Back to Journals » Cancer Management and Research » Volume 11
Long noncoding RNA LINC00511 promotes cell growth and invasion in triple-negative breast cancer by interacting with Snail
Authors Liu R, Wang L, Gan T, Pan T, Huang J, Bai M
Received 30 January 2019
Accepted for publication 30 April 2019
Published 24 June 2019 Volume 2019:11 Pages 5691—5699
DOI https://doi.org/10.2147/CMAR.S203455
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
This paper has been retracted
Ruilei Liu,1,* Liang Wang,2,* Tianyu Gan,1 Tao Pan,3 Jianglong Huang,4 Mingjun Bai3
1Department of Thyroid and Breast Surgery, The Third Affiliated Hospital, Sun Yat-sen University, Guangzhou, People’s Republic of China; 2Department of Gastrointestinal Surgery, The First Affiliated Hospital, Sun Yat-sen University, Guangzhou, People’s Republic of China; 3Department of Vascular Interventional Radiology, The Third Affiliated Hospital, Sun Yat-sen University, Guangzhou, People’s Republic of China; 4Department of Gastrointestinal Surgery, The Third Affiliated Hospital of Sun Yat-sen University, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Purpose: Aberrant long noncoding RNA expression has been frequently reported in cancer research, including in triple-negative breast cancer (TNBC). The aim of the present study was to investigate the involvement of LINC00511 in the progression and prognosis of TNBC.
Materials and methods: The expression level of LINC00511 was examined by RT-PCR in TNBC tissues and in cell lines. MTT and colony formation assays were used to examine the cell growth ability. A Boyden assay was used to examine the cell invasion ability. RNA pull-down and RNA immunoprecipitation (RIP) assays were used to examine the proteins that interacted with LINC00511.
Results: We demonstrated that the LINC00511 expression level was elevated in TNBC tissues when compared with that in normal breast tissues. The downregulation of LINC00511 decreased TNBC cell growth and invasion compared to those of the controls. To explore the molecular mechanisms underlying the biological activity of LINC00511, we identified proteins that bound to LINC00511 with RNA pull-down experiments. We showed that LINC00511 binds to the β-transducin repeat containing (BTRC) E3 ubiquitin protein. Mechanistically, LINC00511 maintained the stability of Snail by impeding its ubiquitination and degradation by the BTRC E3 ubiquitin protein.
Conclusion: Our data suggested that LINC00511 might serve as a novel molecular target for the treatment of TNBC.
Keywords: triple-negative breast cancer, LINC00511, Snail
Plain language summary
Our study aimed to examine the biological role and the underlying mechanism of LINC00511 in TNBC. We demonstrated that LINC00511 expression levels were elevated in TNBC tissues compared to those in the controls. The inhibition of LINC00511 decreased TNBC cell growth and invasion compared to those of the controls. In addition, the downregulation of LINC00511 reversed the epithelial-mesenchymal transition (EMT) phenotype. Mechanistically, LINC00511 maintained the stability of Snail by impeding its ubiquitination and degradation via the BTRC E3 ubiquitin protein. We propose that the LINC00511/Snail axis may be a useful molecular target for the treatment of TNBC.
Introduction
Breast cancer, with its high incidence rate over the past decades, is the most common type of cancer and the second most frequent cause of cancer-related mortality among women.1 During the earliest stages of tumor development, breast cancer cells usually enter the circulation, therefore resulting in the formation of metastatic lesions, which are responsible for ~90% of the breast cancer-related mortality.2,3 Triple-negative breast cancer (TNBC), a subtype of breast cancer, does not grow in response to the hormones estrogen, progesterone, or HER2/neu.4 TNBC characteristically has a higher grade than that of other breast cancer subtypes and tends to metastasize. The five-year survival rate for TNBC is approximately 77% vs the rate of 93% that is observed for other breast cancer types.5 The molecular mechanisms of tumorigenesis in TNBC are still not fully understood. Therefore, it is an urgent need to elucidate the molecular mechanisms underlying breast cancer processes.
Long noncoding RNAs (lncRNAs), a class of noncoding RNAs that are over 200 nucleotides in length, play a significant role in a series of biological processes.6 Recently, studies have reported that the dysregulation of lncRNAs is involved in several pathological states, including cancer.7 LncRNAs exert their function by affecting chromatin remodeling and transcriptional and posttranscriptional regulation.
Snail is a key regulator of the epithelial-mesenchymal transition (EMT) of tumor cells. Mechanistically, Snail can repress the transcription of the cell adhesion molecule E-cadherin and ultimately contributes to the EMT phenotype.8 The elevated expression of Snail has been observed in a series of cancers, including TNBC.9,10
In the present study, we explored the expression and role of LINC00511 in breast cancer. In addition, we investigated the interaction of LINC00511 and Snail in breast cancer cells. Our findings revealed a novel mechanism of LINC00511/Snail interaction, which promoted cell growth and invasion in breast cancer.
Materials and methods
Cell culture and tissue sample collection
The MDA-MB-231 cell line was obtained from the Institute of Chemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China). These cells were cultured in DMEM supplemented with 10% fetal bovine serum.
The TNBC patient samples were collected from the Third Affiliated Hospital of Sun Yat-sen University. This experiment was approved by the Institutional Research Ethics Committee of Sun Yat-sen University. All patients provided written informed consent, and this experiment was conducted in accordance with the Declaration of Helsinki.
RT-PCR analysis
Total RNA was isolated from cells using Trizol reagent (Invitrogen). The first chain cDNA was synthesized using HiScript II Q-RT SuperMix for qPCR (+gDNA wiper, Vanzym, China). RT-PCR analyses were performed on a LightCycler 96 detection system (Roche) according to the manufacturer’s instructions. All mRNA expression levels were normalized to the GAPDH signal.
Production of lentivirus and cell infection
For the lentivirus-mediated suppression of LINC00511, the lentivirus containing LINC00511 shRNA were synthesized by Ruibo Co. Ltd. (Guangzhou, China). A nontargeting scrambled shRNA was also generated as a negative control. The shRNAs were inserted into the pMKO.1-puro vector. Lentiviral particles were obtained by transfecting 293T cells. Viral supernatants were collected 72 hrs after transfection. For cell infection, MDA-MB-231 cells were seeded in six-well plates and infected with lentiviral particles expressing LINC00511 shRNA and scrambled shRNA. After 72 hrs, puromycin (10 µg/mL) was used to select the puromycin-resistant clones.
Colony formation and MTT assay
To perform colony formation assays, 200 cells were seeded in six-well culture plates and cultured for two weeks. After that, we washed the cells three times with PBS and stained them with Giemsa solution. Subsequently, the number of colonies containing ≥50 cells was counted under a microscope. The plate clone formation efficiency was evaluated by using the formula: = (number of colonies/number of cells inoculated)×100%.
An MTT assay was carried out as previously described.11 Briefly, cells were seeded into a 96-well plate and were allowed to grow for 24 hrs. Then, the media were aspirated, and MTT solution was added into each well. After incubation for 30 mins, 150 µL DMSO was added into each well. Finally, the absorbance was read at OD=590 nm.
Cell invasion ability assay
The cell invasion ability was examined with a Boyden assay. The cells were seeded into the upper chambers of the plate (Millipore), which were coated with 150 µg Matrigel (BD Biosciences, Boston, MA, USA). Under the upper chambers, the lower chambers were filled with 500 µL DMEM supplemented with 10% FBS. After incubation for 12 hrs, the cells adhering to the lower surface were fixed with methanol, stained with Giemsa solution and counted.
Flow cytometry assay
To carry out the cell-cycle assay, TNBC cells were harvested after 36 hrs incubation and then washed with PBS. Subsequently, cells were fixed with 70% ice-cold ethanol at 4°C overnight, followed by incubation with propidium iodide and RNase A for 15 mins at 37°C. After washed with cold PBS three times, FACS caliber flow cytometry (BD Biosciences) was used to gain the DNA content of labeled cells.
Western blot assay
Total proteins were extracted from cells with RIPA buffer (Beyotime, China) and were then separated on SDS-PAGE gels, followed by transfer to polyvinylidene fluoride (PVDF) membranes. We blocked the membranes with 3% BSA/TBST and incubated them with primary antibodies at 4°C overnight. We then rinsed the PVDF membranes three times for 5 mins each with TBST and incubated the membranes in HRP-conjugated secondary antibodies for 1 hr at room temperature. We detected the levels of total protein with enhanced chemiluminescence reagents.
RNA pull-down assay
Biotinylated RNA was synthesized in vitro with T7 RNA polymerase and then purified on G-50 Sephadex RNA Columns (Roche). After validating the size with agarose gel electrophoresis, proper RNA secondary structure was folded from the biotin-labeled RNAs following an incubation at different temperatures. Meanwhile, nuclear lysates were harvested from TNBC cancer cells with the addition of prewashed streptavidin–agarose beads. A mixture of folded biotinylated RNAs and nuclear lysates were then sequentially incubated with tRNA and with prewashed streptavidin–agarose beads. The pellets were washed and then boiled with loading buffer. The pulled-down BTRC protein was measured with Western blotting.
RNA immunoprecipitation (RIP) assay
Cells were grown in a 10 cm plate and were lysed in 0.5 mL RIP buffer (25 mM Tris pH 7.4, 150 mM KCl, 0.5 mM DTT, and 0.5% NP-40) containing 100 U/mL RNase inhibitor and protease inhibitors and were centrifuged at 12,000 rpm for 10 mins. The supernatants were incubated separately with anti-Flag, anti-Slug, anti-mouse IgG, or anti-rabbit IgG antibodies at 4°C for 2 hrs with gentle rotation. Forty microliters of protein A/G beads were added and incubated at 4°C for 1 hr. Beads were washed three times with RIP buffer and then once with PBS. RNA was extracted using Trizol, and RT-PCR was performed to analyze the samples.
Tumor xenograft experiments
Female BALB/c nude mice (five-weeks-old) were fed under standard conditions and cared for according to the institutional guidelines for animal care. The animal experiments were approved by the Institutional Animal Care and Use Committee of Sun Yat-sen University. The sh-ctrl and sh-LINC00511 cells were injected subcutaneously into the posterior flank of the mice. We calculated the tumor volumes by using the formula (volume = length × width2/2). Five weeks after implantation, the xenografts were removed from the mice, and the xenografts were weighed. Then, a Ki-67 staining assay was carried out to evaluate the proliferation index.
Results
LINC00511 expression was upregulated in TNBC tissues and was associated with a worse prognosis
First, we used the TCGA database to examine the different lncRNAs in TNBC and normal breast tissues. A total of 1,030 lncRNAs were upregulated, while 369 lncRNAs were downregulated, in TNBC tissues when compared with those in normal breast tissues (fold change>2, Figure S1A and B). Among the elevated lncRNAs, LINC00511 was the most significantly different between the two tissues. Thus, we chose LINC00511 for further study.
Next, we analyzed the expression level of LINC00511 in a cohort of 87 TNBC patients. We found that the expression level of LINC00511 was upregulated in TNBC tissues compared with that in adjacent tissues (Figure 1A). In addition, we found that LINC00511 tended to be elevated in advanced-stage TNBC compared with that in early-stage TNBC patient samples (Figure 1B).
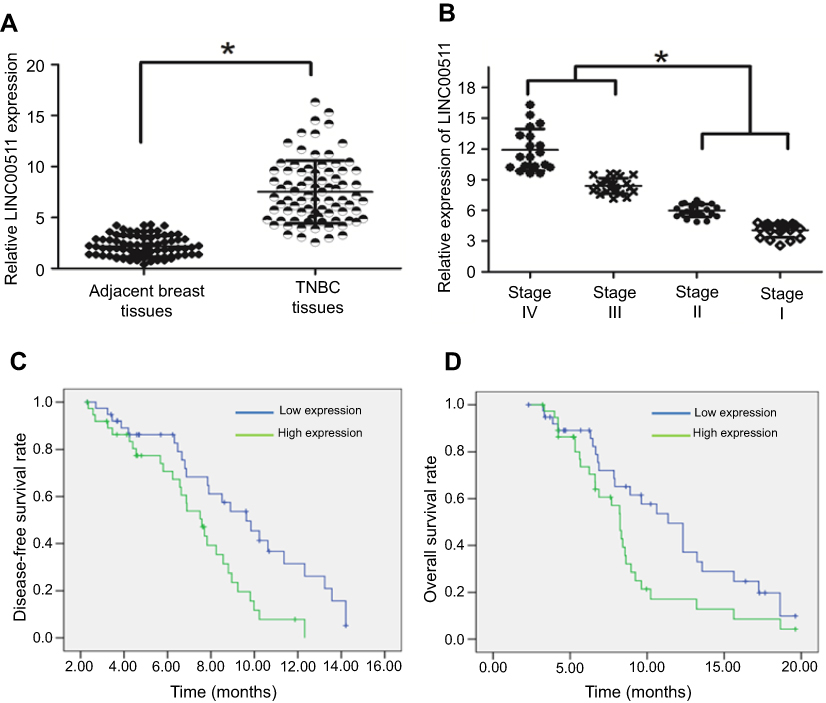 | Figure 1 The LINC00511 expression level was upregulated in TNBC tissues and was associated with a worse prognosis. (A) The expression level of LINC00511 was upregulated in TNBC tissues compared with that in adjacent tissues. (B) The expression levels of LINC00511 are shown in early-stage and advanced-stage TNBC tissue samples compared to those in normal breast tissues. (C) Patients with lower LINC00511 expression had a higher disease-free survival rate than those with higher LINC00511 expression. (D) TNBC patients with higher levels of LINC00511 had shorter overall survival than those with lower levels of LINC00511. *p<0.05. |
Subsequently, we examined whether LINC00511 was a noncoding RNA. Indeed, LINC00511 did not have coding capability, as revealed by an online bioinformatics analysis (
In addition, we explored the association between LINC00511 expression and the clinicopathological features of TNBC patients. As shown in Table 1, elevated LINC00511 expression was significantly associated with tumor size (P=0.000), lymph node metastasis (P=0.014), and late clinical stage (P=0.007). However, LINC00511 expression was not associated with age or smoking. TNBC patients with high expression levels of LINC00511 had worse overall survival rates and disease-free survival rates than those in patients with low expression levels of LINC00511 (Figure 1C and D).
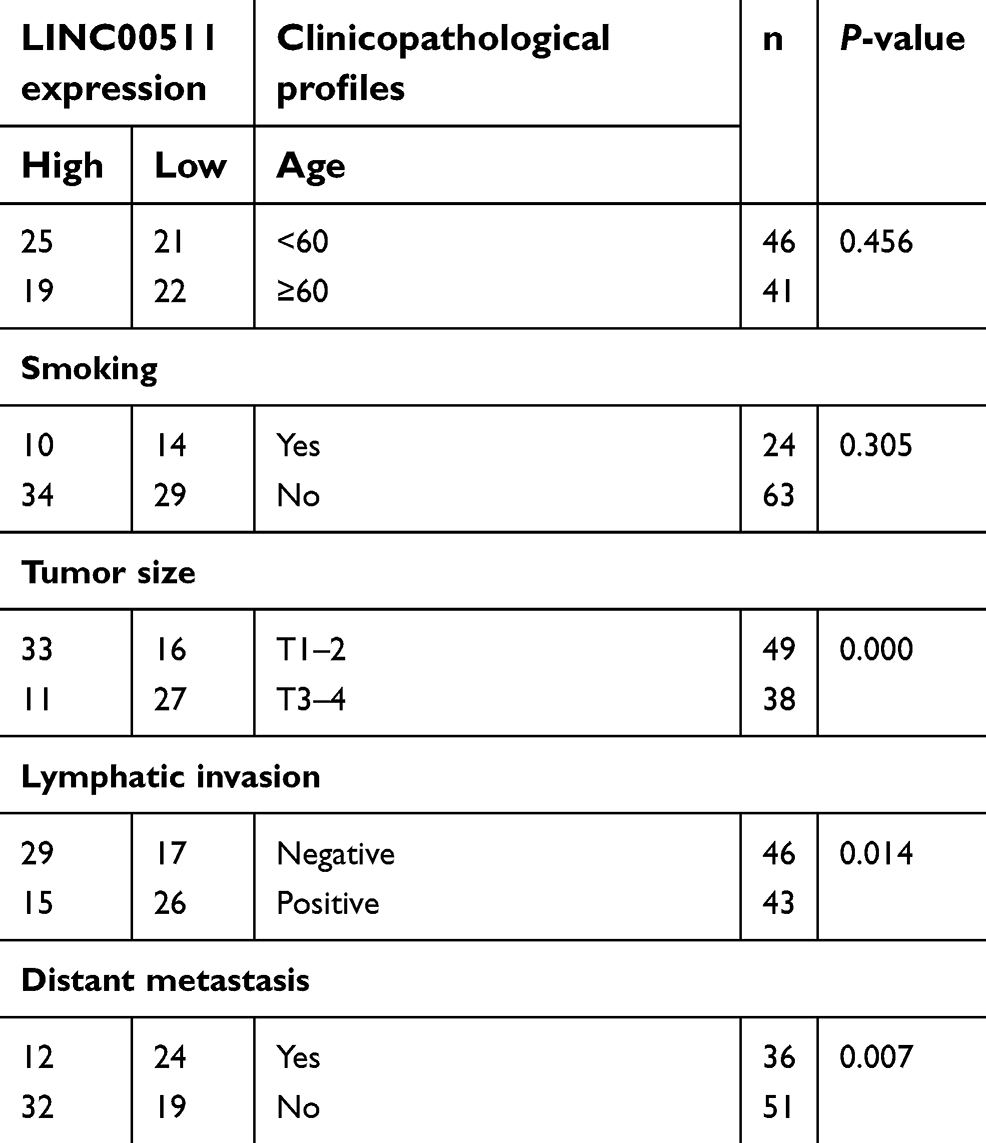 | Table 1 Correlation between LINC00511 expression and the clinicopathological profiles |
Downregulation of LINC00511 inhibited MDA-MB-231 cell growth and invasion
These data suggested that a high expression level of LINC00511 was associated with tumor size and lymph node metastasis; thus, we examined whether LINC00511 affected TNBC cell growth and invasion. The MDA-MB-231 cell line was used to explore the biological role of LINC00511. First, we established MDA-MB-231 cells in which LINC00511 was stably knocked down (sh-LINC00511) (Figure 2A). An MTT assay revealed that LINC00511 downregulation decreased cell viability (Figure 2B) compared to that of the controls. A colony formation assay demonstrated that sh-LINC00511 cells formed smaller and fewer colonies than those formed by sh-ctrl cells (Figure 2C). We subsequently asked whether LINC00511 affected the cell cycle distribution. Flow cytometry analysis demonstrated that MDA-MB-231 cells presented a significantly higher percentage of cells in the G1 phase and a lower percentage of cells in the S phase in the sh-LINC00511 group compared with those in the sh-ctrl group (Figure 2D). The G1/S cell cycle checkpoint proteins (eg, cyclin D1, CDK4, and CDK6) were downregulated in sh-LINC00511 cells compared to those in the controls, as revealed by a Western blot assay (Figure 2E). We speculated that LINC00511 accelerated the cell cycle transition from the G1 phase to the S phase and thus promoted MDA-MB-231 cell growth.
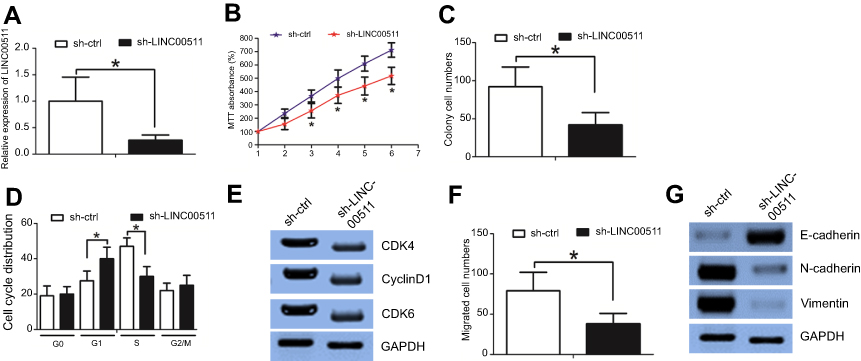 | Figure 2 Downregulation of LINC00511 inhibited MDA-MB-231 cell growth and invasion. (A) The transduction efficiency was validated by an RT-PCR assay. (B) An MTT assay revealed that LINC00511 downregulation decreased cell viability compared to that of the controls. (C) A colony formation assay demonstrated that sh-LINC00511 cells formed smaller and fewer colonies than those formed by sh-ctrl cells. (D) Flow cytometry analysis demonstrated that MDA-MB-231 cells presented a significantly higher percentage of cells in the G1 phase and a lower percentage of cells in the S phase in the sh-LINC00511 group than in the sh-ctrl group. (E) Western blot assays showed altered protein levels. (F) A Boyden assay revealed that LINC00511 downregulation decreased cell invasion ability compared to that in the controls. (G) Western blot assays indicated that LINC00511 downregulation increased E-cadherin expression and decreased N-cadherin and Vimentin expression compared to those of the controls. *p<0.05. |
Subsequently, we examined whether LINC00511 affected the MDA-MB-231 cell invasion ability. A Boyden assay revealed that LINC00511 downregulation decreased the cell invasion ability compared to that of the controls (Figure 2F). It is well known that the EMT phenotype contributes to cell invasion,12 and we then examined whether LINC00511 downregulation affected the EMT phenotype. LINC00511 downregulation increased E-cadherin expression and decreased N-cadherin and vimentin expression compared to those of the controls (Figure 2G).
Taken together, these data revealed that the knockdown of LINC00511 decreased MDA-MB-231 cell growth and invasion.
LINC00511 stabilized Snail by impeding Snail ubiquitination
To explore the molecular mechanisms underlying the biological activity of LINC00511, we examined the protein expression levels of key EMT inducers (eg, Snail, Slug, ZEB1, ZEB2, and Twist) in sh-LINC00511 and sh-ctrl cells. Interestingly, LINC00511 downregulation decreased the Snail protein expression level (Figure 3A) compared to that in the controls. However, LINC00511 downregulation did not affect the Snail mRNA expression level (data not shown). Snail is highly unstable, and the stability of Snail is regulated by several ubiquitin E3 ligases.13 To determine whether the lncRNA LINC00511 affects Snail stability in association with proteasome-mediated degradation, we treated cells with the proteasome inhibitor MG132 to prevent Snail degradation. Compared to those of the controls, in the presence of both MG132 and the lncRNA LINC00511, the Snail protein levels increased (Figure 3B), whereas these levels were significantly decreased following the lncRNA LINC00511 knockdown by siRNA in the absence of MG132 (Figure 3C). However, the addition of MG132 to LINC00511-depleted cells prevented the degradation of endogenous Snail compared to that of the controls (Figure 3C). These results suggested that LINC00511 is involved in the maintenance of Snail stability in a manner that is related to proteasomal degradation. Moreover, LINC00511 knockdown shortened the half-life (Figure 3D), whereas LINC00511 overexpression extended the Snail half-life, compared to that in the controls (Figure 3E).
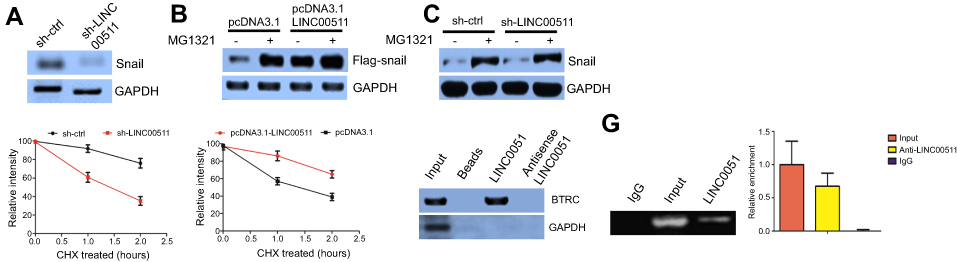 | Figure 3 LINC00511 stabilizes Snail by impeding Snail ubiquitination. (A) LINC00511 downregulation decreased the Snail protein expression level compared to that of the control. (B) Snail protein levels increased in the presence of both MG132 and lncRNA LINC00511 compared to those in the controls. (C) Snail protein levels decreased following lncRNA LINC00511 knockdown by siRNA in the absence of MG132 compared to those of the controls. (D) LINC00511 downregulation shortened the half-life of Snail compared to that of the controls. (E) LINC00511 overexpression extended Snail half-life compared to that of the controls. (F) Western blot assays revealed that BTRC could be detected through its LINC00511 interaction. (G) RIP assays indicated that BTRC is a LINC00511-associated protein. |
To investigate the mechanisms associated with the LINC00511-mediated inhibition of Snail ubiquitination and to identify the factors targeting Snail ubiquitination, we performed pull-down assays to screen for proteins that were potentially associated with LINC00511. RNA-protein complexes that were incubated with in vitro-transcribed biotinylated-LINC00511 and cell lysates were purified with streptavidin magnetic beads. The proteins in the LINC00511-associated protein complexes were then separated with SDS-PAGE, with the corresponding antisense lncRNAs acting as the controls. Among these proteins, the β-transducin repeat containing (BTRC) E3 ubiquitin protein ligase was selected for further study (Figure S1D). To verify the above-mentioned findings, we analyzed LINC00511 pull-down proteins with Western blotting using a BTRC-specific antibody. WB revealed that BTRC could be detected among the proteins that interacted with LINC00511 but not among the proteins associated with either the antisense LINC00511 or with the beads alone (Figure 3F). Furthermore, to verify the interaction between LINC00511 and BTRC, we performed RIP assays for the RNA-protein complexes using BTRC-directed antibodies. Based on the qRT-PCR analysis, compared with the IgG or the anti-BTRC controls, the BTRC-bound complexes showed a significant enrichment of LINC00511. No enrichment of LINC00511 was detected in the IgG complexes (Figure 3G). In summary, the data indicated that BTRC is a LINC00511-associated protein.
Subsequently, we knocked down the expression of the BTRC E3 ubiquitin protein ligase in MDA-MB-231 cells, resulting in increased Snail protein levels (Figure S1E). Consistent with this finding, BTRC overexpression decreased Snail protein levels in MDA-MB-231 cells compared to those in the controls. Furthermore, LINC00511 overexpression rescued BTRC-induced Snail degradation (Figure S1F). Importantly, Snail ubiquitination was significantly diminished by the overexpression of LINC00511 in 293T cells (Figure S1G), whereas the knockdown of endogenous LINC00511 in MDA-MB-231 cells increased the levels of endogenous Snail ubiquitination compared to those of the controls (Figure S1H). These results indicated that LINC00511 blocks BTRC-mediated Snail ubiquitination by inhibiting the interaction between BTRC and Snail, thereby preventing Snail proteasomal degradation.
Inhibition of LINC00511 decreased cell proliferation in vivo
We further examined whether the inhibition of LINC00511 decreased cell proliferation in vivo. sh-LINC00511 or sh-ctrl cells were inoculated into the back of the nude mice. Compared with the sh-ctrl cell-derived xenograft tumors, the sh-LINC00511 cell-derived xenograft tumors grew more slowly (Figure 4A). In addition, the mean weight of the sh-LINC00511 cell-derived xenograft tumors was lower than the sh-ctrl cell-derived xenograft tumors (Figure 4B). Interestingly, a Ki-67 staining assay also revealed that sh-LINC00511 cells had a lower proliferation index than sh-ctrl cells (Figure 4C). Taken together, these results suggested that the inhibition of LINC00511 decreased cell proliferation in vivo.
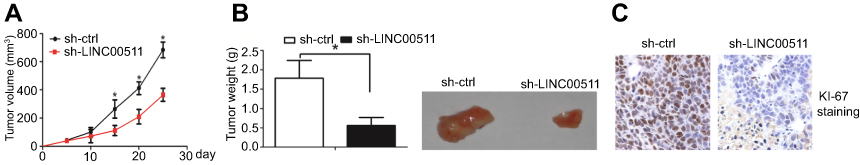 | Figure 4 Inhibition of LINC00511 decreased cell growth in vivo. (A) Compared with the sh-ctrl cell-derived xenograft tumors, the sh-LINC00511 cell-derived xenograft tumors grew more slowly. (B) The mean weight of the sh-LINC00511 cell-derived xenograft tumors was lower than that of the sh-ctrl cell-derived xenograft tumors. (C) A Ki-67 staining assay also revealed that sh-LINC00511 cells had a lower proliferation index than that of sh-ctrl cells. *p<0.05. |
Discussion
Recently, lncRNAs have been reported to influence breast tumorigenesis and progression via different mechanisms, such as regulating gene transcription, splicing, and binding to specific proteins. LncRNAs play a regulatory role in the coordination of protein molecules, so the identification of the specific protein targets of lncRNAs has become the main strategy for elucidating the lncRNA functions and mechanisms of action. In the present study, we performed a series of confirmatory experiments to demonstrate that LINC00511 is a lncRNA that associates with the Snail protein.
EMT is governed by a cohort of transcription factors, including members of the Snail, Zeb, and Twist families. The Snail protein is a core inducer of EMT. Snail directly inhibits the transcription of E-cadherin and of several other intercellular adhesion components. The direct repression of E-cadherin by Snail has been considered to be archetypal of the transcriptional regulation of EMT. In human cancer, the expression of Snail and E-cadherin is inversely correlated, and elevated levels of Snail are associated with tumor aggressiveness and metastasis. Snail is highly unstable, and its stability has emerged as a key determinant of the epithelial phenotype. The stability of Snail is regulated by several ubiquitin E3 ligases, including β-Trcp1, Fbxl14/Fbxl5, and Mdm2.13
In the present study, we demonstrated that LINC00511 elevated the Snail protein expression level but did not affect the Snail mRNA expression level. We speculated that LINC00511 may stabilize Snail by impeding Snail ubiquitination. Using RNA pull-down and RIP assays, we showed that LINC00511 interacted with the BTRC protein. BTRC encodes the βTrCP protein, a member of the F-box protein family and a key component of the SCF (Skp1-Cullin1-F-box)-type ubiquitin ligase E3.14 BTRC was also demonstrated to be an important factor in the process of EMT because of the βTrCP-mediated ubiquitination of Snail in cancer, in which the inhibition of βTrCP resulted in the upregulation of Snail, which could induce EMT.15 We identified a novel mechanism associated with the regulation of Snail stability by demonstrating that LINC00511 binds to BTRC and blocks Snail degradation. Our findings are similar to previous documents, which demonstrated that LINC00511 acted as an oncogene during cancer progression.16,17
In summary, our findings shed light on potential therapeutic strategies by targeting the oncogenic lncRNA00511 in TNBC patients.
Conclusion
In summary, our data revealed that the inhibition of LINC00511 decreased TNBC cell growth and invasion compared to those of the controls. Mechanistically, LINC00511 maintained Snail stability by impeding its ubiquitination and degradation via the BTRC E3 ubiquitin protein. We propose that the LINC00511/Snail axis may be a useful molecular target for the treatment of TNBC.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7–30. doi:10.3322/caac.21332
2. Gupta GP, Massague J. Cancer metastasis: building a framework. Cell. 2006;127:679–695. doi:10.1016/j.cell.2006.11.001
3. Chiang AC, Massague J, Parish S, et al. Molecular basis of metastasis. N Engl J Med. 2008;359:2814–2823. doi:10.1056/NEJMoa0801936
4. Lu L, Mao X, Shi P, et al. MicroRNAs in the prognosis of triple-negative breast cancer: a systematic review and meta-analysis. Medicine. 2017;96:e7085. doi:10.1097/MD.0000000000007085
5. Turashvili G, Lightbody ED, Tyryshkin K, et al. Novel prognostic and predictive MicroRNA targets for triple-negative breast cancer .Faseb J. 2018. fj201800120R. doi:10.1096/fj.201800120R
6. Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function. Nat Rev Genet. 2016;17:47–62. doi:10.1038/nrg.2015.10
7. Prensner JR, Chinnaiyan AM. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011;1:391–407. doi:10.1158/2159-8290.CD-11-0209
8. Cano A, Perez-Moreno MA, Rodrigo I, et al. The transcription factor Snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol. 2000;2:76–83. doi:10.1038/35000025
9. Blanco MJ, Moreno-Bueno G, Sarrio D, et al. Correlation of Snail expression with histological grade and lymph node status in breast carcinomas. Oncogene. 2002;21:3241–3246. doi:10.1038/sj.onc.1205416
10. Muenst S, Daster S, Obermann EC, et al. Nuclear expression of Snail is an independent negative prognostic factor in human breast cancer. Dis Markers. 2013;35:337–344. doi:10.1155/2013/902042
11. Deng X, Ma L, Wu M, et al. miR-124 radiosensitizes human glioma cells by targeting CDK4. J Neurooncol. 2013;114:263–274. doi:10.1007/s11060-013-1179-2
12. Kong P, Chen L, Yu M, et al. miR-3178 inhibits cell proliferation and metastasis by targeting Notch1 in triple-negative breast cancer. Cell Death Dis. 2018;9:1059. doi:10.1038/s41419-018-1091-y
13. Diaz VM, Vinas-Castells R, Garcia de Herreros A. Regulation of the protein stability of EMT transcription factors. Cell Adhes Migration. 2014;8:418–428. doi:10.4161/19336918.2014.969998
14. He N, Li C, Zhang X, et al. Regulation of lung cancer cell growth and invasiveness by beta-TRCP. Mol Carcinog. 2005;42:18–28. doi:10.1002/(ISSN)1098-2744
15. Zhou BP, Deng J, Xia W, et al. Dual regulation of Snail by GSK-3beta-mediated phosphorylation in control of epithelial-mesenchymal transition. Nat Cell Biol. 2004;6:931–940. doi:10.1038/ncb1173
16. Lu G, Li Y, Ma Y, et al. Long noncoding RNA LINC00511 contributes to breast cancer tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J Exp Clin Cancer Res. 2018;37:289. doi:10.1186/s13046-018-0945-6
17. Xu S, Kong D, Chen Q, Ping Y, Pang D. Oncogenic long noncoding RNA landscape in breast cancer. Mol Cancer. 2017;16:129. doi:10.1186/s12943-017-0696-6
Supplementary Material
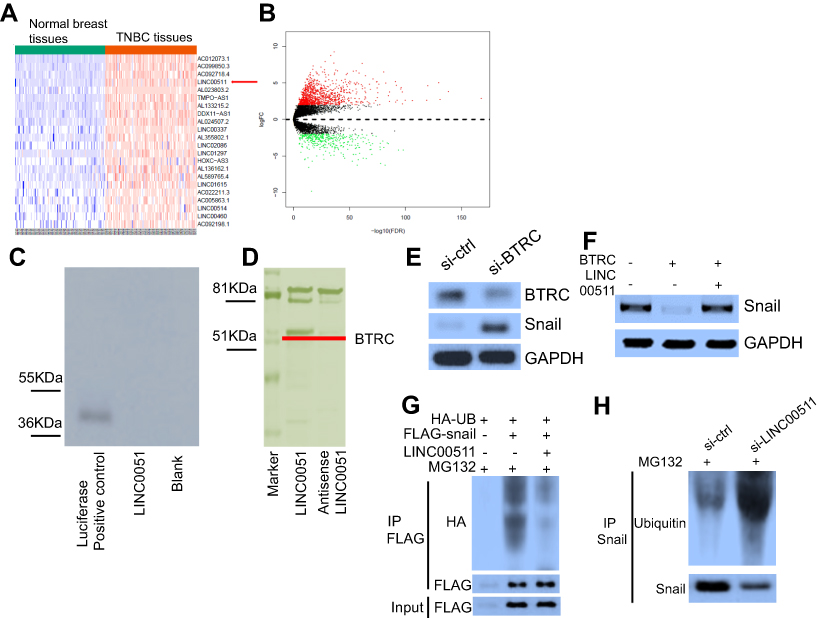 | Figure S1 (A) and (B) The downregulated and upregulated lncRNAs between the TNBC and normal tissues in the TCGA database, as shown by the heat map and volcano plot. (C) An in vitro translation assay further confirmed that LINC00511 did not have protein-coding capability. (D) Identification of the LINC00511–protein complex with an incubation of biotinylated-LINC00511 with protein extracts by coomassie blue staining assay. The β-transducin repeat containing (BTRC) E3 ubiquitin protein ligase was therefore selected as the candidate protein. (E) BTRC E3 ubiquitin protein ligase downregulation resulted in increased Snail protein levels compared to those of the controls. (F) BTRC overexpression decreased Snail protein levels in MDA-MB-231 cells compared to those of the controls. LINC00511 overexpression rescued BTRC-induced Snail degradation. (G) and (H) 293T or MDA-MB-231 cells were transfected with the indicated vectors and treated with MG132 for 6 hrs. Then, cell lysates were immunoprecipitated using an antibody against Flag (G) or HA antibody (H). The precipitates and inputs were analyzed by immunoblotting. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.