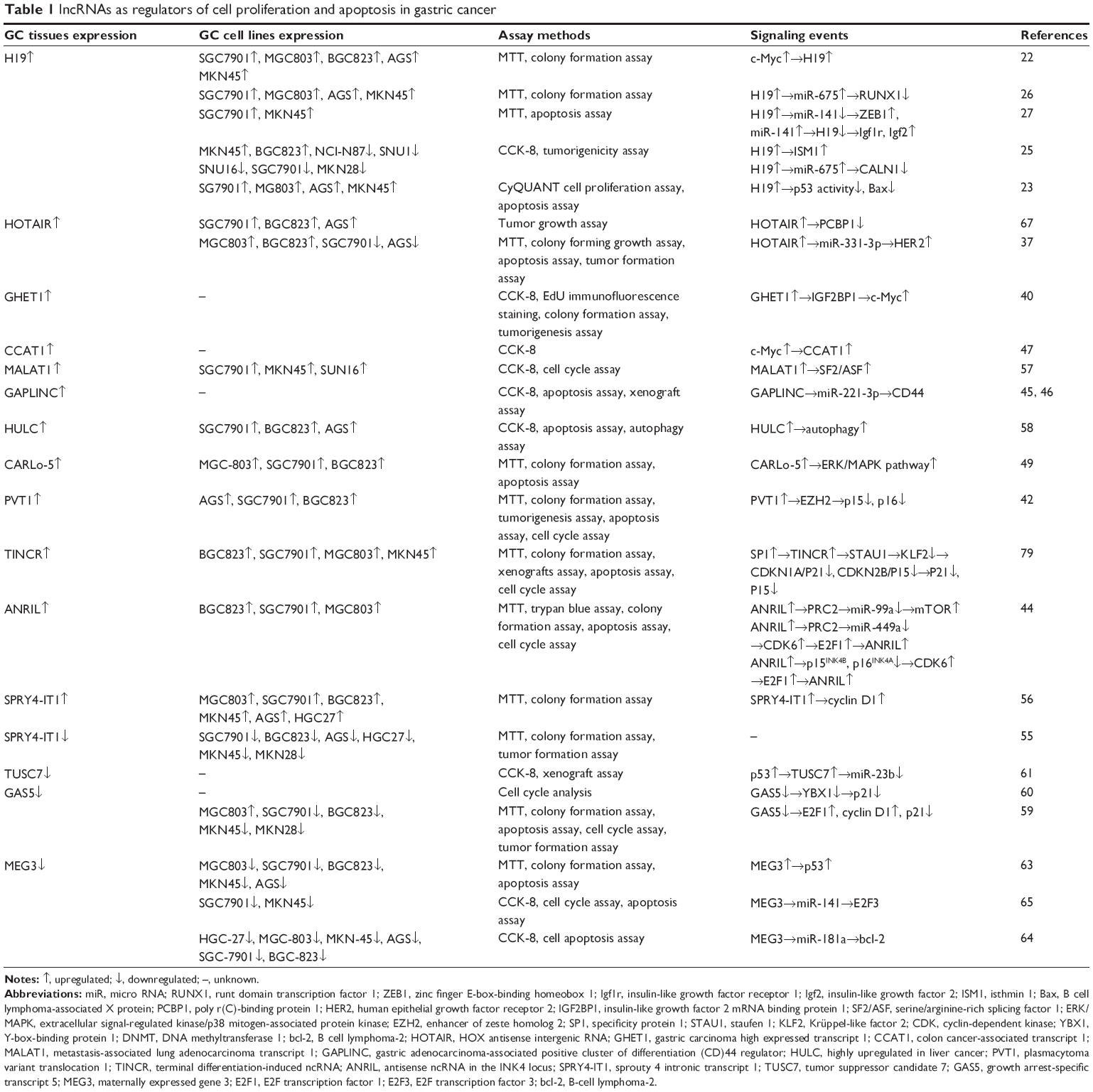 | Table 1 lncRNAs as regulators of cell proliferation and apoptosis in gastric cancer |
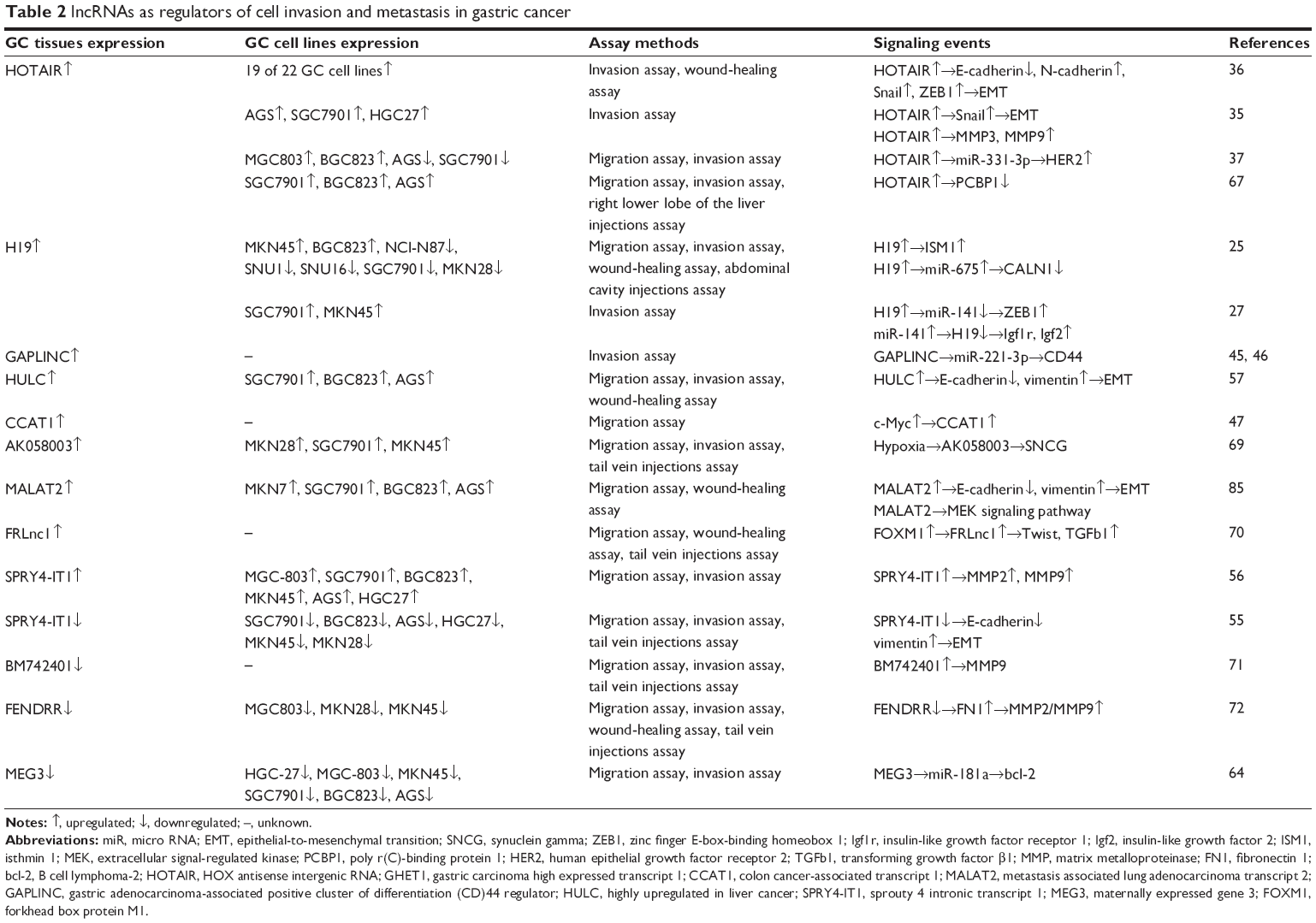 | Table 2 lncRNAs as regulators of cell invasion and metastasis in gastric cancer |
Proliferation and apoptosis
Upregulated lncRNAs
H19
H19 is located on human chromosome 11 (11p15.5) and is highly expressed during embryogenesis.12–15 Aberrant H19 expression is observed in many types of cancer, including esophageal, cervical, bladder, lung, and breast cancers.16–21 H19 was shown to be upregulated in GC relative to normal adjacent tissues (NATs) as well as in five human GC cell lines (MGC-803, BGC-823, SGC-7901, AGS, and MKN-45) compared with the normal gastric epithelial cell line GES-1.22 H19 is considered as an oncogenic RNA that can stimulate cell proliferation, and its overexpression induces GC cell proliferation and inhibits apoptosis.22,23 H19 induces proliferation by inhibiting p53 and suppressing the expression of the p53 target B-cell lymphoma-associated X protein.23 H19 expression is induced by c-Myc, which regulates GC cell proliferation: transfection of the GC cell lines SGC-7901 and BGC-823 with a c-Myc plasmid resulted in a 3.2- and 2.9-fold upregulation of H19, respectively.22
H19 function as the precursor of microRNA (miR)-675.24 Both are overexpressed in GC tissues and promote cell proliferation in vitro and in vivo. H19 knockdown resulted in greater inhibition of cell proliferation, implying a mechanism other than one involving miR-675. H19 regulates isthmin (ISM)1 directly and CALN1 indirectly via miR-675 to promote cell proliferation.25 Runt domain transcription factor 1, a tumor suppressor, was shown to be a direct target of miR-675.26
H19 and miR-141 act as competing endogenous RNAs (ceRNAs) in GC. H19 is upregulated, while miR-141 is downregulated in GC tissues, and H19 and miR-141 levels are negatively correlated, which is consistent with the fact that miR-141 inhibits cell proliferation. H19 and miR-141 modulate cell proliferation by competing for binding to their target genes insulin-like growth factor (Igf)2, Igf receptor 1, and zinc finger E-box-binding homeobox 1.27
H19 is a well-known lncRNA, and many studies have analyzed its role in GC. H19 modulates the expression of p53, ISM1, CALN1, miR675, and miR141 in GC. However, other proteins or lncRNAs are regulated by H19 in other cancers, such as EGR1, 17β-estradiol, and HNF1A-AS1 among others.28–30 The H19 network should be further investigated in the future.
HOX antisense intergenic RNA
HOX antisense intergenic RNA (HOTAIR) was first identified in breast cancer and is associated with metastasis and poor survival.31 It has been implicated in tumorigenesis in lung, pancreatic, liver, and gastric cancers.32–38 HOTAIR was found to be overexpressed in GC relative to NAT as well as in 19 of 22 GC cell lines as compared to normal gastric RNA.36 However, the expression of HOTAIR in SGC-7901 cells is controversial, and Liu et al37 showed that HOTAIR is downregulated in SGC-7901 cells. However, to confirm these results, assessment of all cell lines and SGC-7901 cells should be performed to ensure lack of contamination. In a soft agar assay, cancer cells with high levels of HOTAIR formed larger colonies than those expressing low levels of the protein.39 HOTAIR knockdown inhibited cell proliferation in KATO III, MKN74, and MKN28, but not in AGS cells, by arresting the cell cycle at the G0/G1 phase;36 however, these results are controversial, since another study found that HOTAIR did not influence MKN74 or KATO III cell proliferation.39
Recent studies showed that HOTAIR promotes cell proliferation and inhibits apoptosis in vitro and in vivo. It has been shown to act as a sponge for miR-331-3p, which suppresses GC cell proliferation, thereby relieving the inhibition of HER2 by miR-331-3p.37
Gastric carcinoma high expressed transcript 1
Gastric carcinoma high expressed transcript (GHET)1 is upregulated in GC relative to NAT and was shown to promote cell proliferation in vitro by using the Cell Counting Kit 8 and colony formation assay and by ethynyl deoxyuridine incorporation. GHET1 also promotes xenograft tumor growth in vivo. RNA immunoprecipitation and pull-down experiments demonstrated a specific association between GHET1 and Igf2 mRNA-binding protein (BP)1; GHET1 modulates the physical interaction between c-Myc mRNA and Igf2BP1 by binding the latter, resulting in increased c-Myc expression, which in turn promotes cell proliferation.40 Heterogeneous nuclear ribonucleoprotein U (HNRNPU), synaptotagmin binding, cytoplasmic RNA interacting protein (SYNCRIP), Y-box binding protein 1 (YBX1), and DEAH (Asp–Glu–Ala–His) box helicase 9 (DHX9) were suggested to cooperate with IGF2BP1 in promoting the stabilization of c-Myc mRNA.41 The relationship between GHET1, c-Myc mRNA, and the RNA-BPs (mentioned earlier) requires further investigation.
Plasmacytoma variant translocation 1
Plasmacytoma variant translocation (PVT)1 expression is upregulated in GC tissues, and PVT1 knockdown in SGC-7901 and BGC-823 cells suppresses proliferation by inducing G1 arrest and apoptosis and affects tumorigenesis in vivo. PVT1 was implicated in epigenetic regulation through association with enhancer of zeste homologue (EZH)2, a subunit of the polycomb repressive complex (PRC)2. Moreover, p15 and p16, which control cell cycle progression, are silenced by overexpression of PVT1, resulting in cell cycle arrest via EZH2 recruitment. Therefore, PVT1 along with EZH2 regulates p15 and p16 to promote GC cell proliferation.42 These studies indicate that PVT1 plays a role in PRC2-mediated epigenetic regulation and is thus involved in the progression of GC.
Terminal differentiation-induced ncRNA
Terminal differentiation-induced ncRNA (TINCR) is upregulated in GC tissues, and gain- and loss-of-function studies showed that it promotes cell growth by arresting cells at G0–G1 phase and inducing apoptosis. The nuclear transcription factor specificity protein 1 increases the expression of TINCR, which recruits and binds staufen (STAU)1 to form a complex that binds to the 3′-untranslated region of Kruppel-like factor (KLF)2 mRNA, thereby decreasing its stability and expression. The consequent degradation of KLF2 downregulates the cell cycle inhibitory genes cyclin-dependent kinase (CDK)N1A/P21 and CDKN2B/P15. These results demonstrate that TINCR indirectly regulates CDKN2B/P15 and CDKN1A/P21 at the posttranscriptional level.43
Antisense ncRNA in the INK4 locus
Antisense ncRNA in the INK4 locus (ANRIL; also known as CDKN2B-AS1) is a 3.8 kb lncRNA that is upregulated in GC tissues. ANRIL knockdown induces cell apoptosis and arrests cells at G1–G0 phase through a mechanism involving epigenetic silencing of p15INK4B and p16INK4A via EZH2 binding and H3K27 trimethylation. ANRIL may also epigenetically regulate the expression of miR-99a/miR-449a by binding to PRC2; ANRIL overexpression leads to the downregulation of p15INK4B, p16INK4A, and miR-449a. Since p15INK4B and p16INK4A are inhibitors of CDK6, a target of miR-449a, this results in an increase in CDK6 expression, dephosphorylation of retinoblastoma protein, and release of E2F1 from inhibition, which induces ANRIL expression. This positive feedback loop promotes GC cell proliferation.44 ANRIL, as a member of PRC2-mediated epigenetic regulation, is involved in the development of GC. Moreover, the crosstalk between ANRIL and miRNAs at the epigenetic level is an important discovery.
Gastric adenocarcinoma-associated positive cluster of differentiation 44 regulator, long intergenic ncRNA
Gastric adenocarcinoma-associated positive cluster of differentiation (CD)44 regulator, long intergenic ncRNA (GAPLINC) is overexpressed in GC relative to NAT and is associated with increased proliferation in vitro and in vivo. A strong correlation between GAPLINC and CD44 expression was reported. MiR-211-3p is a target of both GAPLINC and CD44, which compete for binding to this miRNA; miR-211-3p downregulation inhibits the degradation of CD44 mRNA and increases translation of the protein. Thus, GAPLINC in conjunction with CD44 and miR-211-3p promotes cancer cell proliferation.45,46
Colon cancer-associated transcript 1
Colon cancer-associated transcript (CCAT)1 is overexpressed in GC tissues.47,48 A correlation has been observed between CCAT1 and c-Myc mRNA expression; c-Myc binds directly to E-box elements in the CCAT1 promoter to induce its expression. Gain- and loss-of-function approaches showed that CCAT1 promotes the proliferation of AGS and MKN45 cells.47
CARLo-5
CARLo-5 levels are higher in the BGC-823, MGC-803, and SGC-7901 cell lines than in GES-1 cells, and CARLo-5 knockdown in the latter inhibits proliferation by inducing apoptosis and G0/G1 arrest. CARLo-5 knockdown also leads to the dephosphorylation and inhibition of extracellular signal-regulated kinase (ERK) and p38 mitogen-associated protein kinase (MAPK), indicating that CARLo-5 regulates cell proliferation and apoptosis via modulation of ERK/MAPK signaling.49
Sprouty 4 intronic transcript 1
Sprouty 4 intronic transcript (SPRY4-IT)1 is highly expressed in melanoma cells, trophoblast cells, clear cell renal cell carcinoma, and esophageal squamous cell carcinoma,50–53 and downregulated in non-small-cell lung cancer.54 Its expression and function in GC is controversial. Xie et al55 confirmed that SPRY4-IT1 is downregulated in GC and represses cell proliferation in the SGC-7901 and BGC-823 cell lines in vitro and tumorigenesis in vivo. However, Peng et al56 found that SPRY4-IT1 is significantly overexpressed in GC tissues. MKN45 cell proliferation and colony formation were suppressed by SPRY4-IT1 knockdown via a mechanism that likely involves the regulation of cyclin D1.
In the future, additional studies should be performed with a larger sample size and other types of cancer cells to investigate the function of SPRY4-IT1.
Metastasis-associated lung adenocarcinoma transcript 1
Metastasis-associated lung adenocarcinoma transcript (MALAT)1 and serine/arginine-rich splicing factor 1 (SF2/ASF) were found to be upregulated in the SGC-7901, MKN-45, and SUN-16 GC cell lines relative to the levels in GES-1 cells. MALAT1 knockdown resulted in the downregulation of SF2/ASF and induced SGC-7901 cell cycle arrest at G0/G1 phase, thereby inhibiting proliferation. SF2/ASF acts downstream and is a target of MALAT1. Thus, MALAT1 acts as an oncogene in human GC and is a potential therapeutic target.57
Highly upregulated in liver cancer
Overexpression of highly upregulated in liver cancer (HULC) promotes the proliferation of SGC7901 cells. Interestingly, the level of microtubule-associated protein 1 light chain 3-II, an indicator of autophagy, was increased following HULC overexpression, suggesting that HULC stimulates autophagy in these cells, thereby inhibiting apoptosis and contributing to proliferation.58
Downregulated lncRNAs
Growth arrest-specific transcript 5
Growth arrest-specific transcript (GAS)5 is downregulated in GC tissues and cell lines, including SGC7901, BGC823, MKN45, and MKN28. GAS5 overexpression suppresses cell proliferation and promotes apoptosis in vitro and inhibits tumorigenesis in vivo, whereas knockdown of GAS5 induces the expression of E2F1 and cyclin D1 and inhibits that of p21. Overexpression of E2F1 induces tumorigenesis by stimulating cell proliferation and p21 expression.59 GAS5 was also shown to bind to the transcriptional activator YBX1 by an RNA pull-down assay; GAS5 knockdown reduced YBX1 protein level by accelerating its degradation, leading to the downregulation of p21 and progression through the G1 phase of the cell cycle. YBX1 plays a critical role in the GAS5-mediated regulation of the GAS5/YBX1/p21 pathway, which regulates the cell cycle and modulates GC cell proliferation.60
Tumor suppressor candidate 7
Tumor suppressor candidate (TUSC)7 is downregulated in GC as compared to NAT and inhibits cell growth in vitro and in vivo. TUSC7 is activated by p53 through p53-responsive elements in its promoter. In addition, a mutually repressive interaction between TUSC7 and miR-23b has been reported. The activation of TUSC7 by p53 plays a key role in cell growth inhibition through the suppression of miR-23b in GC.61
Maternally expressed gene 3
Maternally expressed gene (MEG)3 expression is downregulated in GC relative to NAT, and its expression is lower in SGC7901, AGS, MGC803, MKN45, and MKN28 cells than in GES-1 cells. miR-148a stimulates MEG3 by inhibiting DNA methyltransferase 1, thereby suppressing cell proliferation and growth.62 Another study showed that MEG3 inhibits cell proliferation by activating p53 signaling in GC.63 MEG3 functions as a ceRNA by competitively binding miR-181a to regulate Bcl-2 and inhibit cell proliferation.64 Another research reported by Zhou et al65 indicated that MEG3 is positively correlated with miR-141 and inversely correlated with E2F3.
Invasion and metastasis
Upregulated lncRNAs
HOTAIR
Knockdown of HOTAIR inhibits cell invasion, motility, and migration in vitro.35–37,66,67 On the other hand, the overexpression of HOTAIR in a mouse model induced metastasis and peritoneal dissemination.39 Xu et al35 found that HOTAIR could inhibit cell invasion by decreasing the expression of matrix metalloproteinase (MMP)1 and 3, and loss of HOTAIR reversed EMT by suppressing Snail expression. Liu et al37 elucidated the mechanism by which HOTAIR regulates the expression of Snail. They found that HOTAIR could recruit the PRC2 complex to silence miR34a, thereby inhibiting its expression. First, Snail is a target gene of miR34a, and the downregulation of miR34a could directly promote Snail translation. Second, miR34a could indirectly induce Snail gene transcription via facilitating C-Met transcription.68 Another study showed that HOTAIR could promote GC metastasis by repressing poly r(C)-binding protein (PCBP)1. They confirmed a direct interaction between HOTAIR and PCBP1 by RNA immunoprecipitation experiments.67 Similar to the mechanism by which it regulates proliferation, HOTAIR regulates HER2 via sponging miR-331-3p.37
H19
H19 not only promotes GC cell proliferation, but also enhances GC metastasis. Similar to the mechanism by which it regulates proliferation, H19 controls ISM1 directly and CLAN1 indirectly by modulating miR-675, thereby promoting cell invasion and migration.25 In addition, miR-141 binds H19 as a ceRNA to regulate target genes involved in cell invasion.27
GAPLINC
Similar to its effect on cell proliferation, GAPLINC in conjunction with CD44 and miR-211-3p promotes cancer cell migration and GC invasion.45,46
HULC
HULC is not only involved in GC cell proliferation, but also promotes cell invasion and blocks EMT. HULC promotes SGC-7901 cell migration and invasion in vitro, while HULC knockdown reverses EMT through the modulation of E-cadherin and vimentin expression.58
AK058003
AK058003 is overxpressed in GC tissues, and AK058003 knockdown suppresses SGC7901 and MKN45 cell migration, invasion, and motility. GC cell migration and invasion were shown to increase under hypoxic relative to normoxic conditions; however, this effect was lost upon AK058003 knockdown. In addition, low levels of AK058003 expression are linked to a decrease in the number and size of lung and liver metastatic nodules in vivo. Synuclein gamma (SNCG), a metastasis-related gene, is upregulated under conditions of hypoxia and is an effector of hypoxia-induced GC metastasis, whereas loss of AK058003 decreases SNCG expression via methylation of the SNCG promoter.69
FRLnc1
FRLnc1 expression is inhibited by Forkhead box protein (FOX)M1 knockdown in MGC803 and AGS cells. Transfection of siFRLnc1- and FRLnc1-overexpressing lentiviruses promoted cell migration. Moreover, in vivo overexpression by direct injection of SGC7901-FRLnc1-expressing cells into mice revealed a role in pulmonary metastasis. The regulation of transforming growth factor β1 and Twist was found to be regulated by FRLnc1, thus mediating its role in cell migration and distant tumor metastasis.70
SPRY4-IT1
SPRY4-IT1 promotes cell migration and invasion. SPRY4-IT1 knockdown strongly inhibits migration and invasion in vitro via regulating MMP2 and MMP9 expression.56 However, other studies showed that SPRY4-IT1 plays a role in the inhibition of GC cell migration and invasion and the EMT process in vitro, and cell metastasis in vivo.55
Downregulated lncRNAs
MEG3
Knockdown of MEG3 inhibits cell invasion, motility, and migration in vitro. MEG3 upregulates Bcl-2 by competitively binding miR-181a, which is similar to the mechanism by which it regulates cell proliferation.64
BM742401
BM742401 is downregulated in GC relative to NAT, and its overexpression inhibits the migration and invasion of AGS and MKN-1 cells and suppresses metastasis in vivo, a process involving MMP9.71 Further studies are required to clarify the underlying molecular mechanism and to identify the effector molecules that interact directly and indirectly with BM742401.
FENDRR
FENDRR is expressed at low levels in GC relative to NAT, and its expression is lower in MKN28, MKN45, and MGC803 cells than in GES-1 cells. Treatment with the histone deacetylase inhibitor trichostatin A altered FENDRR expression. FENDRR suppresses GC cell metastasis in vitro and in vivo, and a negative correlation between fibronectin (FN)1 and FENDRR expression was reported. FENDRR likely inhibits cell migration and invasion by suppressing the levels of MMP2 and MMP9 and FN1.72 Further insight into the function and clinical application of FENDRR and its regulation targets FN1 and MMP2/MMP9 may be helpful in designing treatment strategies for GC.
Clinical applications of lncRNAs in GC
GC is one of the most common gastrointestinal malignant tumors worldwide, with an overall survival (OS) rate of 20%–25%.73,74 Patients are often diagnosed at late stages of the disease, underscoring the need to identify new biomarkers that would allow early detection before metastasis has occurred. Aberrant expression of GC-specific DNAs, mRNAs, miRNAs, and lncRNAs can be detected in body fluids, including plasma or serum, gastric juice, and urine, which can aid in the early diagnosis of GC.75–77 We briefly summarize the current state of knowledge on the role of lncRNAs in GC. lncRNAs that have been linked to GC prognosis and diagnosis are listed in Table 3.
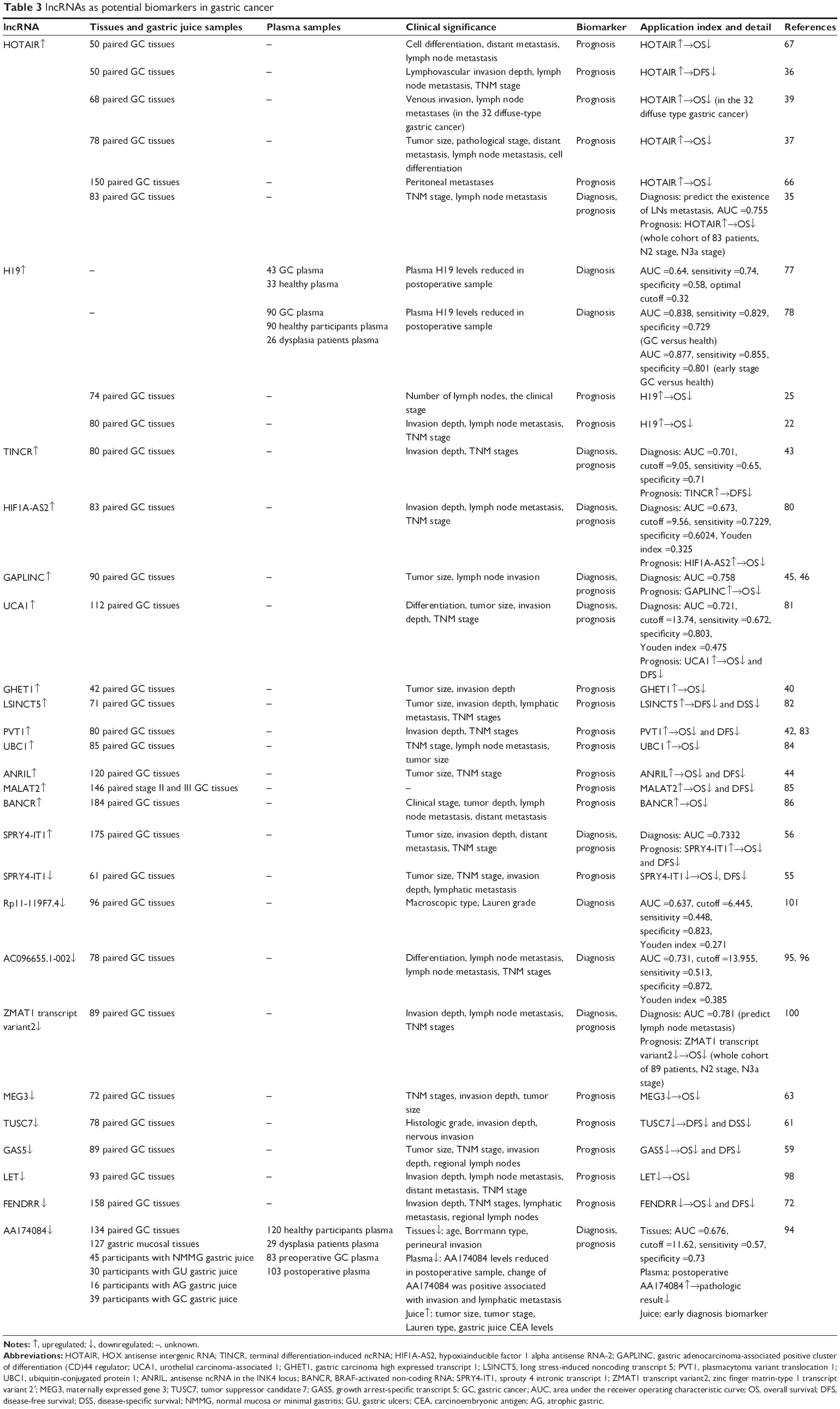 | Table 3 lncRNAs as potential biomarkers in gastric cancer |
Upregulated lncRNAs
HOTAIR expression is associated with tumor size, pathological stage, distant and lymph node metastasis, and tumor cell differentiation, as well as lymphovascular invasion.35–38,67 Another study confirmed that HOTAIR expression levels predict lymph node metastasis, as determined by an area under the receiver operating characteristic (ROC) curve (AUC) of 0.755.35 HOTAIR expression also predicts poor patient outcome, with higher levels associated with a worse prognosis.35–37,67
Plasma H19 levels are higher in GC patients than in healthy controls; ROC curve analysis showed that the AUC was 0.64, with a sensitivity and specificity of 0.74 and 0.58, respectively. In addition, postoperative plasma H19 levels were decreased relative to the preoperative levels.77 This was confirmed in another study which showed that plasma H19 levels can differentiate early-stage GC from healthy patients, with an AUC of 0.877 and a sensitivity and specificity of 0.855 and 0.801, respectively.78 As with HOTAIR, patients with higher H19 levels had a worse prognosis.22,25
TINCR, a 3.7 kb lncRNA, is downregulated in human squamous cell carcinoma,79 whereas it is highly upregulated in GC relative to NAT. TINCR expression level was associated with the degree of invasiveness and tumor–node–metastasis (TNM) stage, and it may be a diagnostic and prognostic biomarker in GC patients, with an AUC of 0.701 and a sensitivity and specificity of 0.65 and 0.71, respectively. The Kaplan–Meier analysis and log-rank test indicated that GC patients with high TINCR expression had higher recurrence rates, suggesting that it is an indicator of disease-free survival (DFS) in GC.43
HIF1A-AS2 overexpression in GC tissues was found to be closely correlated with TNM stage, tumor invasion, and lymph node metastasis, with an AUC of 0.673, and a sensitivity and specificity of 0.7229 and 0.6024, respectively. Kaplan–Meier analysis revealed that high levels of HIF1A-AS2 were associated with poor outcome in GC patients.80
GAPLINC, a 924 bp intergenic ncRNA, is highly expressed in GC tissues; patients with high GAPLINC expression have on average larger tumors and more frequent occurrence of lymph node invasion than those with low expression. The AUC was 0.758. In addition, GAPLINC levels are associated with patient survival, supporting its utility as a biomarker for GC diagnosis and prognosis.45
Urothelial carcinoma-associated (UCA)1, which was first identified in urinary bladder cancer tissue and found to be linked to increased tumorigenicity and invasion, is upregulated in GC tissues. UCA1 levels are higher in SGC-7901, BGC-823, MKN-28, and AGS cells than in GES-1 cells. UCA1 levels were associated with cancer differentiation, tumor size, invasion, and TNM stage. The levels of UCA1 in gastric juice were found to be higher in GC patients than in normal individuals, with an AUC of 0.721 and a sensitivity and specificity of 0.672 and 0.803, respectively. Kaplan–Meier analysis showed that increased UCA1 expression contributes to poor OS and DFS in GC patients, whereas multivariate survival analysis showed that UCA1 is an independent prognostic marker for GC.81
GHET1 is overexpressed in GC tissues and is correlated with tumor size and invasion, as well as GC patient outcome, with high GHET1 levels associated with short OS.40
Long stress-induced noncoding transcript (LSINCT)5 was found to be overexpressed in GC relative to NAT as well as in five GC cell lines relative to GES-1 cells. LSINCT5 levels are associated with tumor size, tumor invasion, lymphatic metastasis, and TNM stage. Patients with high levels of LSINCT5 have worse outcomes, including shorter OS and DFS, than those with lower LSINCT5 expression.82
PVT1 is upregulated in GC, and its expression is correlated with lymph node invasion and TNM stage. PVT1 is associated with poor prognosis, as GC patients with high PVT1 expression levels have worse OS and DFS than those exhibiting low PVT1 levels. Uni- and multivariate survival analyses indicated that PVT1 expression is an independent prognostic factor for GC.42,83
E2 ubiquitin-conjugated protein (UBC)1 is upregulated in GC, and high levels of UBC1 are associated with poor prognosis in GC as well as with lymph node metastasis, tumor size, and TNM stage.84
ANRIL is a 3.8 kb ncRNA that is upregulated in GC tissues relative to NAT in 77.5% of cases and is strongly associated with advanced TNM stage and tumor size. GC patients with low levels of expression of ANRIL have better OS and DFS than those with high levels.44
MALAT2 overexpression in GC tissues is correlated with lymph node metastasis and tumor stage, as well as with shorter DFS.85
BRAF-activated noncoding RNA (BANCR) levels are higher in GC than in NAT, and its expression is associated with clinical stage, tumor depth, and lymph node and distant metastasis. Kaplan–Meier analysis and log-rank test showed that higher BANCR expression in GC tissues is associated with shorter OS in GC patients.86
GACAT3, also known as AC130710, is upregulated in GC relative to NAT, and its expression levels are associated with tumor size, TNM stage, distant metastasis, and tissue carcinoembryonic antigen (CEA) expression level.87,88
LINC00152 expression in gastric juice, plasma, and tissue may also provide useful information for the diagnosis of GC. Plasma and gastric juice LINC00152 levels are higher in GC patients than in normal controls. ROC curve analysis revealed an AUC of 0.657, with a sensitivity and specificity of 0.481 and 0.852, respectively. Postoperative plasma LINC00152 levels are higher than preoperative levels, and LINC00152 upregulation in GC tissues is correlated with greater invasiveness, with an AUC of 0.645 and a sensitivity and specificity of 0.625 and 0.681, respectively.89
SPRY4-IT1 expression was shown to be elevated in GC compared with that in NAT as well as in six GC cell lines relative to GES-1 cells. Moreover, SPRY4-IT1 expression is positively correlated with tumor size, invasion, distant metastasis, and TNM stage. ROC curve analysis revealed an AUC of 0.7332, and GC patients with higher SPRY4-IT1 expression had worse prognosis. SPRY4-IT1 expression is an independent prognostic factor for OS and DFS and may also be a useful diagnostic and prognostic marker in GC patients.56 In contrast with these findings, Xie et al55 found lower expression of SPRY4-IT1 in GC tissues compared with NAT and also demonstrated an association with poor prognosis.
HULC is overexpressed in GC tissues and is associated with lymph node and distant metastasis and TNM stage with an AUC of 0.769 and a sensitivity and specificity of 0.707 and 0.724, respectively. Therefore, HULC is a novel potential prognostic biomarker in GC.58
Small ubiquitin-like modifier 1 pseudogene (SUMO1P)3 can also provide useful information for GC diagnosis; SUMO1P3 levels are higher in GC than in NAT, and its expression is associated with tumor size, differentiation, lymphatic metastasis, and invasion. The AUC was 0.666, with a sensitivity and specificity of 0.659 and 0.636, respectively.90
ABHD11-AS1 was found to be overexpressed in 64% of GC tissue samples compared with NAT in one study; this was associated with differentiation, histological classification, and CA19-9 levels. The AUC was 0.613 and sensitivity and specificity were 0.67 and 0.64, respectively.91
ncRuPAR expression is higher in GC than in NAT, and its expression is associated with TNM stage, tumor invasiveness, lymph node and distant metastasis, and tumor size with an AUC of 0.84; therefore, ncRuPAR can serve as a biomarker to differentiate GC from normal tissue.92
Downregulated lncRNAs
Fer-1-like protein (FER1L)4 expression in GC tissues is linked to tumor diameter, differentiation, general classification, invasion, lymphatic and distant metastasis, TNM stage, vessel or nerve invasion, and serum levels of the tumor marker carbohydrate antigen (CA)72-4. The AUC was 0.778, and sensitivity and specificity were 0.672 and 0.803, respectively. Postoperative plasma FER1L4 levels are reduced relative to preoperative levels.93
AC138128.1, a 1,981 nt antisense lncRNA, is located on chromosome 19 along with FBJ murine osteosarcoma viral oncogene homologue B. Its expression is decreased in 70% of GC samples compared with NAT specimens; on average, the level was 0.548-fold lower in cancerous tissue, with an AUC of 0.688.94
AA174084 levels in the gastric juice of GC patients are higher than those in the normal mucosa or in patients with minimal gastritis, gastric ulcers, or atrophic gastritis. The AUC was 0.848, with a sensitivity and specificity of 0.46 and 0.93, respectively. AA174084 expression in gastric juice is associated with tumor size, tumor stage, histological type, and gastric juice CEA levels. In addition, plasma AA174084 levels decline by 76% postoperatively compared with preoperative levels in GC patients; this reduction is associated with invasion and lymphatic metastasis, while high postoperative plasma AA174084 levels are linked to poor prognosis. On the other hand, AA174084 expression was found to be lower in GC tissues than in NAT, with an AUC of 0.676 and a sensitivity and specificity of 0.57 and 0.73, respectively. Tissues AA174084 levels are associated with various clinicopathologic factors, including age, Borrmann type, and perineural invasion. Therefore, AA174084 is a candidate biomarker for early diagnosis and for predicting prognosis in GC.95
MEG3 expression is lower in GC tissues than in NAT, and MEG3 level is correlated with tumor size, TNM stage, and invasion. Kaplan–Meier survival analysis and log-rank test revealed that lower MEG3 expression is correlated with worse prognosis in GC patients.63
Gastric cancer-associated transcript (GACAT)1, also known as AC096655.1-002, is downregulated in GC tissues, and its expression is associated with lymph node and distant metastasis, TNM stage, and differentiation, suggesting that it plays an important role in GC metastasis. The AUC was 0.731, with a sensitivity and specificity of 0.513 and 0.872, respectively. Therefore, GACAT1 expression can predict GC progression.96,97
GACAT2, also known as HMlincRNA717, is downregulated in GC compared with NAT as well as in five GC cell lines relative to GES-1 cells. Its expression was found to be associated with distant metastasis and venous and perineural invasion.88,98
LET expression is reduced in several cancers including hepatocellular carcinoma, cervical and gallbladder cancers, and GC. Kaplan–Meier analysis and log-rank test showed that lower LET expression levels are associated with decreased OS. In addition, a Cox proportional hazards model showed that LET expression was an independent prognostic marker for predicting poor outcome in GC patients.99
GAS5 was also a prognostic biomarker for GC. Its levels were found to be lower in GC tissues than in NAT in 89% of cases. In addition, GAS5 expression was closely correlated with tumor size and pathological stage. Patients with higher GAS5 levels have longer OS and DFS. GAS5 expression was also an independent risk factor for GC prognosis.59
TUSC7 is downregulated in GC relative to NAT, and TUSC7 levels are associated with histological grade and tumor invasion, including invasion of the nervous system. Patients with high levels of TUSC7 show longer disease-specific survival and DFS, indicating that TUSC7 is a prognostic marker for GC.61
FENDRR expression is lower in GC than in NAT and is correlated with tumor invasion, tumor stage, and lymphatic metastasis. Patients with high FENDRR expression have a lower recurrence rate and longer OS than those with low FENDRR expression. Uni- and multivariate analyses showed that low FENDRR levels were an independent prognostic factor for OS and DFS.72
AI364715 is downregulated in GC relative to NAT and gastric precancerous lesions, and AI364715 expression is associated with tumor size, differentiation, and venous invasion. Poorly differentiated GC and a large tumor size are correlated with poor prognosis, and AI364715 expression also serves as a potential biomarker for GC prognosis.100
Zinc finger matrin-type (ZMAT)1 transcript variant 2 is downregulated in GC tissues compared with NAT, and its expression was associated with tumor invasion, lymph node metastasis, and TNM stage. Real-time reverse transcriptase polymerase chain reaction (RT-PCR) analysis showed that ZMAT1 transcript variant 2 expression was 70.3% lower in metastatic than in matched nonmetastatic lymph nodes, indicating that it could be a biomarker for predicting lymph node metastasis. The AUC was 0.781, and patients at N2 and N3a stages exhibiting higher levels of ZMAT1 transcript variant 2 had better OS than those with lower expression.101
RP11-119F7.4, an antisense lncRNA located on chromosome 10 with a length of 349 bp, was found to be downregulated in GC tissues compared with NAT, with an AUC of 0.637 and a sensitivity and specificity of 0.448 and 0.823, respectively. RP11-119F7.4 expression level was associated with macroscopic tumor type, histological grade, and invasion into lymphatic vessels.102
Conclusion
lncRNAs regulate gene expression at the transcriptional, posttranscriptional, and epigenetic levels, and they are implicated in the occurrence, development, and progression of GC. Dysregulation of lncRNAs in GC is associated with tumor size, macroscopic type, histological grade, tumor invasion, and metastasis. A broad range of lncRNAs have been identified as potential markers for the early detection of GC and for predicting patient outcome, with some already being used in clinical trials. The utility of lncRNAs for cancer diagnosis and prognosis and as therapeutic targets requires further exploration; this knowledge can contribute to the development of more effective lncRNA-based therapies for the treatment of GC.
Future directions
lncRNAs have recently attracted the interest of researchers worldwide. Some lncRNAs have been suggested for use in clinical applications, such as diagnosis, prognosis, and treatment.103 However, the relationship between lncRNAs and GC has only recently begun to be studied in detail.
Health organizations from many countries have focused on preventing the occurrence of GC. Helicobacter is regarded as the pivotal causative agent of gastritis and GC.104 However, there are currently few studies analyzing the relation of the immune response to Helicobacter pylori infection with lncRNAs. Mizrahi et al48 reported that CCAT1 is upregulated in GC tissues compared with NATs, and they further studied the relationship between H. pylori infection and GC. The results showed no significant differences in CCAT1 expression between H. pylori-negative and -positive patients. Yang et al105 identified 23 upregulated and 21 downregulated lncRNAs from microarray data. Further quantitative RT-PCR was used to evaluate the expression of five lncRNAs, which showed that XLOC_004562, XLOC_005912, and XLOC_000620 were upregulated, whereas XLOC_004122 and XLOC_014388 were downregulated in the gastric mucosal tissues of H. pylori-positive patients. These lncRNAs may provide novel targets for the treatment of H. pylori infection, which could contribute to reducing the incidence of gastritis and GC.105
lncRNAs have more restricted tissue-specific expression than protein-coding transcripts in different types of tissues.106 Therefore, researchers should validate and explore novel lncRNAs that may play a role as biomarkers with high specificity, similar to the use of AFP to diagnose liver cancer with high specificity.107,108 Furthermore, lncRNAs play a role in the occurrence, development, and progression of GC.109 Therefore, the expression levels and functions of lncRNAs differ during the different stages of GC. In particular, in the early stages of GC, when tumor sizes are too small for accurate detection using imaging modalities, the levels of lncRNAs could be of value to distinguish patients with early GC from healthy individuals. In the future, researchers should use large sample sizes to verify the utility of lncRNAs as biomarkers in large cohorts. As a noninvasive method, measuring the expression levels of lncRNAs in plasma, gastric juice, and urine could be a new novel strategy to screen cancer patients and healthy individuals.110,111
The identification of therapeutic targets is still a new field, and more studies and efforts in the future are needed to explore the function and molecular mechanism of lncRNAs. The interaction network of lncRNA–miRNA–protein provides additional information and provides novel ideas for GC-targeted treatments. New therapeutic targets of lncRNAs can be identified for drug development. However, the delivery of lncRNAs into cancer cells directly is difficult by conventional RNA interference (RNAi) methods because of the large size and extensive secondary structures of these lncRNAs. Screening for appropriate therapeutic targets and targeting them to cancer cells with high specificity should be the research strategy in the future.
Acknowledgments
This work was supported by National Science Foundation of China (numbers 81201888, 81372549 and 81172370), the Special Prophase Program for National Key Basic Research Program of China (No. 2014CB560712), and Clinical Capability Construction Project for Liaoning Provincial Hospitals (LNCCC-A01-2014).
Author contributions
Jiajun Wang and Jingxu Sun contributed equally to this work. Jun Wang and Yongxi Song contributed to drafting and editing of the manuscript. Peng Gao and Jinxin Shi participated in the conception of the idea. Jiajun Wang and Ping Chen contributed to literature search. Zhenning Wang participated in the conception and coordination. All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Herszenyi L, Tulassay Z. Epidemiology of gastrointestinal and liver tumors. Eur Rev Med Pharmacol Sci. 2010;14(4):249–258. | ||
Saka M, Morita S, Fukagawa T, Katai H. Present and future status of gastric cancer surgery. Jpn J Clin Oncol. 2011;41(3):307–313. | ||
Consortium EP. An integrated encyclopedia of DNA elements in the human genome. Nature. 2012;489(7414):57–74. | ||
Hattori M. Finishing the euchromatic sequence of the human genome. Tanpakushitsu Kakusan Koso. 2005;50(2):162–168. Japanese. | ||
Perez DS, Hoage TR, Pritchett JR, et al. Long, abundantly expressed non-coding transcripts are altered in cancer. Hum Mol Genet. 2008;17(5):642–655. | ||
Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. | ||
Han D, Wang M, Ma N, Xu Y, Jiang Y, Gao X. Long noncoding RNAs: novel players in colorectal cancer. Cancer Lett. 2015;361(1):13–21. | ||
Jia H, Osak M, Bogu GK, Stanton LW, Johnson R, Lipovich L. Genome-wide computational identification and manual annotation of human long noncoding RNA genes. RNA. 2010;16(8):1478–1487. | ||
Kaikkonen MU, Lam MT, Glass CK. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc Res. 2011;90(3):430–440. | ||
Li M, Izpisua Belmonte JC. Roles for noncoding RNAs in cell-fate determination and regeneration. Nat Struct Mol Biol. 2015;22(1):2–4. | ||
Zhang H, Chen Z, Wang X, Huang Z, He Z, Chen Y. Long non-coding RNA: a new player in cancer. J Hematol Oncol. 2013;6:37. | ||
Matouk I, Ayesh B, Schneider T, et al. Oncofetal splice-pattern of the human H19 gene. Biochem Biophys Res Commun. 2004;318(4):916–919. | ||
Zemel S, Bartolomei MS, Tilghman SM. Physical linkage of two mammalian imprinted genes, H19 and insulin-like growth factor 2. Nat Genet. 1992;2(1):61–65. | ||
Pachnis V, Brannan CI, Tilghman SM. The structure and expression of a novel gene activated in early mouse embryogenesis. EMBO J. 1988; 7(3):673–681. | ||
Poirier F, Chan CT, Timmons PM, Robertson EJ, Evans MJ, Rigby PW. The murine H19 gene is activated during embryonic stem cell differentiation in vitro and at the time of implantation in the developing embryo. Development. 1991;113(4):1105–1114. | ||
Hibi K, Nakamura H, Hirai A, et al. Loss of H19 imprinting in esophageal cancer. Cancer Res. 1996;56(3):480–482. | ||
Douc-Rasy S, Barrois M, Fogel S, et al. High incidence of loss of heterozygosity and abnormal imprinting of H19 and IGF2 genes in invasive cervical carcinomas. Uncoupling of H19 and IGF2 expression and biallelic hypomethylation of H19. Oncogene. 1996;12(2):423–430. | ||
Ariel I, Sughayer M, Fellig Y, et al. The imprinted H19 gene is a marker of early recurrence in human bladder carcinoma. Mol Pathol. 2000;53(6):320–323. | ||
Elkin M, Shevelev A, Schulze E, et al. The expression of the imprinted H19 and IGF-2 genes in human bladder carcinoma. FEBS Lett. 1995;374(1):57–61. | ||
Kondo M, Suzuki H, Ueda R, et al. Frequent loss of imprinting of the H19 gene is often associated with its overexpression in human lung cancers. Oncogene. 1995;10(6):1193–1198. | ||
Adriaenssens E, Dumont L, Lottin S, et al. H19 overexpression in breast adenocarcinoma stromal cells is associated with tumor values and steroid receptor status but independent of p53 and Ki-67 expression. Am J Pathol. 1998;153(5):1597–1607. | ||
Zhang EB, Han L, Yin DD, Kong R, De W, Chen J. c-Myc-induced, long, noncoding H19 affects cell proliferation and predicts a poor prognosis in patients with gastric cancer. Med Oncol. 2014;31(5):914. | ||
Yang F, Bi J, Xue X, et al. Up-regulated long non-coding RNA H19 contributes to proliferation of gastric cancer cells. FEBS J. 2012;279(17):3159–3165. | ||
Cai X, Cullen BR. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA. 2007;13(3):313–316. | ||
Li H, Yu B, Li J, et al. Overexpression of lncRNA H19 enhances carcinogenesis and metastasis of gastric cancer. Oncotarget. 2014;5(8):2318–2329. | ||
Zhuang M, Gao W, Xu J, Wang P, Shu Y. The long non-coding RNA H19-derived miR-675 modulates human gastric cancer cell proliferation by targeting tumor suppressor RUNX1. Biochem Biophys Res Commun. 2014;448(3):315–322. | ||
Zhou X, Ye F, Yin C, Zhuang Y, Yue G, Zhang G. The interaction between MiR-141 and lncRNA-H19 in regulating cell proliferation and migration in gastric cancer. Cell Physiol Biochem. 2015;36(4):1440–1452. | ||
Li H, Li J, Jia S, et al. miR675 upregulates long noncoding RNA H19 through activating EGR1 in human liver cancer. Oncotarget. 2015;6(31):31958–31984. | ||
Sun H, Wang G, Peng Y, et al. H19 lncRNA mediates 17beta-estradiol-induced cell proliferation in MCF-7 breast cancer cells. Oncol Rep. 2015;33(6):3045–3052. | ||
Yang X, Song JH, Cheng Y, et al. Long non-coding RNA HNF1A-AS1 regulates proliferation and migration in oesophageal adenocarcinoma cells. Gut. 2014;63(6):881–890. | ||
Gupta RA, Shah N, Wang KC, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076. | ||
Zhuang Y, Wang X, Nguyen HT, et al. Induction of long intergenic non-coding RNA HOTAIR in lung cancer cells by type I collagen. J Hematol Oncol. 2013;6:35. | ||
Kim K, Jutooru I, Chadalapaka G, et al. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene. 2013;32(13):1616–1625. | ||
Ishibashi M, Kogo R, Shibata K, et al. Clinical significance of the expression of long non-coding RNA HOTAIR in primary hepatocellular carcinoma. Oncol Rep. 2013;29(3):946–950. | ||
Xu ZY, Yu QM, Du YA, et al. Knockdown of long non-coding RNA HOTAIR suppresses tumor invasion and reverses epithelial-mesenchymal transition in gastric cancer. Int J Biol Sci. 2013;9(6):587–597. | ||
Lee NK, Lee JH, Park CH, et al. Long non-coding RNA HOTAIR promotes carcinogenesis and invasion of gastric adenocarcinoma. Biochem Biophys Res Commun. 2014;451(2):171–178. | ||
Liu XH, Sun M, Nie FQ, et al. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331–3p in gastric cancer. Mol Cancer. 2014;13:92. | ||
Hajjari M, Behmanesh M, Sadeghizadeh M, Zeinoddini M. Up-regulation of HOTAIR long non-coding RNA in human gastric adenocarcinoma tissues. Med Oncol. 2013;30(3):670. | ||
Endo H, Shiroki T, Nakagawa T, et al. Enhanced expression of long non-coding RNA HOTAIR is associated with the development of gastric cancer. PLoS One. 2013;8(10):e77070. | ||
Yang F, Xue X, Zheng L, et al. Long non-coding RNA GHET1 promotes gastric carcinoma cell proliferation by increasing c-Myc mRNA stability. FEBS J. 2014;281(3):802–813. | ||
Weidensdorfer D, Stohr N, Baude A, et al. Control of c-myc mRNA stability by IGF2BP1-associated cytoplasmic RNPs. RNA. 2009;15(1):104–115. | ||
Kong R, Zhang EB, Yin DD, et al. Long noncoding RNA PVT1 indicates a poor prognosis of gastric cancer and promotes cell proliferation through epigenetically regulating p15 and p16. Mol Cancer. 2015;14(1):82. | ||
Xu TP, Liu XX, Xia R, et al. SP1-induced upregulation of the long noncoding RNA TINCR regulates cell proliferation and apoptosis by affecting KLF2 mRNA stability in gastric cancer. Oncogene. 2015;34(45):5648–5661. | ||
Zhang EB, Kong R, Yin DD, et al. Long noncoding RNA ANRIL indicates a poor prognosis of gastric cancer and promotes tumor growth by epigenetically silencing of miR-99a/miR-449a. Oncotarget. 2014;5(8):2276–2292. | ||
Hu Y, Wang J, Qian J, et al. Long noncoding RNA GAPLINC regulates CD44-dependent cell invasiveness and associates with poor prognosis of gastric cancer. Cancer Res. 2014;74(23):6890–6902. | ||
[No authors]. Correction: long noncoding RNA GAPLINC regulates CD44-dependent cell invasiveness and associates with poor prognosis of gastric cancer. Cancer Res. Epub August 18, 2015. | ||
Yang F, Xue X, Bi J, et al. Long noncoding RNA CCAT1, which could be activated by c-Myc, promotes the progression of gastric carcinoma. J Cancer Res Clin Oncol. 2013;139(3):437–445. | ||
Mizrahi I, Mazeh H, Grinbaum R, et al. Colon Cancer Associated Transcript-1 (CCAT1) expression in adenocarcinoma of the stomach. J Cancer. 2015;6(2):105–110. | ||
Zhang Y, Ma M, Liu W, Ding W, Yu H. Enhanced expression of long noncoding RNA CARLo-5 is associated with the development of gastric cancer. Int J Clin Exp Pathol. 2014;7(12):8471–8479. | ||
Mazar J, Zhao W, Khalil AM, et al. The functional characterization of long noncoding RNA SPRY4-IT1 in human melanoma cells. Oncotarget. 2014;5(19):8959–8969. | ||
Zou Y, Jiang Z, Yu X, et al. Upregulation of long noncoding RNA SPRY4-IT1 modulates proliferation, migration, apoptosis, and network formation in trophoblast cells HTR-8SV/neo. PLoS One. 2013;8(11):e79598. | ||
Zhang HM, Yang FQ, Yan Y, Che JP, Zheng JH. High expression of long non-coding RNA SPRY4-IT1 predicts poor prognosis of clear cell renal cell carcinoma. Int J Clin Exp Pathol. 2014;7(9):5801–5809. | ||
Tong YS, Wang XW, Zhou XL, et al. Identification of the long non-coding RNA POU3F3 in plasma as a novel biomarker for diagnosis of esophageal squamous cell carcinoma. Mol Cancer. 2015;14:3. | ||
Sun M, Liu XH, Lu KH, et al. EZH2-mediated epigenetic suppression of long noncoding RNA SPRY4-IT1 promotes NSCLC cell proliferation and metastasis by affecting the epithelial-mesenchymal transition. Cell Death Dis. 2014;5:e1298. | ||
Xie M, Nie FQ, Sun M, et al. Decreased long noncoding RNA SPRY4-IT1 contributing to gastric cancer cell metastasis partly via affecting epithelial-mesenchymal transition. J Transl Med. 2015;13:250. | ||
Peng W, Wu G, Fan H, Wu J, Feng J. Long noncoding RNA SPRY4-IT1 predicts poor patient prognosis and promotes tumorigenesis in gastric cancer. Tumour Biol. 2015;36(9):6751–6758. | ||
Wang J, Su L, Chen X, et al. MALAT1 promotes cell proliferation in gastric cancer by recruiting SF2/ASF. Biomed Pharmacother. 2014;68(5):557–564. | ||
Zhao Y, Guo Q, Chen J, Hu J, Wang S, Sun Y. Role of long non-coding RNA HULC in cell proliferation, apoptosis and tumor metastasis of gastric cancer: a clinical and in vitro investigation. Oncol Rep. 2014;31(1):358–364. | ||
Sun M, Jin FY, Xia R, et al. Decreased expression of long noncoding RNA GAS5 indicates a poor prognosis and promotes cell proliferation in gastric cancer. BMC Cancer. 2014;14:319. | ||
Liu Y, Zhao J, Zhang W, et al. lncRNA GAS5 enhances G1 cell cycle arrest via binding to YBX1 to regulate p21 expression in stomach cancer. Sci Rep. 2015;5:10159. | ||
Qi P, Xu MD, Shen XH, et al. Reciprocal repression between TUSC7 and miR-23b in gastric cancer. Int J Cancer. 2015;137(6):1269–1278. | ||
Yan J, Guo X, Xia J, et al. MiR-148a regulates MEG3 in gastric cancer by targeting DNA methyltransferase 1. Med Oncol. 2014; 31(3):879. | ||
Sun M, Xia R, Jin F, et al. Downregulated long noncoding RNA MEG3 is associated with poor prognosis and promotes cell proliferation in gastric cancer. Tumour Biol. 2014;35(2):1065–1073. | ||
Peng W, Si S, Zhang Q, et al. Long non-coding RNA MEG3 functions as a competing endogenous RNA to regulate gastric cancer progression. J Exp Clin Cancer Res. 2015;34:79. | ||
Zhou X, Ji G, Ke X, Gu H, Jin W, Zhang G. MiR-141 inhibits gastric cancer proliferation by interacting with long noncoding RNA MEG3 and down-regulating E2F3 expression. Dig Dis Sci. 2015;60(11):3271–3282. | ||
Okugawa Y, Toiyama Y, Hur K, et al. Metastasis-associated long non-coding RNA drives gastric cancer development and promotes peritoneal metastasis. Carcinogenesis. 2014;35(12):2731–2739. | ||
Zhang ZZ, Shen ZY, Shen YY, et al. HOTAIR long noncoding RNA promotes gastric cancer metastasis through suppression of poly r(C)-binding protein (PCBP) 1. Mol cancer Ther. 2015;14(5):1162–1170. | ||
Liu YW, Sun M, Xia R, et al. LincHOTAIR epigenetically silences miR34a by binding to PRC2 to promote the epithelial-to-mesenchymal transition in human gastric cancer. Cell Death Dis. 2015;6:e1802. | ||
Wang Y, Liu X, Zhang H, et al. Hypoxia-inducible lncRNA-AK058003 promotes gastric cancer metastasis by targeting gamma-synuclein. Neoplasia. 2014;16(12):1094–1106. | ||
Cai H, Chen J, He B, Li Q, Li Y, Gao Y. A FOXM1 related long non-coding RNA contributes to gastric cancer cell migration. Mol Cell Biochem. 2015;406(1–2):31–41. | ||
Park SM, Park SJ, Kim HJ, et al. A known expressed sequence tag, BM742401, is a potent lincRNA inhibiting cancer metastasis. Exp Mol Med. 2013;45:e31. | ||
Xu TP, Huang MD, Xia R, et al. Decreased expression of the long non-coding RNA FENDRR is associated with poor prognosis in gastric cancer and FENDRR regulates gastric cancer cell metastasis by affecting fibronectin1 expression. J Hematol Oncol. 2014;7:63. | ||
Hartgrink HH, Jansen EP, van Grieken NC, van de Velde CJ. Gastric cancer. Lancet. 2009;374(9688):477–490. | ||
Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127(12):2893–2917. | ||
Roobol MJ, Schroder FH, van Leenders GL, et al. Performance of prostate cancer antigen 3 (PCA3) and prostate-specific antigen in Prescreened men: reproducibility and detection characteristics for prostate cancer patients with high PCA3 scores (≥100). Eur Urol. 2010;58(6):893–899. | ||
Graham LD, Pedersen SK, Brown GS, et al. Colorectal Neoplasia Differentially Expressed (CRNDE), a novel gene with elevated expression in colorectal adenomas and adenocarcinomas. Genes Cancer. 2011;2(8):829–840. | ||
Arita T, Ichikawa D, Konishi H, et al. Circulating long non-coding RNAs in plasma of patients with gastric cancer. Anticancer Res. 2013;33(8):3185–3193. | ||
Zhou X, Yin C, Dang Y, Ye F, Zhang G. Identification of the long non-coding RNA H19 in plasma as a novel biomarker for diagnosis of gastric cancer. Sci Rep. 2015;5:11516. | ||
Kretz M, Siprashvili Z, Chu C, et al. Control of somatic tissue differentiation by the long non-coding RNA TINCR. Nature. 2013; 493(7431):231–235. | ||
Chen WM, Huang M, Kong R, et al. Antisense long noncoding RNA HIF1A-AS2 is upregulated in gastric cancer and associated with poor prognosis. Dig Dis Sci. 2015;60(6):1655–1662. | ||
Zheng Q, Wu F, Dai WY, et al. Aberrant expression of UCA1 in gastric cancer and its clinical significance. Clin Transl Oncol. 2015;17(8):640–646. | ||
Xu MD, Qi P, Weng WW, et al. Long non-coding RNA LSINCT5 predicts negative prognosis and exhibits oncogenic activity in gastric cancer. Medicine. 2014;93(28):e303. | ||
Ding J, Li D, Gong M, et al. Expression and clinical significance of the long non-coding RNA PVT1 in human gastric cancer. Onco Targets Ther. 2014;7:1625–1630. | ||
Hu Y, Pan J, Wang Y, Li L, Huang Y. Long noncoding RNA linc-UBC1 is negative prognostic factor and exhibits tumor pro-oncogenic activity in gastric cancer. Int J Clin Exp Pathol. 2015;8(1):594–600. | ||
Chen F, Tian Y, Pang EJ, Wang Y, Li L. MALAT2-activated long noncoding RNA indicates a biomarker of poor prognosis in gastric cancer. Cancer Gene Ther. Epub February 27, 2015. | ||
Li L, Zhang L, Zhang Y, Zhou F. Increased expression of LncRNA BANCR is associated with clinical progression and poor prognosis in gastric cancer. Biomed Pharmacother. 2015;72:109–112. | ||
Xu C, Shao Y, Xia T, et al. lncRNA-AC130710 targeting by miR-129–5p is upregulated in gastric cancer and associates with poor prognosis. Tumour Biol. 2014;35(10):9701–9706. | ||
Chen S, Li P, Xiao B, Guo J. Long noncoding RNA HMlincRNA717 and AC130710 have been officially named as gastric cancer associated transcript 2 (GACAT2) and GACAT3, respectively. Tumour Biol. 2014;35(9):8351–8352. | ||
Pang Q, Ge J, Shao Y, et al. Increased expression of long intergenic non-coding RNA LINC00152 in gastric cancer and its clinical significance. Tumour Biol. 2014;35(6):5441–5447. | ||
Mei D, Song H, Wang K, et al. Up-regulation of SUMO1 pseudogene 3 (SUMO1P3) in gastric cancer and its clinical association. Med Oncol. 2013;30(4):709. | ||
Lin X, Yang M, Xia T, Guo J. Increased expression of long noncoding RNA ABHD11-AS1 in gastric cancer and its clinical significance. Med Oncol. 2014;31(7):42. | ||
Liu L, Yan B, Yang Z, Zhang X, Gu Q, Yue X. ncRuPAR inhibits gastric cancer progression by down-regulating protease-activated receptor-1. Tumour Biol. 2014;35(8):7821–7829. | ||
Liu Z, Shao Y, Tan L, Shi H, Chen S, Guo J. Clinical significance of the low expression of FER1L4 in gastric cancer patients. Tumour Biol. 2014;35(10):9613–9617. | ||
Chen X, Sun J, Song Y, et al. The novel long noncoding RNA AC138128.1 may be a predictive biomarker in gastric cancer. Med Oncol. 2014;31(11):262. | ||
Shao Y, Ye M, Jiang X, et al. Gastric juice long noncoding RNA used as a tumor marker for screening gastric cancer. Cancer. 2014; 120(21):3320–3328. | ||
Xiao B, Guo J. Long noncoding RNA AC096655.1–002 has been officially named as gastric cancer-associated transcript 1, GACAT1. Tumour Biol. 2013;34(5):3271. | ||
Sun W, Wu Y, Yu X, et al. Decreased expression of long noncoding RNA AC096655.1–002 in gastric cancer and its clinical significance. Tumour Biol. 2013;34(5):2697–2701. | ||
Shao Y, Chen H, Jiang X, et al. Low expression of lncRNA- HMlincRNA717 in human gastric cancer and its clinical significances. Tumour Biol. 2014;35(10):9591–9595. | ||
Zhou B, Jing XY, Wu JQ, Xi HF, Lu GJ. Down-regulation of long non-coding RNA LET is associated with poor prognosis in gastric cancer. Int J Clin Exp Pathol. 2014;7(12):8893–8898. | ||
Zhu S, Mao J, Shao Y, et al. Reduced expression of the long non-coding RNA AI364715 in gastric cancer and its clinical significance. Tumour Biol. 2015;36(10):8041–8045. | ||
Lai Y, Xu P, Li Q, et al. Downregulation of long noncoding RNA ZMAT1 transcript variant 2 predicts a poor prognosis in patients with gastric cancer. Int J Clin Exp Pathol. 2015;8(5):5556–5562. | ||
Sun J, Song Y, Chen X, et al. Novel long non-coding RNA RP11–119F7.4 as a potential biomarker for the development and progression of gastric cancer. Oncol Lett. 2015;10(1):115–120. | ||
Liu G, Neumeister M, Reichensperger J, Yang RD. Therapeutic potential of human adipose stem cells in a cancer stem cell-like gastric cancer cell model. Int J Oncol. 2013;43(4):1301–1309. | ||
Salama NR, Hartung ML, Muller A. Life in the human stomach: persistence strategies of the bacterial pathogen Helicobacter pylori. Nat Rev Microbiol. 2013;11(6):385–399. | ||
Yang L, Long Y, Li C, et al. Genome-wide analysis of long noncoding RNA profile in human gastric epithelial cell response to Helicobacter pylori. Jpn J Infect Dis. 2015;68(1):63–66. | ||
Derrien T, Johnson R, Bussotti G, et al. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Res. 2012;22(9):1775–1789. | ||
Fox H, Kondi A, Linsell CA, Parker AM, Sizaret P. The alpha 1-fetoprotein test in hepatocellular carcinoma. Lancet. 1970; 1(7643):411. | ||
Kouvalainen K, Linder E, Nanto V. Significance of alpha 1-fetoprotein in primary hepatomas. Duodecim. 1970;86(20):1171–1173. | ||
Yang YR, Zang SZ, Zhong CL, Li YX, Zhao SS, Feng XJ. Increased expression of the lncRNA PVT1 promotes tumorigenesis in non-small cell lung cancer. Int J Clin Exp Pathol. 2014;7(10):6929–6935. | ||
Huang X, Yuan T, Tschannen M, et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genomics. 2013;14:319. | ||
Lee GL, Dobi A, Srivastava S. Prostate cancer: diagnostic performance of the PCA3 urine test. Nat Rev Urol. 2011;8(3):123–124. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.