Back to Journals » Cancer Management and Research » Volume 12
Long Non-Coding RNA LINC00491 Contributes to the Malignancy of Non-Small-Cell Lung Cancer via Competitively Binding to microRNA-324-5p and Thereby Increasing Specificity Protein 1 Expression
Authors Zhang X, Zhao X, Wang Y, Xing L
Received 28 May 2020
Accepted for publication 19 July 2020
Published 6 August 2020 Volume 2020:12 Pages 6779—6793
DOI https://doi.org/10.2147/CMAR.S264681
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
This paper has been retracted
Xiaoning Zhang 1 ,* Xia Zhao 1 ,* Yanqing Wang, 1 Liqun Xing 2
1Department of Chest Surgery, Weifang Yidu Central Hospital, Weifang, Shandong 262500, People’s Republic of China; 2Department of Surgical, Jinan Infectious Disease Hospital, Jinan, Shandong 250021, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Liqun Xing
Department of Surgery, Jinan Infectious Disease Hospital, 22029 Jingshi Road, Jinan, Shandong 250021, People’s Republic of China
Email [email protected]
Background: A long non-coding RNA termed as long intergenic non-protein coding RNA 491 (LINC00491) has been validated as an oncogene to promote cancer progression in colon adenocarcinoma. The goal of this study was to determine the expression and carcinogenic functions of LINC00491 in non-small-cell lung cancer (NSCLC). Besides, it was aimed to understand how LINC00491 affects the malignant processes of NSCLC cells.
Methods: The expression of LINC00491 in NSCLC was investigated by bioinformatic analysis and reverse transcription-quantitative PCR. After LINC00491 knockdown, cell counting kit-8 assay, flow cytometry, migration and invasion detection assays as well as nude mice xenograft assay were conducted to test the roles of LINC00491 in NSCLC cells. Two online databases, StarBase 3.0 and miRDB, were utilized to determine the putative target miRNA of LINC00491, and the prediction was subsequently confirmed by luciferase reporter assay, RNA immunoprecipitation assay, reverse transcription-quantitative PCR, Western blotting, and rescue assays.
Results: LINC00491 was overexpressed in both NSCLC tissues and cell lines. Functional investigation revealed that depleted LINC00491 facilitated cell apoptosis and decreased cell proliferation, migration, and invasion in vitro. Additionally, the downregulation of LINC00491 impaired NSCLC cell tumor growth in vivo. Mechanistically, LINC00491 functioned as a competing endogenous RNA by sponging microRNA-324-5p (miR-324-5p) in NSCLC cells. miR-324-5p was weakly expressed in NSCLC and exerted tumor-suppressing actions during cancer progression. Furthermore, specificity protein 1 (SP1) was validated as the direct target of miR-324-5p in NSCLC and was under the regulation of LINC00491 via sponging miR-324-5p. Rescue experiments reconfirmed that miR-324-5p inhibition and SP1 overexpression both abrogated the suppressive roles of LINC00491 deficiency in NSCLC cells.
Conclusion: LINC00491 promoted the oncogenicity of NSCLC via serving as a miR-324-5p sponge, which further upregulated the expression of SP1. The LINC00491/miR-324-5p/SP1 pathway disclosed a new mechanism of NSCLC pathogenesis and may provide effective targets for better NSCLC treatment.
Keywords: non-small-cell lung cancer management, long non-coding RNA, ceRNA hypothesis
Background
Lung cancer ranks as the most common type of malignancy and the leading cause of tumor-associated mortalities worldwide.1 Annually, lung cancer affects more than 2 million novel cases and causes nearly 1.7 million deaths reported globally.2 Non-small-cell lung cancer (NSCLC) is the primary pathology subtype of lung cancer and accounts for over 85% of all lung cancer cases.3 Over the past decades, despite tremendous advancements in diagnostic and therapeutic strategies, the clinical efficiency of NSCLC is only slightly improved, and the 5-year overall survival rate of patients with NSCLC is still less than 15%.4 Tumor recurrence and distant metastasis in the progression of NSCLC are in charge of about 90% of the cases succumbed to NSCLC.5,6 Another major cause of a poor prognosis is that a large number of patients with NSCLC are diagnosed in the middle or advanced stages and consequently miss the best opportunities for surgical excision.7 Therefore, adequate studying of the molecular processes behind NSCLC pathogenesis is imperative and of great importance for the identification of attractive novel diagnostic and therapeutic targets.
Long non-coding RNAs (lncRNAs) are a family of evolutionarily conserved RNA transcripts with over 200 nucleotides in length.8 LncRNAs are short of protein coding capacity and, therefore, considered initially as the “noise” of genomic transcription.9 In recent years, emerging evidence supports the importance of lncRNAs in the biological and pathological processes, such as development, differentiation, angiogenesis, and oncogenesis.10–12 The differentially expressed lncRNAs have been revealed to be closely related to genesis and progression of various human cancer types.13,14 Increasing literature has identified lncRNAs as crucial contributors in regulating the malignant characteristic of NSCLC through executing oncogenic or anti-oncogenic activities.15–17
microRNAs (miRNAs) belong to a group of non-coding RNA transcripts, which are 17–24 nucleotides.18 They are capable of affecting gene expression via complementarily base pairing to the 3′-untranslated regions (3′-UTRs) of their target mRNAs, thereby resulting in either mRNA degradation or translational suppression.19 The aberrant expression of miRNAs is relevant to human diseases, including cancers.20,21 An abundance of miRNAs is found to be dysregulated in NSCLC and perform tumor-suppressing or tumor-inhibiting roles during NSCLC oncogenesis and progression.22–24 The competing endogenous RNA (ceRNA) hypothesis suggests that lncRNA can competitively bind to miRNAs, thus spared the negative regulation of miRNAs on their target mRNAs.25 Hence, a thorough investigation of the specific roles of lncRNA and miRNAs in NSCLC may be of help for developing effective targets for cancer diagnosis and treatment.
A lncRNA termed as LINC00491 has been validated as an oncogene to promote cancer progression in colon adenocarcinoma.26 Nevertheless, the expression and roles of LINC00491 in NSCLC remain mostly elusive. The goal of this research was to determine the expression and carcinogenic functions of LINC00491 in NSCLC cells. Additionally, the underlying molecular mechanism was determined, and it was confirmed that LINC00491 competitively binds to miR-324-5p in NSCLC cells and therefore improves SP1 expression.
Methods
Patients and Samples
Human NSCLC tissues and corresponding adjacent normal tissues were collected from 57 patients with NSCLC at Weifang Yidu Central Hospital (Weifang, China). All participants had not received preoperative chemotherapy, radiotherapy, or other anti-cancer therapies. Tissues were immediately immersed in liquid nitrogen at the time of surgery and kept in liquid nitrogen until use. This work was carried out with the approval of the Ethics Committee of Weifang Yidu Central Hospital (2015.#0216) and performed in accordance with the Declaration of Helsinki. Written informed consent was collected from all patients enrolled.
Cell Lines
Human NSCLC cell lines, including H522, H460, SK-MES-1, and A549, and a human nontumorigenic bronchial epithelial cell line, BEAS-2B, were obtained from the Institute of Biochemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China). Cell lines H522, H460, and A549 were grown in RPMI 1640 Media (Gibco; Thermo Fisher Scientific, Waltham, MA, USA), while Minimal Essential Medium (Gibco; Thermo Fisher Scientific) was used for the SK-MES-1 cells. Both basal media were supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific) and 1% penicillin-streptomycin mixture (Gibco; Thermo Fisher Scientific). BEAS-2B cells were maintained in the BEGMTM Bronchial Epithelial Cell Growth Medium (Lonza/Clonetics Corporation, Walkersville, MD, USA) containing 0.5 ng/mL epidermal growth factor, 500 ng/mL hydrocortisone, 0.035 ng/mL bovine pituitary extract, 500 mM ethanolamine, 500 nM ethanolamine phosphate, 0.01 mg/mL adrenaline, and 0.1g/mL retinoic acid. All cells were cultured in a humidified atmosphere at 37°C in the presence of 5% CO2.
Plasmids, Oligonucleotides, and Cell Transfection
The negative control small interfering RNA (siRNA) (si-NC) and LINC00491-specific siRNA (si-LINC00491) were chemically produced by RiboBio (Guangzhou, China). The overexpression and suppression of miR-324-5p were conducted by separate transfection of miR-324-5p mimic, and miR-324-5p inhibitor (anti-miR-324-5p) purchased from GenePharma (Shanghai, China). The miRNA mimic negative control (miR-NC) and negative control miRNA inhibitor (anti-miR-NC) served as the control for miR-324-5p mimic and anti-miR-324-5p, respectively. SP1 overexpression plasmid was established using the pcDNA3.1, which was provided by Sangon (Shanghai, China). Lipofectamine® 2000 (Invitrogen; Thermo Fisher Scientific) was utilized to transfect the NSCLC cells with the abovementioned plasmids or oligonucleotides.
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
Total RNA was extracted using Beyozol (Beyotime, Shanghai, China) and reverse transcribed into cDNA using the PrimeScript™ RT Reagent Kit (TaKaRa Bio, Dalian, China). For the detection of LINC00491 and SP1, quantitative polymerase chain reaction was performed using an ABI 7500 Real-Time PCR System (Thermo Fisher Scientific) by using a TB Green Premix Ex Taq II (Tli RNaseH Plus) (TaKaRa Bio). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) functioned as the internal control for LINC00491 and SP1 expression.
To quantify miR-324-5p, reverse transcription was performed using the miScript Reverse Transcription Kit (Qiagen GmbH, Hilden, Germany). The resultant cDNA was subjected to quantitative polymerase chain reaction using a miScript SYBR Green PCR Kit (Qiagen GmbH). The expression of miR-324-5p was normalized to that of U6 small nuclear RNA. The 2−ΔΔCt method was used to calculate gene expression.
The primers were designed as follows: LINC00491, 5′- CTGGCACTCCCTAGTGAGATGAA-3′ (forward) and 5′-GGTTGAGATACACAATGGATTATCCT-3′ (reverse); SP1, 5′-TGCCCCTACTGTAAAGACAGTGAA-3′ (forward) and 5′-CCACAGTATGACCAGGTACACATAAA-3′ (reverse); GAPDH, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ (forward) and 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′ (reverse); miR-324-5p, 5′- TCGGCAGGCGCAUCCCCUAG −3′ (forward) and 5′- CACTCAACTGGTGTCGTGGA −3′ (reverse); and U6, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ (forward) and 5′-CGCTTCACGAATTTGCGTGTCAT-3′ (reverse).
Subcellular Fractionation
NSCLC cells in the logarithmic growth phase were collected, and the abruption of nuclear and cytoplasmic fractions was performed using the Cytoplasmic & Nuclear RNA Purification Kit (Norgen, Belmont, CA, USA). Next, RT-qPCR was performed to detect the relative LINC00491 expression in both fractions.
Cell Counting Kit-8 (CCK-8) Assay
Transfected cells were trypsinized with trypsin after 24 h of transfection. After centrifugation, the cells were resuspended in the complete culture medium, and the cell density was adjusted to 2.5 × 104 cells/mL. A total of 2500 cells were plated in the 96-well plates and incubated at 37°C in the presence of 5% CO2. At 0, 24, 48, and 72 h after cell inoculation, 10 µL of CCK-8 reagent (Sigma-Aldrich, St. Louis, MO, USA) was added to each well, and the cells were incubated at 37°C for another 2 h. The optical density of the medium was determined at 450 nm wavelength using a microplate reader (Bio-Rad Laboratories, Inc.).
Flow Cytometry
Cellular apoptosis was detected by an Annexin V–fluorescein isothiocyanate (FITC) apoptosis detection kit (BioLegend, San Diego, CA, USA). Transfected cells were collected after treatment with trypsin without ethylene diamine tetraacetic acid (Gibco; Thermo Fisher Scientific) and washed twice with ice-cooled phosphate-buffered saline (Gibco; Thermo Fisher Scientific), followed by resuspending the cells in 100 μL of 1× Binding. Next, 5 μL of Annexin V–FITC and 5 μL of propidium iodide were added and incubated at room temperature in the dark for 15 min to stain the transfected cells. Finally, the stained cells were analyzed using a flow cytometer (FACScan; BD Biosciences, Franklin Lakes, NJ, USA).
Migration and Invasion Detection Assays
In the migration assay, 5 × 104 transfected cells, which were starved for 24 h, were suspended in 200 μL of FBS-free culture medium and inoculated in the upper chambers of the Transwell insert (BD Biosciences, San Jose, CA, USA). In the invasion assay, an equal number of cells were placed in the upper chambers, which were pre-coated with Matrigel (BD Biosciences). For both assays, the bottom of the chambers contained 500 μL of the complete culture medium supplemented with 10% FBS. Following plate incubation at 37°C for 24 h, the cells that failed to pass through the 8-μm pores on the membranes were carefully removed with a ball of cotton wool. The migrated and invaded cells adhering to the surface of membranes were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. The stained cells were then photographed and counted under an inverted light microscope (Olympus Corporation, Tokyo, Japan).
Xenograft Tumor Growth Assay
Lentiviral stably expressing negative control short hairpin RNA (shRNA)(sh-NC) or LINC00491-specific shRNA (sh-LINC00491) was designed and packaged by GenePharma. H522 cells were transduced with the lentivirus, and stable cells expressing high levels of sh-LINC00491 or sh-NC were selected by treating with puromycin.
4-week-old male BALB/c nude mice were obtained from SLAC Laboratory Animal Center (Shanghai, China) and housed in specific pathogen-free conditions. The H522 cells stably transduced with sh-LINC00491 or sh-NC were collected, rinsed with phosphate-buffered saline, and subcutaneously inoculated into nude mice. The diameter of formed subcutaneous xenografts was detected weekly using a vernier caliper, and tumor volumes were analyzed by following the formula: Tumor volume = 0.5 × (length × width2). All mice were euthanized after 5 weeks of cell inoculation, and the subcutaneous xenografts were excised and weighed. All experimental steps involving animals were authorized by the Experimental Animal Ethics Committee of Weifang Yidu Central Hospital (2019.#0502), and carried out in compliance with the Animal Protection Law of the People’s Republic of China-2009 for experimental animals.
Bioinformatic Analysis
Gene Expression Profiling Interactive Analysis (GEPIA; http://gepia.cancer-pku.cn/) containing the TCGA and GTEx databases was employed to determine the expression profile of the LINC00491 in NSCLC. Moreover, patient survival analysis of C1QTNF1-AS1 was also conducted using GEPIA. The localization of the LINC00491 was predicted via lncLocator (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/). YM500v3 (http://120.110.158.132:8787/ym500v3/), a smRNA-Seq database for cancer research, was used to determine the expression correlation between miR-324-5p and overall survival in NSCLC. The association between LINC00491 or SP1 expression and overall survival in NSCLC was analyzed by Kaplan-Meier Plotter (www.kmplot.com/lung).
The putative miRNAs that may interact with LINC00491 were searched using StarBase version 3.0 (http://starbase.sysu.edu.cn/) and miRDB (http://mirdb.org/). Three online databases for miRNA target prediction: StarBase version 3.0, miRDB, and TargetScan (http://www.targetscan.org/), were used to find potential targets for miR-324-5p.
Luciferase Reporter Assay
The wild-type (WT) fragments of LINC00491 with the predictive miR-324-5p binding site and the mutant (MUT) LINC00491 fragments were amplified and inserted into the psiCHECK-2 luciferase plasmid (Promega, Madison, WI, USA), termed as LINC00491-WT and LINC00491-MUT. Similarly, the SP1-WT and SP1-MUT reporter plasmids were synthesized as described in the above experimental steps. NSCLC cells were seeded into 24-well plates and cotransfected with miR-324-5p mimic or miR-NC and constructed reporter plasmids. After 48 h, transfected cells were harvested and analyzed with a Dual-Luciferase Reporter Assay System (Promega) for determining the luciferase activity.
RNA Immunoprecipitation (RIP) Assay
The interaction between miR-324-5p and LINC00491 in NSCLC cells was tested using the Magna RIP RNA Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA, USA). Firstly, the NSCLC cells were lysed in the RIP lysis buffer, and then the acquired 100 μL cell lysate was diluted in 900 μL RIP lysis buffer, and cultivated with magnetic beads coated with anti-Ago2 or anti-IgG control antibodies (Millipore). 10 μL cell lysate was regarded as the Input control. Following Proteinase K treatment, immunoprecipitated RNA was isolated, and RT-qPCR analysis was achieved to evaluate the miR-324-5p and LINC00491 enrichment.
Western Blot Analysis
Total proteins were extracted using RIPA lysis buffer (Beyotime), and the concentration of proteins was detected using an Enhanced BCA Protein Assay Kit (Beyotime). Next, 10% SDS-PAGE gel was used to conduct electrophoresis by using equal amounts of proteins, which were loaded in each lane of the gel, followed by transferring to PVDF membranes (Beyotime). The membranes were blocked with 5% skimmed milk in Tris-buffered saline (TBS) containing 0.05% Tween-20 (TBST) for 2 h at room temperature. Afterward, TBST was adopted to rinse the membranes, and primary antibodies were utilized to incubate the membranes overnight at 4°C. The primary antibodies against SP1 (ab124804; Abcam, Cambridge, UK.) and GAPDH (ab128915; Abcam) were used after diluting 1000-fold. As the secondary antibody, horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G secondary antibody (ab205718; Abcam) was employed. The cultivation was carried out at room temperature for 2 h. Finally, the target signals were developed by the use of BeyoECL Star Western Blotting Detection Reagents (Beyotime). Here, GAPDH worked as an endogenous control.
Statistical Analysis
For all the results, mean ± standard deviation (SD) values were calculated using three biological replicates in each experiment. The t-test was performed to analyze the comparisons between the two groups. Besides, the differences among multiple groups were analyzed using one-way analysis of variance alongside Tukey’s posthoc test. Kaplan–Meier method and Log rank test were also utilized to plot and compare the overall survival curves in patients with NSCLC. The correlation between the expressions of LINC00491 and miR-324-5p in NSCLC tissues was examined by conducting the Pearson’s correlation coefficient. Statistical significance was set at P < 0.05.
Results
LINC00491 is Overexpressed in NSCLC and Executes Oncogenic Roles During Cancer Progression
The expression profile of LINC00491 in NSCLC was determined using the GEPIA database. Its expression in lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) was evaluated. LINC00491 was expressed highly in both LUAD and LUSC (Figure 1A). Additionally, the expression of LINC00491 was determined in 57 pairs of NSCLC tissues and corresponding adjacent normal tissues. It was observed that the LINC00491 levels were higher in NSCLC tissues than that in corresponding adjacent normal tissues (Figure 1B).
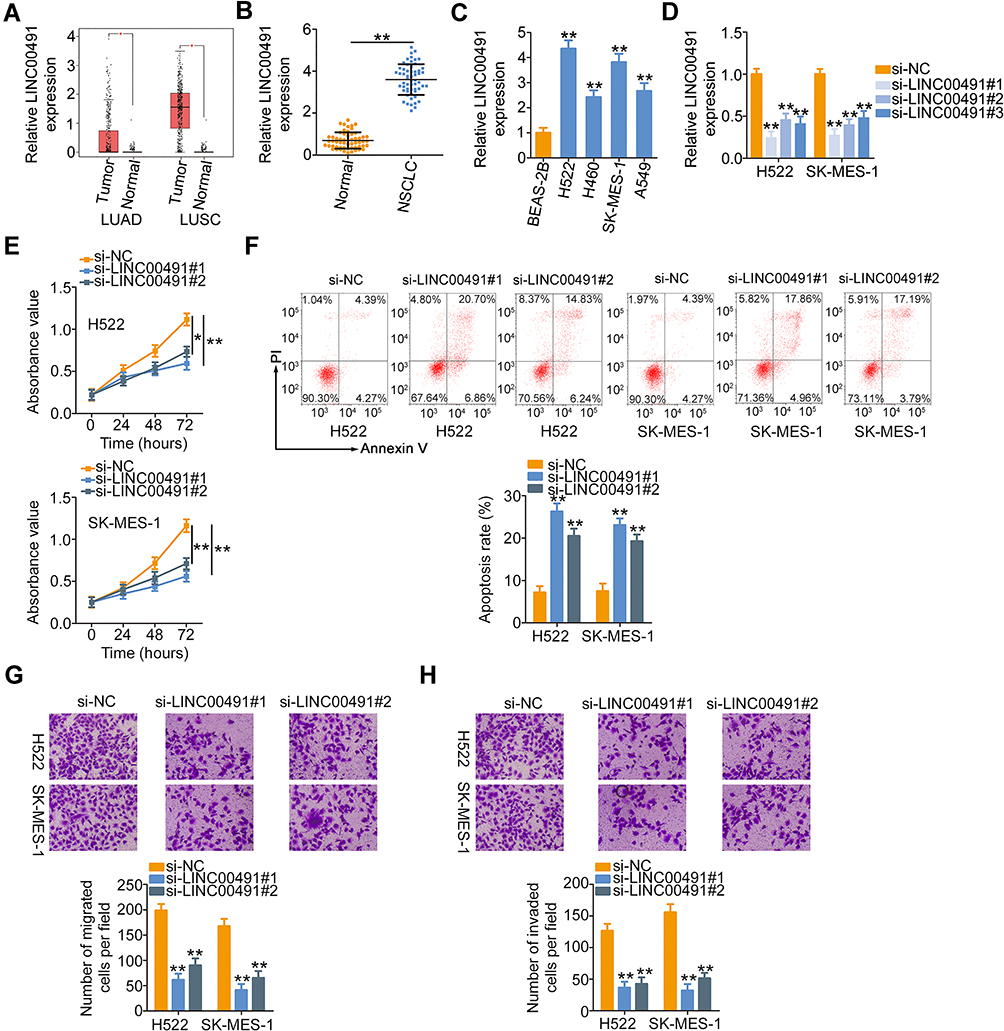 |
Figure 1 LINC00491 was upregulated in NSCLC, and LINC00491 silencing inhibited the biological activities of NSCLC cells. (A) GEPIA database was used to analyze LINC00491 expression in LUAD and LUSC. (B) RT-qPCR was performed to measure the LINC00491 expression in 57 pairs of NSCLC tissues and corresponding adjacent normal tissues. (C) The expression of LINC00491 in NSCLC cells was detected via RT-qPCR. A human nontumorigenic bronchial epithelial cell line BEAS-2B served as the control. (D) H522 and SK-MES-1 cells were transfected with specific siRNAs targeting the LINC00491 or si-NC and subjected to RT-qPCR analysis to determine transfection efficiency. (E, F) The proliferation and apoptosis were tested in H522 and SK-MES-1 cells after si- LINC00491 or si-NC transfection via CCK-8 assay and flow cytometry analysis. (G, H) Migration and invasion detection assays uncovered the migratory and invasive capacities of H522 and SK-MES-1 cells after LINC00491 downregulation. *P < 0.05 and **P < 0.01. Abbreviations: LINC00491, long intergenic non-protein coding RNA 491; LUAD, in lung adenocarcinoma; LUSC, lung squamous cell carcinoma; NSCLC, non-small cell lung cancer; si-LINC00491, LINC00491-specific small interfering RNA; si-NC, negative control small interfering RNA; PI, propidium iodide. |
To investigate the role of LINC00491 in NSCLC, the expression of LINC00491 was first detected in NSCLC cell lines (H522, H460, SK-MES-1, and A549). The results revealed that LINC00491 was overexpressed in all the four NSCLC cell lines as compared with that of a human nontumorigenic bronchial epithelial cell line BEAS-2B (Figure 1C). Cell lines H522 and SK-MES-1, which presented the highest LINC00491 levels among the four NSCLC cell lines, were transfected with specific siRNAs targeting LINC00491. RT-qPCR affirmed that transfection of si-LINC00491 was successful in H522 and SK-MES-1 cells (Figure 1D). The si-LINC00491#1 manifested the highest efficiency in silencing LINC00491 expression and was therefore used in the functional experiments. CCK-8 assay showed that the loss of LINC00491 restricted the proliferative ability of H522 and SK-MES-1 cells (Figure 1E). Furthermore, knockdown of LINC00491 dramatically increased H522 and SK-MES-1 cell apoptosis (Figure 1F). Besides, migration and invasion detection assays were performed to assess the effects of LINC00491 downregulation on the migration and invasion of NSCLC cells. Inhibition of LINC00491 decreased the migratory (Figure 1G) and invasive (Figure 1H) abilities of H522 and SK-MES-1 cells. These results implied that LINC00491 was a tumor-suppressing lncRNA in NSCLC cells.
LINC00491 Functions as a miR-324-5p Sponge in NSCLC Cells
To explicate the molecular events of LINC00491 involving in the oncogenicity of NSCLC, the location of LINC00491 was first predicted by the lncLocator. The prediction indicated that LINC00491 was mostly located in the cytoplasm (Figure 2A), and this observation was further demonstrated by the subcellular fractionation (Figure 2B). The cytoplasmic lncRNAs might operate as ceRNAs to sponge miRNAs and consequently regulate miRNAs’ target mRNAs at the posttranscriptional level.27 Bioinformatics analysis was performed to identify the target miRNAs that have a chance to interact with LINC00491. The StarBase 3.0 and miRDB online databases revealed that miR-324-5p was the common target miRNA of LINC00491 (Figure 2C). To substantiate the prediction, luciferase reporter assay was performed to determine the binding between miR-324-5p and LINC00491 in NSCLC cells. The WT and MUT miR-324-5p binding sites within LINC00491 was displayed in Figure 2D. Luciferase reporter assay results indicated that miR-324-5p upregulation suppressed the luciferase activity of LINC00491-WT in H522 and SK-MES-1 cells but exerted no impact on the luciferase activity of LINC00491-MUT (Figure 2E). Furthermore, RIP assay showed that LINC00491 and miR-324-5p were coimmunoprecipitated by anti-Ago2 antibody in H522 and SK-MES-1 cells (Figure 2F), which further verified that miR-324-5p was the downstream target miRNA of LINC00491.
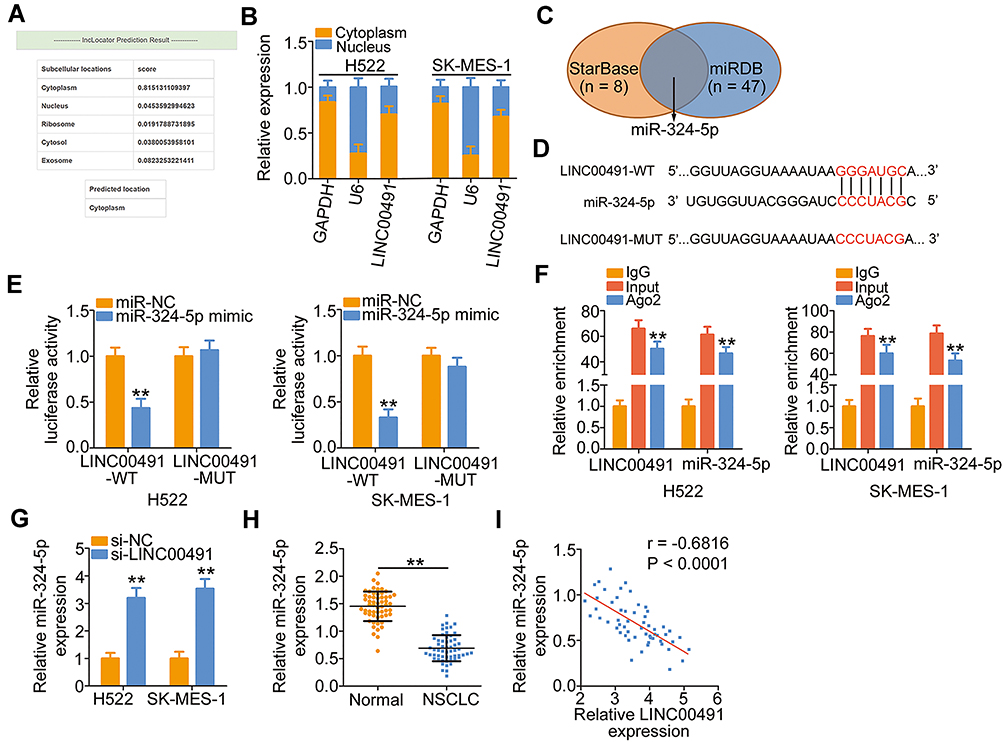 |
Figure 2 LINC00491 functioned as a molecular sponge of miR-324-5p in NSCLC cells. (A) The predicted distribution of LINC00491 by the lncLocator. (B) The nuclear and cytoplasmic fractions of H522 and SK-MES-1 cells were separated by subcellular fractionation. Then, RT-qPCR was performed to determine the location of LINC00491 in H522 and SK-MES-1 cells. (C) Venn diagrams show the common target miRNAs by StarBase 3.0 and miRDB. (D) The complementary wild-type and mutant binding of the miR-324-5p within LINC00152. (E) Luciferase reporter assay was utilized to affirm the binding of LINC00491 to miR-324-5p in NSCLC cells. H522 and SK-MES-1 cells were cotransfected with LINC00491-WT or LINC00491-MUT and miR-324-5p mimic or miR-NC, and luciferase activity was detected after 48 h. (F) The interaction between LINC00491 and miR-324-5p in NSCLC cells was determined through the RIP assay. (G) RT-qPCR analysis illustrated the expression of miR-324-5p in LINC00491 silenced-H522 and SK-MES-1 cells. (H) miR-324-5p expression in 57 pairs of NSCLC tissues and corresponding adjacent normal tissues was measured by RT-qPCR. (I) The expression correlation between LINC00491 and miR-324-5p was assessed in 57 NSCLC tissues by Pearson’s correlation coefficient. **P < 0.01. Abbreviations: LINC00491, long intergenic non-protein coding RNA 491; miR-324-5p, microRNA-324-5p; WT, wild-type; MUT, mutant; Ago2, Argonaute 2; IgG, immunoglobulin G; NSCLC, non-small cell lung cancer; si-LINC00491, LINC00491-specific small interfering RNA; si-NC, negative control small interfering RNA; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; U6, U6 small nuclear RNA. |
To explore the regulatory action of LINC00491 on miR-324-5p, LINC00491 was silenced in H522 and SK-MES-1 cells, and its effect on miR-324-5p expression was analyzed by the RT-qPCR. Depleted LINC00491 caused a noticeable increase of miR-324-5p expression in H522 and SK-MES-1 cells (Figure 2G). Furthermore, miR-324-5p was downregulated in NSCLC tissues in contrast to corresponding adjacent normal tissues (Figure 2H). Pearson’s correlation coefficient was then adopted to examine the expression correlations between LINC00491 and miR-324-5p in the 57 NSCLC tissues, and a notable inverse correlation between them was identified (Figure 2I; r = −0.6816, P < 0.0001). In brief, LINC00491 worked as a ceRNA for sponging miR-324-5p in NSCLC cells.
SP1 is a Direct Target of miR-324-5p in NSCLC Cells and is Under the Control of LINC00491 via Sponging miR-324-5p
The weak expression of miR-324-5p in NSCLC implied its involvement in the malignancy of NSCLC. To test the biological activities of miR-324-5p in NSCLC progression, miR-324-5p mimic was utilized to increase miR-324-5p expression in H522 and SK-MES-1 cells (Figure 3A), and the miR-324-5p overexpressed-H522 and SK-MES-1 cells were then used in functional studies. Transfection with miR-324-5p mimic hindered H522 and SK-MES-1 cell proliferation (Figure 3B) and increased cell apoptosis (Figure 3C and D), as evidenced by the CCK-8 assay and flow cytometry analysis. Furthermore, the migratory (Figure 3E) and invasive (Figure 3F) capabilities of H522 and SK-MES-1 cells were dramatically impaired by miR-324-5p overexpression. These results suggested that miR-324-5p has a cancer-inhibiting role during NSCLC progression.
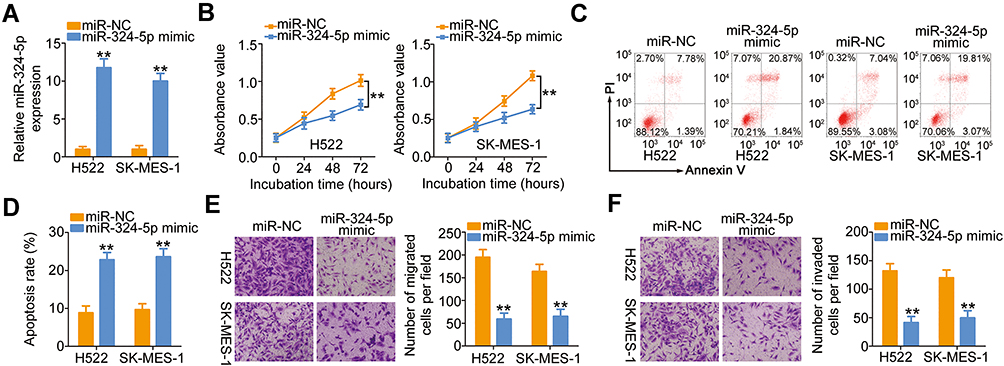 |
Figure 3 LINC00491 positively regulated SP1 expression in NSCLC cells via sponging miR-324-5p. (A) RT-qPCR was conducted to detect miR-324-5p expression in H522 and SK-MES-1 cells after introducing the miR-324-5p mimic or miR-NC. (B–D) The proliferation and apoptosis of H522 and SK-MES-1 cells after the overexpression of miR-324-5p was analyzed via the CCK-8 assay and flow cytometry analysis. (E, F) Cell migration and invasion were detected in H522 and SK-MES-1 cells after being transfected with miR-324-5p mimic or miR-NC. **P < 0.01. Abbreviations: miR-324-5p, microRNA-324-5p; PI, propidium iodide; miR-NC, miRNA mimic negative control; WT, wild-type; MUT, mutant. |
Using the bioinformatics analysis, multiple targets of miR-324-5p were found. Among them, SP1 (Figure 4A) was chosen for further corroboration due to its well-characterized pro-oncogenic roles during NSCLC progression.28–30 Luciferase reporter assay was employed to verify the predicted results. The results revealed that ectopic miR-324-5p expression reduced the luciferase activity driven by the SP1-WT in H522 and SK-MES-1 cells, while it did not affect the luciferase activity of SP1-MUT (Figure 4B). Synchronously, transfection with miR-324-5p mimic resulted in a significant downregulation of SP1 mRNA (Figure 4C) and protein (Figure 4D) expression in H522 and SK-MES-1 cells, whereas transfection with anti-miR-324-5p clearly increased the expression of SP1 mRNA (Figure 4E) and protein (Figure 4F) levels. Furthermore, the expression of SP1 mRNA was elevated in NSCLC tissues (Figure 4G) and showed a negative correlation with miR-324-5p expression (Figure 4H; r = −0.6588, P < 0.0001).
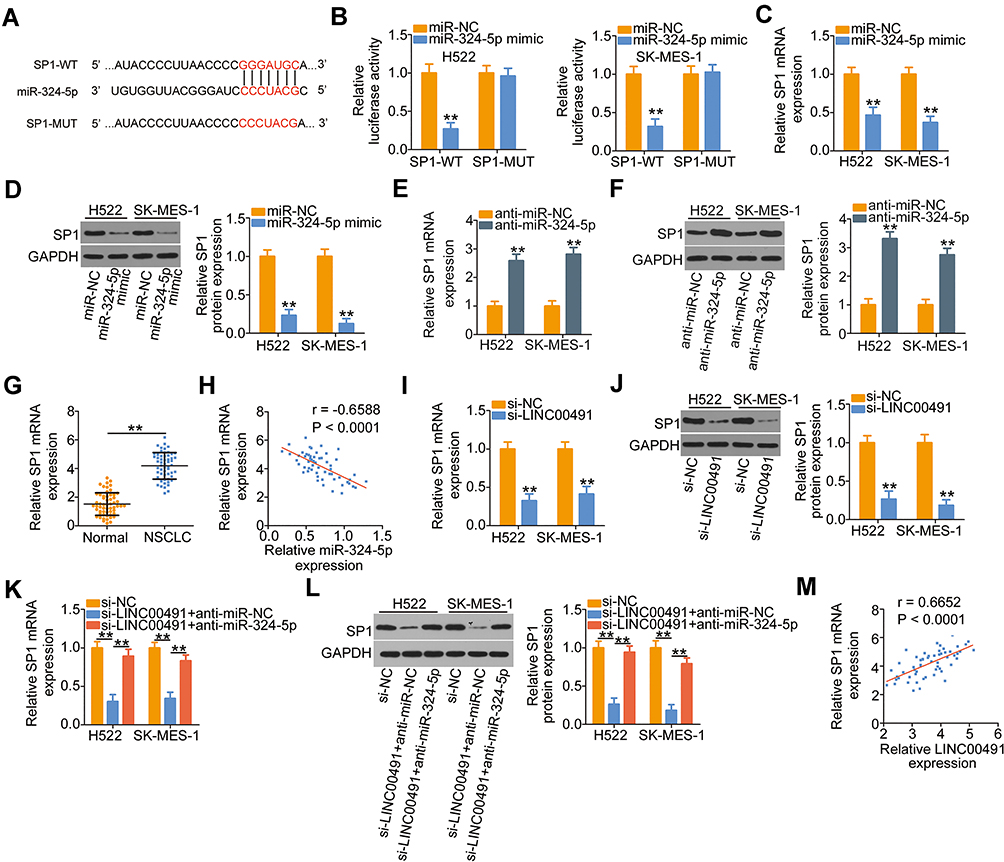 |
Figure 4 SP1 was a direct target of miR-324-5p and could be regulated by LINC00491 in NSCLC cells via sponging miR-324-5p. (A) A potential binding site for miR-324-5p on the 3′-UTR of SP1. The mutant binding sequences were also presented. (B) SP1-WT or SP1-MUT, in combination with miR-324-5p mimic or miR-NC, was introduced into H522 and SK-MES-1 cells. Detection of the luciferase activity was conducted at 48 h post-transfection. (C, D) The expression of SP1 mRNA and protein was detected in H522 and SK-MES-1 cells after miR-324-5p upregulation by RT-qPCR and Western blotting, respectively. (E, F) RT-qPCR and Western blotting were utilized to determine SP1 mRNA and protein levels in H522 and SK-MES-1 cells that were transfected with anti-miR-324-5p or anti-miR-NC. (G) RT-qPCR analysis revealed the SP1 mRNA levels in 57 pairs of NSCLC tissues and corresponding adjacent normal tissues. (H) The correlation between the expressions of SP1 mRNA and miR-324-5p in 57 NSCLC tissues was explored by Pearson’s correlation coefficient. (I, J) The regulatory roles of LINC00491 knockdown on the levels of SP1 mRNA and protein in H522 and SK-MES-1 cells were investigated by RT-qPCR and Western blotting. (K, L) H522 and SK-MES-1 cells were transfected with anti-miR-324-5p or anti-miR-NC in the presence of si-LINC00491. RT-qPCR and Western blotting were performed to determine the expression change of SP1 mRNA and protein, respectively. (M) The association of expression levels of LINC00491 and SP1 mRNA in 57 NSCLC tissues analyzed by Pearson’s correlation coefficient. **P < 0.01. Abbreviations: LINC00491, long intergenic non-protein coding RNA 491; miR-324-5p, microRNA-324-5p; NSCLC, non-small cell lung cancer; si-LINC00491, LINC00491-specific small interfering RNA; si-NC, negative control small interfering RNA; WT, wild-type; MUT, mutant; SP1, specificity protein 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; anti-miR-NC, negative control miRNA inhibitor; anti-miR-324-5p, miR-324-5p inhibitor. |
To investigate how LINC00491 affects the expression of SP1, the LINC00491 deficient-H522 and SK-MES-1 cells were subjected to RT-qPCR and Western blotting. The mRNA (Figure 4I) and protein (Figure 4J) levels of SP1 were considerably decreased in H522 and SK-MES-1 cells after LINC00491 depletion, and the regulatory actions were abrogated by the cotransfection with anti-miR-324-5p (Figure 4K and L). Furthermore, Pearson’s correlation coefficient revealed that the expressions of SP1 mRNA in 57 NSCLC tissues were positively correlated with the level of LINC00491 (Figure 4M; r = 0.6652, P < 0.0001). Based on these findings, it was suggested that SP1 was a direct target of miR-324-5p in NSCLC cells and could be positively regulated by LINC00491 via sponging miR-324-5p.
LINC00491 Promotes Cell Carcinogenesis in NSCLC by Modulating miR-324-5p/SP1 Axis
The results suggested that LINC00491 served as a ceRNA in positively regulating SP1 expression through sponging miR-324-5p. Hence, rescue experiments were also performed to determine whether miR-324-5p/SP1 axis was required for LINC00491 mediated tumor-promoting roles in NSCLC. First, H522 and SK-MES-1 cells were transfected with anti-miR-324-5p or anti-miR-NC, and then RT-qPCR corroborated that the expression of miR-324-5p was decreased in H522 and SK-MES-1 cells after anti-miR-324-5p transfection (Figure 5A). Next, anti-miR-324-5p or anti-miR-NC in parallel with si-LINC00491 was cotransfected into H522 and SK-MES-1 cells, and the functional studies demonstrated that inhibition of miR-324-5p could partially abolish the impacts of LINC00491 deletion on the proliferation (Figure 5B), apoptosis (Figure 5C), migration (Figure 5D), and invasion (Figure 5E) of H522 and SK-MES-1 cells.
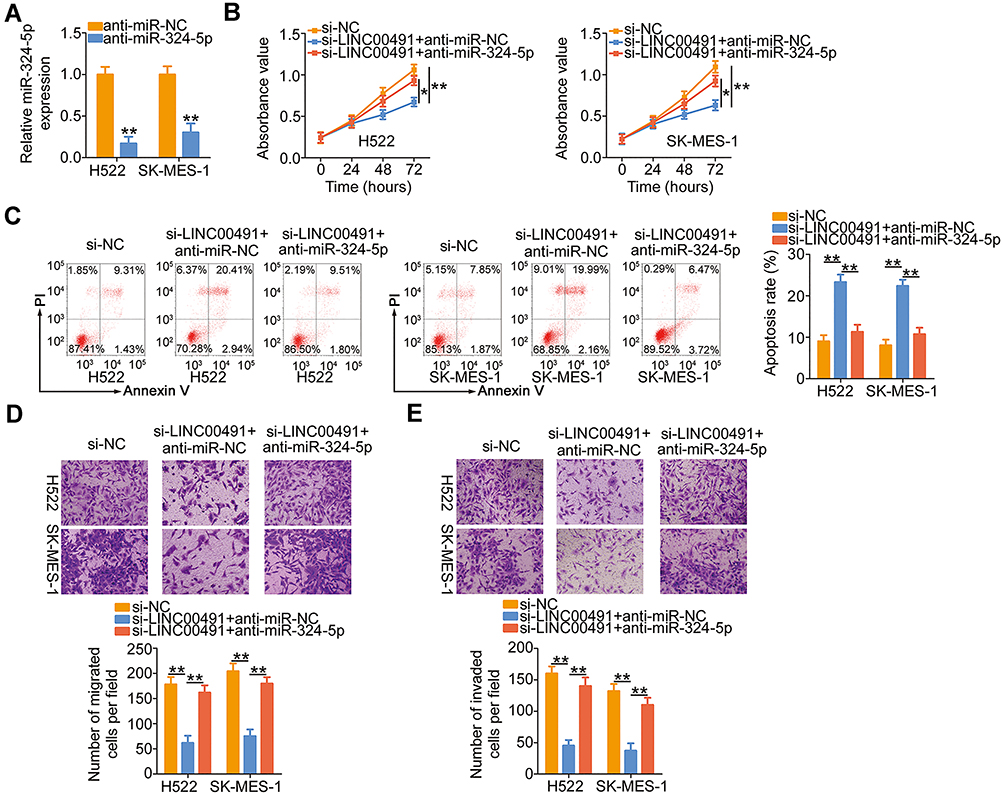 |
Figure 5 miR-324-5p inhibition reverses the impacts of LINC00491 depletion on NSCLC cells. (A) The expression of miR-324-5p was detected in H522 and SK-MES-1 cells transfected with anti-miR-324-5p or anti-miR-NC. (B–E) CCK-8 assay, flow cytometry analysis, and migration and invasion detection assays were carried out to, respectively, determine the proliferation, apoptosis, migration, and invasion of H522 and SK-MES-1 cells after cotransfection with anti-miR-324-5p or anti-miR-NC and si-LINC00491. *P < 0.05 and **P < 0.01. Abbreviations: LINC00491, long intergenic non-protein coding RNA 491; si-LINC00491, LINC00491-specific small interfering RNA; si-NC, negative control small interfering RNA; PI, propidium iodide; miR-324-5p, microRNA-324-5p; anti-miR-NC, negative control miRNA inhibitor; anti-miR-324-5p, miR-324-5p inhibitor. |
Similarly, si-LINC00491 was cotransfected with SP1 overexpressed plasmid pcDNA3.1-SP1 or empty pcDNA3.1 plasmid in H522 and SK-MES-1 cells. Western blotting confirmed the efficiency of pcDNA3.1-SP1 in increasing SP1 expression (Figure 6A). Upregulation of SP1 restored the proliferation of H522 and SK-MES-1 cells suppressed by LINC00491 downregulation (Figure 6B). Additionally, the regulatory actions of LINC00491 silencing on H522 and SK-MES-1 cell apoptosis were abated by the addition of SP1 (Figure 6C). Furthermore, cotransfection with pcDNA3.1-SP1 attenuated the inhibitory influences of si-LINC00491 on the migration (Figure 6D) and invasion (Figure 6E) of the two cell lines. Collectively, LINC00491 aggravated the biological activities of NSCLC by regulating the output of miR-324-5p/SP1 axis.
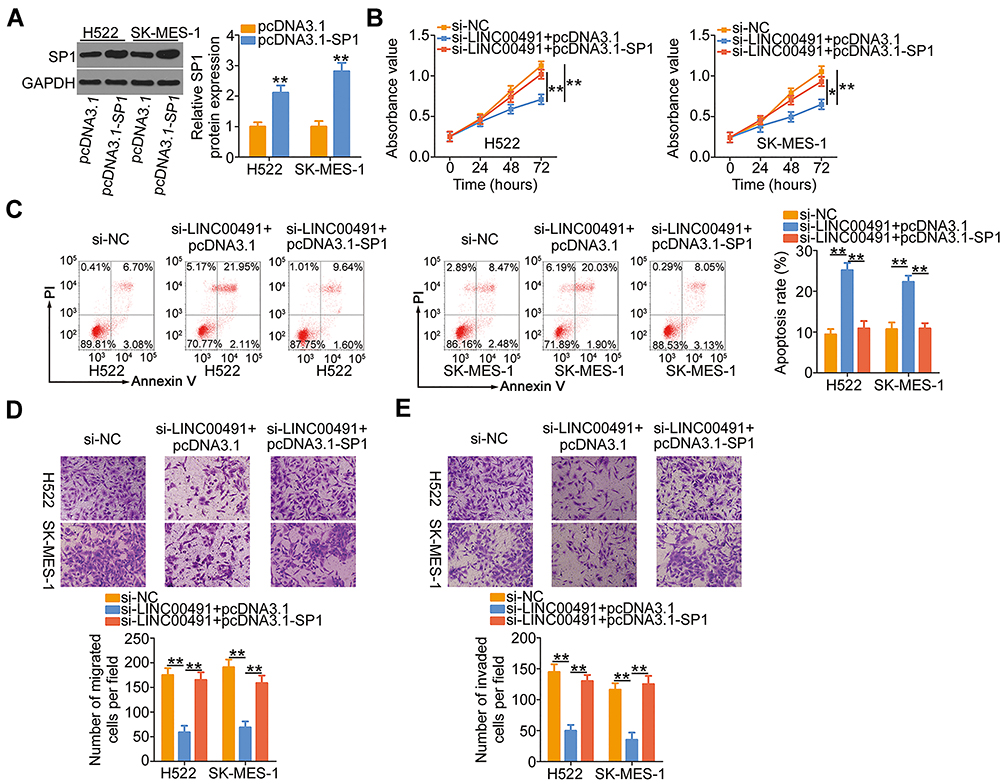 |
Figure 6 SP1 overexpression abrogated the inhibitory functions of si-LINC00491 on NSCLC cells. (A) The overexpression efficiency of pcDNA3.1-SP1 in H522 and SK-MES-1 cells was evaluated by Western blotting. The empty pcDNA3.1 served as the control. (B–E) The pcDNA3.1-SP1 or pcDNA3.1, together with si-LINC00491, was injected into H522 and SK-MES-1 cells. The proliferation, apoptosis, migration, and invasion were analyzed by CCK-8 assay, flow cytometry, and migration and invasion detection assay, respectively. *P < 0.05 and **P < 0.01. Abbreviations: LINC00491, long intergenic non-protein coding RNA 491; si-LINC00491, LINC00491-specific small interfering RNA; si-NC, negative control small interfering RNA; PI, propidium iodide; SP1, specificity protein 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; pcDNA3.1-SP1, SP1 overexpression plasmid. |
LINC00491 Depletion Inhibits Tumor Growth of NSCLC Cells in vivo
To further demonstrate the oncogenic roles of LINC00491 in NSCLC, H522 cells stably expressing sh-LINC00491 or sh-NC were subcutaneously inoculated into the flank of nude mice. The subcutaneous xenografts in the sh-LINC00491 group manifested lower volume (Figure 7A and B) and weight (Figure 7C) compared with those in the sh-NC group. In addition, the tumor xenografts were removed, and LINC00491, miR-324-5p, and SP1 levels were detected. The expression of LINC00491 in LINC00491 stably inference-tumor xenografts was lower than that in the sh-NC group (Figure 7D). Furthermore, the tumor xenografts originated from H522 cells stably transfected with sh-LINC00491 had higher miR-324-5p (Figure 7E) and lower SP1 mRNA (Figure 7F) and protein levels (Figure 7G) than that in the sh-NC groups. Accordingly, LINC00491 knockdown suppressed NSCLC cell tumor growth in vivo by targeting the miR-324-5p/SP1 axis.
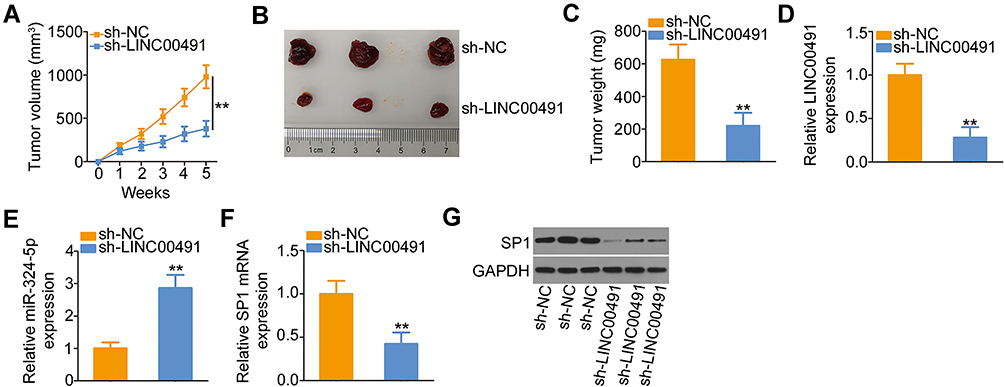 |
Figure 7 The depletion of LINC00491 hindered NSCLC cell tumor growth in vivo. (A) Tumor volume recorded weekly, and growth curves plotted. (B) The picture of tumor xenografts harvested at 5 weeks after the cell injection. (C) Tumor xenografts were harvested in the fifth week, and tumor weight was detected. (D, E) LINC00491 and miR-324-5p expression in tumor xenografts was examined by RT-qPCR. (F, G) RT-qPCR and Western blotting were performed to measure the SP1 mRNA and protein expressions, respectively, in the tumor xenografts. **P < 0.01. Abbreviations: LINC00491, long intergenic non-protein coding RNA 491; sh-LINC00491, LINC00491-specific short hairpin RNA; sh-NC, negative control short hairpin RNA; SP1, specificity protein 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; miR-324-5p, microRNA-324-5p. |
The Clinical Relevance of LINC00491/miR-324-5p/SP1 in NSCLC
The prognostic relevance of LINC00491/miR-324-5p/SP1 was then examined via different database. High expression of LINC00491 was not related to the overall survival of patients with LUAD or LUSC (Figure 8A, P = 0.920). To confirm this observation, the median value of LINC00491 in NSCLC tissues was set as the cut-off line, and all 57 patients with NSCLC were divided into either LINC00491 high (n = 29) or LINC00491 low (n = 28) expression groups. Consistently, the Kaplan–Meier plot revealed that there was no apparent association between the LINC00491 expression and overall survival in the 57 patients with NSCLC (Figure 8B, P = 0.442). However, according to Kaplan-Meier Plotter, a high LINC00491 expression closely related with a shorter overall survival in patients with NSCLC (Figure 8C, P = 8.4e-10). Next, YM500v3 was applied to determine the expression correlation between miR-324-5p and overall survival in NSCLC. The results indicated that the expression of miR-324-5p presented a significant correlation with the overall survival of patients with LUAD (Figure 8D, P = 0.860) or LUSC (Figure 8D, P = 0.594). The correlation between SP1 expression and overall survival in NSCLC was examined via Kaplan-Meier Plotter. As shown in Figure 8E (P = 2.8e-09), patients with NSCLC showing high SP1 expression had shorter overall survival in contrast to patients with low SP1 expression.
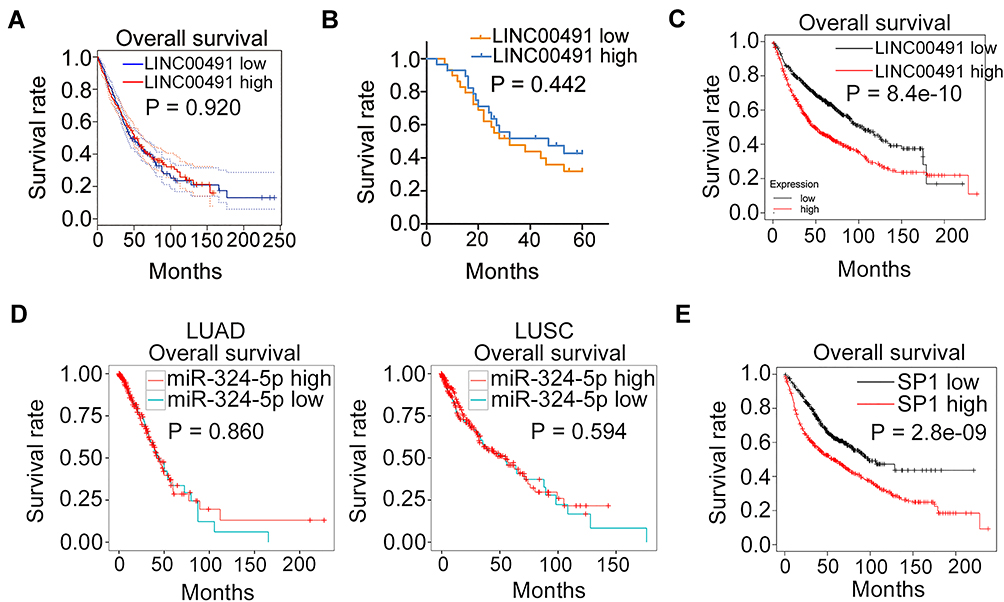 |
Figure 8 The clinical relevance of LINC00491/miR-324-5p/SP1 in NSCLC. (A) GEPIA database revealed the association between the LINC00491 expression and the overall survival in patients with LUAD and LUSC. (B) Kaplan–Meier plots based on 57 patients with NSCLC were examined whether a high LINC00491 expression was related to the overall survival. (C) Kaplan-Meier Plotter database revealed the correlation between the LINC00491 expression and the overall survival in patients with NSCLC. (D) YM500v3 analyzed the expression correlation between miR-324-5p and overall survival in patients with LUAD or LUSC. (E) The correlation between SP1 expression and overall survival in NSCLC was examined via Kaplan-Meier Plotter. Abbreviations: LUAD, in lung adenocarcinoma; LUSC, lung squamous cell carcinoma; LINC00491, long intergenic non-protein coding RNA 491; miR-324-5p, microRNA-324-5p; SP1, specificity protein 1. |
Discussion
Numerous studies have uncovered the dysregulation of lncRNAs in NSCLC and claimed that their abnormal expression could be developed as prediction biomarkers.31–33 The aberrantly expressed lncRNAs have been closely associated with physiological signs of progress in NSCLC and implicated in the cancer oncogenesis and progression.34 Hence, a comprehensive investigation of cancer-associated lncRNAs in NSCLC might be of great significance in treating NSCLC. Many lncRNAs have been validated to date; yet, our understanding of their roles in NSCLC is still tremendously incomplete. In this study, we first determined the expression and specific roles of LINC00491 in NSCLC. More importantly, the relevant molecular events responsible for the cellular response to LINC00491 in NSCLC were elucidated thoroughly.
LINC00491 is overexpressed in colon adenocarcinoma tissues and cell lines.26 Functionally, interference of LINC00491 reduces cell proliferation, colony formation, migration, and invasion in colon adenocarcinoma.26 Nevertheless, it remains unknown whether LINC00491 is differently expressed in NSCLC. In the present study, LINC00491 was upregulated in NSCLC, suggesting that the elevated level of LINC00491 might exert essential roles in NSCLC progression. With regard to clinical value, our results and data obtained from GEPIA database revealed no significant correlation between LINC00491 expression and overall survival in patients with NSCLC. On the contrary, Kaplan-Meier Plotter indicated that patients with NSCLC showing high LINC00491 expression manifested a shorter overall survival in contrast to patients with low LINC00491 expression. The conflicting in-house study may be attributed to different race, eating and living habits and medical condition. In our further studies, we will collect more NSCLC samples and address the issue. Functional investigations showed that the downregulation of LINC00491 promoted NSCLC cell apoptosis and suppressed cell proliferation, migration, and invasion in vitro. Furthermore, in vivo experiments showed that loss of LINC00491 hindered NSCLC cell tumor growth in vivo.
After verifying the roles of LINC00491 in NSCLC, we further unveiled how lncRNA affects the malignant processes of NSCLC cells. Mechanistically, lncRNA has multiple different molecular mechanisms, which is mainly decided by their cellular distribution.35 The nuclear lncRNAs are capable to control cellular behaviors through chromatin interactions, transcriptional regulation, and RNA processing.36 For instance, lncRNA AB074169 interacts with KHSRP protein to regulate CDKN1a mRNA stability, and is involved in the regulation of papillary thyroid carcinoma cell proliferation.37
The ceRNA hypothesis proposed in recent years has been an extraordinary mediatory mechanism of lncRNAs, which indicates that cytoplasmic lncRNAs can competitively bind to miRNAs to decrease the miRNAs-mediated inhibition of target genes.38 Herein, we first predicted the localization of LINC00491 in human cells by the use of the lncLocator, and the prediction suggested that LINC00491 mainly distributed in the cytoplasm. After that, subcellular fractionation, followed by RT-qPCR analysis, verified the accuracy of the prediction results. These results implied that LINC00491 might act as a ceRNA in NSCLC.
To find out the putative target miRNA, two online databases, including StarBase 3.0 and miRDB, was used for the lncRNA target prediction, and miR-324-5p was the common target miRNA of LINC00491. Luciferase reporter and RIP assays were performed and identified direct binding and interaction between miR-324-5p and LINC00491 in NSCLC cells. Meanwhile, miR-324-5p was weakly expressed in NSCLC tissues and manifested a negative correlation with LINC00491 expression. Additionally, miR-324-5p expression was increased in NSCLC cells after LINC00491 silencing. Besides, SP1 was demonstrated as the direct target of miR-324-5p in NSCLC cells, and LINC00491 could raise SP1 expression by adsorbing the miR-324-5p. All these results identified a new ceRNA pathway in NSCLC involving LINC00491, miR-324-5p, and SP1.
miR-324-5p is known to be aberrantly expressed in a variety of human cancers;39–41 however, the expression status and specific roles of miR-324-5p in NSCLC have not been interrogated. The expression of miR-324-5p was decreased in NSCLC, while miR-324-5p mimic transfected into NSCLC cells inhibited cell proliferation, migration, and invasion as well as accelerated cell apoptosis. In this mechanism, SP1, a sequence-specific DNA-binding protein,42 was verified as a direct target of miR-324-5p in NSCLC cells. SP1 was reported to be expressed at high levels in NSCLC and involved in NSCLC progression by regulating multiple aggressive behaviors.28–30 Our rescue experiments further demonstrated that the miR-324-5p inhibition or SP1 overexpression partially reversed the tumor-inhibiting impacts of LINC00491 knockdown in NSCLC cells. These results suggested that the miR-324-5p/SP1 axis was required for LINC00491-triggered regulatory mechanism in the malignancy of NSCLC.
BEAS-2B are bronchial epithelium-derived, and can only serve as a “normal” comparison for LUSC. It was a limitation of our study. In addition, some of the foundational experiments in Figures 1 and 2 could be analyzed in parallel to alveolar epithelial tissue extract and compared to the A549 & H522 cell lines. The two limitations will be addressed in the near future.
Conclusion
In this study, we uncovered the novel aspects of the tumor-promoting activities of LINC00491 in NSCLC. Our results indicated that LINC00491 promoted the oncogenicity of NSCLC by performing as ceRNA for miR-324-5p, which further upregulated the expression of SP1. Our study disclosed a new mechanism of NSCLC pathogenesis and may guide new efforts to counteract cancer progression in the NSCLC.
Abbreviations
FBS, fetal bovine serum; GEPIA, Gene Expression Profiling Interactive Analysis; LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma; NSCLC, non-small-cell lung cancer; SD, standard deviation; TBS, tris-buffered saline; WT, wild-type; XIST, X-inactive specific transcript; LINC00491, long intergenic non-protein coding RNA 491; miR-324-5p, microRNA-324-5p; SP1, specificity protein 1.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34. doi:10.3322/caac.21551
2. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
3. Saintigny P, Burger JA. Recent advances in non-small cell lung cancer biology and clinical management. Discov Med. 2012;13(71):287–297.
4. Custodio A, Mendez M, Provencio M. Targeted therapies for advanced non-small-cell lung cancer: current status and future implications. Cancer Treat Rev. 2012;38(1):36–53. doi:10.1016/j.ctrv.2011.04.001
5. Yousefi M, Bahrami T, Salmaninejad A, Nosrati R, Ghaffari P, Ghaffari SH. Lung cancer-associated brain metastasis: molecular mechanisms and therapeutic options. Cell Oncol. 2017;40(5):419–441. doi:10.1007/s13402-017-0345-5
6. Gupta GP, Massague J. Cancer metastasis: building a framework. Cell. 2006;127(4):679–695. doi:10.1016/j.cell.2006.11.001
7. Harris K, Dhillon SS. Focused issue on diagnosis and management of lung cancer patients. Ann Transl Med. 2019;7(15):347. doi:10.21037/atm.2019.07.34
8. Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways. Cancer Cell. 2016;29(4):452–463. doi:10.1016/j.ccell.2016.03.010
9. Patrushev LI, Kovalenko TF. Functions of noncoding sequences in mammalian genomes. Biochemistry (Mosc). 2014;79(13):1442–1469. doi:10.1134/S0006297914130021
10. Lin YH. Crosstalk of lncRNA and cellular metabolism and their regulatory mechanism in cancer. Int J Mol Sci. 2020;21:8. doi:10.3390/ijms21082947
11. Huang Z, Zhou J-K, Peng Y, He W, Huang C. The role of long noncoding RNAs in hepatocellular carcinoma. Mol Cancer. 2020;19(1):77. doi:10.1186/s12943-020-01188-4
12. Wang Y, Sun B, Wen X, et al. The roles of lncRNA in cutaneous squamous cell carcinoma. Front Oncol. 2020;10:158. doi:10.3389/fonc.2020.00158
13. Luo L, Wang M, Li X, et al. Long non-coding RNA LOC285194 in cancer. Clin Chim Acta. 2020;502:1–8. doi:10.1016/j.cca.2019.12.004
14. Ren X. Genome-wide analysis reveals the emerging roles of long non-coding RNAs in cancer. Oncol Lett. 2020;19(1):588–594. doi:10.3892/ol.2019.11141
15. Chen Z, Lei T, Chen X, et al. Long non-coding RNA in lung cancer. Clin Chim Acta. 2020;504:190–200. doi:10.1016/j.cca.2019.11.031
16. Ding X, Wang Q, Tong L, Si X, Sun Y. Long non-coding RNA FOXO1 inhibits lung cancer cell growth through down-regulating PI3K/AKT signaling pathway. Iran J Basic Med Sci. 2019;22(5):491–498. doi:10.22038/ijbms.2019.31000.7480
17. Niazi Z, Garazhian E, Esfandi F, Hassani ZM, Taheri M, Ghafouri-Fard S. Expression analysis of the long non-coding RNA LINC01433 in lung cancer. Klin Onkol. 2019;32(6):453–455. doi:10.14735/amko2019453
18. He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531. doi:10.1038/nrg1379
19. Mendell JT. MicroRNAs: critical regulators of development, cellular physiology and malignancy. Cell Cycle. 2005;4(9):1179–1184. doi:10.4161/cc.4.9.2032
20. Labbe M, Hoey C, Ray J, et al. microRNAs identified in prostate cancer: correlative studies on response to ionizing radiation. Mol Cancer. 2020;19(1):63. doi:10.1186/s12943-020-01186-6
21. Dilsiz N. Role of exosomes and exosomal microRNAs in cancer. Future Sci OA. 2020;6(4):FSO465. doi:10.2144/fsoa-2019-0116
22. Petrek H, Yu AM. MicroRNAs in non-small cell lung cancer: gene regulation, impact on cancer cellular processes, and therapeutic potential. Pharmacol Res Perspect. 2019;7(6):e00528. doi:10.1002/prp2.528
23. Weidle UH, Birzele F, Nopora A. MicroRNAs as potential targets for therapeutic intervention with metastasis of non-small cell lung cancer. Cancer Genomics Proteomics. 2019;16(2):99–119. doi:10.21873/cgp.20116
24. Leonetti A, Assaraf YG, Veltsista PD, El Hassouni B, Tiseo M, Giovannetti E. MicroRNAs as a drug resistance mechanism to targeted therapies in EGFR-mutated NSCLC: current implications and future directions. Drug Resist Updat. 2019;42:1–11. doi:10.1016/j.drup.2018.11.002
25. Wang L, Cho KB, Li Y, Tao G, Xie Z, Guo B. Long noncoding RNA (lncRNA)-mediated competing endogenous RNA networks provide novel potential biomarkers and therapeutic targets for colorectal cancer. Int J Mol Sci. 2019;20(22):5758.
26. Wan J, Deng D, Wang X, Wang X, Jiang S, Cui R. LINC00491 as a new molecular marker can promote the proliferation, migration and invasion of colon adenocarcinoma cells. Onco Targets Ther. 2019;12:6471–6480. doi:10.2147/OTT.S201233
27. Ye Y, Shen A, Liu A. Long non-coding RNA H19 and cancer: a competing endogenous RNA. Bull Cancer. 2019;106(12):1152–1159. doi:10.1016/j.bulcan.2019.08.011
28. Li X, Fu Y, Xia X, et al. Knockdown of SP1/Syncytin1 axis inhibits the proliferation and metastasis through the AKT and ERK1/2 signaling pathways in non-small cell lung cancer. Cancer Med. 2019;8(12):5750–5759. doi:10.1002/cam4.2448
29. Xie JJ, Guo QY, Jin JY, Jin D. SP1-mediated overexpression of lncRNA LINC01234 as a ceRNA facilitates non-small-cell lung cancer progression via regulating OTUB1. J Cell Physiol. 2019;234(12):22845–22856. doi:10.1002/jcp.28848
30. Jiang J, Lv X, Fan L, et al. MicroRNA-27b suppresses growth and invasion of NSCLC cells by targeting Sp1. Tumour Biol. 2014;35(10):10019–10023. doi:10.1007/s13277-014-2294-1
31. Tian LJ, Wu YP, Wang D, et al. Upregulation of Long Noncoding RNA (lncRNA) X-Inactive Specific Transcript (XIST) is associated with cisplatin resistance in Non-Small Cell Lung Cancer (NSCLC) by downregulating MicroRNA-144-3p. Med Sci Monit. 2019;25:8095–8104. doi:10.12659/MSM.916075
32. Qiu HB, Yang K, Yu HY, Liu M. Downregulation of long non-coding RNA XIST inhibits cell proliferation, migration, invasion and EMT by regulating miR-212-3p/CBLL1 axis in non-small cell lung cancer cells. Eur Rev Med Pharmacol Sci. 2019;23(19):8391–8402. doi:10.26355/eurrev_201910_19150
33. Zhou X, Xu X, Gao C, Cui Y. XIST promote the proliferation and migration of non-small cell lung cancer cells via sponging miR-16 and regulating CDK8 expression. Am J Transl Res. 2019;11(9):6196–6206.
34. Lu T, Wang Y, Chen D, Liu J, Jiao W. Potential clinical application of lncRNAs in non-small cell lung cancer. Onco Targets Ther. 2018;11:8045–8052. doi:10.2147/OTT.S178431
35. Tsagakis I, Douka K, Birds I, Aspden JL. Long non-coding RNAs in development and disease: conservation to mechanisms. J Pathol. 2020;250(5):480–495. doi:10.1002/path.5405
36. Pirogov SA, Gvozdev VA, Klenov MS. Long noncoding RNAs and stress response in the nucleolus. Cells. 2019;8:7. doi:10.3390/cells8070668
37. Gou Q, Gao L, Nie X, et al. Long noncoding RNA AB074169 inhibits cell proliferation via modulation of KHSRP-mediated CDKN1a expression in papillary thyroid carcinoma. Cancer Res. 2018;78(15):4163–4174. doi:10.1158/0008-5472.CAN-17-3766
38. Cai Y, Wan J. Competing endogenous RNA regulations in neurodegenerative disorders: current challenges and emerging insights. Front Mol Neurosci. 2018;11:370. doi:10.3389/fnmol.2018.00370
39. Xu HS, Zong HL, Shang M, et al. MiR-324-5p inhibits proliferation of glioma by target regulation of GLI1. Eur Rev Med Pharmacol Sci. 2014;18(6):828–832.
40. Gu C, Zhang M, Sun W, Dong C. Upregulation of miR-324-5p inhibits proliferation and invasion of colorectal cancer cells by targeting ELAVL1. Oncol Res. 2019;27(5):515–524. doi:10.3727/096504018X15166183598572
41. Lin H, Zhou AJ, Zhang JY, Liu SF, Gu JX. MiR-324-5p reduces viability and induces apoptosis in gastric cancer cells through modulating TSPAN8. J Pharm Pharmacol. 2018;70(11):1513–1520. doi:10.1111/jphp.12995
42. Chang WC, Hung JJ. Functional role of post-translational modifications of Sp1 in tumorigenesis. J Biomed Sci. 2012;19:94. doi:10.1186/1423-0127-19-94
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.