Back to Journals » OncoTargets and Therapy » Volume 13
Long Non-Coding RNA HAND2-AS1 Inhibits Growth and Migration of Gastric Cancer Cells Through Regulating the miR-590-3p/KCNT2 Axis
Authors Yu L, Li H, Li Z, Jia J, Wu Z, Wang M, Li F, Feng Z, Xia H ![]() , Gao G
, Gao G
Received 2 October 2019
Accepted for publication 16 February 2020
Published 16 April 2020 Volume 2020:13 Pages 3187—3196
DOI https://doi.org/10.2147/OTT.S233256
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Federico Perche
This paper has been retracted.
Lan Yu, 1 Hui Li, 2 Zhiying Li, 1 Jianchao Jia, 1 Zhouying Wu, 1 Min Wang, 1 Feng Li, 1 Zongqi Feng, 1 Huilin Xia, 3 Guanxin Gao 3
1Clinical Medical Research Center, Inner Mongolia People’s Hospital, Hohhot 010017, People’s Republic of China; 2Department of Oncology, Inner Mongolia People’s Hospital, Hohhot 010017, People’s Republic of China; 3Medical Engineering Department, Inner Mongolia People’s Hospital, Hohhot 010017, People’s Republic of China
Correspondence: Huilin Xia; Guanxin Gao
Medical Engineering Department, Inner Mongolia People’s Hospital, No. 20 Zhaowuda Road, Saihan District, Hohhot 010017, People’s Republic of China
Email [email protected]; [email protected]
Introduction: Long non-coding RNAs (lncRNAs) are regarded as crucial regulators for cancer initiation and progression. Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1 (HAND2-AS1) was recently proposed to function as tumor suppressor in several human cancers. However, its role in gastric cancer (GC) remains unclear.
Methods: HAND2-AS1 expression in GC tissues and normal tissues was analyzed at GEPIA (a web server for gene expression profiling analysis). Moreover, RT-qPCR method was utilized to explore HAND2-AS1 expression in GC cells and normal cell. In vitro experiments were carried out using cell counting kit-8 assay, colony formation assay, and flow cytometry assay, respectively. Bioinformatic analysis and luciferase activity reporter assay were performed to identify the downstream targets of HAND2-AS1.
Results: We found HAND2-AS1 has decreased expression in both GC tissues and cells. Overexpression of HAND2-AS1 was able to inhibit GC cell proliferation, colony formation, but promote apoptosis. On the contrary, knockdown of HAND2-AS1 could cause the opposite effects on GC cells. Furthermore, HAND2-AS1 was shown to function as a competitive RNA that binds with microRNA-590-3p (miR-590-3p) to affect the expression of potassium sodium-activated channel subfamily T member 2 (KCNT2).
Discussion: Our results indicated the tumor suppressive role of HAND2-AS1 in GC. Also, the newly identified HAND2-AS1/miR-590-3p/KCNT2 axis will help us to understand the role of HAND2-AS1 in cancer.
Keywords: HAND2-AS1, miR-590-3p, KCNT2, gastric cancer
Introduction
Among all the cancer types in the digestive system, gastric cancer (GC) is one of the leading causes of newly occurring cancer types each year.1 As predicted, the numbers for newly diagnosed and cancer deaths each year are 1,033,071 and 782,685, respectively.1 The 5-year overall survival of GC patients receiving treatment at early stages can be about 95% owing to the improvements in surgery and targeted therapy methods.2 However, for patients diagnosed at late stages, the best treatment window is closed.
Long noncoding RNAs (lncRNAs) are a family of RNAs with lengths of 200 nucleotides to 100 kilobases.3 LncRNAs have typically been regarded as junk genes as they lack the ability to code proteins.3 In 2011, Salmena et al4 proposed a competitive RNA (ceRNA) theory that helped us to understand the functions of non-coding RNAs including lncRNA.
In recent years, investigations of the biological roles of lncRNAs in disease progression, especially in cancers, have been the hotspots.5 Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1 (HAND2-AS1), located at chromosome 4q33-34, has been demonstrated to be a crucial regulator for cancer progression in the past 2 years.6–9 In endometrial carcinoma, HAND2-AS1 was found to have decreased expression in tumor tissues.6 Moreover, they found HAND2-AS1 overexpression could suppress cancer cell migration and invasion via inactivating neuromedin U 6. In colorectal cancer, downregulation of HAND2-AS1 was found to have a negative correlation with tumor stages.7 Also, they showed that the proliferation and invasion abilities of colorectal cancer cells can be suppressed by HAND2-AS1 overexpression.7 A recent work performed on non-small cell lung cancer showed HAND2-AS1 can suppress cancer cell malignant behaviors with transforming growth factor β as a mediator.8 Importantly, a similar expression trend of HAND2-AS1 was found in esophagus squamous cell carcinoma.9 However, until now, the expression and functions of HAND2-AS1 in GC remains to be elucidated.
In this study, we explored the expression of HAND2-AS1 in both GC tissues and cell lines. Also, the effects of HAND2-AS1 on GC cell behaviors were explored using in vitro experiments. Importantly the possible involvement of microRNA-590-3p (miR-590-3p) and potassium sodium-activated channel subfamily T member 2 (KCNT2) in the HAND2-AS1-mediated GC cell events was investigated.
Materials and Methods
Cell Lines and Cell Culture
Normal gastric mucosal epithelial cell (GES-1) and GC cells (SGC-7901 and BGC-823) were bought from the Cell Collection Center of Chinese Academy of Sciences (Shanghai, People's Republic of China). RPMI 1640 with 10% fetal bovine serum (Invitrogen, Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplement was used to incubate these cells at a 37°C humidified incubator supplemented with 5% of CO2.
Cell Transfection
Small interfering RNA targeting HAND2-AS1 (si-HAND2-AS1), negative control (si-NC), miR-590-3p mimic, and the corresponding negative control (mi-NC) were provided by GenePharm (Shanghai, People's Republic of China). The pcDNA3.1 with open reading frame of HAND2-AS1 or KCNT2 inserted was bought from Generay (Shanghai, People's Republic of China). These siRNAs, miRNAs, or pcDNAs were transfected into GC cells using Lipofectamine 2000 (Invitrogen) after incubating these cells to about 60% of confluence.
Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
RNA from cells were prepared using Trizol reagent (Invitrogen) according to the provided protocols. Complementary DNA was synthesized using PrimerScript reagent kit (Invitrogen). RT-qPCR was performed at ABIT 7500 system (Applied Biosystems, Thermo Fisher Scientific, Inc., Waltham, MA, USA) to detect the relative expression level of HAND2-AS1, miR-590-3p, and KCNT2 using SYBR Green Mix (Takara, Dalian, Liaoning, People's Republic of China). The primers used were as follows: HAND2-AS1: 5ʹ-GGGTGTTTACGTAGACCAGAACC-3ʹ (forward) and 5ʹ-CTTCCAAAAGCCTTCTGCCTTAG-3ʹ (reverse); KCNT2: 5ʹ-TGCCTCCCAGGTACAGATTCCGTGAT-3ʹ (forward) and 5ʹ-TTGTTTCAAATAGACTTATCAATGCCACCGAGA-3ʹ (reverse); β-actin: 5ʹ-GACCTCTATGCCAACACAGT-3ʹ (forward) and 5ʹ-AGTACTTGCGCTCAGGAGGA-3ʹ (reverse); miR-590-3p: 5ʹ-GCGCTAATTTTATGTATAA-3ʹ (forward) and 5ʹ-GTGCAGGGTCCGAGGT-3ʹ (reverse); U6 snRNA: 5ʹ-AGAGCCTGTGGTGTCCG-3ʹ (forward) and 5ʹ-CATCTTCAAAGCACTTCCCT-3ʹ (reverse). β-actin was used as internal control for HAND2-AS1 and KCNT2, while U6 snRNA was used as endogenous control for miR-590-3p.
Cell Counting Kit-8 (CCK-8) Assay
Cells were incubated into 96-well plates with the density of 5,000 cells/well. After 0, 24, 48, and 72 hours of incubation, CCK-8 reagent obtained from Beyotime (Haimen, Jiangsu, People's Republic of China) was added to each well and further incubated for 2 hours. Microplate reader was used to measure optical density at 450 nm.
Colony Formation Assay
Cells were seeded into 6-well plates with the density of 1,000 cells/well and incubated at the above-mentioned condition for 2 weeks to form colonies. Subsequently, colonies formed were fixed, stained, and counted under a microscope.
Cell Apoptosis Assay
The cultured cells were treated using trypsin, and then stained with Annexin V Fluorescein Isothiocyanate (FITC)/propidium iodide (PI) at room temperature for 15 minutes obtained from Beyotime. Cell apoptosis rate was analyzed at flow cytometry (BD Biosciences, San Jose, MA, USA). The cells with FITC staining were regarded as apoptosis cells.
Targets Prediction Using Bioinformatic Analysis Tools
The miRNA target for HAND2-AS1 was analyzed at LncBase Predicted v.2 (http://carolina.imis.athena-innovation.gr/diana_tools/) and miR-590-3p was found to be a putative target. The targets for miR-590-3p were analyzed at TargetScan V7.2 (http://www.targetscan.org/vert_72/) and KCNT2 was identified as a highly potential target for miR-590-3p.
Luciferase Activity Reporter Assay
According to the bioinformatic analysis results, luciferase activity vectors were constructed using pmirGLO vector (Promega, Madison, WI, USA). The wild-type and mutant sequences of HAND2-AS1 and KCNT2 were cloned into pmirGLO and named as wt/mt HAND2-AS1 or KCNT2, respectively. Cells were co-transfected with luciferase activity vectors and synthetic miRNAs using Lipofectamine 2000. A dual-luciferase activity system was used to measure relative luciferase activity.
RNA Immunoprecipitation (RIP)
RIP assay was utilized to investigate the connection of HAND2-AS1 and miR-590-3p using Magna RI RNA-Binding Protein Immunoprecipitation Kit (Millipore, Sigma Aldrich, Merck, Billerica, MA, USA). RIP buffer was used to incubate cultured cells and treated with anti-Ago2 antibody and control IgG. Samples were then treated with Proteinase K and subjected to RT-qPCR analysis to analyze HAND2-AS1 and miR-590-3p levels.
RNA Pull-Down Assay
Cells were transfected with biotin-labeled miR-590-3p-wt and miR-590-3p-mt (bio-miR-590-3p-wt/mt). Then, cells were treated with lysis buffer and incubated with streptavidin magnetic beads (Sigma, St Louis, MO, USA) and rinsed with pre-cold lysis buffer and salt buffer. Trizol was used to extract the bound RNA and then detect the expression of HAND2-AS1 using RT-qPCR.
Gene Expression Level Exploration in GC Tissues at GEPIA or StarBase
Expression of HAND2-AS1 in GC tissues and normal tissues was analyzed at GEPIA (http://gepia2.cancer-pku.cn/#index).10 Expression of miR-590-3p, and KCNT2 in GC tissues and normal tissues was analyzed at StarBase (http://starbase.sysu.edu.cn/).11
Statistical Analysis
SPSS 21.0 software (IBM Corporation, Armonk, NY, USA) was used for data analysis. Data were presented as mean ± SD. Differences in groups were analyzed with Student’s t-test and one-way ANOVA with Tukey post-hoc test. A p-value less than 0.05 was considered as statistically significant.
Results
Expression of HAND2-AS1 in GC
We first explored the expression of HAND2-AS1 in GC tissues and normal tissues. As presented in Figure 1A, we found HAND2-AS1 expression level was significantly lower in GC tissues than in normal tissues. Furthermore, a similar result was observed in GC cells. As expected, we revealed that HAND2-AS1 was downregulated in GC cells compared with normal cell line (Figure 1B).
 |
Figure 1 Downregulation of HAND2-AS1 was found in GC. Notes: Downregulation of HAND2-AS1 was found in (A) GC tissues and normal tissues, and (B) GC cell lines and a normal cell line. *P < 0.05; ***P < 0.001. Abbreviations: HAND2-AS1, Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1; GC, gastric cancer. |
Overexpression of HAND2-AS1 Inhibits GC Cell Proliferation and Invasion but Promotes Apoptosis
The upregulation of HAND2-AS1 in GC cells transfected with pHAND2-AS1 transfection was explored by RT-qPCR (Figure 2A). CCK-8 assay was utilized to explore cell proliferation rate with pHAND2-AS1 transfection. We found the overexpression of HAND2-AS1 decreased cell proliferation ability of GC cells (Figure 2B). Colony formation assay was conducted to evaluate colony formation ability, and the results are presented in Figure 2C, which revealed colony formation ability was significantly suppressed by pHAND2-AS1. Moreover, flow cytometry assay revealed that cell apoptosis ability was significantly enhanced by pHAND2-AS1 transfection (Figure 2D).
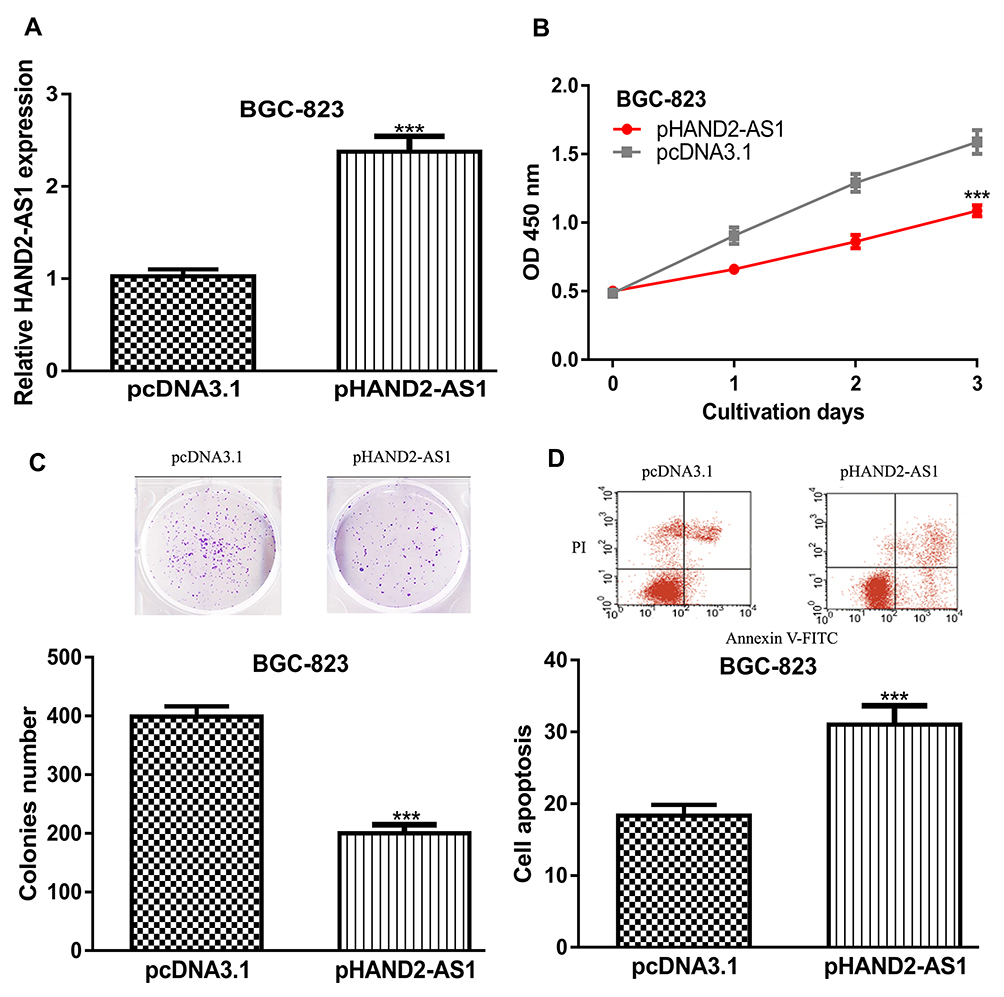 |
Figure 2 HAND2-AS1 overexpression inhibits GC cell proliferation and colony formation but promotes cell apoptosis. Notes: (A) HAND2-AS1 expression, (B) Cell proliferation, (C) Colony formation, and (D) Cell apoptosis in GC cell with pHAND2-AS1 or pcNA3.1 transfection. ***P < 0.001. Abbreviations: HAND2-AS1, Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1; GC, gastric cancer. |
Knockdown of HAND2-AS1 Promotes GC Cell Proliferation and Invasion but Inhibits Apoptosis
Furthermore, loss-of-function experiments were performed to fully understand the role of HAND2-AS1 in GC. The introduction of si-HAND2-AS1 significantly decreased the expression level of HAND2-AS1 in GC cell compared with si-NC (Figure 3A). CCK-8 assay and colony formation assay revealed that cell proliferation ability and colony formation ability were significantly inhibited by si-HAND2-AS1 (Figure 3B and C). Furthermore, flow cytometry assay revealed that si-HAND2-AS1 transfection could inhibit cell apoptosis (Figure 3D).
 |
Figure 3 HAND2-AS1 knockdown promotes GC cell proliferation and colony formation but inhibits cell apoptosis. Notes: (A) HAND2-AS1 expression, (B) Cell proliferation, (C) Colony formation, and (D) Cell apoptosis in GC cell with si-HAND2-AS1 or si-NC transfection. ***P < 0.001. Abbreviations: HAND2-AS1, Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1; GC, gastric cancer; si-NC, negative control small interfering RNA. |
miR-590-3p Was a Direct Target for HAND2-AS1
The ceRNA theory suggested that lncRNA can function as a miRNA sponge to regulate miRNA expression. Hence, we analyzed the potential miRNA target for HAND2-AS1 using LncBase Predicted v.2, and we found miR-590-3p may bind with HAND2-AS1 (Figure 4A). Luciferase activity reporter assay showed miR-590-3p mimic transfection decreased luciferase activity in GC cells with wt-HAND2-AS1 transfection (Figure 4B). RIP assay showed HAND2-AS1 and miR-590-3p was enriched in anti-Ago2 groups compared with anti-IgG groups (Figure 4C). RNA pull-down assay showed HAND2-AS1 enrichment was significantly increased in bio-miR-590-3p-wt group (Figure 4D). Then, we explored the expression of miR-590-3p in GC tissues and cells. We showed that miR-590-3p expression level was significantly upregulated in both GC tumor tissues and cell lines compared with the normal tissues and cell line (Figure 4E and F).
 |
Figure 4 HAND2-AS1 direct bind with miR-590-3p. Notes: (A) The binding site between HAND2-AS1 and miR-590-3p. (B) Luciferase activity reporter assay revealed the relative luciferase activity in cells with wt/mt HAND2-AS1 or miR-590-3p mimic/mi-NC transfection. (C) RNA immunoprecipitation analysis for the enrichment of HAND2-AS1 and miR-590-3p in anti-Ago2 and anti-IgG groups. (D) HAND2-AS1 binding to miR-590-3p analyzed by RNA pull-down assay. Expression of miR-590-3p in (E) GC tissues and normal tissues, and (F) GC cell lines and a normal cell line. ***P < 0.001. Abbreviations: HAND2-AS1, Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1; GC, gastric cancer; miR-590-3p, microRNA-590-3p; wt, wild-type; mt, mutant; mi-NC, negative control miRNA; Ago2, Argonaute 2. |
KCNT2 Was a Direct Target for miR-590-3p
Subsequently, we analyzed the targets for miR-590-3p using TargetScan, and we found KCNT2 was a putative target for miR-590-3p (Figure 5A). Dual-luciferase activity reporter assay revealed that overexpression of miR-590-3p inhibited luciferase activity in GC cells with wt-KCNT2 expression (Figure 5B). Moreover, expression level of KCNT2 in GC tissues and cells was further explored. We showed KCNT2 expression level was significantly decreased in GC tissues and cell lines (Figure 5C and D).
 |
Figure 5 KCNT2 was a direct target for miR-590-3p. Notes: (A) The binding site between KCNT2 and miR-590-3p. (B) Luciferase activity reporter assay revealed the relative luciferase activity in cells with wt/mt KCNT2 or miR-590-3p mimic/mi-NC transfection. Expression of KCNT2 in (C) GC tissues and normal tissues, and (D) GC cell lines and a normal cell line. **P < 0.01; ***P < 0.001. Abbreviations: KCNT2, potassium sodium-activated channel subfamily T member 2; GC, gastric cancer; miR-590-3p, microRNA-590-3p; wt, wild-type; mt, mutant; mi-NC, negative control miRNA. |
HAND2-AS1 Regulates GC Cell Behaviors via Regulating miR-590-3p/KCNT2 Axis
At length, rescue experiments were performed to analyze whether HAND2-AS1 regulates GC cell behaviors via regulating the miR-590-3p/KCNT2 axis. We showed KCNT2 expression was enhanced by pKCNT2 but decreased by miR-590-3p mimic (Figure 6A). CCK-8 assay and colony formation assay revealed that cell proliferation ability and colony formation ability of GC cells were decreased by pKCNT2 but increased by miR-590-3p mimic (Figure 6B and C). The analysis of cell apoptosis rate revealed that cell apoptosis rate was increased by pKCNT2 but decreased by miR-590-3p mimic (Figure 6D). More importantly, we showed the introduction of pHAND2-AS1 or pKCNT2 could reverse the effects of miR-590-3p on GC cells (Figure 6B–D).
 |
Figure 6 HAND2-AS1 regulates GC cell behaviors via regulating miR-590-3p/KCNT2 axis. Notes: (A) KCNT2 expression, (B) Cell proliferation, (C) Colony formation, and (D) Cell apoptosis in GC cells with miR-590-3p mimic, pKCNT2, mi-NC, pcDNA3.1+mi-NC, miR-590-3p mimic+pKCNT2, and miR-590-3p mimic+pHAND2-AS1 transfection. *P < 0.05; **P < 0.01; ***P < 0.001. Abbreviations: HAND2-AS1, Heart and Neural Crest Derivatives Expressed 2 antisense RNA 1; GC, gastric cancer; miR-590-3p, microRNA-590-3p; KCNT2, potassium sodium-activated channel subfamily T member 2; mi-NC, negative control miRNA. |
Discussion
lncRNA has been reported to be a crucial mediator for tumorigenesis and the recent studies showed that their roles are partially mediated by ceRNA cross-talk.12 In GC, the ceRNA network including lncRNA, miRNA, and message RNA was reported to play crucial roles in tumor initiation and progression.13 lncRNAs were found to be abnormally expressed in multiple human cancers, however, the role of lncRNAs in GC remains to be defined. For instance, lncRNA TRPM2 antisense RNA (TRPM2-AS) expression in GC tumor tissues was found to be elevated by ELK1, and correlated with advanced tumor stages and poorer tumor stages.14 In addition, it was revealed that TRPM2-AS could promote GC cell proliferation, migration, and invasion via regulating miR-195 and high-mobility group AT-hook 1.14 Moreover, Gan et al15 reported that lncRNA H19 had upregulated expression in both GC tissues and cells. They also reported H19 functions as an oncogenic lncRNA to promote GC progression in vitro and in vivo via targeting the miR‑22‑3p/Snail1 axis.15 Liu et al16 revealed the upregulation status of lncRNA FLVCR1-AS1 in GC and its correlation with overall survival of cancer patients. Also, FLVCR1-AS1 was found to functions as ceRNA in regulating myc expression via sponging miR-155 to promote GC cell proliferation and invasion.16
In this study, through analysis of the expression of HAND2-AS1 in GC tumor tissues and normal tissues in GEPIA, we showed HAND2-AS1 was significantly decreased in tumor tissues compared with normal tissues. Moreover, we showed that HAND2-AS1 expression was lower in GC cells than in normal cell lines. Through gain and loss-of-function experiments, we showed the overexpression of HAND2-AS1 could inhibit GC cell proliferation and colony formation but promote apoptosis, while the knockdown of HAND2-AS1 caused the opposite effects on GC cell behaviors. Subsequently, ceRNA theory was employed to investigate the mechanisms behind HAND2-AS1-mediated GC cell behaviors. We showed that HAND2-AS1 may function as an miR-590-3p sponge to regulate KCNT2 expression. miR-590-3p is a miRNA reported to have elevated expression in colorectal cancer and predict the poor overall survival of cancer patients.17 Also, miR-590-3p was demonstrated to function as an oncogenic miRNA in regulating colorectal cancer cell malignancy behaviors via targeting the Hippo pathway.17 In colon cancer, miR-590-3p was found to have elevated expression in tumor tissues and cell lines.18 Here, we have reported that miR-590-3p also had elevated expression in GC tumor tissues and cell lines. Also, we confirmed miR-590-3p was a direct target for HAND2-AS1 using dual-luciferase activity reporter assay, RIP assay, and RNA pull-down assay. Previously, we identified miR-338-5p as having decreased expression in GC stem cells, and its overexpression could inhibit cell growth.19 Also, we showed an anticancer bioactive peptide-3 could exert its biological roles via regulating miR-338-5p.19 Here, we identified miR-590-3p was also able to regulate GC cell behaviors and was regulated by upstream lncRNA to exert biological roles.
KCNT2 was revealed to contain binding sites for Nuclear Factor-κB.20 Yang et al21 recently performed a bioinformatic-based study and revealed low KCNT2 level was associated with the worse survival of patients with melanoma.21 We demonstrated that KCNT2 had decreased expression in tumor tissues and cell lines, and was a target of miR-590-3p. The roles of KCNT2 in GC development has not been reported until now. Here, we showed overexpression of KCNT2 could inhibit GC cell proliferation, colony formation and promote apoptosis. Moreover, functional assays revealed that HAND2-AS1 regulate GC cell behaviors via regulating miR-590-3p/KCNT2. Hence, we believe our work may advance our understanding of the mechanisms behind GC tumorigenesis. However, we should acknowledge the limitations of this work: 1) the lack of in vivo animal model results to strengthen the importance of HAND2-AS1/miR-590-3p/KCNT2 triplets in GC development; and 2) we did not investigate the upstream mechanism that is responsible for the elevated expression status of HAND2-AS1 in GC. There are studies showed epigenetic modifications can affect lncRNA expression in cancers,22,23 which may be a mechanism for the decreased expression of HAND2-AS1 and deserves to be deeply investigated in the future.
Conclusion
To sum up, our work presented here showed that HAND2-AS1 functions as oncogenic lncRNA to accelerate GC cell proliferation and colony formation but inhibit cell apoptosis via regulating KCNT2 through sponging miR-590-3p. Therefore, our results indicate that HAND2-AS1 may serve as a therapeutic target for GC.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This work was supported by National Natural Science Foundation of China (Grant No. 81560405), Research Foundation of Inner Mongolia People’s Hospital (Grant No. 201415), Inner Mongolia Natural Science Foundation (Grant No. 2018MS08060), Scientific Research Project Foundation of Inner Mongolia Health Commission (Grant No. 201702003), and PhD Science Foundation of Inner Mongolia People’s Hospital (Grant No. BS201805).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.v68.6
2. Song Z, Wu Y, Yang J, Yang D, Fang X. Progress in the treatment of advanced gastric cancer. Tumour Biol. 2017;39(7):1010428317714626. doi:10.1177/1010428317714626
3. Kornienko AE, Guenzl PM, Barlow DP, Pauler FM. Gene regulation by the act of long non-coding RNA transcription. BMC Biol. 2013;11(1):59. doi:10.1186/1741-7007-11-59
4. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
5. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
6. Yang X, Wang CC, Lee WYW, Trovik J, Chung TKH, Kwong J. Long non-coding RNA HAND2-AS1 inhibits invasion and metastasis in endometrioid endometrial carcinoma through inactivating neuromedin U. Cancer Lett. 2018;413:23–34. doi:10.1016/j.canlet.2017.10.028
7. Zhou J, Lin J, Zhang H, Zhu F, Xie R. LncRNA HAND2-AS1 sponging miR-1275 suppresses colorectal cancer progression by upregulating KLF14. Biochem Biophys Res Commun. 2018;503(3):1848–1853. doi:10.1016/j.bbrc.2018.07.125
8. Miao F, Chen J, Shi M, Song Y, Chen Z, Pang L. LncRNA HAND2-AS1 inhibits non-small cell lung cancer migration, invasion and maintains cell stemness through the interactions with TGF-β1. Biosci Rep. 2019;39(1):
9. Yan Y, Li S, Wang S, et al. Long noncoding RNA HAND2-AS1 inhibits cancer cell proliferation, migration, and invasion in esophagus squamous cell carcinoma by regulating microRNA-21. J Cell Biochem. 2019;120(6):9564–9571. doi:10.1002/jcb.v120.6
10. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
11. Li JH, Liu S, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42(D1):D92–D97. doi:10.1093/nar/gkt1248
12. Qi X, Zhang DH, Wu N, Xiao JH, Wang X, Ma W. ceRNA in cancer: possible functions and clinical implications. J Med Genet. 2015;52(10):710–718. doi:10.1136/jmedgenet-2015-103334
13. Guo LL, Song CH, Wang P, Dai LP, Zhang JY, Wang KJ. Competing endogenous RNA networks and gastric cancer. World J Gastroenterol. 2015;21(41):11680–11687. doi:10.3748/wjg.v21.i41.11680
14. Huang B, Chang C, Wang BL, Li H. ELK1-induced upregulation of lncRNA TRPM2-AS promotes tumor progression in gastric cancer by regulating miR-195/HMGA1 axis. J Cell Biochem. 2019;120(10):16921–16933. doi:10.1002/jcb.28951
15. Gan L, Lv L, Liao S. Long non‑coding RNA H19 regulates cell growth and metastasis via the miR‑22‑3p/Snail1 axis in gastric cancer. Int J Oncol. 2019;54(6):2157–2168. doi:10.3892/ijo.2019.4773
16. Liu Y, Guo G, Zhong Z, et al. Long non-coding RNA FLVCR1-AS1 sponges miR-155 to promote the tumorigenesis of gastric cancer by targeting c-Myc. Am J Transl Res. 2019;11(2):793–805.
17. Sun ZQ, Shi K, Zhou QB, et al. MiR-590-3p promotes proliferation and metastasis of colorectal cancer via Hippo pathway. Oncotarget. 2017;8(35):58061–58071. doi:10.18632/oncotarget.22429
18. Feng ZY, Xu XH, Cen DZ, Luo CY, Wu SB. miR-590-3p promotes colon cancer cell proliferation via Wnt/β-catenin signaling pathway by inhibiting WIF1 and DKK1. Eur Rev Med Pharmacol Sci. 2017;21(21):4844–4852.
19. Xing Z, Yu L, Li X, Su X. Anticancer bioactive peptide-3 inhibits human gastric cancer growth by targeting miR-338-5p. Cell Biosci. 2016;6(1):53. doi:10.1186/s13578-016-0112-8
20. Tomasello DL, Gancarz-Kausch AM, Dietz DM, Bhattacharjee A. Transcriptional regulation of the Sodium-activated Potassium Channel SLICK (KCNT2) promoter by nuclear factor-κB. J Biol Chem. 2015;290(30):18575–18583. doi:10.1074/jbc.M115.643536
21. Yang L, Xu Y, Yan Y, et al. Common nevus and skin cutaneous melanoma: prognostic genes identified by gene co-expression network analysis. Genes (Basel). 2019;10(10):
22. Yang JJ, Yang Y, Zhang C, Li J, Yang Y. Epigenetic silencing of LncRNA ANRIL enhances liver fibrosis and HSC activation through activating AMPK pathway. J Cell Mol Med. 2020;24(4):2677–2687. doi:10.1111/jcmm.14987
23. Hanniford D, Ulloa-Morales A, Karz A, et al. Epigenetic silencing of CDR1as drives IGF2BP3-mediated melanoma invasion and metastasis. Cancer Cell. 2020;37(1):55–70.e15. doi:10.1016/j.ccell.2019.12.007
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.