Back to Journals » Cancer Management and Research » Volume 12
Long Noncoding RNA ZFPM2-AS1 Enhances the Malignancy of Cervical Cancer by Functioning as a Molecular Sponge of microRNA-511-3p and Consequently Increasing FGFR2 Expression
Authors Dai J, Wei R, Zhang P, Liu P
Received 13 November 2019
Accepted for publication 18 December 2019
Published 23 January 2020 Volume 2020:12 Pages 567—580
DOI https://doi.org/10.2147/CMAR.S238373
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Eileen O'Reilly
This paper has been retracted.
Jun Dai, 1 Rujia Wei, 2 Peihai Zhang, 3 Peishu Liu 1
1Department of Gynaecology and Obstetrics, Qilu Hospital of Shandong University, Jinan, Shandong 250012, People’s Republic of China; 2School of Life Sciences, Liaocheng University, Liaocheng, Shandong 252004, People’s Republic of China; 3Department of Gynaecology and Obstetrics, Qilu Hospital of Shandong University (Qingdao), Qingdao, Shandong 266035, People’s Republic of China
Correspondence: Jun Dai
Department of Gynaecology and Obstetrics, Qilu Hospital of Shandong University, 107 Wenhua West Road, Jinan, Shandong 250012, People’s Republic of China
Email [email protected]
Peihai Zhang
Department of Gynaecology and Obstetrics, Qilu Hospital of Shandong University (Qingdao), 758 Hefei Road, Qingdao, Shandong 266035, People’s Republic of China
Email [email protected]
Purpose: A long noncoding RNA called ZFPM2 antisense RNA 1 (ZFPM2-AS1) has been verified as a key modulator in multiple human cancer types. Nonetheless, the expression and functions of ZFPM2-AS1 in cervical cancer remain poorly understood. Therefore, our purpose was to characterize the expression pattern, clinical value, and detailed roles of ZFPM2-AS1 in cervical cancer.
Methods: Reverse-transcription quantitative PCR was carried out to measure ZFPM2-AS1 expression in cervical cancer. A Cell Counting Kit-8 assay, flow cytometry, Transwell migration and invasion assays, and a tumor xenograft experiment were conducted to determine the influence of ZFPM2-AS1 on cervical cancer cell proliferation, apoptosis, migration, and invasion in vitro and on tumor growth in vivo, respectively.
Results: ZFPM2-AS1 was found to be aberrantly upregulated in cervical cancer, and its upregulation was associated with unfavorable values of clinical parameters. A ZFPM2-AS1 knockdown significantly reduced cervical cancer cell proliferation, migration, and invasion and increased apoptosis in vitro. The ZFPM2-AS1 knockdown decelerated tumor growth of cervical cancer cells in vivo. Molecular investigation indicated that ZFPM2-AS1 acts as a molecular sponge of microRNA-511-3p (miR-511-3p) in cervical cancer cells. Fibroblast growth factor receptor 2 (FGFR2) mRNA was validated as a direct target of miR-511-3p in cervical cancer, and its expression was positively modulated by ZFPM2-AS1. The effects of the ZFPM2-AS1 knockdown on malignant characteristics of cervical cancer cells were greatly attenuated by miR-511-3p inhibition.
Conclusion: ZFPM2-AS1 promotes cervical cancer progression through upregulation of miR-511-3p–FGFR2 axis output, thereby pointing to possible diagnostics and therapeutics based on the ZFPM2-AS1–miR-511-3p–FGFR2 pathway.
Keywords: ZFPM2 antisense RNA 1, cervical cancer therapy, fibroblast growth factor receptor 2, microRNA-511-3p
Introduction
Cervical cancer ranks the second most frequent cancer among women and the fourth leading cause of gynecological-cancer–related deaths worldwide.1 A total of 569,847 new cervical cancer cases were diagnosed and 311,365 patients died of cervical cancer per year, as estimated by Global Cancer Statistics 2018.2 At present, the major therapeutic approaches to cervical cancer include surgical treatments, radiotherapy, cytotoxic chemotherapy, and adjuvant therapy.3 In addition, natural anti-cancerous drug is also widely used in therapeutic strategies against cervical cancer.4 In spite of tremendous developments in the diagnostic techniques and therapeutic approaches to cervical cancer, long-term prognosis of the patients, especially cases diagnosed at an advanced stage, remains unsatisfactory.5 It is generally believed that human papillomavirus infection is a major but not the only factor causing cervical cancer initiation and progression.6,7 Accordingly, full characterization of the molecular events of cervical cancer, especially of the pathogenesis, is important for the identification of novel and promising therapeutic techniques.
Long noncoding RNAs (lncRNAs) are transcripts with length of over 200 nucleotides.8 LncRNAs have no protein-coding capacity but can regulate gene expression at transcriptional, post-transcriptional, and chromosomal levels.9 Recently, lncRNAs received much attention because of their crucial regulatory functions in various biological processes.10,11 Moreover, a growing body of evidence suggests that lncRNAs may play complicated and crucial roles during tumorigenesis and tumor progression.12–14 Many lncRNAs have turned out to be abnormally expressed in cervical cancer. For instance, TUG1,15 PVT1,16 and CRNDE17 are highly expressed in cervical cancer; on the contrary, HAND2-AS1,18 PTCSC3,19 and ZNF667-AS120 are underexpressed in this cancer. Functionally, the aberrantly expressed lncRNAs exert cancer-inhibiting or cancer-promoting actions and thereby serve as critical modulators of cervical carcinogenesis by affecting a number of malignant characteristics.21–23 Therefore, an in-depth understanding of the functions of lncRNAs in cervical cancer is crucial for improving the disease diagnosis, prognosis, and treatments.
An lncRNA called ZFPM2-AS1 has been verified as a key modulator in gastric cancer,24 lung adenocarcinoma,25 and renal cell cancer.26 Nevertheless, the expression and functions of ZFPM2-AS1 in cervical cancer remain poorly understood. Therefore, our purpose was to characterize the expression pattern, clinical value, and detailed roles of ZFPM2-AS1 in cervical cancer. Moreover, the molecular mechanisms behind ZFPM2-AS1–mediated promotion of the aggressive phenotype of cervical cancer cells were explored in detail in vitro and in vivo.
Materials and Methods
Collection of Clinical Samples
This study was carried out with the approval of Ethics Committees of Qilu Hospital of Shandong University and in accordance with the Declaration of Helsinki. All participating patients provided written informed consent. Cervical cancer tissue samples and tumor adjacent tissue samples were collected from 47 patients in Qilu Hospital of Shandong University (Shandong, China). None of the patients had received chemotherapy, radiotherapy, or other antitumor therapies before the surgical operation. The obtained tissue specimens were quickly frozen in liquid nitrogen and stored in liquid nitrogen until analysis.
Cell Lines
Four cervical cancer cell lines, HeLa, SiHa, C-33A, and CaSki, were purchased from the Institute of Biochemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China), and a normal human cervix epithelial cell line (Ect1/E6E7) from the American Type Culture Collection (Manassas, VA, USA). All the aforementioned cell lines were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing 10% of fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco; Thermo Fisher Scientific, Inc.) at 37 °C in a humidified atmosphere supplied with 5% of CO2.
Transient Transfection
The small interfering RNA (siRNA) specific to ZFPM2-AS1 (si-ZFPM2-AS1) and negative control siRNA (si-NC) were synthesized by RiboBio (Guangzhou, China). An miR-511-3p mimic, microRNA (miRNA) mimic negative control (miR-NC), an miR-511-3p inhibitor, and its NC were purchased from GeneCopoeia (Guangzhou, China). A plasmid encoding FGFR2 (called pcDNA3.1-FGFR2) and the empty pcDNA3.1 vector were designed and constructed by GenePharma Technology (Shanghai, China). Cells were seeded in 24-well plates and incubated at 37 °C and 5% CO2 for 24 h. The cells were transfected with the above siRNA, miRNA mimic, miRNA inhibitor, or plasmid by means of Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA).
Isolation of Cytoplasmic and Nuclear RNA
As described previously,27 the isolation of the cytoplasmic and nuclear fractions of cervical cancer cells was performed with the PARIS Kit (Invitrogen; Thermo Fisher Scientific, Inc.).
Reverse-Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
RT-qPCR was performed as described previously.28 TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) was employed for total-RNA extraction. The concentration and purity of total RNA were evaluated on a NanoDrop 2000 spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific, Inc.). For the quantification of miR-511-3p expression, complementary DNA (cDNA) was synthesized using the miScript Reverse Transcription Kit (Qiagen GmbH, Hilden, Germany). The qPCR was then conducted with the miScript SYBR Green PCR Kit (Qiagen GmbH, Hilden, Germany). For the analysis of ZFPM2-AS1 and FGFR2 mRNA expression, total RNA was reversely transcribed into cDNA by means of the PrimeScript RT-Reagent Kit (Takara Bio, Kusatsu, Japan). The cDNA was then subjected to PCR amplification with the SYBR Premix Ex Taq™ Kit (Takara Bio, Kusatsu, Japan). U6 small nuclear RNA served as the internal control for miR-511-3p, whereas GAPDH for other RNAs. Relative gene expression was analyzed with the comparative quantification cycle (2−ΔΔCq) method.
Cell Counting Kit-8 (CCK-8) Assay
CCK-8 assay was applied to determine cellular proliferative ability as described previously.29 At 24 h post-transfection, preparation of cell suspension was performed, and cell concentration was adjusted to 2 × 103 cells/mL. In total, 100 μL of the cell suspension was inoculated into wells of 96-well plates. To test cellular proliferation, 10 μL of the CCK-8 reagent (Dojindo Molecular Technologies, Inc.) was added into each well, after which the plates were incubated at 37 °C and 5% CO2 for another 2 h. The absorbance at 450 nm wavelength was measured on a microplate reader (Bio-Rad Laboratories, Benicia, CA, USA). The CCK-8 assay was carried out at 0, 24, 48, and 72 h after cell seeding.
Flow-Cytometric Analysis of Apoptosis
The apoptosis of transfected cells was evaluated by menas of flow-cytometric analysis.30 After cultivation for 48 h, transfected cells were harvested using trypsin without EDTA and rinsed with precooled phosphate-buffered saline, followed by quantification of apoptotic cells using the Annexin V–Fluorescein Isothiocyanate (FITC) Apoptosis Detection Kit (BioLegend, San Diego, CA, USA). Namely, the transfected cells were resuspended in 100 μL of Annexin-V-binding buffer prior to double staining with 5 µL of Annexin V–FITC and 5 µL of the propidium iodide solution. After 15 min incubation at room temperature in darkness, a flow cytometer (FACScan; BD Biosciences, Franklin Lakes, NJ, USA) was utilized to quantify the apoptotic cells.
Transwell Migration and Invasion Assays
The migratory capacity was assessed in 24-well Transwell® chambers (pore size: 8 µm; BD Biosciences, San Jose, CA, USA) as described by previous studies.31,32 A total of 5 × 104 transfected cells were resuspended in 100 μL of FBS-free DMEM and were seeded in the upper compartments. The complete medium (containing 20% of FBS) was added into the basolateral chambers. After 24 h incubation, nonmigratory cells (those remaining on the upper side of the membranes) were gently wiped off with a cotton-tipped swab, while the migratory cells were fixed in a methanol solution and stained with 0.1% crystal violet. The counting of migratory cells was conducted under an inverted optical microscope (x200 magnification; Olympus Corporation) in five randomly selected fields of view for each chamber. Similar experimental steps were performed for testing the invasive ability, except that the chambers were precoated with Matrigel (BD Biosciences).
Tumor Xenograft Model
As described previously,33 tumor xenograft model was utilized to test the influence of ZFPM2-AS1 knockdown on tumor growth in vivo. The plasmids encoding a short hairpin RNA (shRNA) specifically targeting ZFPM2-AS1 (pLKO.1- sh-ZFPM2-AS1) or negative control shRNA (pLKO.1-sh-NC) were designed and manufactured by GenePharma Technology. HeLa cells were transfected with a lentivirus carrying either pLKO.1-sh-ZFPM2-AS1 or pLKO.1-sh-NC and were selected with 2 μg/mL puromycin, resulting in a stable ZFPM2-AS1 knockdown cell line.
Female BALB/c nude mice (4–6 weeks of age) were acquired from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai Laboratory Animal Center of Chinese Academy of Sciences, Shanghai, China). HeLa cells stably transfected with either sh-ZFPM2-AS1 or sh-NC were inoculated subcutaneously into a flank of the mice. The size of subcutaneous tumors was determined every 4 days by measurement of their length and width. The volume of tumors was calculated via the following formula: Volume (mm3) = width2 (mm2) × length (mm)/2. On Day 28 post-injection, all the mice were euthanized with the method of cervical dislocation. The tumor xenografts were resected for weighing and further experiments. The animal experiments were approved by the Institutional Experimental Animal Review Board of Qilu Hospital of Shandong University, and performed in compliance with the Animal Protection Law of the People’s Republic of China-2009 for experimental animals.
RNA Immunoprecipitation (RIP) Assay
The binding between ZFPM2-AS1 and miR-511-3p in cervical cancer cells was examined by the RIP assay based on the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore, Bedford, MA, USA), as described by previous study.34 Cells were collected and probed in RIPA cell lysis buffer. Following 20 min incubation on ice, each cell lysate was incubated overnight at 4 °C with magnetic beads conjugated with an anti-Argonaute 2 (AGO2) antibody or IgG control (Millipore, Bedford, MA, USA). The beads were treated with 150 μL proteinase K for digestion of the protein. Finally, RT-qPCR was performed to analyze the purified RNA.
Bioinformatic Prediction
The miRNA(s) that may interact with ZFPM2-AS1 was predicted in starBase 3.0 software (http://starbase.sysu.edu.cn/).35 Two online miRNA target prediction databases, miRDB36 (http://mirdb.org/) and TargetScan37 (http://www.targetscan.org/), were employed for miR-511-3p target prediction.
Luciferase Reporter Assay
Luciferase reporter assay was conducted to assess the binding between ZFPM2-AS1 and miR-511-3p in cervical cancer cells.38 FGFR2 3′ untranslated region (UTR) fragments containing either the predicted wild-type (WT) miR-511-3p–binding site or a mutant (MUT) binding site were amplified by GenePharma Technology. The amplified fragments were cloned into the pmirGLO luciferase reporter vector (Promega, Madison, WI, USA) to respectively create luciferase reporter plasmids WT-FGFR2 and MUT-FGFR2. For testing the interaction between miR-511-3p and ZFPM2-AS1, the luciferase reporter plasmids WT-ZFPM2-AS1 and MUT-ZFPM2-AS1 were constructed in a similar way. Cotransfection of cervical cancer cells was performed with the combination of either a WT or MUT reporter plasmid and either the miR-511-3p mimic or miR-NC. After 48 h, we harvested the transfected cells and detected the luciferase activities via a Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA). Renilla luciferase activity was utilized as an internal control for normalization.
Western Blotting
Western blotting was conducted to detect the protein expression.39 Cultured cells were washed with phosphate-buffered saline and lysed with radioimmunoprecipitation assay lysis buffer (Beyotime, Shanghai, China) for total-protein isolation. Equal amounts of protein were separated by sodium dodecyl sulfate 10% polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes, followed by blocking at room temperature for 2 h with 5% nonfat milk diluted in Tris-buffered saline (TBS) supplemented with 0.1% of Tween 20. After overnight incubation at 4 °C with primary antibodies, the membranes were probed with a horseradish peroxidase–conjugated goat anti-mouse IgG antibody (1:5000 dilution; cat. No. ab205719; Abcam) (secondary antibody). The ECL™ Western Blotting Detection Reagent (GE Healthcare) was employed for immunoblot visualization. The following primary antibodies were used in this study: anti-FGFR2 (1:1000 dilution; cat. No. ab58201; Abcam) and anti-GAPDH (1:1000 dilution; cat. No. ab9482; Abcam). GAPDH served as an endogenous control for the quantification of FGFR2 protein expression.
Statistical Analysis
All results are expressed as the mean ± standard deviation from three independent experiments. The relation between ZFPM2-AS1 and clinical parameters of the patients with cervical cancer was tested via the chi-squared test. The Kaplan–Meier method was utilized for survival analysis, and differences between survival curves were assessed by the logrank test. Student’s t test was conducted to evaluate differences between two groups. The comparison among multiple groups was made by one-way analysis of variance (ANOVA) plus the Bonferroni–Dunn test. All statistical analyses were conducted in the SPSS software, version 21.0 (Chicago, IL, USA), and P < 0.05 was assumed to indicate a statistically significant difference.
Results
ZFPM2-AS1 Is Upregulated in Cervical Cancer and Is Related to Shorter Survival of Patients with Cervical Cancer
Cervical cancer tissue samples and tumor adjacent tissue samples were collected from 45 patients and used for the determination of ZFPM2-AS1 expression by RT-qPCR. The results indicated that ZFPM2-AS1 expression was higher in the cervical cancer tissue samples than in tumor adjacent tissues (Figure 1A, P < 0.05). The expression status of ZFPM2-AS1 in the normal human cervix epithelial cell line (Ect1/E6E7) and four cervical cancer cell lines (HeLa, SiHa, C-33A, and CaSki) was analyzed via RT-qPCR. ZFPM2-AS1 turned out to be upregulated in all four tested cervical cancer cell lines relative to Ect1/E6E7 cells (Figure 1B, P < 0.05).
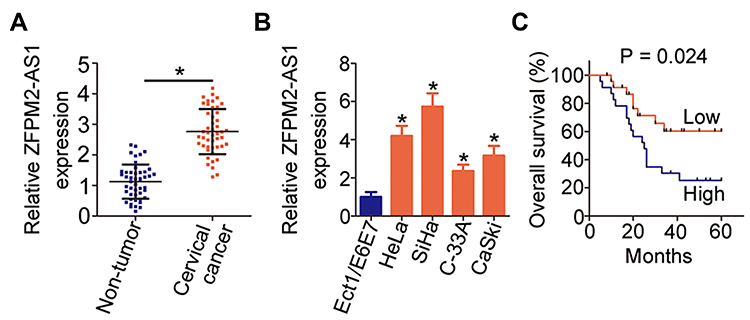 |
Figure 1 ZFPM2-AS1 is highly expressed in cervical cancer, and this overexpression correlates with a poor prognosis of patients with cervical cancer. (A) RT-qPCR analysis was carried out to measure ZFPM2-AS1 expression in 45 pairs of cervical cancer tissue samples and tumor adjacent tissues. *P < 0.05 vs tumor adjacent tissue samples. (B) The expression of ZFPM2-AS1 in the four cervical cancer cell lines (HeLa, SiHa, C-33A, and CaSki) was determined in an RT-qPCR assay. A normal human cervix epithelial cell line (Ect1/E6E7) served as the control. *P < 0.05 vs Ect1/E6E7 cells. (C) All 45 participating patients were subdivided into two groups: the ZFPM2-AS1 low-expression group and ZFPM2-AS1 high-expression group. The overall survival of the two groups was analyzed with the Kaplan–Meier method and compared by the logrank test. P = 0.024. |
We subsequently subdivided all the patients with cervical cancer into either ZFPM2-AS1 low-expression group or ZFPM2-AS1 high-expression group, according to the median value of ZFPM2-AS1 among the cervical cancer tissue samples. Analysis the correlation between ZFPM2-AS1 expression and clinical parameters revealed that increased ZFPM2-AS1 expression obviously correlated with tumor size (P = 0.041), FIGO stage (P = 0.020), and lymph node metastasis (P = 0.017) among these 45 patients with cervical cancer (Table 1). Furthermore, patients with cervical cancer in the ZFPM2-AS1 high-expression group showed shorter overall survival than did the patients in the ZFPM2-AS1 low-expression group (Figure 1C, P = 0.024). Taken together, these results implied that ZFPM2-AS1 is overexpressed in cervical cancer and is closely associated with poor clinical outcomes of patients with cervical cancer, suggesting that ZFPM2-AS1 might be implicated in the malignancy of this disease.
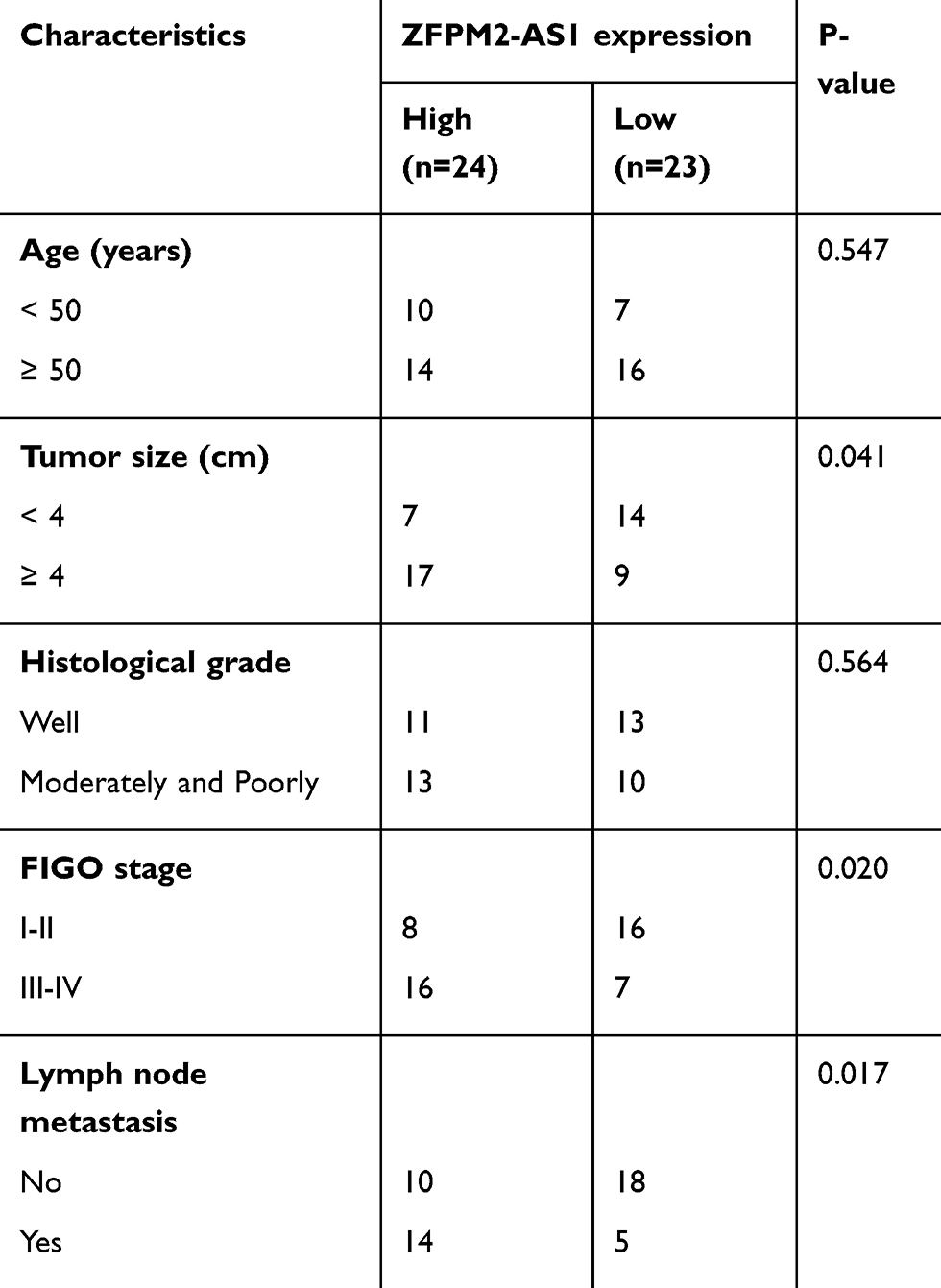 |
Table 1 The relation between ZFPM2-AS1 expression and clinicopathological characteristics in patients with cervical cancer. |
Knockdown of ZFPM2-AS1 Restricts Cervical Cancer Cell Proliferation, Migration, and Invasion and Induces Apoptosis
Given that ZFPM2-AS1 was more highly expressed in HeLa and SiHa cell lines among the four tested cervical cancer cell lines, the two cell lines were chosen as cell models for further experiments. To evaluate the significance of ZFPM2-AS1 in cervical cancer, its expression was silenced in HeLa and SiHa cells using the siRNA specific to ZFPM2-AS1 (si-ZFPM2-AS1). Obvious downregulation of ZFPM2-AS1 in HeLa and SiHa cells was verified via RT-qPCR analysis (Figure 2A, P < 0.05). The CCK-8 assay showed that the knockdown of ZFPM2-AS1 remarkably suppressed the proliferative ability of HeLa and SiHa cells (Figure 2B, P < 0.05). In addition, the knockdown of ZFPM2-AS1 strongly promoted the apoptosis of HeLa and SiHa cells, as evidenced by flow-cytometric analysis (Figure 2C and D, P < 0.05). We next conducted Transwell migration and invasion assays to test whether ZFPM2-AS1 is involved in the mobility of cervical cancer cells. According to the results, the knockdown of ZFPM2-AS1 significantly reduced the migratory (Figure 2E, P < 0.05) and invasive abilities (Figure 2F, P < 0.05) of HeLa and SiHa cells. Therefore, ZFPM2-AS1 may play a cancer-promoting part in the malignant phenotype of cervical cancer cells.
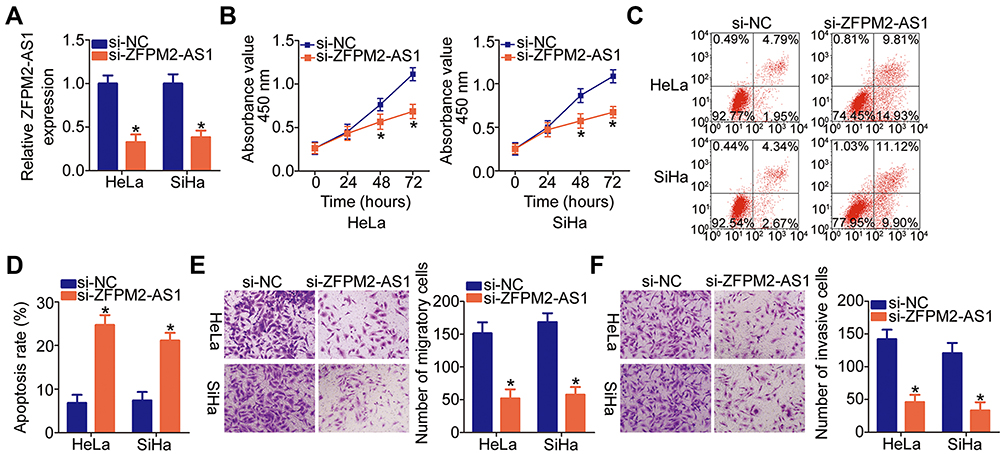 |
Figure 2 The ZFPM2-AS1 knockdown inhibits HeLa and SiHa cell proliferation, migration, and invasion but promotes apoptosis. (A) HeLa and SiHa cells were transfected with either si-ZFPM2-AS1 or si-NC. The efficiency of si-ZFPM2-AS1 transfection was evaluated by RT-qPCR. *P < 0.05 vs the si-NC group. (B) The CCK-8 assay was applied to determine the proliferative ability of HeLa and SiHa cells after either si-ZFPM2-AS1 or si-NC transfection. *P < 0.05 vs the si-NC group. (C, D) The apoptosis of ZFPM2-AS1–deficient HeLa and SiHa cells was determined by flow cytometry. *P < 0.05 vs group si-NC. (E, F) The influence of the ZFPM2-AS1 knockdown on the migratory and invasive abilities of HeLa and SiHa cells was assessed by Transwell migration and invasion assays (x200 magnification). *P < 0.05 vs the si-NC group. |
ZFPM2-AS1 Directly Targets miR-511-3p in Cervical Cancer Cells
LncRNA can directly interact with miRNAs to modulate the expression of miRNAs’ targets during carcinogenesis and cancer progression.40 We first evaluated the expression distribution of ZFPM2-AS1 in HeLa and SiHa cells. As indicated in Figure 3A, ZFPM2-AS1 was observed both in the cell nucleus and cytoplasm but was mostly located in the cytoplasm of HeLa and SiHa cells, suggesting that activities of specific miRNAs may be regulated by ZFPM2-AS1. Some studies have revealed that the roles of ZFPM2-AS1 are dependent on miR-137 in renal cell cancer26 and on miR-18b-5p in lung adenocarcinoma;25 accordingly, we next determined whether ZFPM2-AS1 may directly target specific miRNA in cervical cancer. First, bioinformatic prediction was performed to search for the potential miRNA-binding sites in ZFPM2-AS1. The analysis suggested that ZFPM2-AS1 harbors a potential binding site for miR-511-3p (Figure 3B). Next, after confirming the efficiency of miR-511-3p mimic transfection (Figure 3C, P < 0.05), we conducted the luciferase reporter assay in HeLa and SiHa cells that were cotransfected with either plasmid WT-ZFPM2-AS1 or MUT-ZFPM2-AS1 and either the miR-511-3p mimic or miR-NC. Transfection with the miR-511-3p mimic notably reduced the luciferase activity of the plasmid containing the WT miR-511-3p–binding site in HeLa and SiHa cells (P < 0.05), whereas there was no significant alteration in the MUT-ZFPM2-AS1–transfected cell group (Figure 3D). After that, the RIP assay revealed that ZFPM2-AS1 and miR-511-3p were significantly enriched in the AGO2 immunoprecipitation complex (Figure 3E, P < 0.05), meaning that ZFPM2-AS1 and miR-511-3p can associate with AGO2 and that miR-511-3p can directly bind to ZFPM2-AS1 in cervical cancer cells.
 |
Figure 3 ZFPM2-AS1 directly targets miR-511-3p in cervical cancer cells. (A) The expression distribution of ZFPM2-AS1 in the HeLa and SiHa cells was analyzed via isolation of cytoplasmic and nuclear RNA followed by RT-qPCR. (B) Bioinformatics analysis uncovered an miR-511-3p–binding site within ZFPM2-AS1. The mutant binding site is shown too. (C) The expression of miR-511-3p in HeLa and SiHa cells was quantified after either miR-511-3p mimic or miR-NC introduction. *P < 0.05 vs the miR-NC group. (D) Either luciferase reporter plasmid WT-ZFPM2-AS1 or MUT-ZFPM2-AS1 as well as either the miR-511-3p mimic or miR-NC were cotransfected into HeLa and SiHa cells. The luciferase reporter assay was conducted at 48 h post-transfection. *P < 0.05 vs group miR-NC. (E) The RIP assay was conducted to evaluate the enrichment of ZFPM2-AS1 and miR-511-3p in AGO2 immunoprecipitates from HeLa and SiHa cell lysates. *P < 0.05 as compared with the IgG group. (F) Assessment of miR-511-3p expression in 45 pairs of cervical cancer tissue samples and tumor adjacent tissues was conducted by RT-qPCR. *P < 0.05 vs nontumor tissue samples. (G) Spearman correlation analysis was performed to test the expression correlation between ZFPM2-AS1 and miR-511-3p in the cervical cancer tissue samples (n = 45). r = −0.6011, P < 0.0001. (H) The expression of miR-511-3p was measured in HeLa and SiHa cells after the transfection with either si-ZFPM2-AS1 or si-NC. *P < 0.05 as compared to the si-NC group. |
Furthermore, the expression of miR-511-3p was measured in the 45 pairs of cervical cancer tissue samples and tumor adjacent tissue samples. The RT-qPCR data showed that miR-511-3p was weakly expressed in the cervical cancer tissue samples compared with the tumor adjacent tissues (Figure 3F, P < 0.05). Moreover, the expression of miR-511-3p inversely correlated with ZFPM2-AS1 expression in these 45 cervical cancer tissue samples (Figure 3G; r = –0.6011, P < 0.0001), as evidenced by Spearman correlation analysis. Finally, the miR-511-3p amount in ZFPM2-AS1–deficient HeLa and SiHa cells was determined, and we investigated whether the expression of miR-511-3p can be sponged by ZFPM2-AS1. Notably, miR-511-3p expression was increased by the transfection of si-ZFPM2-AS1 in HeLa and SiHa cells (Figure 3H, P < 0.05). These results collectively indicated that ZFPM2-AS1 may directly interact with miR-511-3p and can act as a molecular sponge of miR-511-3p in cervical cancer cells.
miR-511-3p Suppresses the Malignant Phenotype of Cervical Cancer Cells
The miR-511-3p mimic or miR-NC was introduced into HeLa and SiHa cells, and functional experiments were conducted on the transfected cells to explore the functions of miR-511-3p in cervical cancer progression. The CCK-8 assay and flow cytometry suggested that the ectopic miR-511-3p expression significantly decreased the proliferation (Figure 4A, P < 0.05) and increased apoptosis (Figure 4B, P < 0.05) of HeLa and SiHa cells. Additionally, we found that the migration (Figure 4C, P < 0.05) and invasiveness (Figure 4D, P < 0.05) of HeLa and SiHa cells were obviously suppressed by miR-511-3p overexpression, as determined by Transwell migration and invasion assays. In summary, these results indicated that miR-511-3p plays a tumor-suppressive role in cervical cancer cells.
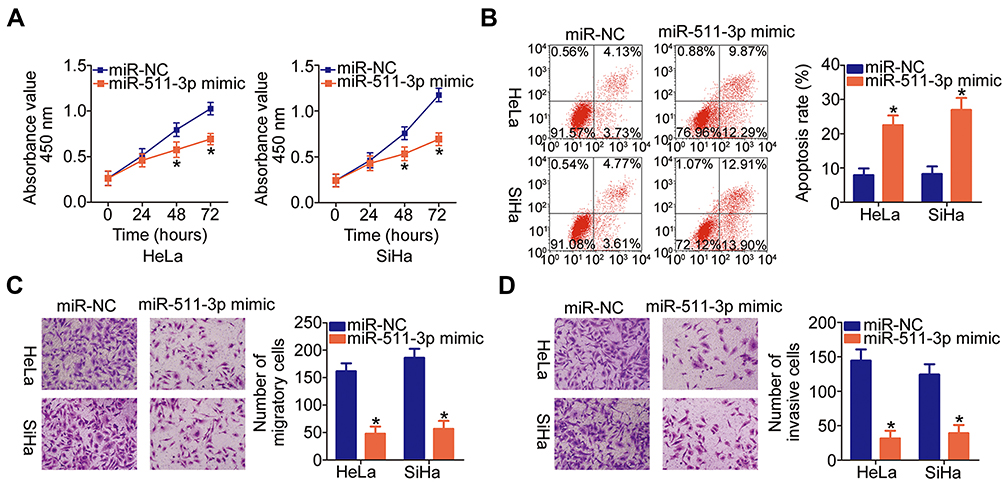 |
Figure 4 MiR-511-3p overexpression has an inhibitory influence on the malignant phenotype of HeLa and SiHa cells. (A, B) HeLa and SiHa cells were transfected with either the miR-511-3p mimic or miR-NC. The proliferation and apoptosis of miR-511-3p–overexpressing HeLa and SiHa cells were respectively examined by the CCK-8 assay and flow-cytometric analysis. *P < 0.05 vs the miR-NC group. (C, D) The migration and invasiveness of the aforementioned cells were assessed in Transwell migration and invasion assays (x200 magnification). *P < 0.05 vs the miR-NC group. |
FGFR2 Is a Direct Target Gene of miR-511-3p in Cervical Cancer Cells
To decipher the mechanism underlying the involvement of miR-511-3p in cervical cancer progression, we carried out bioinformatics analysis to find a downstream target of miR-511-3p. The 3′-UTR of FGFR2 mRNA was found to contain complementary binding sequences for the seed region of miR-511-3p (Figure 5A), and FGFR2 was selected for further experimental verification because this gene is involved in the cervical carcinogenesis and cancer progression41–43 in addition to being regulated by multiple miRNAs.44–46 The luciferase reporter assay was performed to verify the above prediction. The luciferase activity of HeLa and SiHa cells transfected with WT-FGFR2 was substantially impaired by the miR-511-3p mimic (P < 0.05); however, when the binding sequences of miR-511-3p were mutated in the luciferase reporter plasmid (resulting in plasmid MUT-FGFR2), upregulation of miR-511-3p failed to inhibit the luciferase activity in the transfected HeLa and SiHa cells (Figure 5B). In addition, transfection with the miR-511-3p mimic significantly diminished FGFR2 expression in HeLa and SiHa cells at both mRNA (Figure 5C, P < 0.05) and protein levels (Figure 5D, P < 0.05). Next, using RT-qPCR, we revealed that the expression of FGFR2 mRNA was much higher in cervical cancer tissue samples (Figure 5E, P < 0.05), manifesting an inverse correlation with miR-511-3p expression (Figure 5F; r = –0.5810, P < 0.0001). In a word, FGFR2 mRNA is a direct target of miR-511-3p in cervical cancer cells.
 |
Figure 5 FGFR2 mRNA is a direct target of miR-511-3p in cervical cancer cells. (A) Schematic representation of the wild-type and mutant binding sites for miR-511-3p in the 3′-UTR of FGFR2 mRNA. (B) The luciferase reporter assay was carried out in HeLa and SiHa cell lysates after cotransfection with either reporter plasmid WT-FGFR2 or MUT-FGFR2 and the miR-511-3p mimic or miR-NC. *P < 0.05 vs the miR-NC group. (C, D) The impact of transfection with the miR-511-3p mimic on FGFR2 expression was examined by RT-qPCR and Western blotting in HeLa and SiHa cells. MiR-NC served as the control. *P < 0.05 vs group miR-NC. (E) RT-qPCR analysis was performed to evaluate FGFR2 mRNA expression in the 45 pairs of cervical cancer tissue samples and tumor adjacent tissues. *P < 0.05 vs tumor adjacent tissue samples. (F) The correlation between FGFR2 mRNA and miR-511-3p expression levels in the 45 cervical cancer tissue samples was determined by Spearman correlation analysis. r = −0.5810, P < 0.0001. |
The Oncogenic Actions of ZFPM2-AS1 in Cervical Cancer are Dependent on the Enhancement of the miR-511-3p–FGFR2 Axis Output
To verify the relation between ZFPM2-AS1 and FGFR2 in cervical cancer, the expression of FGFR2 was quantitated in ZFPM2-AS1–deficient HeLa and SiHa cells. Expression levels of the FGFR2 mRNA (Figure 6A, P < 0.05) and protein (Figure 6B, P < 0.05) were lower in the ZFPM2-AS1–deficient HeLa and SiHa cells. Moreover, ZFPM2-AS1 expression was found to positively correlate with FGFR2 mRNA expression in cervical cancer tissue samples judging by Spearman correlation analysis results (Figure 6C; r = 0.5243, P = 0.0002). We subsequently investigated whether ZFPM2-AS1 controls FGFR2 expression through miR-511-3p sponging. To this end, either the miR-511-3p inhibitor or NC inhibitor as well as si-ZFPM2-AS1 were cotransfected into HeLa and SiHa cells, and the expression of FGFR2 was then determined. MiR-511-3p inhibitor introduction obviously decreased the expression of miR-511-3p in HeLa and SiHa cells (Figure 6D, P < 0.05). The downregulation of FGFR2 mRNA (Figure 6E, P < 0.05) and protein (Figure 6F, P < 0.05) mediated by the ZFPM2-AS1 knockdown was found to be greatly recovered by miR-511-3p inhibition in HeLa and SiHa cells. Therefore, these results proved that ZFPM2-AS1 acts as a competing endogenous RNA (ceRNA) on miR-511-3p and thereby positively regulates FGFR2 expression in cervical cancer.
 |
Figure 6 The miR-511-3p–FGFR2 axis is responsible for the cancer-promoting activities of ZFPM2-AS1 in cervical cancer cells. (A, B) The mRNA and protein levels of FGFR2 were determined in HeLa and SiHa cells treated with either si-ZFPM2-AS1 or si-NC using RT-qPCR and Western blotting, respectively. *P < 0.05 vs the si-NC group. (C) Spearman correlation analysis was applied to demonstrate the positive correlation between FGFR2 mRNA and ZFPM2-AS1 expression levels in the cervical cancer tissue samples. r = 0.524., P = 0.0002. (D) RT-qPCR was carried out to measure miR-511-3p expression in HeLa and SiHa cells that were transfected with either the miR-511-3p inhibitor or NC inhibitor. *P < 0.05 vs the NC inhibitor group. (E, F) Either the miR-511-3p inhibitor or NC inhibitor along with si-ZFPM2-AS1 was introduced into HeLa and SiHa cells. After the transfection, RT-qPCR and Western blotting were carried out to evaluate the change in FGFR2 expression. *P < 0.05 vs group si-NC. #P < 0.05 vs group si-ZFPM2-AS1+NC inhibitor. (G–K) A series of functional experiments, including the CCK-8 assay, flow-cytometric analysis, and Transwell migration and invasion assays (x200 magnification), was conducted to respectively examine the proliferation, apoptosis, migration, and invasiveness of HeLa and SiHa cells that were treated as descried above. *P < 0.05 vs the si-NC group. #P < 0.05 vs group si-ZFPM2-AS1+NC inhibitor. |
To uncover the importance of the miR-511-3p–FGFR2 pathway for the promotion of cervical cancer progression by ZFPM2-AS1, functional experiments were performed on HeLa and SiHa cells that were transfected with either the miR-511-3p inhibitor or NC inhibitor in the presence of si-ZFPM2-AS1. The ZFPM2-AS1 knockdown suppressed HeLa and SiHa cell proliferation (Figure 6G, P < 0.05), promoted their apoptosis (Figure 6H and I, P < 0.05), and decreased their migration (Figure 6J, P < 0.05) and invasiveness (Figure 6K, P < 0.05) in vitro. These phenomena were counteracted by miR-511-3p inhibitor cotransfection. Taken together, these findings revealed that ZFPM2-AS1 contributes to the aggressiveness of cervical cancer in part via upregulation of the output of the miR-511-3p–FGFR2 axis.
Knockdown of ZFPM2-AS1 Reduces the Tumor Growth of Cervical Cancer Cells in vivo
The in vivo tumor xenograft model was set up to address the impact of the ZFPM2-AS1 knockdown on the tumor growth of cervical cancer cells. The results showed that the nude mice injected with the HeLa cells stably transfected with sh-ZFPM2-AS1 manifested slower tumor growth compared with those in the sh-NC group (Figure 7A and B, P < 0.05). In addition, the weight of subcutaneous tumors was significantly lower in the sh-ZFPM2-AS1 group than in the sh-NC group (Figure 7C, P < 0.05). The expression levels of ZFPM2-AS1, miR-511-3p, and FGFR2 in the subcutaneous tumors were determined to verify whether the miR-511-3p–FGFR2 axis is responsible for the tumor growth inhibition caused by the ZFPM2-AS1 knockdown. ZFPM2-AS1 (Figure 7D, P < 0.05) and FGFR2 protein (Figure 7E, P < 0.05) expression was lower, while miR-511-3p (Figure 7F, P < 0.05) expression was higher in the tumor xenografts derived from HeLa cells stably transfected with sh-ZFPM2-AS1. These results indicated that the reduction in ZFPM2-AS1 expression slowed down the tumor growth of cervical cancer cells in vivo, and this effect was mediated by the downregulation of miR-511-3p–FGFR2 axis output.
 |
Figure 7 Downregulation of ZFPM2-AS1 restricts cervical cancer cell growth through the miR-511-3p–FGFR2 axis. (A) HeLa cells stably transfected with either sh-ZFPM2-AS1 or sh-NC were subcutaneously injected into nude mice. The growth curves of groups sh-ZFPM2-AS1 and sh-NC were plotted and analyzed. *P < 0.05 vs the sh-NC group. (B) Representative images of tumor xenografts derived from the HeLa cells stably transduced with either sh-ZFPM2-AS1 or sh-NC. (C) Tumor xenografts were excised from the mice and then weighed. *P < 0.05 vs group sh-NC. (D) ZFPM2-AS1 expression in the tumor xenografts was analyzed via RT-qPCR. *P < 0.05 vs the sh-NC group. (E) Western blotting analysis of FGFR2 protein expression in the tumor xenografts derived from HeLa cells stably transfected with either sh-ZFPM2-AS1 or sh-NC. *P < 0.05 vs group sh-NC. (F) The expression of miR-511-3p was estimated in subcutaneous tumors collected in groups sh-ZFPM2-AS1 and sh-NC. *P < 0.05 vs the sh-NC group. |
Discussion
Extensive research has confirmed aberrant expression of lncRNAs in cervical cancer.21,47,48 The dysregulation of lncRNAs has been demonstrated to be crucial for cervical carcinogenesis and cervical cancer progression through the control of a wide range of aggressive characteristics of cancer cells.49–51 Accordingly, lncRNAs may be promising RNAs for the identification of novel chemotherapeutic medications. In this study, we hypothesized that one of cancer-associated lncRNAs, ZFPM2-AS1, is aberrantly expressed in cervical cancer and is functionally implicated in the malignancy of cervical cancer. Therefore, we employed systematic experimental methods to determine the expression profile of ZFPM2-AS1 in cervical cancer and to characterize the role of ZFPM2-AS1 in cervical cancer.
ZFPM2-AS1 is upregulated in gastric cancer, and its upregulation is significantly associated with tumor size, depth of tumor invasion, differentiation grade, and TNM stage.24 Patients with gastric cancer featuring high ZFPM2-AS1 expression show shorter overall survival and disease-free survival than do the patients with low ZFPM2-AS1 expression.24 Lung adenocarcinoma25 and renal cell cancer26 also manifest high expression of ZFPM2-AS1. Increased ZFPM2-AS1 expression is related to lymph node metastasis, tumor stage, and survival time of patients with renal cell cancer.26 By contrast, the expression profile of ZFPM2-AS1 in cervical cancer has been poorly studied until now. Herein, our results suggest that ZFPM2-AS1 is strongly expressed in both cervical cancer tissue samples and cell lines. Upregulation of ZFPM2-AS1 highly correlated with tumor size, FIGO stage, lymph node metastasis, and shorter overall survival among the patients with cervical cancer.
ZFPM2-AS1 plays tumor-promoting roles in human cancers. For instance, upregulation of ZFPM2-AS increases gastric cancer cell proliferation and reduces apoptosis in vitro and promotes tumor growth in vivo.24 In lung adenocarcinoma, resumption of ZFPM2-AS1 expression can increase cancer cell viability and colony-forming ability in vitro.25 In renal cell cancer, ZFPM2-AS1 overexpression accelerates cancer cell growth and metastasis as well as diminishes cancer cell apoptosis in vitro.26 On the other hand, the functions of ZFPM2-AS1 in cervical cancer have not been clarified. In this study, our results showed that a reduction in ZFPM2-AS1 expression decreased cervical cancer cell proliferation, migration, and invasion and induced apoptosis in vitro as well as diminished tumor growth in vivo.
The molecular mechanisms of action of lncRNAs are diverse. LncRNAs can work as ceRNAs by competitively binding to miRNAs through their miRNA response elements, thereby upregulating miRNA target genes.52 Recently, researchers reported that the cancer-promoting activities of ZFPM2-AS1 are mediated by the MIF–p53 signaling pathway in gastric cancer,24 by the miR-18b-5p–VMA21 axis in lung adenocarcinoma,25 and by miR-137 in renal cell cancer.26 After identifying the expression status and roles of ZFPM2-AS1 in cervical cancer, we next sought to explore the mechanisms of ZFPM2-AS1 knockdown–mediated suppression of the aggressive phenotype of cervical cancer cells in vitro and in vivo.
In this study, we demonstrated a network regulating cervical cancer malignancy; this network is composed of ZFPM2-AS1, miR-511-3p, and FGFR2. First, ZFPM2-AS1 was found to be mainly located in the cytoplasm of cervical cancer cells. Second, bioinformatic analysis indicated that ZFPM2-AS1 harbors a potential binding site for miR-511-3p. Third, luciferase reporter and RIP assays revealed that ZFPM2-AS1 can directly bind to miR-511-3p and interact with the latter in cervical cancer cells. Fourth, miR-511-3p was found to be only weakly expressed in cervical cancer, showing a negative expression correlation with ZFPM2-AS1. Fifth, the ZFPM2-AS1 knockdown raised endogenous miR-511-3p expression and decreased the expression of miR-511-3p’s target (FGFR2) in cervical cancer cells. Sixth, FGFR2 upregulation was proven in cervical cancer and positively correlated with the expression of ZFPM2-AS1. Finally, miR-511-3p inhibition abrogated the regulatory effects of the ZFPM2-AS1 knockdown on FGFR2 expression and on the malignant phenotype of cervical cancer cells.
MiR-511-3p expression is low in prostate cancer and plays an inhibitory part in the aggressiveness of tumor cells in vitro and in vivo.53 Herein, we for the first time showed that miR-511-3p expression is low in cervical cancer tissue samples and cell lines. Further experiments indicated that ectopic miR-511-3p expression slowed the proliferation of cervical cancer cells, induced their apoptosis, and impaired their migratory and invasive abilities in vitro. Further molecular investigation identified FGFR2 as a direct target gene of miR-511-3p in cervical cancer cells. FGFR2, located in human chromosomal region 10q26, is upregulated in cervical cancer tissue samples and cell lines, with an obvious correlation with lymph node metastasis, disease-free survival, and overall survival.41,42 Functionally, FGFR2 is believed to serve as an oncogene enhancing the malignant characteristics of cervical cancer.43 Here, our results confirmed that ZFPM2-AS1 can positively regulate FGFR2 expression by functioning as a ceRNA of miR-511-3p in cervical cancer, thereby pointing to possible diagnostics and therapeutics based on the ZFPM2-AS1–miR-511-3p–FGFR2 axis.
Conclusion
Thus, we validated an oncogenic function of lncRNA ZFPM2-AS1 in cervical cancer. The regulation of miR-511-3p and FGFR2 expression by ZFPM2-AS1 is a mechanism behind the oncogenic role of ZFPM2-AS1 in cervical carcinogenesis and cervical cancer progression. Hence, the ZFPM2-AS1–miR-511-3p–FGFR2 regulatory pathway might be a therapeutic target in cervical cancer.
Ethics Approval and Informed Consent
This study was carried out with the approval of Ethics Committees of Qilu Hospital of Shandong University and in accordance with the Declaration of Helsinki. All participating patients provided written informed consent. The animal experiments were approved by the Institutional Experimental Animal Review Board of Qilu Hospital of Shandong University.
Data Sharing Statement
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Funding
This research was supported by the Analysis of Cervical Cancer Screening in Women of the Shandong Province (2017ws302).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
2. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
3. Wright JD, Huang Y, Ananth CV, et al. Influence of treatment center and hospital volume on survival for locally advanced cervical cancer. Gynecol Oncol. 2015;139(3):506–512. doi:10.1016/j.ygyno.2015.07.015
4. Ma YL, Zhang YS, Zhang F, et al. Methyl protodioscin from Polygonatum sibiricum inhibits cervical cancer through cell cycle arrest and apoptosis induction. Food Chem Toxicol. 2019;132:110655. doi:10.1016/j.fct.2019.110655
5. Handler AS, Henderson VA, Rosenfeld A, Rankin K, Jones B, Issel LM. Illinois breast and cervical cancer program: implementing effective public-private partnerships to assure population health. J Public Health Manage Pract. 2015;21(5):459–466. doi:10.1097/PHH.0000000000000191
6. Banister CE, Liu C, Pirisi L, Creek KE, Buckhaults PJ. Identification and characterization of HPV-independent cervical cancers. Oncotarget. 2017;8(8):13375–13386. doi:10.18632/oncotarget.v8i8
7. He C, Lv X, Huang C, et al. A human papillomavirus-independent cervical cancer animal model reveals unconventional mechanisms of cervical carcinogenesis. Cell Rep. 2019;26(10):2636–2650 e2635. doi:10.1016/j.celrep.2019.02.004
8. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–159. doi:10.1038/nrg2521
9. Fang Y, Fullwood MJ. Roles, functions, and mechanisms of long non-coding RNAs in cancer. Genomics Proteomics Bioinformatics. 2016;14(1):42–54. doi:10.1016/j.gpb.2015.09.006
10. Zhao J, Li L, Han ZY, Wang ZX, Qin LX. Long noncoding RNAs, emerging and versatile regulators of tumor-induced angiogenesis. Am J Cancer Res. 2019;9(7):1367–1381.
11. Lecerf C, Le Bourhis X, Adriaenssens E. The long non-coding RNA H19: an active player with multiple facets to sustain the hallmarks of cancer. Cell Mol Life Sci. 2019;76:4673–4687. doi:10.1007/s00018-019-03240-z
12. Yao F, Wang Q, Wu Q. The prognostic value and mechanisms of lncRNA UCA1 in human cancer. Cancer Manag Res. 2019;11:7685–7696. doi:10.2147/CMAR.S200436
13. Huang J, Chen J, Liu L, Cai X, Yao Z. Progress in the study of long noncoding RNA in tongue squamous cell carcinoma. Oral Surg Oral Med Oral Pathol Oral Radiol. 2019.
14. Lv Y, Huang S. Role of non-coding RNA in pancreatic cancer. Oncol Lett. 2019;18(4):3963–3973. doi:10.3892/ol.2019.10758
15. Duan W, Nian L, Qiao J, Liu NN. LncRNA TUG1 aggravates the progression of cervical cancer by binding PUM2. Eur Rev Med Pharmacol Sci. 2019;23(19):8211–8218. doi:10.26355/eurrev_201910_19128
16. Chang QQ, Chen CY, Chen Z, Chang S. LncRNA PVT1 promotes proliferation and invasion through enhancing Smad3 expression by sponging miR-140-5p in cervical cancer. Radiol Oncol. 2019;53:443–452. doi:10.2478/raon-2019-0048
17. Bai X, Wang W, Zhao P, et al. LncRNA CRNDE acts as an oncogene in cervical cancer through sponging miR-183 to regulate CCNB1 expression. Carcinogenesis. 2019. doi:10.1093/carcin/bgz166
18. Jin L, Ji J, Shi L, Jin S, Pei L. lncRNA HAND2-AS1 inhibits cancer cell proliferation, migration and invasion by downregulating ROCK1 in HPV-positive and negative cervical squamous cell carcinoma. Exp Ther Med. 2019;18(4):2512–2518. doi:10.3892/etm.2019.7830
19. Tong R, Zhang J, Wang C, Li X, Yu T, Wang L. LncRNA PTCSC3 inhibits the proliferation, invasion and migration of cervical cancer cells via sponging miR-574-5p. Clin Exp Pharmacol Physiol. 2019. doi:10.1111/1440-1681.13186
20. Li YJ, Yang Z, Wang YY, Wang Y. Long noncoding RNA ZNF667-AS1 reduces tumor invasion and metastasis in cervical cancer by counteracting microRNA-93-3p-dependent PEG3 downregulation. Mol Oncol. 2019;13:2375–2392. doi:10.1002/mol2.v13.11
21. Zhao H, Zheng GH, Li GC, et al. Long noncoding RNA LINC00958 regulates cell sensitivity to radiotherapy through RRM2 by binding to microRNA-5095 in cervical cancer. J Cell Physiol. 2019;234(12):23349–23359. doi:10.1002/jcp.28902
22. Zhang JJ, Fan LP. Long non-coding RNA CRNDE enhances cervical cancer progression by suppressing PUMA expression. Biomed Pharmacother. 2019;117:108726. doi:10.1016/j.biopha.2019.108726
23. Shao S, Wang C, Wang S, Zhang H, Zhang Y. LncRNA STXBP5-AS1 suppressed cervical cancer progression via targeting miR-96-5p/PTEN axis. Biomed Pharmacother. 2019;117:109082. doi:10.1016/j.biopha.2019.109082
24. Kong F, Deng X, Kong X, et al. ZFPM2-AS1, a novel lncRNA, attenuates the p53 pathway and promotes gastric carcinogenesis by stabilizing MIF. Oncogene. 2018;37(45):5982–5996. doi:10.1038/s41388-018-0387-9
25. Xue M, Tao W, Yu S, et al. lncRNA ZFPM2-AS1 promotes proliferation via miR-18b-5p/VMA21 axis in lung adenocarcinoma. J Cell Biochem. 2019.
26. Liu JG, Wang HB, Wan G, Yang MZ, Jiang XJ, Yang JY. Long noncoding RNA ZFPM2-AS1 promotes the tumorigenesis of renal cell cancer via targeting miR-137. Eur Rev Med Pharmacol Sci. 2019;23(13):5675–5681. doi:10.26355/eurrev_201907_18304
27. Ouyang T, Zhang Y, Tang S, Wang Y. Long non-coding RNA LINC00052 regulates miR-608/EGFR axis to promote progression of head and neck squamous cell carcinoma. Exp Mol Pathol. 2019;111:104321. doi:10.1016/j.yexmp.2019.104321
28. Li S, Zheng K, Pei Y, Wang W, Zhang X. Long noncoding RNA NR2F1-AS1 enhances the malignant properties of osteosarcoma by increasing forkhead box A1 expression via sponging of microRNA-483-3p. Aging. 2019;11.
29. Pan S, Shen M, Zhou M, et al. Long noncoding RNA LINC01111 suppresses pancreatic cancer aggressiveness by regulating DUSP1 expression via microRNA-3924. Cell Death Dis. 2019;10(12):883. doi:10.1038/s41419-019-2123-y
30. Song H, Song J, Lu L, Li S. SNHG8 is upregulated in esophageal squamous cell carcinoma and directly sponges microRNA-411 to increase oncogenicity by upregulating KPNA2. Onco Targets Ther. 2019;12:6991–7004. doi:10.2147/OTT.S214881
31. Ma X, Qi S, Duan Z, et al. Long non-coding RNA LOC554202 modulates chordoma cell proliferation and invasion by recruiting EZH2 and regulating miR-31 expression. Cell Prolif. 2017;50:6. doi:10.1111/cpr.12388
32. Sun T, Yang P, Gao Y. Long non-coding RNA EPB41L4A-AS2 suppresses progression of ovarian cancer by sequestering microRNA-103a to upregulate transcription factor RUNX1T1. Exp Physiol. 2019.
33. Yan J, Jia Y, Chen H, Chen W, Zhou X. Long non-coding RNA PXN-AS1 suppresses pancreatic cancer progression by acting as a competing endogenous RNA of miR-3064 to upregulate PIP4K2B expression. J Exp Clin Cancer Res. 2019;38(1):390. doi:10.1186/s13046-019-1379-5
34. Yan P, Su Z, Zhang Z, Gao T. LncRNA NEAT1 enhances the resistance of anaplastic thyroid carcinoma cells to cisplatin by sponging miR95p and regulating SPAG9 expression. Int J Oncol. 2019;55(5):988–1002. doi:10.3892/ijo.2019.4868
35. Li JH, S L, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42(Database issue):D92–97. doi:10.1093/nar/gkt1248
36. Wong N, Wang X. miRDB: an online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015;43(Database issue):D146–152. doi:10.1093/nar/gku1104
37. Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB. Prediction of mammalian microRNA targets. Cell. 2003;115(7):787–798. doi:10.1016/S0092-8674(03)01018-3
38. Pang W, Zhai M, Wang Y, Li Z. Long noncoding RNA SNHG16 silencing inhibits the aggressiveness of gastric cancer via upregulation of microRNA-628-3p and consequent decrease of NRP1. Cancer Manag Res. 2019;11:7263–7277. doi:10.2147/CMAR.S211856
39. Han S, Han B, Li Z, Sun D. Downregulation of long noncoding RNA CRNDE suppresses drug resistance of liver cancer cells by increasing microRNA-33a expression and decreasing HMGA2 expression. Cell Cycle. 2019;18(19):2524–2537. doi:10.1080/15384101.2019.1652035
40. Bayoumi AS, Sayed A, Broskova Z, et al. Crosstalk between long noncoding RNAs and MicroRNAs in health and disease. Int J Mol Sci. 2016;17(3):356. doi:10.3390/ijms17030356
41. Choi CH, Chung JY, Kim JH, Kim BG, Hewitt SM. Expression of fibroblast growth factor receptor family members is associated with prognosis in early stage cervical cancer patients. J Transl Med. 2016;14(1):124. doi:10.1186/s12967-016-0874-0
42. Kawase R, Ishiwata T, Matsuda Y, et al. Expression of fibroblast growth factor receptor 2 IIIc in human uterine cervical intraepithelial neoplasia and cervical cancer. Int J Oncol. 2010;36(2):331–340.
43. Sun Y, Cheng Y, Zhang Y, Han K. MicroRNA-889-3p targets FGFR2 to inhibit cervical cancer cell viability and invasion. Exp Ther Med. 2019;18(2):1440–1448. doi:10.3892/etm.2019.7675
44. Fu YT, Zheng HB, Zhang DQ, Zhou L, Sun H. MicroRNA-1266 suppresses papillary thyroid carcinoma cell metastasis and growth via targeting FGFR2. Eur Rev Med Pharmacol Sci. 2018;22(11):3430–3438. doi:10.26355/eurrev_201806_15166
45. Li M, Qian Z, Ma X, et al. MiR-628-5p decreases the tumorigenicity of epithelial ovarian cancer cells by targeting at FGFR2. Biochem Biophys Res Commun. 2018;495(2):2085–2091. doi:10.1016/j.bbrc.2017.12.049
46. Yang X, Ruan H, Hu X, Cao A, Song L. miR-381-3p suppresses the proliferation of oral squamous cell carcinoma cells by directly targeting FGFR2. Am J Cancer Res. 2017;7(4):913–922.
47. Mao BD, Xu P, Xu P, Zhong Y, Ding WW, Meng QZ. LINC00511 knockdown prevents cervical cancer cell proliferation and reduces resistance to paclitaxel. J Biosci. 2019;44:2.
48. Feng LL, Shen FR, Zhou JH, Chen YG. Expression of the lncRNA ZFAS1 in cervical cancer and its correlation with prognosis and chemosensitivity. Gene. 2019;696:105–112. doi:10.1016/j.gene.2019.01.025
49. Xu Y, Zhou W, Zhang C, et al. Long non-coding RNA RP11-552M11.4 favors tumorigenesis and development of cervical cancer via modulating miR-3941/ATF1 signaling. Int J Biol Macromol. 2019;130:24–33. doi:10.1016/j.ijbiomac.2019.02.083
50. Zhang L, Liu SK, Song L, Yao HR. SP1-induced up-regulation of lncRNA LUCAT1 promotes proliferation, migration and invasion of cervical cancer by sponging miR-181a. Artif Cells Nanomed Biotechnol. 2019;47(1):556–564. doi:10.1080/21691401.2019.1575840
51. Gao F, Feng J, Yao H, Li Y, Xi J, Yang J. LncRNA SBF2-AS1 promotes the progression of cervical cancer by regulating miR-361-5p/FOXM1 axis. Artif Cells Nanomed Biotechnol. 2019;47(1):776–782. doi:10.1080/21691401.2019.1577883
52. Chan JJ, Tay Y. Noncoding RNA: RNA regulatory networks in cancer. Int J Mol Sci. 2018;19:5. doi:10.3390/ijms19051310
53. Zhang F, Wu Z. Significantly altered expression of miR-511-3p and its target AKT3 has negative prognostic value in human prostate cancer. Biochimie. 2017;140:66–72. doi:10.1016/j.biochi.2017.06.007
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.