Back to Journals » Cancer Management and Research » Volume 11
Long Non-Coding RNA HCP5 Facilitates Cell Invasion And Epithelial-Mesenchymal Transition In Oral Squamous Cell Carcinoma By miR-140-5p/SOX4 Axis
Authors Zhao J, Bai X, Feng C, Shang X, Xi Y
Received 9 September 2019
Accepted for publication 25 October 2019
Published 12 December 2019 Volume 2019:11 Pages 10455—10462
DOI https://doi.org/10.2147/CMAR.S230324
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Jianling Zhao,1,* Xijing Bai,2,* Chong Feng,3 Xinhua Shang,4 Yuli Xi5
1Experimental Center of Dental Hospital Affiliated to Jiamusi University, Jiamusi City 154002, Heilongjiang Province, People’s Republic of China; 2Department of Oral and Maxillofacial Surgery, Tianjin Stomatology Hospital, Tianjin City 300041, People’s Republic of China; 3Department of Orthodontics, Tianjin Stomatology Hospital, Tianjin City 300041, People’s Republic of China; 4Binhu Outpatient Department, Hefei Stomatological Hospital, Hefei City 230001, Anhui Province, People’s Republic of China; 5Department of Stomatology, Hongqi Hospital Affiliated to Mudanjiang Medical University, Mudanjiang City 157000, Heilongjiang Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yuli Xi
Department of Stomatology, Hongqi Hospital Affiliated to Mudanjiang Medical University, No. 1 Taiping Road, Xi’an District, Mudanjiang City 157000, Heilongjiang Province, People’s Republic of China
Email [email protected]
Background: Oral squamous cell carcinoma (OSCC) is the predominant histological type of human oral cancer. In this study, we sought to investigate the functional role of lncRNA HCP5 in OSCC progression.
Methods: The HCP5 and miR-140-5p expression level was determined in 73 paired OSCC tissues and their adjacent normal tissues. Knockdown or overexpression of HCP5 was conducted to investigate the effects of HCP5 on malignant behaviors of OSCC cells. Then, bioinformatic prediction and dual-luciferase reporter assay were conducted to study the interaction between HCP5 and miR-140-5p in OSCC.
Results: Our results demonstrated that HCP5 expression was significantly increased in OSCC tissues and cell lines. High HCP5 level was associated with the aggressive clinicopathological characteristics and poor prognosis of OSCC patients. In vitro gain- and loss-of-function experiments showed that HCP5 overexpression promoted, whereas HCP5 knockdown inhibited the proliferation, migration, invasion and epithelial-mesenchymal transition (EMT) of OSCC cells. Mechanistically, we confirmed that HCP5 might serve as a competitive endogenous RNA (ceRNA) for miR-140-5p to alleviate the repression of its downstream target, SOX4, a master regulator of EMT. Furthermore, restoration of miR-140-5p expression diminished the oncogenic effects of HCP5 on OSCC cells.
Conclusion: Overall, the present study indicated that HCP5/miR-140-5p/SOX4 axis might be a ponderable and promising therapeutic target for OSCC.
Keywords: oral squamous cell carcinoma, long non-coding RNA HCP5, epithelial-mesenchymal transition, competitive endogenous RNA
Introduction
Oral cancer, also named mouth cancer, is a malignant neoplasia which arises on the lip or oral cavity, and oral squamous cell carcinoma (OSCC) is the predominant histological type of oral cancer.1 Although considerable progress has been made in research and therapy, the long-term prognosis of patients suffering from OSCC remains largely unfavorable.2 Accordingly, expounding the possible molecular mechanisms involved in OSCC progression is of great clinical significance.
Long non-coding RNAs (lncRNAs), a class of non-protein coding RNA transcripts with more than 200 nucleotides in length, were initially regarded as the “garbage” of genome transcription. But at present, it has been well implicated that lncRNAs frequently participate in a wide variety of human diseases, including cancers.3 Previous studies have shown that as a member of lncRNAs, human leukocyte antigen (HLA) Complex P5 (HCP5) acts as an oncogenic regulator in follicular thyroid carcinoma, osteosarcoma and breast cancer.4–6 In this study, we aimed to investigate the functional role of HCP5 in OSCC progression and to elucidate the underlying mechanisms.
Materials And Methods
Patients And Tissue Specimens
OSCC tissues and their adjacent normal tissues were obtained from 73 cases of patients who had undergone surgical resection at Dental Hospital Affiliated to Jiamusi University (Jiamusi City, China) and Tianjin Stomatology Hospital (Tianjin City, China). All patients did not receive any radiotherapy or chemotherapy prior to surgery. The tissues were snap-frozen in liquid nitrogen and stored at −80°C for further analysis. The use of human tissues was approved by the Ethics Committee of Dental Hospital Affiliated to Jiamusi University and the Ethics Committee of Tianjin Stomatology Hospital. All patients signed the written informed consent.
Cell Culture And Transfection
Human OSCC cell lines, including SCC-4, Tca8113 and SCC-9, and human normal oral keratinocyte NOK cell line were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). These cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Hyclone, Logan, UT, USA) containing 10% fetal bovine serum (FBS; Sigma-Aldrich, St. Louis, MO, USA), 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C in a humidified incubator with 5% CO2.
The specific small interference RNA (siRNA) targeting HCP5 (si-HCP5), miR-140-5p mimics and the scrambled oligonucleotides (NC) were obtained from GenePharma Co., Ltd (Shanghai, China). The full length sequence of human HCP5 cDNA was amplified by PCR, and then subcloned into the vector pcDNA3.1 (Invitrogen, Carlsbad, CA, USA). Cells grown at 70–80% confluence were transfected with the oligonucleotides and vectors using Lipofectamine 2000 (Invitrogen). After 48 hrs, the cells were collected for further experiments.
RT-qPCR Analysis
Total RNA samples were extracted using TRIzol reagent (Invitrogen). Complementary DNA (cDNA) was synthesised using the PrimeScript RT reagent kit (TaKaRa, Dalian, China). Quantitative PCR (qPCR) analysis was then carried out using a SYBR Green PCR Kit (TaKaRa) on a 7500 Fast Real-Time Sequence detection system (Applied Biosystems, Foster City, CA, USA). Relative gene expression levels were calculated using 2−ΔΔCt method.7 GAPDH or U6 was used as an internal control.
MTT Assay
Cell proliferation was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyl-2H-tetrazolium bromide (MTT) assay. Cells were seeded in a 96-well plate at a density of 3×103 cells/well and incubated for 24, 48 or 72 hrs. Then, 20 µL MTT solution (5 mg/L; Sigma-Aldrich) was added to each well. After incubation for additional 4 hrs, the supernatant was discarded and 150 µL DMSO (Sigma-Aldrich) was added to solubilize the formed crystals. The optical density (OD) value of each well was detected at 570 nm on a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Cell Cycle Analysis
Cell cycle analysis was performed using the CycleTEST PLUS DNA Reagent Kit (BD Biosciences, San Jose, CA, USA). Cells were harvested, washed with PBS and stained with propidium iodide solution for 30 mins at room temperature. Then the cells were subjected to FACScan flow cytometer (BD Biosciences).
Transwell Assay
Cells suspended in 200 μL of serum-free medium were seeded into the uncoated (for migration) or Matrigel-coated (for invasion) upper chamber of transwell (8 μm pore size; Corning Inc., Corning, NY, USA). Medium (600 µL) containing 10% FBS was added to the lower chamber. Following incubation for 24 hrs, the cells on the lower membrane surface were fixed by 4% paraformaldehyde and stained with 0.1% crystal violet. Cell number was counted in five randomly selected fields.
Western Blot Analysis
Cells were collected and lysed with RIPA lysis buffer (Beyotime, Shanghai, China). Equal amounts of protein samples were separated by SDS-polyacrylamide gel electrophoresis and then transferred onto polyvinylidene fluoride (PVDF) membranes (GE Healthcare, Piscataway, NJ, USA). After blocking in 5% nonfat milk for 1 hr, the membranes were incubated with the specific primary antibodies overnight at 4°C. The membranes were then incubated with HRP-conjugated secondary antibody for 2 hrs at room temperature. After washes, the bands were visualized using an enhanced chemiluminescence (ECL) substrate kit (Amersham Biosciences, Little Chalfont, UK), and GAPDH was used as the loading control.
Dual-Luciferase Reporter Assay
The fragment from HCP5 or SOX4 containing the predicted miR-140-5p binding site was amplified by PCR and cloned into the psiCHECK2 vector (Promega, Madison, WI, USA). HEK293T cells were seeded into 24-well plates and co-transfected with the recombinant vectors and miR-140-5p mimics or NC using Lipofectamine 2000. After 48 hrs, the cells were lysed and the luciferase activity was detected using the Dual-Luciferase Reporter Assay System (Promega).
Statistical Analysis
Statistical analyses were performed using GraphPad Prism version 6.0 (GraphPad Software, Inc., San Diego, CA, USA) and Statistical Package for Social Sciences version 16.0 (SPSS Inc., Chicago, IL, USA). Experimental data are expressed as the mean ± standard deviation (SD), and the significance of differences between groups was estimated using Student’s t-test or one-way analysis of variance (ANOVA) analysis. Survival curves were generated by Kaplan–Meier analysis and compared using log rank test. Values of P<0.05 were considered to indicate a statistically significant result.
Results
HCP5 Is Overexpressed In OSCC
Through RT-qPCR analysis, we observed that, compared with adjacent normal tissues, HCP5 expression was remarkably increased in OSCC tissues (Figure 1A). Moreover, the HCP5 expression in the human OSCC cell lines (SCC-4, Tca8113 and SCC-9) was also significantly higher than that of NOK cells (Figure 1B).
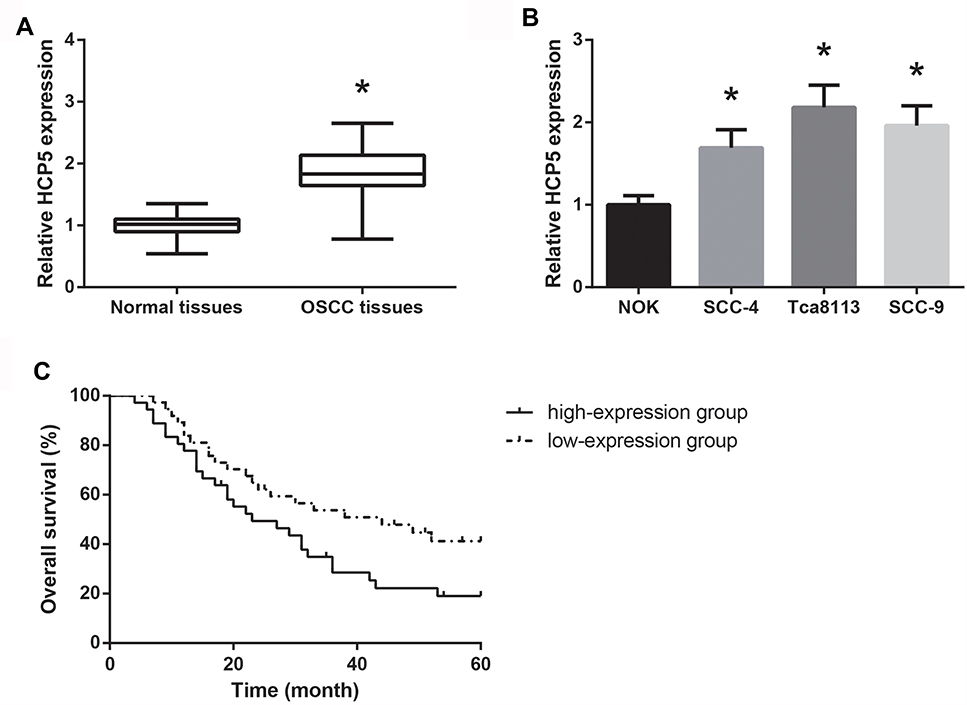 |
Figure 1 HCP5 is overexpressed in OSCC. (A) RT-qPCR analysis of HCP5 expression levels in clinical OSCC tissues. (B) RT-qPCR analysis of HCP5 expression levels in a series of OSCC cell lines. (C) Kaplan–Meier analysis of the association between HCP5 expression and overall survival in OSCC patients. Data are presented as mean ± SD. *P<0.05 versus normal tissues or NOK cells. |
According to the HCP5 expression in OSCC tissues, the patients were then allocated into two groups, including the high-expression group (n=36) and the low-expression group (n=37). As summarized in Table 1, high expression of HCP5 was significantly associated with lymph node metastasis (P=0.025) and advanced TNM stage (P=0.003) of OSCC patients. Moreover, as shown in Figure 1C, Kaplan–Meier analysis showed that high HCP5 expression was also closely associated with the unfavorable overall survival in OSCC patients (P=0.044).
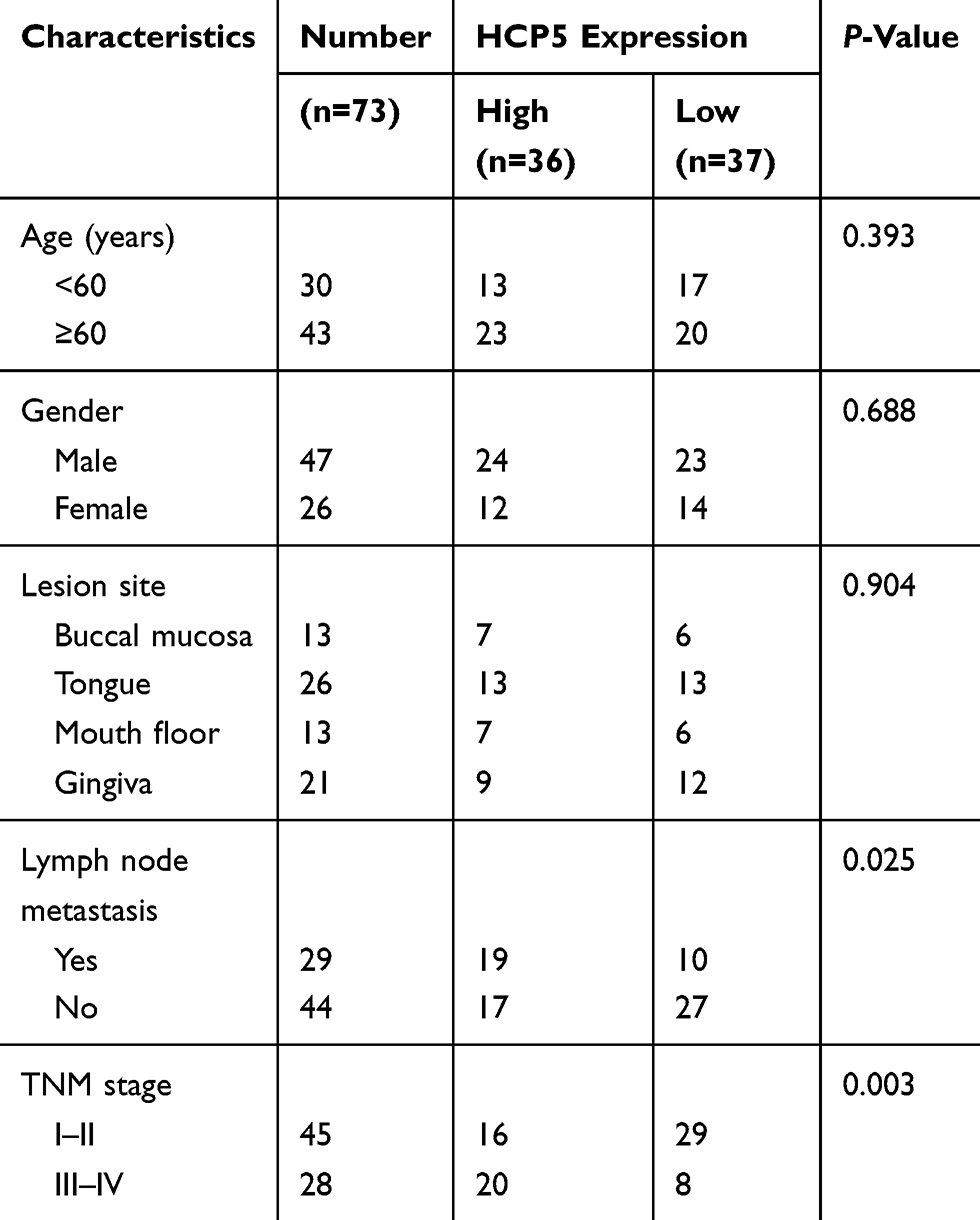 |
Table 1 Relationship Between HCP5 Expression And Clinicopathological Characteristics Of OSCC Patients |
HCP5 Promotes OSCC Cell Proliferation And Cell Cycle Progression
We next investigated the regulatory roles of HCP5 in OSCC cell phenotypes. As confirmed by RT-qPCR analysis, after transfection with si-HCP5, HCP5 was effectively knocked-down in Tca8113 cells (Data not shown). Besides, we also found HCP5 was overexpressed in SCC-4 cells transfected with pcDNA3.1-HCP5. Through MTT assay, it was shown that HCP5 overexpression significantly increased the proliferation rate of SCC-4 cells; in contrast, Tca8113 cells transfected with si-HCP5 grew notably slower (Figure 2A). Moreover, as demonstrated in Figure 2B, HCP5 knockdown significantly induced G0/G1 cell arrest in Tca8113 cells, whereas HCP5 overexpression decreased the number of SCC-4 cells in G0/G1 phase.
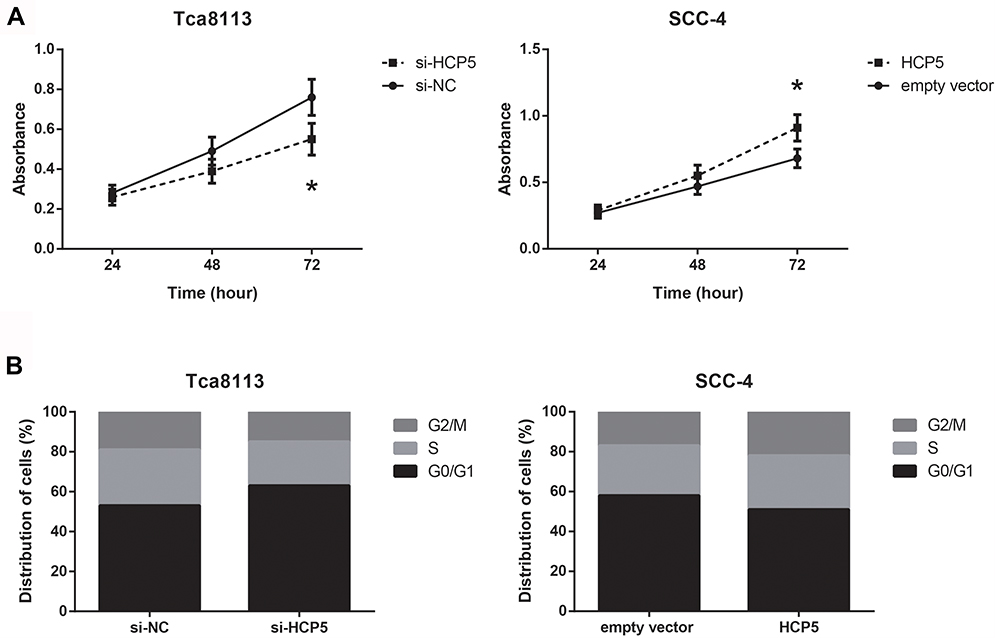 |
Figure 2 HCP5 promotes OSCC cell proliferation and cell cycle progression. (A) Proliferation of OSCC cells after transfection, detected by MTT assay. (B) Flow cytometric analysis of cell cycle distributions in OSCC cells after transfection. Data are presented as mean ± SD. *P<0.05 versus si-NC or empty vector-transfected cells. |
HCP5 Promotes OSCC Cell Migration, Invasion And EMT
Furthermore, as revealed by transwell assay, the capacities of cell migration and invasion were significantly impaired in Tca8113 cells transfected with si-HCP5. On the other hand, HCP5 overexpression markedly enhanced the migration and invasion of SCC-4 cells (Figure 3A). EMT is of key importance for migration and invasion of OSCC cells. We then detected the expression levels of EMT-related proteins by Western blot analysis, and the results indicated that HCP5 knockdown increased the level of E-cadherin, but decreased the levels of N-cadherin and Vimentin in Tca8113 cells (Figure 3B). HCP5 overexpression notably enhanced EMT in SCC-4 cells.
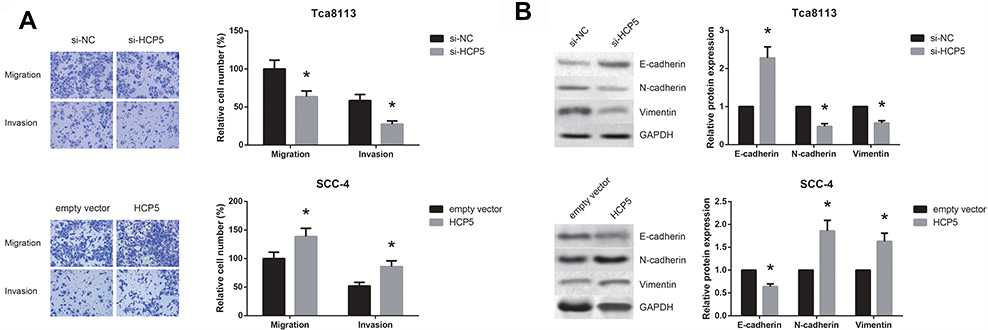 |
Figure 3 HCP5 promotes OSCC cell migration, invasion and EMT. (A) Migration and invasion of OSCC cells after transfection, detected by transwell assay. (B) Western blot analysis of EMT-related proteins in OSCC cells after transfection. Data are presented as mean ± SD. *P<0.05 versus si-NC or empty vector-transfected cells. |
HCP5 Acts As A ceRNA For miR-140-5p In OSCC
Through the Starbase online software (http://starbase.sysu.edu.cn/index.php), we searched for the potential target miRNAs of HCP5, and found that miR-140-5p had putative HCP5 binding sites (Figure 4A). Then, dual-luciferase reporter assay was carried out to further verify the bioinformatical prediction. The results showed that in both Tca8113 and SCC-4 cells, co-transfection with miR-140-5p mimics markedly decreased the luciferase activity of HCP5-WT, rather than HCP5-MUT (Figure 4B). The downregulation of miR-140-5p was observed in OSCC tissues and cell lines (Figure 4C and D). There is a negative correlation between HCP5 expression and miR-140-5p in OSCC tissues (Figure 4E). miR-140-5p expression was decreased in SCC-4 cells with HCP5 overexpression, while it was increased in si-HCP5-transfected Tca8113 cells (Figure 4F).
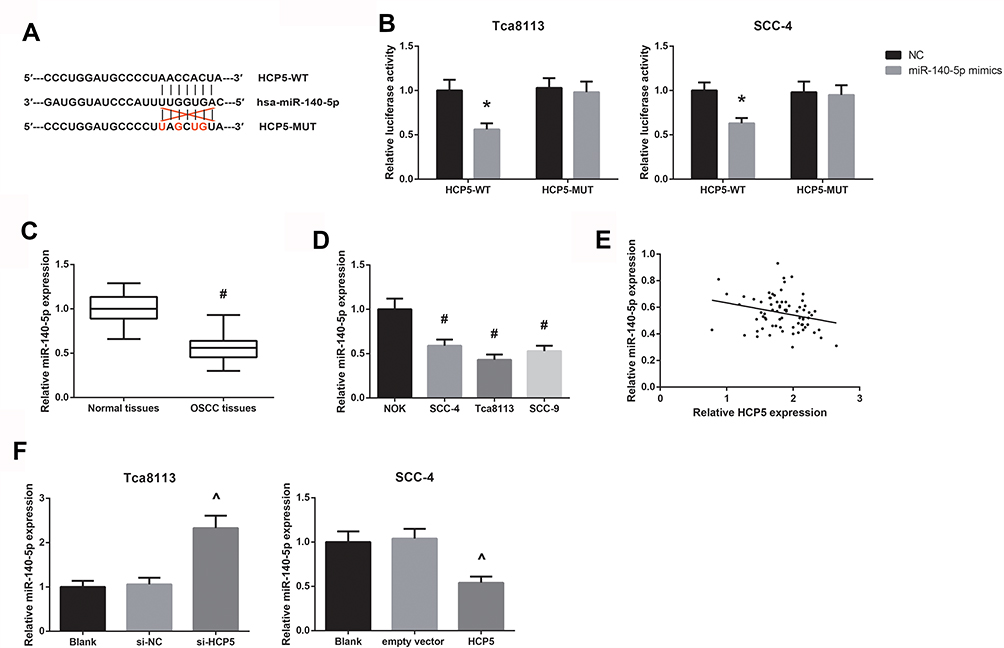 |
Figure 4 HCP5 acts as a ceRNA for miR-140-5p in OSCC. (A) The potential binding sites between miR-140-5p and HCP5. (B) The luciferase activities in OSCC cells co-transfected with HCP5-WT or HCP5-MUT reporter vectors and miR-140-5p mimics or NC. (C) RT-qPCR analysis of miR-140-5p expression levels in clinical OSCC tissues. (D) RT-qPCR analysis of miR-140-5p expression levels in a series of OSCC cell lines. (E) The inverse correlation between miR-140-5p and HCP5 expression in OSCC tissues. (F) RT-qPCR analysis of miR-140-5p expression levels in OSCC cells after transfection. Data are presented as mean ± SD. *P<0.05 versus NC-transfected cells; #P<0.05 versus normal tissues or NOK cells; ^P<0.05 versus si-NC or empty vector-transfected cells. |
miR-140-5p Targets SOX4 In OSCC
Next, through the TargetScan online software (http://www.targetscan.org/vert_71/), we observed that SOX4 might contain a miR-140-5p binding site in its 3′-UTR (Figure 5A). In addition, as shown in Figure 5B, the luciferase activity was significantly reduced in both Tca8113 and SCC-4 cells following co-transfection with SOX4-WT and miR-140-5p mimics. Moreover, the increased SOX4 level in HCP5-overexpressing SCC-4 cells was diminished by miR-140-5p restoration (Figure 5C).
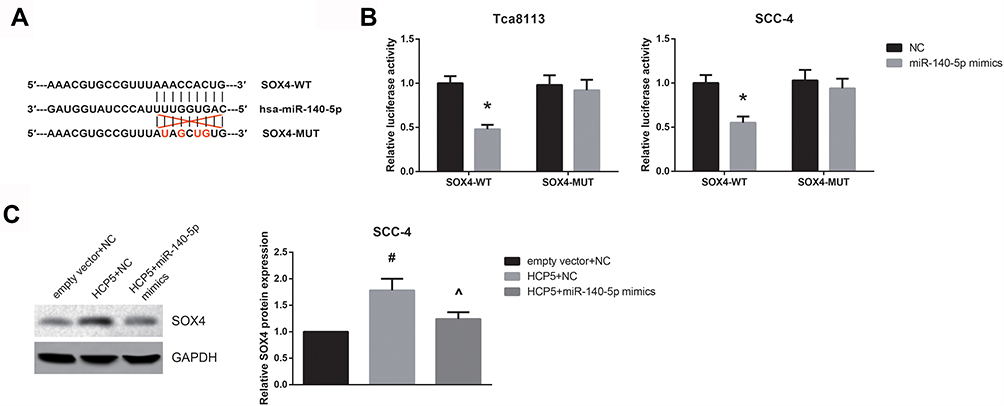 |
Figure 5 miR-140-5p targets SOX4 in OSCC. (A) The potential binding sites between miR-140-5p and SOX4. (B) The luciferase activities in OSCC cells co-transfected with SOX4-WT or SOX4-MUT reporter vectors and miR-140-5p mimics or NC. (C) Western blot analysis of SOX protein expression levels in SCC-4 cells after transfection. Data are presented as mean ± SD. *P<0.05 versus NC-transfected cells; #P<0.05 versus empty vector+NC-transfected cells; ^P<0.05 versus HCP5+NC-transfected cells. |
miR-140-5p Restoration Blocks The Oncogenic Role Of HCP5 In OSCC Cells
Then, rescue experiments were performed to determine whether HCP5 regulated OSCC cell phenotypes in a miR-140-5p-dependent manner. As shown in Figure 6A, miR-140-5p restoration counteracted the effect of HCP5 in promoting the EMT of SCC-4 cells. Moreover, in transwell assay, the migration and invasion of HCP5-overexpressing SCC-4 cells were markedly inhibited by co-transfection with miR-140-5p mimics (Figure 6B).
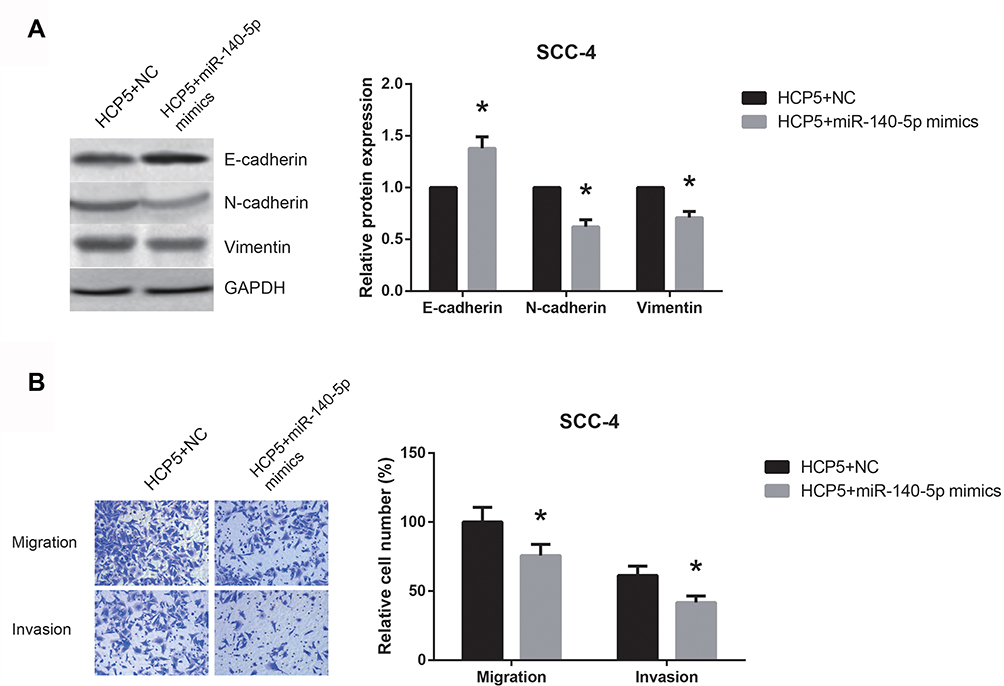 |
Figure 6 miR-140-5p restoration blocks the oncogenic role of HCP5 in OSCC cells. (A) Western blot analysis of EMT-related proteins in SCC-4 cells after transfection. (B) Migration and invasion of SCC-4 cells after transfection, detected by transwell assay. Data are presented as mean ± SD. *P<0.05 versus HCP5+NC-transfected cells. |
Discussion
OSCC develops by the accumulation of molecular and genetic alterations. In recent years, lots of lncRNAs have been discovered. Some of them are abnormally expressed and have multiple effects in all steps of OSCC progression.8 LncRNAs hold promise as prospective novel diagnostic indicators and therapeutic targets for OSCC patients. The oncogenic role of HCP5 has been previously reported in many human cancers, and this study, for the first time, reported novel biological effects of HCP5 in OSCC.
Our results demonstrated that HCP5 was overexpressed in OSCC, and OSCC patients with aggressive clinicopathological characteristics and poor prognosis exhibited higher intratumoral HCP5 expression. Dysregulation of cell proliferation, migration and invasion is a critical trigger of carcinogenesis. Our functional experiments further verified that HCP5 served as an oncogenic regulator in OSCC, and knockdown of HCP5 diminished the malignant traits of OSCC cells. Metastasis is a main cause of cancer-related deaths, and epithelial-mesenchymal transition (EMT), resulting in enhanced cell migration and invasion, is a key process in cancer metastasis.9 EMT is characterized by a loss of epithelial markers, including E-cadherin, and the acquisition of mesenchymal markers, including N-cadherin and Vimentin.10 Some EMT markers have significant prognostic impact for OSCC patients.11 In this study, we confirmed that HCP5 activates EMT in OSCC, consistent with its role in lung adenocarcinoma and colorectal cancer.12,13
microRNAs (miRNAs), another kind of noncoding RNA, are 19–25 nucleotides in length. The ceRNA hypothesis, originally proposed by Pandolfi et al, indicates that lncRNAs can function as a competing endogenous RNAs (ceRNAs) that sequesters miRNAs to block the repression of miRNAs on target mRNAs.14 Peng et al previously reported that miR-140-5p suppressed OSCC tumorigenesis,15 and in this study, we demonstrated that HCP5 directly binds to miR-140-5p and inhibits miR-140-5p expression in OSCC. As a critical transcription factor in EMT, SOX4 is closely associated with lymph node metastasis in OSCC patients,16 and here, it was verified as a direct target of miR-140-5p in OSCC. From rescue experiments, we found that miR-140-5p restoration blocked SOX4-mediated EMT, thereby reducing the oncogenic effects of HCP5 in OSCC cells.
In short, our findings illustrated that HCP5 might serve as an oncogene to facilitate OSCC progression partly through regulating miR-140-5p/SOX4 axis. The discovery of this regulation pattern was meaningful, and we therefore think that HCP5 may represent a potential therapeutic target for OSCC treatment in the future.
Acknowledgment
This work was supported by Research Project on Basic Scientific Research Business Funds of Provincial Colleges and Universities in Heilongjiang Province (2017-KYYWFMY-0686).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rivera C.Essentials of oral cancer. Int J Clin Exp Pathol. 2015;8(9):11884–11894. Epub 2015/12/01.
2. Warnakulasuriya S. Global epidemiology of oral and oropharyngeal cancer. Oral Oncol. 2009;45(4–5):309–316. Epub 2008/09/23. doi:10.1016/j.oraloncology.2008.06.002
3. Renganathan A, Felley-Bosco E. Long noncoding RNAs in cancer and therapeutic potential. Adv Exp Med Biol. 2017;1008:199–222. Epub 2017/08/18.
4. Liang L, Xu J, Wang M, et al. LncRNA HCP5 promotes follicular thyroid carcinoma progression via miRNAs sponge. Cell Death Dis. 2018;9(3):372. Epub 2018/03/09. doi:10.1038/s41419-018-0382-7
5. Zhao W, Li L. SP1-induced upregulation of long non-coding RNA HCP5 promotes the development of osteosarcoma. Pathol Res Pract. 2019;215(3):439–445. Epub 2018/12/18. doi:10.1016/j.prp.2018.12.006
6. Wang L, Luan T, Zhou S, et al. LncRNA HCP5 promotes triple negative breast cancer progression as a ceRNA to regulate BIRC3 by sponging miR-219a-5p. Cancer Med. 2019;8(9):4389–4403. Epub 2019/06/20. doi:10.1002/cam4.v8.9
7. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–408. Epub 2002/02/16. doi:10.1006/meth.2001.1262
8. Zhang L, Meng X, Zhu XW, et al. Long non-coding RNAs in oral squamous cell carcinoma: biologic function, mechanisms and clinical implications. Mol Cancer. 2019;18(1):102. Epub 2019/05/28. doi:10.1186/s12943-019-1021-3
9. Savagner P .The epithelial-mesenchymal transition (EMT) phenomenon. Ann Oncol.2010;21 Suppl 7:vii89–vii92. Epub 2010/10/15. doi:10.1093/annonc/mdq292
10. Acloque H, Adams MS, Fishwick K, Bronner-Fraser M, Nieto MA. Epithelial-mesenchymal transitions: the importance of changing cell state in development and disease. J Clin Invest. 2009;119(6):1438–1449. Epub 2009/06/03. doi:10.1172/JCI38019
11. da Silva SD, Morand GB, Alobaid FA, et al. Epithelial-mesenchymal transition (EMT) markers have prognostic impact in multiple primary oral squamous cell carcinoma. Clin Exp Metastasis. 2015;32(1):55–63. Epub 2014/12/01. doi:10.1007/s10585-014-9690-1
12. Jiang L, Wang R, Fang L, et al. HCP5 is a SMAD3-responsive long non-coding RNA that promotes lung adenocarcinoma metastasis via miR-203/SNAI axis. Theranostics. 2019;9(9):2460–2474. Epub 2019/05/28. doi:10.7150/thno.31097
13. Yang C, Sun J, Liu W, et al. Long noncoding RNA HCP5 contributes to epithelial-mesenchymal transition in colorectal cancer through ZEB1 activation and interacting with miR-139-5p. Am J Transl Res. 2019;11(2):953–963. Epub 2019/03/23.
14. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146(3):353–358. Epub 2011/08/02. doi:10.1016/j.cell.2011.07.014
15. Peng M, Pang C. MicroRNA-140-5p inhibits the tumorigenesis of oral squamous cell carcinoma by targeting p21-activated kinase 4. Cell Biol Int. 2019. Epub 2019/08/09. doi:10.1002/cbin.11213
16. Watanabe M, Ohnishi Y, Wato M, Tanaka A, Kakudo K. SOX4 expression is closely associated with differentiation and lymph node metastasis in oral squamous cell carcinoma. Med Mol Morphol. 2014;47(3):150–155. Epub 2013/09/14. doi:10.1007/s00795-013-0057-5
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.