Back to Journals » OncoTargets and Therapy » Volume 13
Long Non-Coding RNA TP73-AS1 Promotes the Development of Lung Cancer by Targeting the miR-27b-3p/LAPTM4B Axis
Authors Jiang Q, Xing W, Cheng J, Yu Y
Received 12 October 2019
Accepted for publication 17 February 2020
Published 20 July 2020 Volume 2020:13 Pages 7019—7031
DOI https://doi.org/10.2147/OTT.S234443
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Qingfeng Jiang, Wenqun Xing, Jinhua Cheng, Yongkui Yu
Department of Thoracic Surgery, The Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital, Zhengzhou, Henan, People’s Republic of China
Correspondence: Wenqun Xing
Department of Thoracic Surgery, The Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital, No. 127 Dongming Road, Zhengzhou 450008, Henan, People’s Republic of China
Tel +86-0371-65588265
Email [email protected]
Purpose: Long non-coding RNA P73 antisense RNA 1T (TP73-AS1) is a newly discovered lncRNA involved in the occurrence and development of several cancers. However, its role in lung cancer has not been well investigated yet.
Methods: The expressions of TP73-AS1, microRNA-27b-3p (miR-27b-3p) and lysosomal-associated protein transmembrane-4 Beta (LAPTM4B) were detected by quantitative real-time polymerase chain reaction (qRT-PCR). The cell proliferation, apoptosis, migration and invasion were detected by 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), Annexin V-FITC/PI and transwell assays, respectively. Tumor xenografts were applied to explore the role of TP73-AS1 in vivo. The target relationship was predicted by StarBase v.2.0 or TargetScan and confirmed by luciferase reporter assay. Pearson’s coefficient assay was applied to assess the expression correlation between two groups. Protein expression levels were detected by Western blot.
Results: We found that TP73-AS1 was strikingly up-regulated in lung cancer tissues and cells. TP73-AS1 depletion inhibited the growth and metastasis of lung cancer cells in vitro. Furthermore, TP73-AS1 could act as an endogenous sponge by directly binding miR-27b-3p, and a notable inverse correlation between them was also discovered. Importantly, knockdown of miR-27b-3p could reverse the inhibitory effects of TP73-AS1 depletion on the growth and metastasis of lung cancer cells. Besides, LAPTM4B was directly targeted by miR-27b-3p and could be co-regulated by TP73-AS1 and miR-27b-3p in lung cancer cells. Silencing TP73-AS1 hampered tumor growth by regulating miR-27b-3p/LAPTM4B axis in vivo.
Conclusion: TP73-AS1 promoted the progression of lung cancer through regulating miR-27b-3p/LAPTM4B axis and it might be a potential target for diagnosis and treatment of lung cancer.
Keywords: lung cancer, TP73-AS1, miR-27b-3p, LAPTM4B, proliferation, metastasis
Introduction
Lung cancer is a pervasive life-threatening neoplasm worldwide with the topmost morbidity in males and ranks third in females.1 Lung cancer is split into small cell lung cancer and non-small cell lung cancer (NSCLC), includes lung adenocarcinoma (ADC) and squamous cell carcinoma (SCC), which occupy 80–85% proportion in all lung cancer cases.2 There were numerous researches on regulatory factors in lung cancer. However, the underlying molecular regulation mechanisms of these biomarkers are still unclear. So, it is a great challenge in grasping the molecular mechanism of lung cancer in discerning novel molecular markers.
Long non-coding RNA (lncRNA) was discovered as RNA molecule longer than 200 nucleotides. lncRNAs have been widely concerned on account of their rising roles in cancer.3,4 Statistically, many epigenetic modifiers containing lncRNAs are involved in regulating the progression of various cancers.5,6 Besides, lncRNAs have been referred to a mass of transcriptional regulatory processes, for instance, sponging activity of miRNAs.7 Sponging activity suggested that lncRNAs and coding RNAs might co-regulate certain biological processes, during which they were mutually conditioned by their ability to competitively bind to miRNAs, known as competitive endogenous RNA (ceRNA).8 However, the specific molecular mechanism of how lncRNAs competitively bind to miRNAs to regulate lung cancer progression remains unclear.
Long non-coding RNA P73 antisense RNA 1T (TP73-AS1), a lncRNA situated on chromosome 1p36, was implicated in the progression of several cancers. For example, silencing TP73-AS1 retarded cell proliferation and tumor growth in esophageal squamous cell carcinoma.9 TP73-AS1 could act as an oncogene to facilitate cell proliferation and invasion by sponging miR-142 through HMGB1/RAGE pathway in glioma.10 And TP73-AS1 also could promote breast cancer cell growth and metastasis by miR-200a/ZEB1 axis.11 Not only that, but TP73-AS1 also served as a tumor suppressor in bladder cancer through the EMT pathway.12 However, its role and function in lung cancer have rarely been reported.
MicroRNAs (miRNAs) are an endogenous single-stranded ncRNAs with roughly 22 nucleotides.13 They could take part in a variety of cellular processes by targeting the 3ʹUTR of mRNA.14 In previous studies, it has been reported that miR-27b-3p was exceptionally constrained in lung cancer.15 And miR-27b-3p was enormously declined in lung cancer tissues and could be served as a tumor-inhibiting factor to hinder cell growth via targeting Fzd7.16 What is interesting is that high expression of miR-27b was associated with poor prognosis of breast cancer, interference with miR-27b could retard tumor growth, migration and invasion, and miR-27b was a cell-dependent oncogene in breast cancer.17,18 Lysosomal-associated protein transmembrane-4 Beta (LAPTM4B), a novel oncogene, promoting tumorigenesis and might be a potential prognostic biomarker in many types of cancer, including breast cancer,19 gastric cancer,20 ovarian cancer21 and lung cancer.22 Although the function of LAPTM4B has been reported in lung cancer, there was a few report on how LAPTM4B was regulated by lncRNAs and miRNAs to participate in lung cancer progression.
And up to now, the molecular mechanism of TP73-AS1 regulation of lung cancer is not sufficient. Hence, the study mainly to elucidate the role of TP73-AS1/miR-27b-3p/LAPTM4B network in the progression of lung cancer, hoping to provide new therapeutic targets for lung cancer.
Materials and Methods
Clinical Samples
A total of 49 specimens were obtained from the Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital. Small pieces of the samples were maintained in the liquid nitrogen. Every patient in the study had received no other therapies before operation and signed the informed consent. The research was approved by the Ethics Committee of the Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital in accordance with the National Institutes of Health guideline of the experiment.
Cell Lines and Culture
Normal human bronchial epithelial cell line 16HBE14o was purchased from Be Na collection (Beijing, China), and lung cancer cell lines A549 and H1650 were obtained from American Type Culture Collection (ATCC; Manassas, VA, USA). They were grown in RPMI 1640 with 10% fetal bovine serum (Gibco, Waltham, MA, USA) and 1% penicillin/streptomycin at 37°C in 5% CO2.
Transfection Assay
Short hairpin RNA (shRNA) specifically against TP73-AS1 (sh-TP73-AS1) and corresponding control sh-NC were obtained from Santa Cruz Biotechnology (Dallas, TX, USA). MiR-27b-3p mimics, miR-27b-3p inhibitor and their NCs were acquired from RiboBio (Guangzhou, China). Overexpression vectors (TP73-AS1 and LAPTM4B) and control (pcDNA) were bought from SyngenTech (Beijing, China). Transfection was performed with Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA).
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
Total RNA was extracted by Trizol reagent (Invitrogen). PrimeScript RT-polymerase (Takara, Dalian, China) was applied to reverse transcribe RNA into cDNA. Then, RT-qPCR was performed on the cDNA (Sangon, Shanghai, China). For TP73-AS1 and LAPTM4B, GAPDH was used as an internal control. The relative expression of miR-27b-3p was standardized to U6. The results were computed by 2−∆∆Ct means. Sequence of primers: TP73-AS1 (Forward: 5ʹ-CCGGTTTTCCAGTTCTTGCA-3ʹ; Reverse: 5ʹ-GCCTCACAGGGAAACTTCATG-3ʹ); LAPTM4B (Forward: 5ʹ-AAGACCATTAGAAAGCACCAGG-3ʹ; Reverse: 5ʹ-ACCAATCTAGGGCAGA ACACTTA-3ʹ); GAPDH (Forward: 5ʹ-ACGGATTTGGTCGTATTGGGCG-3ʹ; Reverse: 5ʹ-CTCCTGGAAGATGGTGATGG-3ʹ); miR-27b-3p (Forward: 5ʹ-AGGGTTCACAGTGGCTAAG-3ʹ; Reverse: 5ʹ-GAGAGGAGAGGAAGAGGGAA-3ʹ); U6 (Forward: 5ʹ-CGGGTGCTCGCTTCGCAGC-3ʹ; Reverse: 5ʹ-CCAGTGCAGGGTCCGAGGT-3ʹ).
Cell Proliferation and Apoptosis Analyses
Cells were grown in 96-well plates. We recorded the proliferation of cells every 24 h for four times after transfection. The cells were then treated with 20 µL 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (5 mg/mL) and cultured at 37°C for 4 h. After discarding the medium, 150 µL dimethyl sulfoxide (DMSO) was added to lyse the cells. The absorbance was measured at 570 nm by a UV microplate reader (Tecan Austria GmbH, Groedig, Austria).
For detection cell apoptosis, cells were plated in 6-well plates. After 24 h, transfection was carried out and the culture was continued for 48 h. Then, FITC-Annexin V and propidium iodide (Sigma, Santa Clara, CA) were used for double staining, followed by flow cytometry.
Transwell Assay
The 24-well transwell chamber was applied to measure cell migration without Matrigel and invasion with Matrigel (Corning Life Sciences, Corning, NY, USA). Cells were harvested at 24 h after transfection and cell suspensions were produced using serum-free medium and seeded into the upper chamber with 100 µL per well. The lower chamber was filled with DMEM containing 10% FBS to induce cell migration or invasion. Cells were cultivated for one day. Finally, Cells on the lower surface of the chamber were fixed with ethanol and dyed with crystal violet. Migrating and invading cells were counted by an optical microscope.
Dual-Luciferase Reporter Assay
TP73-AS1 wild type (WT-TP73-AS1) and mutant type (MUT-TP73-AS1) containing miR-27b-3p interacting site or not were co-transfected into cells with miR-NC or miR-27b-3p mimics by using the Lipofectamine 2000 (Invitrogen). Wild type (WT-LAPTM4B 3ʹUTR) and mutant type (MUT-LAPTM4B 3ʹUTR) plasmids of LAPTM4B containing miR-27b-3p targeting sites or not were also transfected into cells. Cells were collected two days after transfection and luciferase activity was examined by luciferase reporter system (Promega, Madison, WI, USA).
Western Blot Assay
First, RIPA buffer (Sigma) was used to extract the total protein, and the concentration was detected by BCA reagent (Thermo Fisher Scientific, Waltham, MA, USA). Then, protein lysates (20 µg) were then subjected to SDS-PAGE and Western blotting. PVDF membranes (Thermo Fisher Scientific) were immersed in 5% skim milk powder for 2 h, and the primary antibodies including LAPTM4B, Bcl-2, Bax, cleaved-caspase3 and GAPDH (all from Thermo Fisher Scientific) were incubated at 4°C overnight. Finally, membranes were probed with the HRP-labeled secondary antibody (Thermo Fisher Scientific) for 1 h at 37°C. Antibody binding was detected by enhanced chemiluminescence using the ECL assay (Amersham Biosciences, Buckinghamshire, UK).
Animal Experiments
All animal protocols were approved by the Animal Care and Use Committee of The Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital. Sh-TP73-AS1 or sh-NC transfected H1650 cells (7×106) were injected subcutaneously into the 4-week-old nude mice. The tumor volume was examined every five days for a total of 5 times. Then, the mice were euthanized and the tumor weight was measured. In addition, the tumor tissues were stored at −80°C for RNA and protein extraction.
Statistical Analysis
Each experiment was performed at least three times. Data were expressed as mean ± standard deviation (SD). Statistical analysis was performed by Student’s t-test using SPSS 22.0 software. *P<0.05 was considered statistically significant.
Results
TP73-AS1 Expression Was Up-Regulated in Lung Cancer Tissues and Cell Lines
To explore the effect of TP73-AS1 in lung cancer, we first examined TP73-AS1 expression in tumor tissues and adjacent non-tumor tissues from 49 patients with lung cancer by qRT-PCR. The results showed that TP73-AS1 expression in lung cancer tissues was extremely elevated compared with adjacent non-tumor tissues (Figure 1A). Also, the expression of TP73-AS1 was markedly increased in lung cancer cell lines A549 and H1650 compared to human normal bronchial epithelial cell 16HBE14o (Figure 1B). These data indicated that TP73-AS1 might be a biomarker in lung cancer.
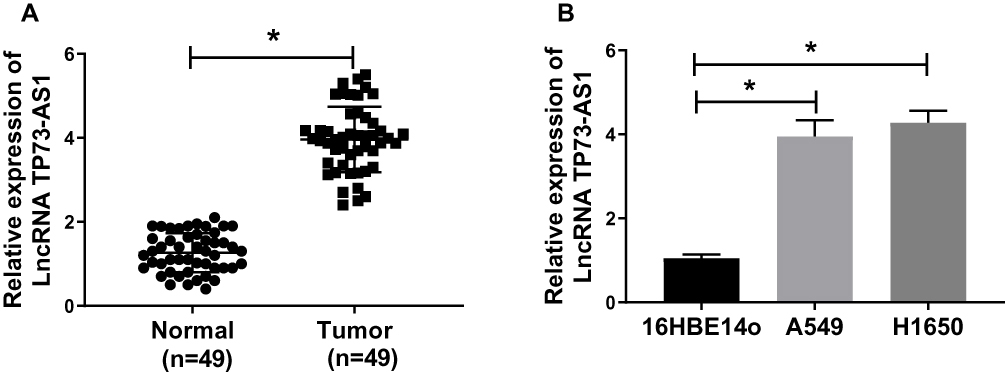 |
Figure 1 TP73-AS1 expression was up-regulated in lung cancer tissues and cell lines. (A) TP73-AS1 expression was measured by qRT-PCR in 49 paired lung cancer tissues compared with adjacent non-tumor tissues. (B) The expression of TP73-AS1 in human normal bronchial epithelial cell 16HBE14o and human lung cancer cell lines A549 and H1650 was detected. *P<0.05. |
Silencing TP73-AS1 Repressed Lung Cancer Cell Proliferation, Migration and Invasion and Facilitated Apoptosis in vitro
To explore the effect of TP73-AS1 on lung cancer cells, we conducted the TP73-AS1 loss-of-function experiments in A549 and H1650 cells. The expression level of TP73-AS1 was significantly declined in A549 and H1650 cells after transfection with sh-TP73-AS1 compared with the cells transfected with sh-NC (Figure 2A). The results of MTT assay demonstrated that knockdown of TP73-AS1 remarkably impeded the cell proliferation in A549 and H1650 cells (Figure 2B and C). Moreover, further apoptosis analysis with Annexin V-FITC/PI showed that the cell apoptosis rate in sh-TP73-AS1 group was prominently promoted in contrast to that in sh-NC group (Figure 2D). To confirm the role of TP73-AS1 in tumor metastasis of lung cancer, we investigated whether TP73-AS1 affected cell migration and invasion in vitro. The results of transwell assay showed that the capacities of cell migration and invasion were evidently impeded under TP73-AS1 inhibition in A549 and H1650 cells (Figure 2E and F). The above data indicated that silencing TP73-AS1 repressed proliferation, migration, invasion and promoted apoptosis of lung cancer cells in vitro.
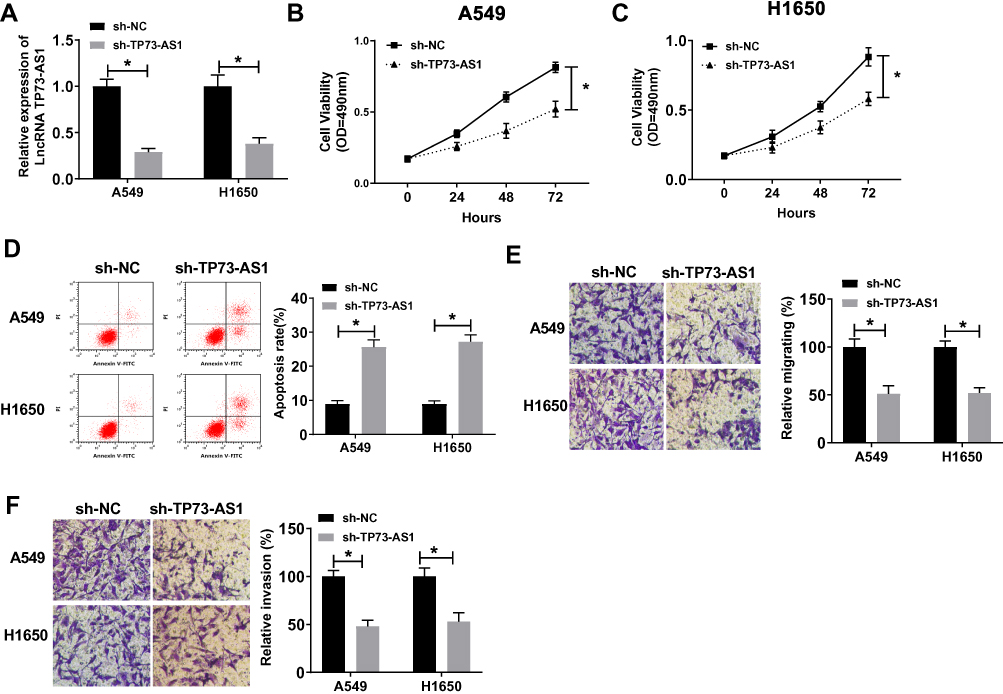 |
Figure 2 Knockdown of TP73-AS1 hindered lung cancer cell proliferation, metastasis and induced apoptosis in vitro. (A) QRT-PCR was used to analyze the transfection efficiency of sh-TP73-AS1 in A549 and H1650 cells. (B, C) MTT assay was conducted to evaluate the impact of interference TP73-AS1 on cell proliferation in A549 and H1650 cells. (D) Flow cytometry analysis was utilized to determine the cell apoptosis rate of A549 and H1650 cells transfected with sh-TP73-AS1 or sh-NC. (E, F) Cell migration and invasion abilities were, respectively, examined by transwell migration and invasion assay both in A549 and H1650 cells. *P<0.05. |
MiR-27b-3p Served as a Direct Target of TP73-AS1
Increasing studies have reported the correlation between lncRNAs and miRNAs in various cancer types, including lung cancer. In this study, we forecasted that miR-27b-3p might be a target gene of TP73-AS1 by StarBase v.2.0 (Figure 3A). To further confirm the correlation between them, the luciferase reporter assay was used in the study. It was found that co-transfection of the luciferase reporter plasmid containing the WT-TP73-AS1 with miR-27b-3p mimics in A549 and H1650 cells obviously decreased the reporter activity compared with the plasmid containing MUT-TP73-AS1 and miR-27b-3p mimics (Figure 3B and C). Besides, miR-27b-3p expression could be inhibited by TP73-AS1 overexpression, while interference with TP73-AS1 enhanced the expression of miR-27b-3p in A549 and H1650 cells (Figure 3D and E). More than that the miR-27b-3p expression level was aberrantly dwindled in lung cancer tissues (Figure 3F). A significant inverse correlation between TP73-AS1 and miR-27b-3p was also observed (Figure 3G). These results revealed that miR-27b-3p could act as a direct target of TP73-AS1 in lung cancer.
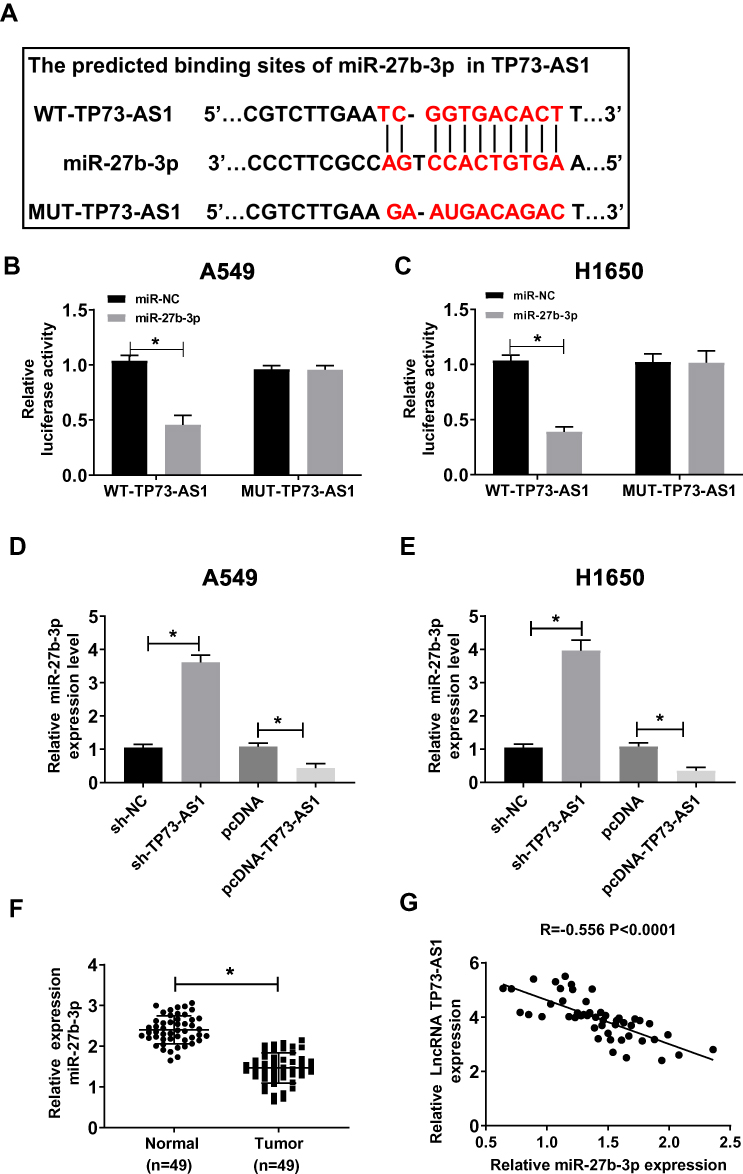 |
Figure 3 miR-27b-3p was a direct target of TP73-AS1. (A) The putative binding sites of TP73-AS1 and miR-27b-3p were predicted by StarBase v.2.0. (B and C) The luciferase activity of A549 and H1650 cells co-transfected WT-TP73-AS1 or MUT-TP73-AS1 with miR-27b-3p or miR-NC was detected. (D, E) miR-27b-3p expression was detected in A549 and H1650 cells transfected with sh-TP73-AS1, pcDNA-TP73-AS1 or their NCs. (F) miR-27b-3p expression was examined by qRT-PCR in 49 paired lung cancer tissues compared to adjacent normal tissues. (G) Correlations between the expression of TP73-AS1 and miR-27b-3p were analyzed by Spearman’s test (R= –0.556, P<0.0001). *P<0.05. |
The Inhibitory Effect of Silencing TP73-AS1 on the Growth and Metastasis of Lung Cancer Cells Was Reversed by Anti-miR-27b-3p
In view of the inhibition effect of TP73-AS1 on miR-27b-3p expression in lung cancer, we further explored whether miR-27b-3p regulating the function of TP73-AS1. We found that silencing TP73-AS1 significantly impeded cell proliferation in A549 and H1650 cells. However, compared with sh-TP73-AS1 + anti-miR-NC groups, the cell proliferation in sh-TP73-AS1 + anti-miR-27b-3p co-transfected A549 and H1650 cells was enormously increased and could basically restore to the original growth activity (Figure 4A and B). In addition, the cell apoptosis rate of A549 and H1650 cells transfected with sh-TP73-AS1 was drastically elevated, whereas this promotion was counteracted by co-transfection of anti-miR-27b-3p (Figure 4C and D). On the other hand, the suppressive impact of silencing TP73-AS1 on the migration and invasion of A549 and H1650 cells was also reversed by anti-miR-27b-3p (Figure 4E and F). The results showed that interference with miR-27b-3p could reverse the inhibition effects of silencing TP73-AS1 on the progression of lung cancer cells in vitro.
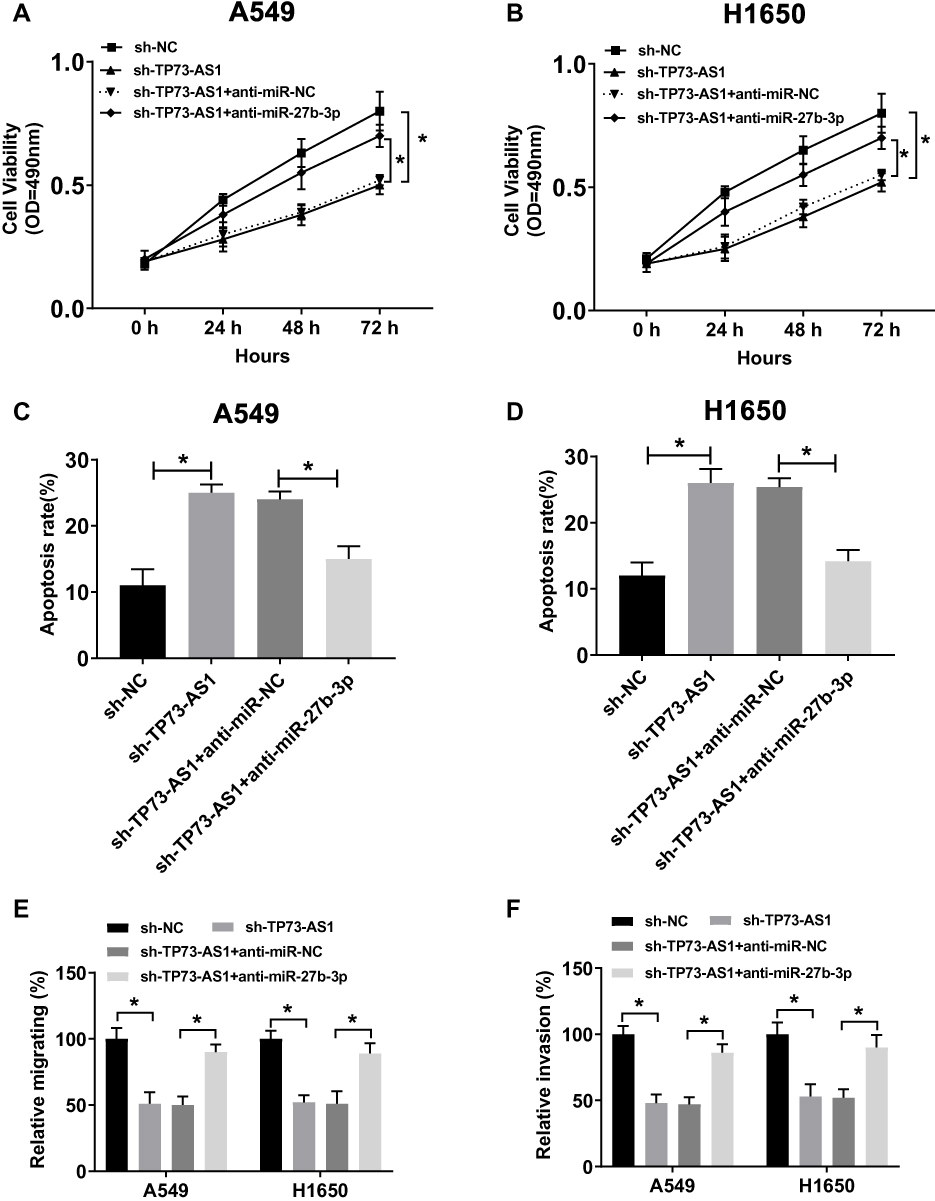 |
Figure 4 Interference with miR-27b-3p inverted the inhibitory effect of silencing TP73-AS1 on cell growth and metastasis of lung cancer cells in vitro. (A, B) Cell viability was assayed in sh-NC, sh-TP73-AS1 transfected or sh-TP73-AS1 + anti-miR-NC or sh-TP73-AS1 + anti-miR-27b-3p co-transfected A549 and H1650 cells by MTT. (C, D) Cell apoptosis rate was examined in sh-NC, sh-TP73-AS1 transfected or sh-TP73-AS1 + anti-miR-NC or sh-TP73-AS1 + anti-miR-27b-3p co-transfected A549 and H1650 cells by flow cytometry. (E, F) Transwell assay was used to investigate changes in migration and invasion of A549 and H1650 cells. *P<0.05. |
LAPTM4B Was a Direct Target of miR-27b-3p
MiRNAs could regulate the expression of target genes posttranscriptionally by targeting 3ʹUTR of mRNAs in various cancers. To explore the detailed mechanism of miR-27b-3p in the progression of lung cancer, TargetScan was used to predict the putative targets of miR-27b-3p and LAPTM4B was predicted as its target (Figure 5A). Luciferase activity assay showed that miR-27b-3p mimics markedly decreased the luciferase activity of the WT-LAPTM4B in A549 and H1650 cells, while there was no distinct change in the cells transfected with MUT-LAPTM4B (Figure 5B and C). What is more, LAPTM4B mRNA and protein expressions were repressed by miR-27b-3p mimics in A549 and H1650 cells, while miR-27b-3p inhibitor promoted the mRNA and protein levels of LAPTM4B (Figure 5D and E). Meanwhile, we found that the mRNA and protein expressions of LAPTM4B were up-regulated in lung cancer cell lines A549 and H1650 compared with 16HBE14o cells (Figure 5F and G). These data indicated that LAPTM4B could be directly targeted by miR-27b-3p in lung cancer cells.
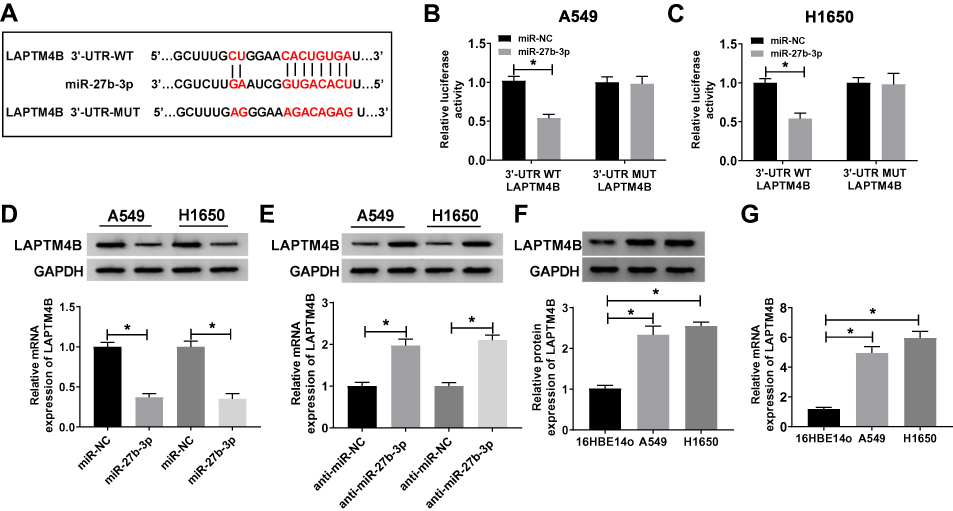 |
Figure 5 MiR-27b-3p was able to directly target LAPTM4B. (A) The putative binding sites of miR-27b-3p and LAPTM4B were predicted by TragetScan. (B, C) The relative luciferase activity of A549 and H1650 cells co-transfected WT-LAPTM4B or MUT-LAPTM4B with miR-27b-3p or miR-NC was detected (D, E) The mRNA and protein expression of LAPTM4B in A549 and H1650 cells were measured by qRT-PCR and Western blot transfected with miR-NC, miR-27b-3p, anti-miR-NC or anti-miR-27b-3p. (F, G) The mRNA and protein expression of LAPTM4B in human normal bronchial epithelial cell 16HBE14o and human lung cancer cell lines A549 and H1650 were detected. *P<0.05. |
MiR-27b-3p Inhibited Lung Cancer Cells Growth and Metastasis by Regulation of LAPTM4B
In consideration of the inhibitory effect of miR-27b-3p on LAPTM4B expression in lung cancer, we further explored whether miR-27b-3p had the same effect on the function of LAPTM4B. The MTT and Annexin V-FITC/PI assays showed that miR-27b-3p could hinder cell proliferation and accelerate apoptosis in A549 and H1650 cells, whereas this facilitation and inhibition could invert by overexpression of LAPTM4B (Figure 6A–C). At the same time, we examined the changes in the levels of apoptosis-related protein Bcl-2, Bax, cleaved-caspase3 and caspase3. MiR-27b-3p-mediated increase in cleaved-caspase3 activity, as well as miR-27b-3p-elicited decrease in the ratio of Bcl-2/Bax, was remarkably attenuated after overexpression of LAPTM4B in A549 and H1650 cells (Figure 6D and E). Analogously, it was found that overexpression of LAPTM4B could alleviate the inhibition of miR-27b-3p on migration and invasion of A549 and H1650 cells (Figure 6F and G). The above results supported that miR-27b-3p impeded the growth and metastasis of lung cancer cells by inhibiting the expression of LAPTM4B.
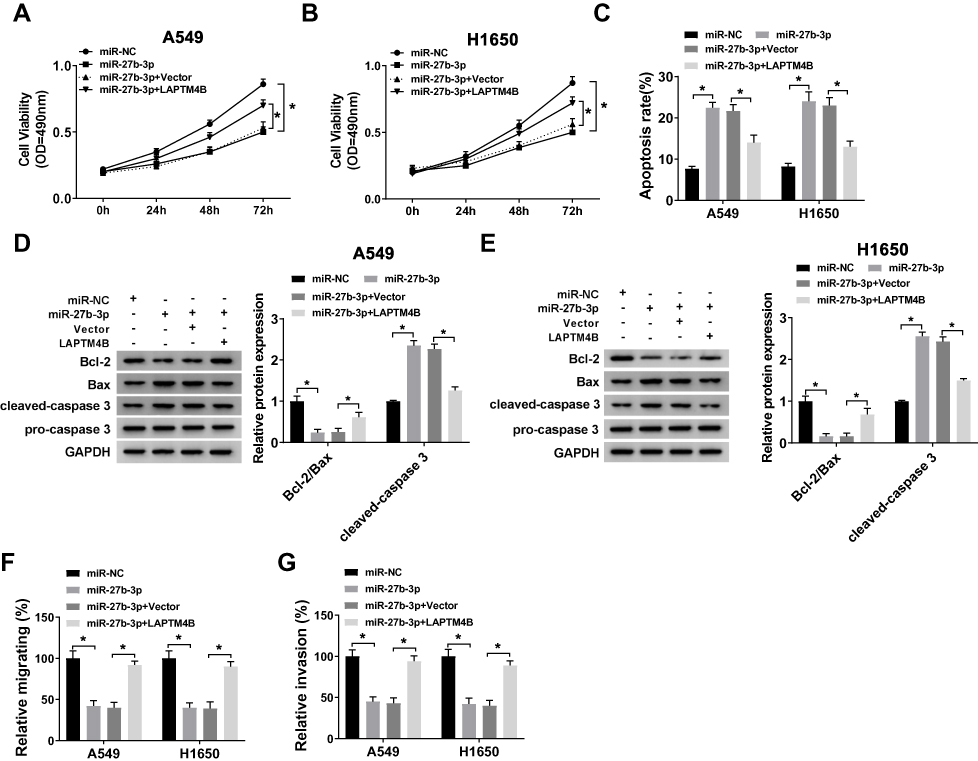 |
Figure 6 Overexpression of LAPTM4B regained the inhibition of miR-27b-3p on the growth and metastasis of lung cancer cells in vitro. (A, B) Cell viability was measured in miR-NC, miR-27b-3p transfected or miR-27b-3p + Vector or miR-27b-3p + LAPTM4B co-transfected A549 and H1650 cells by MTT. (C) Cell apoptosis rate was examined in A549 and H1650 cells by flow cytometry. (D, E) The protein expression levels of Bcl-2, Bax, cleaved-caspase3 and caspase3 were detected in miR-NC, miR-27b-3p transfected or miR-27b-3p + Vector or miR-27b-3p + LAPTM4B co-transfected A549 and H1650 cells using Western blot. (F, G) Transwell assays were performed to measure changes in migration and invasion of A549 and H1650 cells. *P<0.05. |
TP73-AS1 Enhanced LAPTM4B Expression by Sponging miR-27b-3p
In order to investigate whether TP73-AS1 could regulate the target gene LAPTM4B of miR-27b-3p in lung cancer, qRT-PCR and Western blot assays were performed. The results showed that the mRNA and protein expression of LAPTM4B were inhibited by miR-27b-3p and TP73-AS1 augmented LAPTM4B expression in A549 and H1650 cells (Figure 7A–D). These results revealed that TP73-AS1 could enhance the expression of LAPTM4B by binding miR-27b-3p.
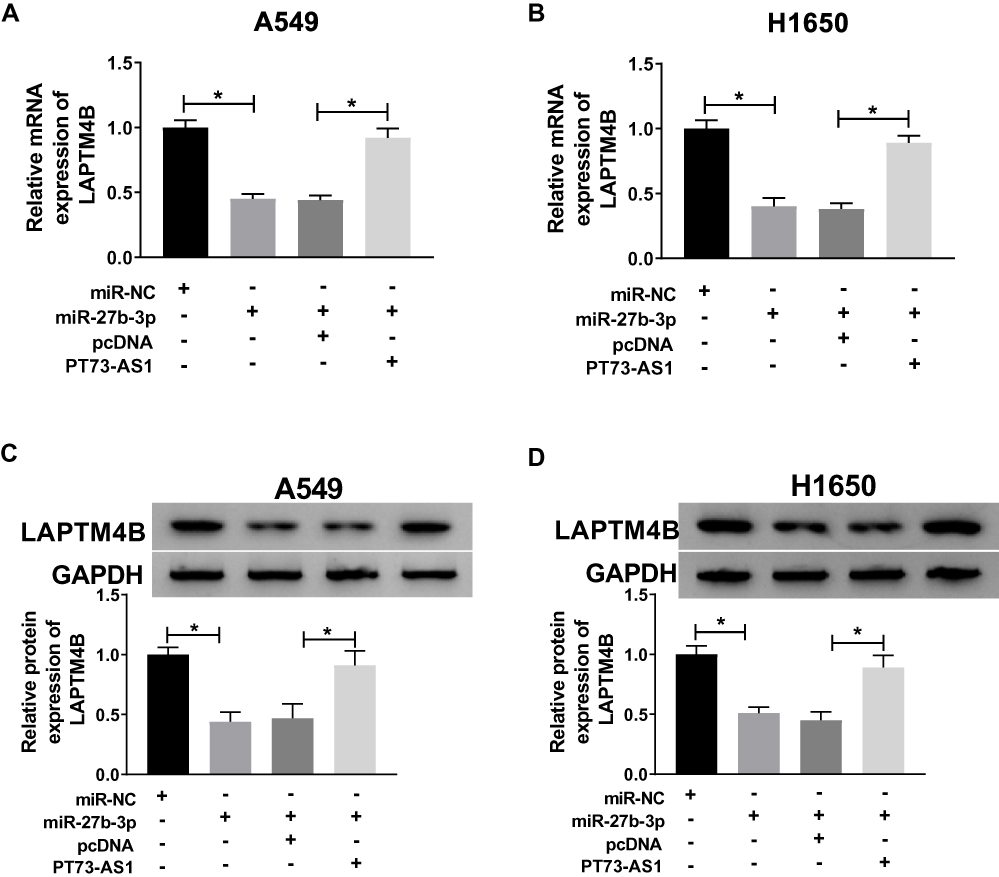 |
Figure 7 TP73-AS1 up-regulated the expression of LAPTM4B in lung cancer cells by regulating miR-27b-3p. (A, B) The mRNA and protein expression of LAPTM4B was measured in A549 and H1650 cells transfected with miR-NC, miR-27b-3p, miR-27b-3p + pcDNA or miR-27b-3p + TP73-AS1. (C, D) The protein level of LAPTM4B was detected in A549 and H1650 cells transfected with miR-NC, miR-27b-3p, miR-27b-3p + pcDNA or miR-27b-3p + TP73-AS1. *P<0.05. |
TP73-AS1 Knockdown Inhibited Tumor Growth in vivo
To further realize the significance of TP73-AS1 in tumor growth in vivo, TP73-AS1 silenced H1650 cells were subcutaneously introduced into the left flanks of nude mice. The results showed that the tumor growth rate was notably reduced after silencing TP73-AS1 (Figure 8A). The tumor weight in sh-TP73-AS1 group was also strikingly lower than that of tumors in sh-NC group (Figure 8B). Simultaneously, we found that TP73-AS1 was down-regulated in sh-TP73-AS1 group (Figure 8C), while miR-27b-3p was increased (Figure 8D), and the mRNA and protein levels of LAPTM4B were restrained (Figure 8E and F). The results supported that TP73-AS1 promoted tumor growth by up-regulating LAPTM4B and decreasing miR-27b-3p in vivo.
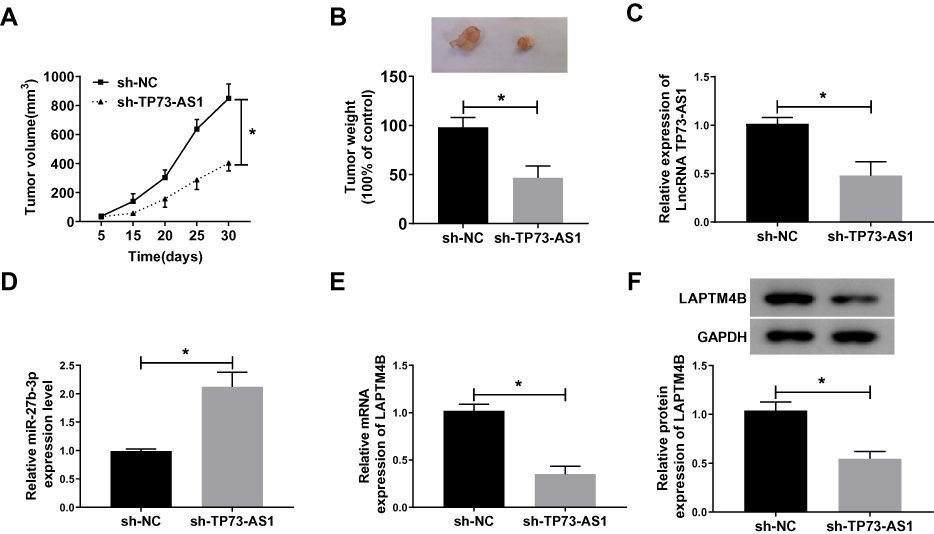 |
Figure 8 TP73-AS1 knockdown inhibited tumor growth in vivo. (A, B) H1650 cells stably transfected sh-TP73-AS1 or sh-NC were used for in vivo experiments. The volume and weight of tumors were measured. (C–E) The levels of TP73-AS1, miR-27b-3p and LAPTM4B in tumor tissues of mice were determined by qRT-PCR. (F) LAPTM4B protein expression was detected by Western blot. *P<0.05. |
Discussion
In recent years, lncRNAs have been validated as momentous cancer biomarkers, including lung cancer.23 Therefore, it is crucial to identify premonitory lncRNAs for lung cancer. In this study, TP73-AS1 was up-regulated, whereas miR-27b-3p was down-regulated in lung cancer cells. Also, LAPTM4B was predicted as a target of miR-27b-3p. So, we speculated that the TP73-AS1/miR-27b-3p/LAPTM4B network was answerable for the development of lung cancer.
TP73-AS1 was a new-found lncRNA, which has been proved to be an oncogene in cervical cancer,24 epithelial ovarian cancer25 and breast cancer.26 But it also has been reported as a tumor-inhibiting factor in bladder cancer.12 And our results showed that TP73-AS1 was exceptionally facilitated in lung cancer tissues and cells. Furthermore, we unveiled that silencing TP73-AS1 attenuated cell proliferation, migration, and invasion and augmented apoptosis in vitro. These data signified that TP73-AS1 served as an oncogene in lung cancer. TP73-AS1 was reported to be enormously aggrandized and predicted to have a low survival rate in NSCLC.27 Besides, in lung adenocarcinoma, TP73-AS1 could promote tumor growth and metastasis via the PI3K/AKT pathway.28 The above data suggested that our results were the same as previous reports and in line with expectations.
Most studies have shown that lncRNA function as a ceRNA sponging miRNA to affect the target gene of miRNA. For example, Shuang et al found that interference with TP73-AS1 hindered NSCLC cell progression and tumor growth by competitively sponging miR-449a to regulate EZH2 in vivo and in vitro.29 Here, the study proved that miR-27b-3p was notably down-regulated in lung cancer tissues and cells. Meanwhile, TP73-AS1 could negatively regulate the expression of miR-27b-3p and their expression was inversely correlated. It was worth mentioning that interference with miR-27b-3p could overturn these inhibitory effects of silencing TP73-AS1 on lung cancer cells. Dong et al also demonstrated that miR-27b-3p was aberrantly declined in NSCLC cells and counteracted the inhibitory impact of KCNQ1OT1 on cell growth and invasion.30 Given these facts, we believed that TP73-AS1 played a carcinogenic role in lung cancer through sponging miR-27b-3p.
Elevated level of LAPTM4B has been found to be associated with poor prognosis in several cancers, including lung cancer.31 In our study, LAPTM4B was directly by targeted miR-27b-3p in lung cancer cells. Similarly, LAPTM4B expression could also be negatively modulated by miR-27b-3p. A significant inverse interplay between miR-27b-3p and LAPTM4B was also observed. Then, we found that LAPTM4B expression was strikingly augmented in lung cancer cells. Functionally, LAPTM4B could attenuate the suppressive impact of miR-27b-3p on the growth and metastasis of lung cancer cells. Another important thing worth mentioning was that TP73-AS1 abolished the decreased expression of LAPTM4B mRNA and protein induced by miR-27b-3p. These data indicated that by binding miR-27b-3p, TP73-AS1 could enhance the expression of LAPTM4B at a post-transcriptional level. Moreover, TP73-AS1 depletion retarded tumor growth in vivo by regulating miR-27b-3p/LAPTM4B axis. Cumulative information suggested that LAPTM4B could promote the growth and metastasis of cells and motivate multidrug resistance in various kinds of tumors.32 Increasing research teams have chosen LAPTM4B as a target for anticarcinogen development.33 Thus, LAPTM4B might be co-expressed with TP73-AS1 in a multitude of lung cancer tissues and the interplay of TP73-AS1, miR-27b-3p and LAPTM4B might be an important network in regulating the progression of lung cancer.
Conclusion
Our study demonstrated that TP73-AS1 was overexpressed in lung cancer tissues and cell lines, and it promoted the progression of lung cancer in vivo and in vitro. TP73-AS1 exerted its carcinogenic effects in lung cancer through modulating the miR-27b-3p/LAPTM4B axis, which might supply new strategies for the precaution and therapeutic target of lung cancer.
Highlights
1. The targeting relationship between TP73-AS1 and miR-27b-3p was first verified.
2. The targeting relationship between miR-27b-3p and LAPTM4B was first verified.
3. TP73-AS1 promoted lung cancer cell progression via miR-27b-3p/LAPTM4B.
Ethics Approval and Consent to Participate
The present study was approved by the ethical review committee of The Affiliated Cancer Hospital of Zhengzhou University. Written informed consent was obtained from all enrolled patients.
Data Sharing Statement
The analyzed data sets generated during the present study are available from the corresponding author on reasonable request.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Torre LA, Freddie B, Siegel RL, Jacques F, Joannie LT, Ahmedin J. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
2. Reck M, Heigener DF, Mok T, Soria J-C, Rabe KF. Management of non-small-cell lung cancer: recent developments. Lancet. 2013;382(9893):709–719. doi:10.1016/S0140-6736(13)61502-0
3. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
4. Pan Y, Li C, Chen J, et al. The emerging roles of long noncoding RNA ROR (lincRNA-ROR) and its possible mechanisms in human cancers. Cell Physiol Biochem. 2016;40(1–2):219–229. doi:10.1159/000452539
5. Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013;154(1):26–46. doi:10.1016/j.cell.2013.06.020
6. Batista PJ, Chang HY. Long noncoding RNAs: cellular address codes in development and disease. Cell. 2013;152(6):1298–1307. doi:10.1016/j.cell.2013.02.012
7. Cazalla D, Yario T, Steitz JA. Down-regulation of a host microRNA by a Herpesvirus saimiri noncoding RNA. Science. 2010;328(5985):1563–1566. doi:10.1126/science.1187197
8. Yvonne T, John R, Pier Paolo P. The multilayered complexity of ceRNA crosstalk and competition. Nature. 2014;505(7483):344–352. doi:10.1038/nature12986
9. Zang W, Wang T, Wang Y, et al. Knockdown of long non-coding RNA TP73-AS1 inhibits cell proliferation and induces apoptosis in esophageal squamous cell carcinoma. Oncotarget. 2016;7(15):19960. doi:10.18632/oncotarget.v7i15
10. Zhang R, Jin H, Lou F. The long non-coding RNA TP73-AS1 interacted with miR-142 to modulate brain glioma growth through HMGB1/RAGE pathway. J Cell Biochem. 2017;119(4):3007–3016. doi:10.1002/jcb.26021
11. Zou Q, Zhou E, Xu F, Zhang D, Yi W, Yao J. A TP73-AS1/miR-200a/ZEB1 regulating loop promotes breast cancer cell invasion and migration. J Cell Biochem. 2017;119(2):2189–2199. doi:10.1002/jcb.26380
12. Tuo Z, Zhang J, Xue W. LncRNA TP73-AS1 predicts the prognosis of bladder cancer patients and functions as a suppressor for bladder cancer by EMT pathway. Biochem Biophys Res Commun. 2018;499:875–881. doi:10.1016/j.bbrc.2018.04.010
13. Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350. doi:10.1038/nature02871
14. Ameres SL, Zamore PD. Diversifying microRNA sequence and function. Nat Rev Mol Cell Biol. 2013;14(8):475–488. doi:10.1038/nrm3611
15. Qin X, Yu S, Xu X, Shen B, Feng J. Comparative analysis of microRNA expression profiles between A549, A549/DDP and their respective exosomes. Oncotarget. 2017;8(26):42125–42135. doi:10.18632/oncotarget.v8i26
16. Sun Y, Xu T, Cao YW, Ding XQ. Antitumor effect of miR-27b-3p on lung cancer cells via targeting Fzd7. Eur Rev Med Pharmacol Sci. 2017;21(18):4113.
17. Lianjin J, Oliver W, Marcusson EG, Cristina I, Calin GA, Alahari SK. Prooncogenic factors miR-23b and miR-27b are regulated by Her2/Neu, EGF, and TNF-α in breast cancer. Cancer Res. 2013;73(9):2884–2896. doi:10.1158/0008-5472.CAN-12-2162
18. Buffa FM, Carme C, Laura W, et al. microRNA-associated progression pathways and potential therapeutic targets identified by integrated mRNA and microRNA expression profiling in breast cancer. Cancer Res. 2011;71(17):5635–5645. doi:10.1158/0008-5472.CAN-11-0489
19. Li S, Wang L, Meng Y, Chang Y, Xu J, Zhang Q. Increased levels of LAPTM4B, VEGF and survivin are correlated with tumor progression and poor prognosis in breast cancer patients. Oncotarget. 2017;8(25):41282–41293. doi:10.18632/oncotarget.17176
20. Liu L, Xu X, Jing L, et al. Lysosomal-associated protein transmembrane 4 Beta-35 overexpression is a novel independent prognostic marker for gastric carcinoma. PLoS One. 1932;10(2):e0118026. doi:10.1371/journal.pone.0118026
21. Mingzhu Y, Cong L, Xia L, et al. Over-expression of LAPTM4B is associated with poor prognosis and chemotherapy resistance in stages III and IV epithelial ovarian cancer. J Surg Oncol. 2011;104(1):29–36. doi:10.1002/jso.21912
22. Yuho M, Junya F, Wenhua L, et al. LAPTM4B is associated with poor prognosis in NSCLC and promotes the NRF2-mediated stress response pathway in lung cancer cells. Sci Rep. 2015;5:13846. doi:10.1038/srep13846
23. Huarte M. The emerging role of lncRNAs in cancer. Nat Med. 2015;21(11):1253. doi:10.1038/nm.3981
24. Xu J, Zhang J. LncRNA TP73‐AS1 is a novel regulator in cervical cancer via miR‐329‐3p/ARF1 axis. J Cell Biochem. 2019;121(1):344–352.
25. Li Y, Jiao Y, Hao J, Xing H, Li C. Long noncoding RNA TP73-AS1 accelerates the epithelial ovarian cancer via epigenetically repressing p21. Am J Transl Res. 2019;11(4):2447.
26. Tao W, Sun W, Zhu H, Zhang J. Knockdown of long non-coding RNA TP73-AS1 suppresses triple negative breast cancer cell vasculogenic mimicry by targeting miR-490-3p/TWIST1 axis. Biochem Biophys Res Commun. 2018;504(4):629–634. doi:10.1016/j.bbrc.2018.08.122
27. Zhu D, Zhou J, Liu Y, Du L, Zheng Z, Qian X. LncRNA TP73-AS1 is upregulated in non-small cell lung cancer and predicts poor survival. Gene. 2019;710:98–102. doi:10.1016/j.gene.2019.05.044
28. Liu C, Ren L, Deng J, Wang S. LncRNA TP73-AS1 promoted the progression of lung adenocarcinoma via PI3K/AKT pathway. Biosci Rep. 2019;39(1):BSR20180999. doi:10.1042/BSR20180999
29. Shuang W, Zhu Y, Qiu R. Long noncoding RNA TP73-AS1 promotes non-small cell lung cancer progression by competitively sponging miR-449a/EZH2. Biomed Pharmacother. 2018;104:705–711. doi:10.1016/j.biopha.2018.05.089
30. Dong Z, Yang P, Qiu X, et al. KCNQ1OT1 facilitates progression of non‐small‐cell lung carcinoma via modulating miRNA‐27b‐3p/HSP90AA1 axis. J Cell Physiol. 2019;234(7):11304–11314. doi:10.1002/jcp.v234.7
31. Qiao M, Hu G. Lysosome-associated protein transmembrane-4β-35 is a novel independent prognostic factor in small cell lung cancer. Tumour Biol. 2015;36(10):7493–7499. doi:10.1007/s13277-015-3467-2
32. Meng Y, Wang L, Chen D, et al. LAPTM4B: an oncogene in various solid tumors and its functions. Oncogene. 2016;35(50):6359–6365. doi:10.1038/onc.2016.189
33. Zhang H, Wei Q, Liu R, et al. Overexpression of LAPTM4B-35: a novel marker of poor prognosis of prostate cancer. PLoS One. 2014;9(3):e91069. doi:10.1371/journal.pone.0091069
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.