Back to Journals » Cancer Management and Research » Volume 13
Long Non-Coding RNA (lncRNA) RAMS11 Promotes Metastatis and Cell Growth of Prostate Cancer by CBX4 Complex Binding to Top2α
Authors Zheng Z, Qiu K, Huang W
Received 11 September 2020
Accepted for publication 2 December 2020
Published 2 February 2021 Volume 2021:13 Pages 913—923
DOI https://doi.org/10.2147/CMAR.S270144
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ahmet Emre Eşkazan
Zhixiong Zheng, Kaiyan Qiu, Weiwen Huang
Urology Department, Quanzhou First Hospital Affiliated to Fujian Medical University, Quanzhou, Fujian 362000, People’s Republic of China
Correspondence: Weiwen Huang Email [email protected]
Introduction: Studies have confirmed that parts of the non-coding genes in the human genome play an important role in the pathogenesis and metastasis of prostate cancer. Among them, long non-coding RNAs (lncRNAs) are vitally involved in the biological regulation of prostate cancer. In addition, lncRNAs are closely associated with the recurrence, metastasis and prognosis of prostate cancer. However, the molecular pathogenesis of lncRNAs in regulating cell growth and metastasis of prostate cancer remains unclear. Therefore, this study was designed to explore the function and mechanism of lncRNA RAMS11 in cell growth and metastasis of prostate cancer.
Methods: Prostate cancer and para-carcinoma tissue samples were obtained from 42 patients who were diagnosed from March 2013 to September 2014 at Quanzhou First Hospital Affiliated to Fujian Medical University. Microarray experiments and real-time polymerase chain reaction (PCR) measured the expression of lncRNA. RWPE-2, LNCap, PC3 and DU145 cells were used for an in vitro model.
Results: The expression of lncRNA RAMS11 was up-regulated in prostate cancer tissue samples. LncRNA RAMS11 promoted cell growth and metastasis of prostate cancer cells. Down-regulation of lncRNA RAMS11 attenuated cell growth and metastasis of prostate cancer cells. We also demonstrated that lncRNA RAMS11 bound to CBX4 to activate expression of Top2α. LncRNA RAMS11 promoted tumor growth of prostate cancer in the mouse model. The inhibition of CBX4 attenuated the pro-cancer effects of lncRNA AMS11 in prostate cancer cells, while the activation of Top2α attenuated the anti-cancer effects of si-lncRNA RAMS11 in prostate cancer cells.
Discussion: Our results indicated that lncRNA RAMS11 promoted cell growth and metastasis of prostate cancer by CBX4 complex via binding to Top2α, and might be developed for the treatment of prostate cancer.
Keywords: lncRNA RAMS11, CBX4, Top2α, prostate cancer
Introduction
Prostate cancer is one of the common malignant tumors in the male urogenital system. According to the European Union Cancer Center, the mortality rate of prostate cancer is estimated to rank the third, with prostate cancer being the 8th cause of death due to cancer.1 Although the incidence of prostate cancer in China is lower than in European and American countries, it also clearly shows a rising trend in recent years.2 With the study of prostate cancer, the treatment of prostate cancer has also been changed to some extent.3,4
Chromobox (CBX) protein homolog protein is an important member of Polycomb group proteins (PcG).5 The PcG protein family is an epigenetic regulatory complex, existing in the form of polymeric transcriptional repression complex PRCs.6 It can inhibit the transcription of target genes by modifying chromatin, thereby playing an important role in regulating cell differentiation, aging, death, tumorigenesis and metastasis.7,8 The CBX protein family is an important member of the classic PRC1, including five members, namely CBX2, CBX4, CBX6, CBX7 and CBX8.9 Increasing evidence has shown that CBX protein plays an important role in tumorigenesis and tumor progression.10
Top2α plays an important role in maintaining the spatial structure of chromosomes during cellular DNA replication and transcription.11 Studies have shown that Top2α might be involved in regulating the apoptosis and proliferation of cancer cells, and may be associated with brain metastasis of non-small cell lung cancer.12,13 In addition, Top2α may also play an important role in the development of prostate cancer. Prostate cancer cells with higher expression of Top2α show stronger invasiveness and proliferation ability.14
LncRNAs are non-coding RNA molecules with a transcript length of over 200 nt, without protein-coding function.15 LncRNAs can exert biological functions through various mechanisms.16 Moreover, lncRNAs are closely associated with tumorigenesis, which can regulate gene expression at the epigenetic, transcriptional and post-transcriptional levels.17 At present, a variety of studies have shown that lncRNAs are widely involved in physiological and pathological processes, and also play important roles in the tumorigenesis and progression of malignant tumors.15–17 LncRNA RAMS11 promotes metastatic colorectal cancer progression. MicroRNAs (miRNAs) are noncoding RNAs with a length of about 22 nucleotides; miRNA plays a vital role in the tumorigenesis and metastasis of prostate cancer. Therefore, this study was designed to explore the function and mechanism of lncRNA RAMS11 in cell growth and metastasis of prostate cancer.
Materials and Methods
Clinical Samples
Prostate cancer and para-carcinoma tissue samples were obtained from 42 patients who were diagnosed from March 2013 to September 2014 at Quanzhou First Hospital Affiliated to Fujian Medical University. Tissues were immersed in liquid nitrogen and were stored at −80°C until use. Approval for this study was provided by the Ethics Committee of Quanzhou First Hospital Affiliated to Fujian Medical University. Written informed consent has been obtained from each subject, and all experiments conform to the Declaration of Helsinki.
Survival Analysis
Overall survival (OS) and disease-free survival (DFS) were performed using the Cox proportional hazard model with R survival package 2.37–7 2014. Kaplan–Meier curves were plotted using the R survplot package 0.0.7 2014.
Microarray Experiments and Real-Time Polymerase Chain Reaction (PCR)
Microarray experiments were performed at Genminix Informatics (China). Gene expression profiles were analyzed with the Human Exon 1.0 ST GeneChip (Affymetrix).
Total RNA was isolated using Takara Bio NucleoSpin RNA (Takara, Mountain View, CA). cDNA was then transcribed using SuperScript III First strand cDNA system (Invitrogen). PCR was executed using Fast Sybr Green Master Mix (Invitrogen).
Cell Culture
RWPE-2, LNCap, PC3 and DU145 cells were obtained from Cell Bank (Shanghai Genechem Co., Ltd., Shanghai, China). Cell was cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco) supplemented with 10% fetal bovine serum (FBS, Gibco) in an incubator at 37°C with 5% CO2. Cell was transfected using Lipofectamine 2000 (Invitrogen, USA) with correlation RAMS11 plasmid (5′-ggcgcgagtttcaggcagcgctgcgtc-3′ and 5′-agtttttcagttttgaaaaaaa-3′), CBX4 plasmid (5′-ccacccagggaacggcaggagcagctgatg-3′ and 5′-cgtcaccgcgaactgcctcaccgttacttt-3′), Top2α plasmid (5′-catggaagtgtcaccattgc-3′ and 5′-agaccttgaagctgatgatg-3′), si-RAMS11 mimics (5′-tggagaccctgagaaggacc-3′), si-CBX4 mimics (sc-270,615, Santa Cruz Biotechnology), or si-Top2α mimics (5′-ttttgtaatcattgtcaagtgcaaaa-3′).
MTT Assays and Cytotoxicity
Cell viability was analyzed using MTT assays (Beyotime) at 0, 24, 48 and 72 h after transfection. 20 μL of MTT solution was added to each well and cells were incubated for 4 h at 37°C. Supernatants were removed and formazan crystals were dissolved in 150 μL dimethylsulfoxide (DMSO). Absorbance was measured using a Multiskan Spectrum Microplate Spectrophotometer (Thermo Scientific™, USA) at a wavelength of 492 nm.
Cytotoxicity (LDH activity) levels were measured using LDH activity kits (Beyotime). Absorbance was measured using a Multiskan Spectrum Microplate Spectrophotometer (Thermo Scientific™, USA) at a wavelength of 450 nm.
Transwell Assays
Cell migration assays was performed using a 24-well Transwell (Corning, New York, NY, USA) with a polycarbonate pre-coated filter (BD Biosciences, Franklin Lakes, NJ, USA). Cells (1 x 104 cell) after transfection at 6 h were added to the upper well of the chamber, and 500 μL of DMEM supplemented with 10% FBS was added to the lower well. The upper well of cells was removed using scraping after 48 h incubation. Invaded cells of the membrane were fixed with 4% paraformaldehyde for 15 min and stained with 0.1% crystal violet solution (BD Biosciences, Franklin Lakes, NJ, USA). Cell numbers were counted using Image-Pro Plus 6.0 software (Media Cybernetics, Silver Springs, MD, USA).
Caspase-3/9 Assay
Total protein was extracted from the cells using RIPA buffer containing protease inhibitors. Proteins (10 µg) were used to measure Caspase-3/Caspase-9 activity levels using Caspase-3/Caspase-9 activity level kits. The absorbance was measured by an EnSpire multimode plate reader (PerkinElmer, Inc., Waltham, MA, USA) at a wavelength of 405 nm. Proteins (10 µg) were used to measure Caspase-3/Caspase-9 activity levels using Caspase-3/Caspase-9 activity level kits. The absorbance was measured by an EnSpire multimode plate reader (PerkinElmer, Inc., Waltham, MA, USA) at a wavelength of 405 nm.
RNA Immunoprecipitation
Isolating nuclear lysates were collected using NERPER Nuclear and Cytoplasmic Extraction Reagent Kit (Thermo Fisher). Nuclear lysates were incubated with 5 µg CBX4 antibody or IgG antibody isotype control in RIPA wash buffer overnight at 4°C. 50 µL of Invitrogen Dynabeads Protein G were rotated for 1 h at 4°C. Beads were subsequently washed six times with RIPA and protein was then digested with RIPA at 55° for 30 min with shaking. The primers of Top2α promoter were 5′-catggaagtgtcaccattgc-3′ and 5′-ttaaaatgtgaggcgattat-3′. gDNA was removed from RNA using ArcticZymes Heat and Run gDNA removal kit. cDNA was then transcribed using SuperScript III First strand cDNA system (Invitrogen). PCR was executed using Fast Sybr Green Master Mix (Invitrogen).
Chromatin Immunoprecipitation (ChIP)-qPCR
ChIP-qPCR was conducted by first sonicating 5 million cells in 1% SDS lysis buffer. Sonicated cells were incubated with 5 µg IgG or CBX4 antibodies in ChIP-qPCR dilution buffer overnight with rotation at 4°C. Dynabeads Protein G (Invitrogen) was added and rotated for 1 h. Mixture was then washed once with low salt wash buffer, then high salt buffer, lithium chloride wash buffer, and finally two washes with Tris-HCl EDTA buffer. DNA was eluted by incubating beads for 30 min at room temperature with SDS elution buffer, at 95°C for 15 min with shaking, and then at 60°C for 15 min with shaking. DNA was then isolated and cDNA was then transcribed using SuperScript III First strand cDNA system (Invitrogen). PCR was executed using Fast Sybr Green Master Mix (Invitrogen).
Western Blot Analysis
Total proteins of cell or tissue samples were extracted using RIPA lysis buffer and protease inhibitor cocktail (1:100, Beyotime). The protein concentration was determined using BCA Kit (Beyotime). 50 μg protein samples were loaded to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred onto polyvinyl difluoride (PVDF, Thermo Scientific™, USA) membranes. Membranes were incubated with CBX4, Top2α and GAPDH at 4°C overnight after blocking with 5% non-fat milk in Tris-buffered saline with 0.1% Tween 20 (TBST). The membrane was washed with TBST and then incubated with anti-rabbit secondary antibody (1:5000) for 2 h at room temperature. Immunoreactive bands were visualized using the ECL kit (Thermo Scientific™, USA) and integrated density of the bands was quantified by Quantity One software (Bio-Rad).
Statistical Analysis
Data were expressed as the mean ± standard deviation (SD). Comparisons between groups of independent samples were assessed by Student’s t-test or one-way analysis of variance (ANOVA) and Tukey’s post test. p values < 0.05 were considered to indicate statistical significance.
Results
LncRNA RAMS11 Expression in Prostate Cancer
To confirm the changes of lncRNA RAMS11 expression in prostate cancer, microarray assay was executed. LncRNA RAMS11 expression was found to be up-regulated in cancer tissue samples, compared with para-carcinoma tissue samples (Figure 1A and B). We also found that the expression of lncRNA RAMS11 was up-regulated in LNCap, PC3 and DU145 cells (prostate cancer cell lines), compared with RWPE-2 cells (normal prostatic cell, Figure 1C). In addition, OS and DFS of patients with high RAMS11 expression were poorer than those with low lncRNA RAMS11 expression (Figure 1D and E). These results showed that lncRNA RAMS11 regulated prostate cancer cell growth.
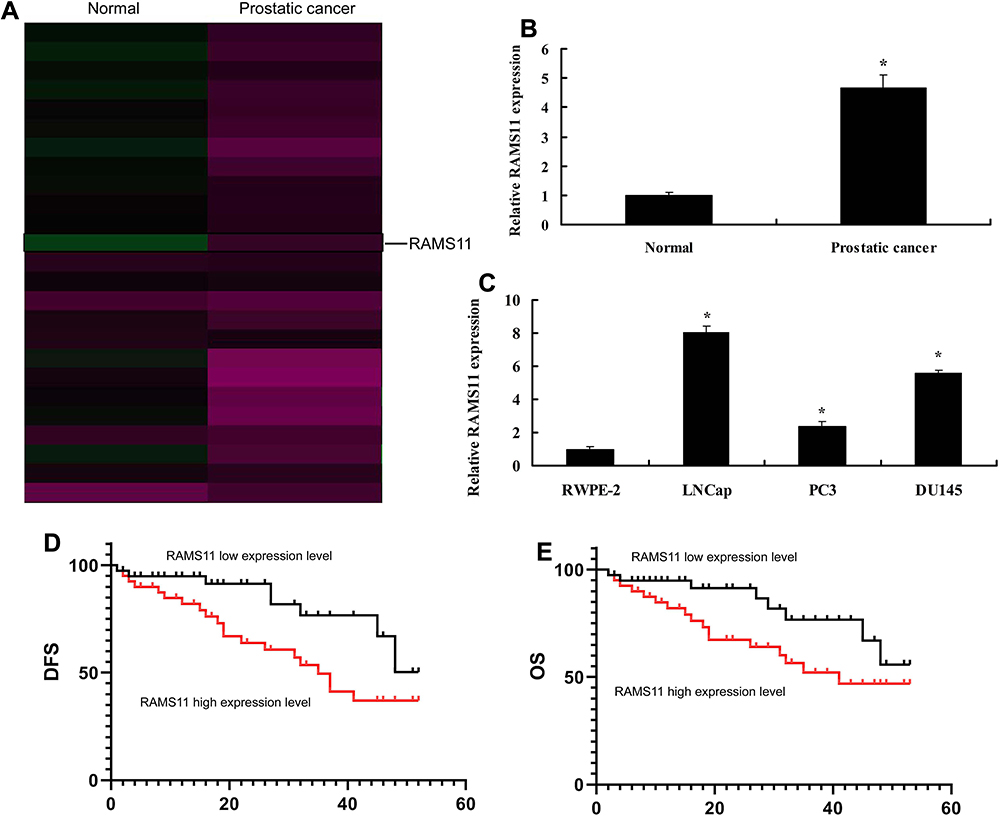 |
Figure 1 LncRNA RAMS11 expression in prostate cancer. Heat map of gene chip (A), lncRNAs RAMS11 expression in cancer tissue samples (B), lncRNA RAMS11 expression in LNCap, PC3 and DU145 cells (C), DFS and OS of lncRNA RAMS11 expression in prostate cancer (D, E). *p<0.01 compared with normal group. Abbreviations: Normal, normal group; prostate cancer, prostate cancer patients. |
LncRNA RAMS11 Promoted Cell Growth and Metastasis of Prostate Cancer Cells
To determine the pro-cancer effects of lncRNA RAMS11 in human prostate cancer, cell growth and cell migration assay were measured. As shown in Figure 2, lncRNA RAMS11 expression was up-regulated, and over-expression of lncRNA RAMS11 promoted cell viability, migration and transfer rate, and decreased LDH activity, caspase-3/9 activity levels and apoptosis rate of prostate cancer cells (PC3 cell). However, we found that the expression of lncRNA RAMS11 was down-regulated. Down-regulation of lncRNA RAMS11 reduced cell viability, migration and transfer rate, and induced LDH activity, caspase-3/9 activity levels and apoptosis rate of liver cancer cells, compared with negative control group (Figure 3).
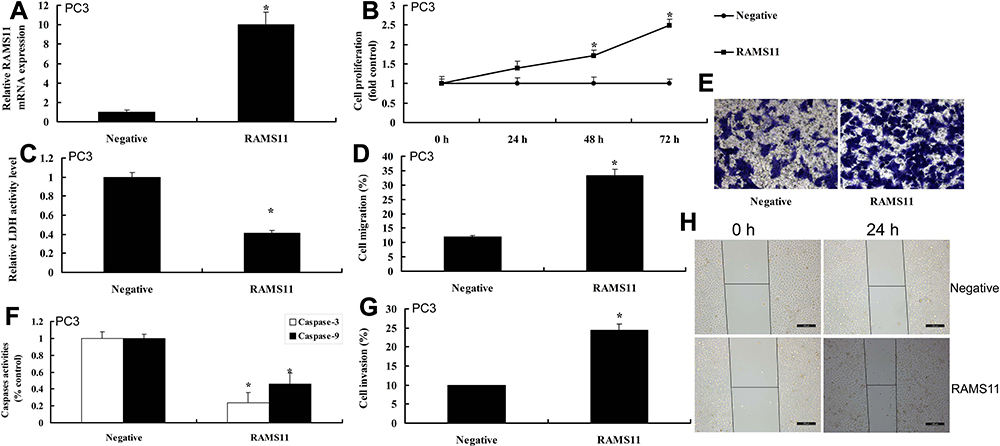 |
Figure 2 LncRNA RAMS11 promotes metastatic and cell growth of prostate cancer cells. LncRNA RAMS11 expression (A), cell proliferation (B), LDH activity cytotoxicity (C), migration rate (D, E), caspase-3/9 activity levels (F), invasion rate (G, H) in vitro model of PC3 cell. *p<0.01 compared with negative group. Abbreviations: Negative, negative mimics group; RAMS11, over-expression of RAMS11 group. |
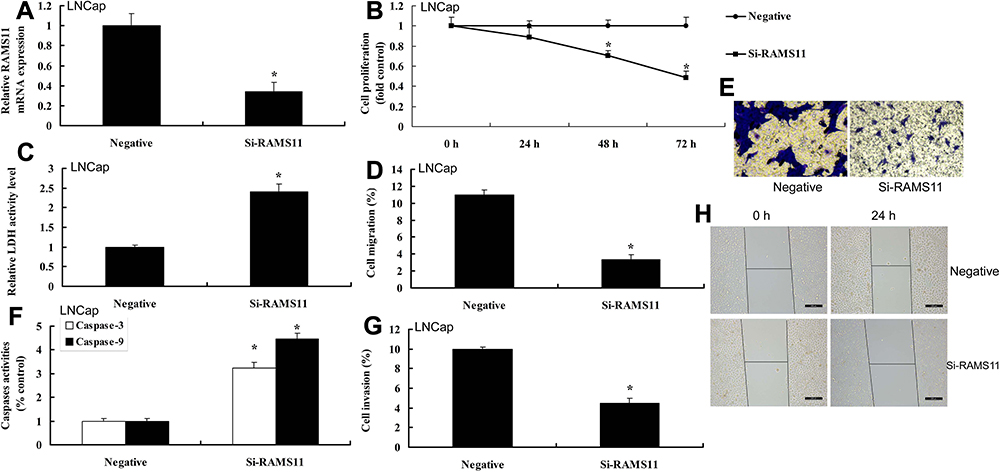 |
Figure 3 Si-LncRNA RAMS11 reduced metastatic and cell growth of prostate cancer cell. LncRNA RAMS11 expression (A), cell proliferation (B), LDH activity cytotoxicity (C), migration rate (D, E), caspase-3/9 activity levels (F), invasion rate (G, H) in vitro model of LNCap cells. *p<0.01 compared with negative group. Abbreviations: Negative, negative mimics group; Si-RAMS11, down-regulation of RAMS11 group. |
LncRNA RAMS11 Bound to CBX4 to Activate the Expression of Top2α
To confirm the mechanism of lncRNA RAMS11 in prostate cancer, microarray assay was performed to analyze the regulatory role of lncRNA RAMS11 in gene expression (Figure 4A). Volcano plot showed that the expression of CBX4 and Top2α were up-regulated by lncRNA RAMS11 (Figure 4B). Network plot showed that CBX4 and Top2α regulated passage (Figure 4C). As shown in Figure 4D, lncRNA RAMS11 bound to CBX4 to activate the expression of Top2α. Over-expression of lncRNA RAMS11 induced the protein expression of CBX4 and Top2α in vitro and down-regulation of lncRNA RAMS11 suppressed the protein expression of CBX4 and Top2α in vitro (Figure 4E and J).
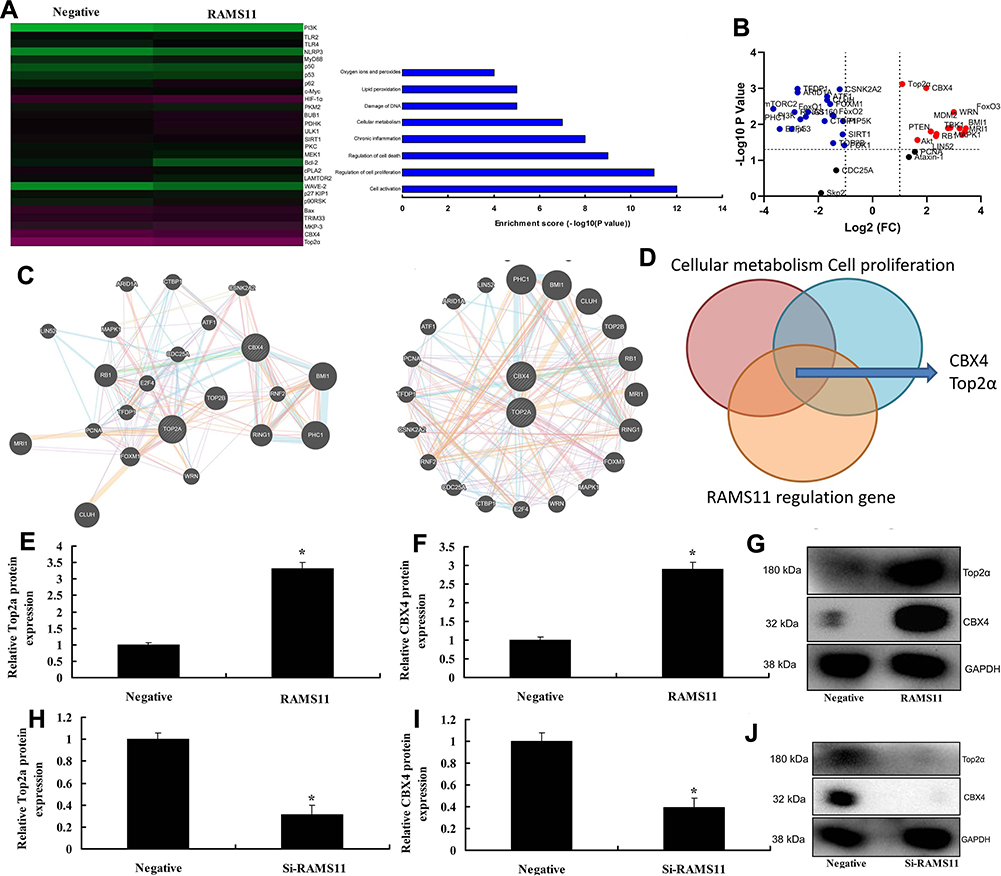 |
Figure 4 LncRNA RAMS11 regulated CBX4 and Top2α expression. Heat map and results map of gene chip (A), volcano figure (B), Network figures (C, D), analyze results map (E), CBX4 and Top2α protein expressions by over-expression of lncRNA RAMS11 (E–G); CBX4 and Top2α protein expression by down-regulation of lncRNA RAMS11 (H–J). *p<0.01 compared with negative group. Abbreviations: Negative, negative mimics group; RAMS11, over-expression of RAMS11 group; Si-RAMS11, down-regulation of RAMS11 group. |
RNA immunoprecipitation (RIP) showed that enrichment levels of lncRNA RAMS11 binding to CBX4 in PC3 and DU145 cells were higher than those of lncRNA RAMS11 binding to lgG (Figure 5A). Chip assay showed that Top2α promoter levels of lncRNA RAMS11 binding to CBX4 or si-CBX4 binding to CBX4 were lower than those of WT binding to CBX4 or WT binding to CBX4 in LNCap or DU145 cells (Figure 5B and C). The promoter levels of Top2α of RAMS11 binding to CBX4 or CBX4 binding to CBX4 were higher than those of WT binding to CBX4 or WT binding to CBX4 in PC3 or DU145 cells (Figure 5D and E). Top2α promoter levels of lncRNA RAMS11+CBX4 binding to CBX4 in PC3 and DU145 cells were higher than those of si-lncRNA RAMS11 binding to CBX4 (Figure 5F and G). Additionally, Top2α promoter levels of si-lncRNA RAMS11+si-CBX4 binding to CBX4 in PC3 and DU145 cells were lower than those of RAMS11 binding to CBX4 (Figure 5H and I). Meanwhile, Top2α promoter levels of si-lncRNA RAMS11+si-CBX4 binding to CBX4 were lower than those of si-lncRNA RAMS11 binding to CBX4 (Figure 5J and K). Top2α promoter levels of si-lncRNAs RAMS11+CBX4 binding to CBX4 were higher than those of si-lncRNA RAMS11 binding to CBX4 (Figure 5L and M). These results showed that lncRNA RAMS11 bound to CBX4 to activate the expression of Top2α.
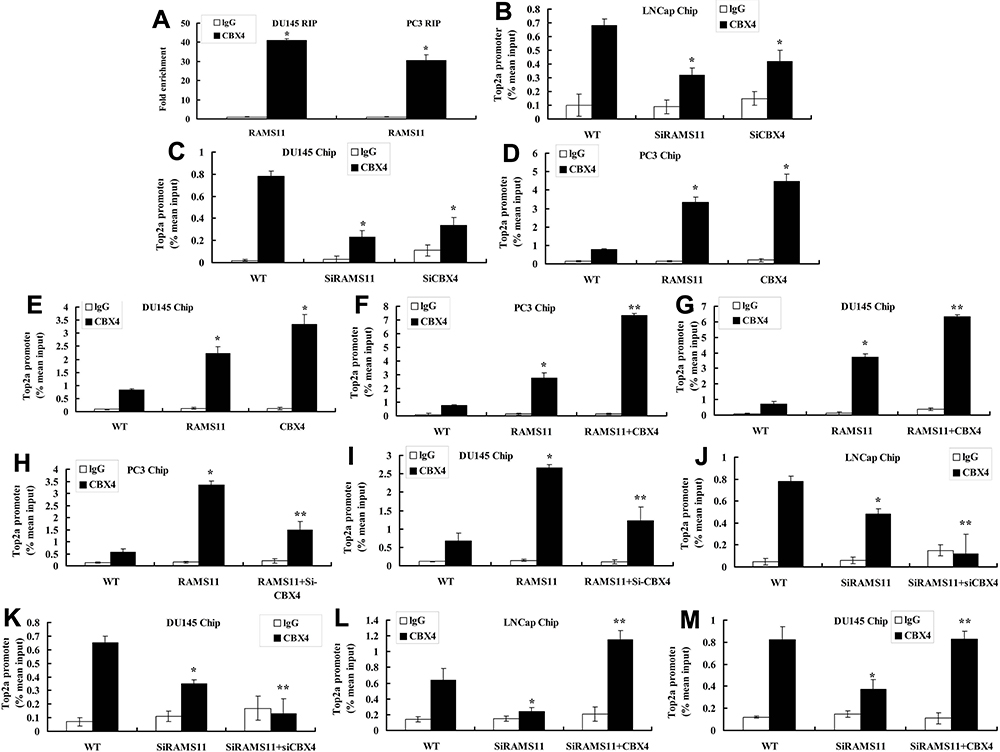 |
Figure 5 LncRNA RAMS11 binds to CBX4 to activate expression of Top2α. LncRNA RAMS11 binds to CBX4 in PC3 and DU145 cells (RIP, A), Top2α promoter levels of lncRNA RAMS11 binds to CBX4 or si-CBX4 binds to CBX4 (Chip, B, C), Top2α promoter levels of RAMS11 binds to CBX4 or CBX4 binds to CBX4 (Chip, D, E), Top2α promoter levels of lncRNA RAMS11+CBX4 binds to CBX4 in PC3 and DU145 cells (Chip, F, G), Top2α promoter levels of si-lncRNA RAMS11+si-CBX4 binds to CBX4 in PC3 and DU145 cells (Chip, H, I), Top2α promoter levels of si-lncRNA RAMS11+si-CBX4 binds to CBX4 (Chip, J, K), Top2α promoter levels of si-lncRNA RAMS11+CBX4 binds to CBX4 (Chip, L, M). *p<0.01 compared with negative group; **p<0.01 compared with over-expression of RAMS11 group or down-regulation of RAMS11 group. Abbreviations: WT, WT negative group; RAMS11, over-expression of RAMS11 group; SiRAMS11, down-regulation of RAMS11 group; CBX4, over-expression of CBX4 group; SiCBX4, down-regulation of CBX4 group. |
LncRNA RAMS11 Promoted Tumor Growth of Prostate Cancer in Mouse Model
We further found that over-expression of lncRNA RAMS11 promoted tumor growth, and decreased caspase-3/9 activity levels in mice with prostate cancer (Figure 6A and C). The gene expression of lncRNA RAMS11 was increased, and the protein expression of CBX4/Top2α was also enhanced in prostate cancer mice with over-expression of lncRNA RAMS11 prostate cancer (Figure 6D and G). Meanwhile, in prostate cancer tissue samples, the protein expression of CBX4 was up-regulated in prostate cancer mice with over-expression of lncRNA RAMS11 prostate cancer (Figure 6H).
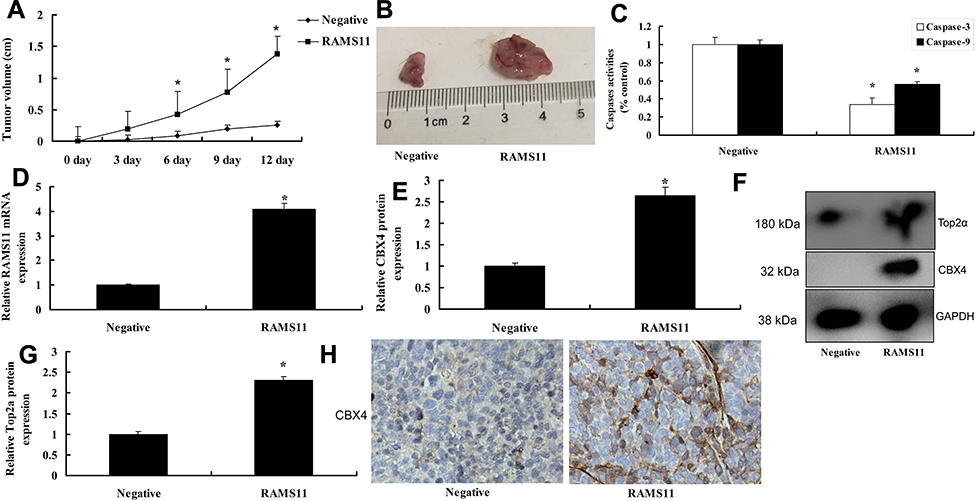 |
Figure 6 LncRNA RAMS11 promotes tumor growth of prostate cancer in mice model. Tumor growth (A, B), caspase-3/9 activity levels (C), LncRNA RAMS11 mRNA expression (D), CBX4 and Top2α protein expressions (E–G), CBX4 protein expression (F, H). *p<0.01 compared with negative group. Abbreviations: Negative, negative mimics group; RAMS11, over-expression of RAMS11 group. |
The Inhibition of CBX4 Attenuated the Pro-Cancer Effects of lncRNA AMS11 in Prostate Cancer Cells
Si-CBX4 suppressed the protein expression of CBX4 and Top2α in vitro by over-expression of lncRNA RAMS11, compared with over-expression of lncRNA RAMS11 group (Figure 7A and C). The inhibition of CBX4 attenuated the pro-cancer effects of lncRNA RAMS11 on cell viability, migration and transfer rate, and decreased LDH activity, caspase-3/9 activity levels and apoptosis rate of prostate cancer cells (PC3 cells) (Figure 7D and J).
 |
Figure 7 The inhibition of CBX4 reduced the pro-cancer effects of lncRNA RAMS11 in prostate cancer cells. CBX4 and Top2α protein expressions (A–C), cell proliferation (D), LDH activity cytotoxicity (E), caspase-3/9 activity levels (F), migration rate (G, H), invasion rate (I, J) in vitro model of PC3 cell. *p<0.01 compared with negative group, #p<0.01 compared with over-expression of RAMS11 group. Abbreviations: Negative, negative mimics group; RAMS11, over-expression of RAMS11 group; SiCBX4, down-regulation of CBX4 group. |
The Activation of Top2α Attenuated the Anti-Cancer Effects of si-lncRNA RAMS11 in Prostate Cancer Cells
Top2α plasmid induced the protein expression of Top2α in the in vitro model of prostate cancer cells by down-regulation of lncRNA RAMS11 (Figure 8A and B). The activation of Top2α attenuated the anti-cancer effects of si-lncRNA RAMS11 on cell viability, migration and transfer rate, and decreased LDH activity, caspase-3/9 activity levels and apoptosis rate of prostate cancer cells (PC3 cell) (Figure 8C and I).
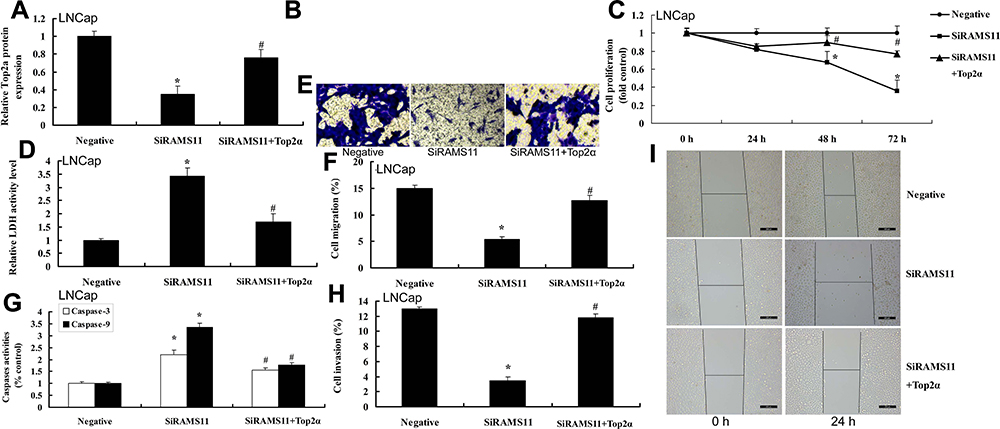 |
Figure 8 The activation of Top2α reduced the anti-cancer effects of si-lncRNA RAMS11 in prostate cancer cells. Top2α protein expressions (A, B), cell proliferation (C), LDH activity cytotoxicity (D), migration rate (E, F), caspase-3/9 activity levels (G), invasion rate (H, I) in vitro model of LNCap cell. *p<0.01 compared with negative group, #p<0.01 compared with down-regulation of RAMS11 group. Abbreviations: Negative, negative mimics group; SiRAMS11, down-regulation of RAMS11 group; Top2α, over-expression of Top2α group. |
Discussion
The treatment of prostate cancer is mainly determined by objective factors, including patient age, physical condition, clinical stage, Gleason score, presence or absence of pelvic lymph nodes, distant metastases, and life expectancy.18 At present, there are various therapeutic methods for prostate cancer, each with advantages and disadvantages.19 Comprehensive consideration should be given towards patients with early-stage prostate cancer according to the patient’s condition. While patients with advanced prostate cancer still cannot be cured, the treatment should aim to relieve clinical symptoms and to prolong survival.20 With the development of molecular biology and immunology, novel therapeutic approaches, such as gene therapy and immunotherapy, will bring more benefits to patients with prostate cancer. MiRNAs play important regulatory roles in tumor development and metastasis.21 In the current study, RAMS11 expression in cancer tissue samples was up-regulated, RAMS11 expression in LNCap, PC3 and DU145 cells. OS and DFS of RAMS11 high expression were lower than those of RAMS11 low expression. Silva-Fisher et al. showed that lncRNA RAMS11 promotes metastatic colorectal cancer progression.22 Therefore, we thought lncRNA RAMS11 expression in cancer tissue samples was up-regulated, and OS and DFS of RAMS11 high expression were lower than those of lncRNA RAMS11 low expression.
In recent years, a large number of studies have shown that lncRNAs are closely associated with prostate cancer, playing an important role in tumorigenesis and progression of tumors.15–17 Our study showed that RAMS11 promotes metastatic and cell growth of prostate cancer cells. Silva-Fisher et al. showed that lncRNA RAMS11 promotes metastatic colorectal cancer progression by CBX4/TOP2α.22 Our results showed that lncRNA RAMS11 promotes metastatis and cell growth of prostate cancer.
The role of CBX4 in tumors is correlated with its unique domains.9 It is the only protein that has enzymatic activity and can act as a SUMO E3 ligase to participate in SUMO modification of the family.23 Meanwhile, abnormal sumoylation has been shown to be related to multiple types of diseases, including neurodegenerative diseases, heart failure, and malignant tumors. CBX4 regulates various important proteins via the action of its SUMO E3 enzyme.24 In recent years, the expression and mechanism of action of CBX4 in malignant tumors has gradually become a research hotspot.7 We also show that RAMS11 binds to CBX4 to activate expression of Top2α.
Top2α plays an important role in many biological processes, such as DNA replication, transcription, repair, and structural maintenance and reconstruction of chromosomes.11,25 in vitro assays have shown that Top2α is generally highly expressed during cell replication, which can be used as one of the proliferative markers.12,13 Our study shows that the inhibition of CBX4 reduced the pro-cancer effects of lncRNA RAMS11 in prostate cancer cells and the activation of Top2α reduced the anti-effects of si-lncRNAs RAMS11 in prostate cancer cells. Silva-Fisher et al. showed that lncRNA RAMS11 promotes metastatic colorectal cancer progression by CBX4/TOP2α.22
Overall, our understanding of how lncRNAs RAMS11 promote metastasis and cell growth of prostate cancer may have tremendous biological and clinical significance. Next, our study found that lncRNAs RAMS11 expression in prostate cancer tissue samples was up-regulated, and lncRNAs RAMS11 promotes metastatis and cell growth of prostate cancer by CBX4 complex binding to Top2α (Figure 9). In addition, the novel role of lncRNA RAMS11 may help lead to the treatment of prostate cancer or to development of novel diagnostic tools for assessing prostate cancer progression.
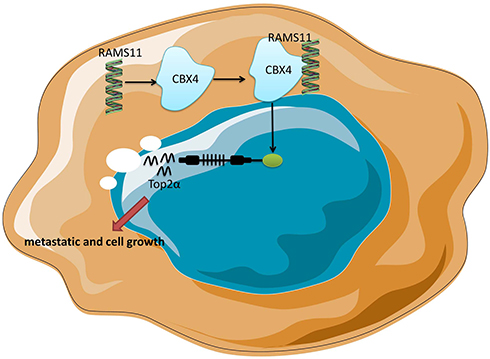 |
Figure 9 Long non-coding RNA (lncRNA) RAMS11 promotes metastatic and cell growth of prostate cancer by CBX4 complex binding to Top2α. |
Funding
There is no funding to report.
Disclosure
The authors declare that they have no conflicts of interest for this work.
References
1. Jin W, Fei X, Wang X, Chen F, Song Y. Circulating miRNAs as biomarkers for prostate cancer diagnosis in subjects with benign prostatic hyperplasia. J Immunol Res. 2020;2020:5873056. doi:10.1155/2020/5873056
2. Ranasinghe WKB. Patterns of metastases of prostatic ductal adenocarcinoma. Mediators Inflamm. 2020.
3. Fredsøe J, Kirkegaard P, Edwards A, Vedsted P, Sørensen KD, Bro F. A genetic risk assessment for prostate cancer influences patients’ risk perception and use of repeat PSA testing: a cross-sectional study in Danish general practice. BJGP Open. 2020;4(2):bjgpopen20X101039. doi:10.3399/bjgpopen20X101039
4. Jin W. Detection and prognosis of prostate cancer using blood-based biomarkers. J Immunol Res. 2020;2020:8730608.
5. Mahira S, Kommineni N, Husain GM, Khan W. Cabazitaxel and silibinin co-encapsulated cationic liposomes for CD44 targeted delivery: a new insight into nanomedicine based combinational chemotherapy for prostate cancer. Biomed Pharmacother. 2019;110:803–817. doi:10.1016/j.biopha.2018.11.145
6. Pessoa RR, Viana PC, Mattedi RL, et al. Value of 3-tesla multiparametric magnetic resonance imaging and targeted biopsy for improved risk stratification in patients considered for active surveillance. BJU Int. 2017;119:535–542. doi:10.1111/bju.13624
7. Jiang N, Niu G, Pan YH, et al. CBX4 transcriptionally suppresses KLF6 via interaction with HDAC1 to exert oncogenic activities in clear cell renal cell carcinoma. EBioMedicine. 2020;53:102692. doi:10.1016/j.ebiom.2020.102692
8. Pu Z, Che Y, Zhang W, et al. Dual roles of IL-18 in colitis through regulation of the function and quantity of goblet cells. Int J Mol Med. 2019;43:2291–2302. doi:10.3892/ijmm.2019.4156
9. Milosevich N, McFarlane J, Gignac MC, et al. Pan-specific and partially selective dye-labeled peptidic inhibitors of the polycomb paralog proteins. Bioorg Med Chem. 2020;28(1):115176. doi:10.1016/j.bmc.2019.115176
10. Wang S, Denton KE, Hobbs KF, Weaver T, McFarlane JMB. Optimization of ligands using focused DNA-encoded libraries to develop a selective, cell-permeable CBX8 chromodomain inhibitor. ACS Chem Biol. 2020;15:112–131. doi:10.1021/acschembio.9b00654
11. Fang Y, Yang C, Zhang L, Wei L, Lin J. Spica prunellae extract enhances fluorouracil sensitivity of 5-fluorouracil-resistant human colon carcinoma HCT-8/5-FU cells via TOP2α and miR-494. Biomed Res Int. 2019;2019:5953619. doi:10.1155/2019/5953619
12. Kanagasabai R, Karmahapatra S, Kientz CA, et al. The novel C-terminal truncated 90-kDa isoform of topoisomerase IIα (TOP2α/90) is a determinant of etoposide resistance in K562 leukemia cells via heterodimerization with the TOP2α/170 isoform. Mol Pharmacol. 2018;93:515–525. doi:10.1124/mol.117.111567
13. Zhao W, Jiang G, Bi C, et al. The dual topoisomerase inhibitor A35 preferentially and specially targets topoisomerase 2α by enhancing pre-strand and post-strand cleavage and inhibiting DNA religation. Oncotarget. 2015;6:37871–37894. doi:10.18632/oncotarget.5680
14. Duzcan F, Duzcan SE, Sen S, et al. Expression and amplification of topoisomerase-2α in type 1 and type 2 papillary renal cell carcinomas and its correlation with HER2/neu amplification. Pathol Oncol Res. 2011;17:697–703. doi:10.1007/s12253-011-9372-0
15. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
16. Castro-Oropeza R, Melendez-Zajgla J, Maldonado V, Vazquez-Santillan K. The emerging role of lncRNAs in the regulation of cancer stem cells. Cell Oncol (Dordr). 2018;41:585–603. doi:10.1007/s13402-018-0406-4
17. Chang YN, Zhang K, Hu ZM, et al. Hypoxia-regulated lncRNAs in cancer. Cell Oncol (Dordr). 2016;575:1–8.
18. Paparo F, Peirano A, Matos J, et al. Diagnostic value of retrospectively fused (64)CuCl(2) PET/MRI in biochemical relapse of prostate cancer: comparison with fused (18)F-Choline PET/MRI, (64)CuCl2 PET/CT, (18)F-Choline PET/CT, and mpMRI. Abdom Radiol (NY). 2020;45(11):3896–3906. doi:10.1007/s00261-020-02591-7
19. van Leenders G, van der Kwast TH, Grignon DJ, et al., ISUP Grading Workshop Panel Members. The 2019 International Society of Urological Pathology (ISUP) consensus conference on grading of prostatic carcinoma. Am J Surg Pathol. 2018;44(8):e87–e99. doi:10.1097/PAS.0000000000001497
20. Wu C, Jiang H, Chen J. A systematic review and meta-analysis about the effect of bisphosphonates on the risk of skeletal-related event in men with prostate cancer. Anticancer Agents Med Chem. 2020;20(13):1604–1612. doi:10.2174/1871520620666200521114815
21. Salles DC, Mata DA, Epstein JI. Significance of paneth-cell-like differentiation in prostatic adenocarcinoma: a retrospective cohort study of 80 cases. Hum Pathol. 2020;
22. Silva-Fisher JM, Dang HX, White NM, et al. Long non-coding RNA RAMS11 promotes metastatic colorectal cancer progression. Nat Commun. 2020;11:2156. doi:10.1038/s41467-020-15547-8
23. Yerokun T, Winfield LL. LLW-3-6 and celecoxib impacts growth in prostate cancer cells and subcellular localization of COX-2. Anticancer Res. 2014;34:4755–4759.
24. Tan C, Bei C, Zhu X, Zhang Y, Qin L, Tan S. Single nucleotide polymorphisms of CBX4 and CBX7 decrease the risk of hepatocellular carcinoma. Biomed Res Int. 2019;2019:6436825. doi:10.1155/2019/6436825
25. Ferreira D, Soares M, Correia J, Adega F, Ferreira F, Chaves R. Assessment of ERBB2 and TOP2α gene status and expression profile in feline mammary tumors: findings and guidelines. Aging. 2019;11(13):4688–4705. doi:10.18632/aging.102079
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.