Back to Journals » Cancer Management and Research » Volume 12
Long Intergenic Non-Protein Coding RNA 1094 Promotes Initiation and Progression of Glioblastoma by Promoting microRNA-577-Regulated Stabilization of Brain-Derived Neurotrophic Factor
Authors Dong X, Fu X, Yu M, Li Z
Received 30 March 2020
Accepted for publication 9 June 2020
Published 9 July 2020 Volume 2020:12 Pages 5619—5631
DOI https://doi.org/10.2147/CMAR.S256147
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ahmet Emre Eşkazan
Xiaoyan Dong,1 Xiuxin Fu,1 Miao Yu,1 Zengfen Li2
1Department of Hepatobiliary Surgery, Weifang People’s Hospital, Weifang, Shandong 261000, People’s Republic of China; 2Brain Hospital, Weifang People’s Hospital, Weifang, Shandong 261000, People’s Republic of China
Correspondence: Zengfen Li
Brain Hospital, Weifang People’s Hospital, 151 Guangwen Road, Weifang, Shandong 261000, People’s Republic of China
Email [email protected]
Purpose: The long intergenic non-protein coding RNA 1094 (LINC01094) plays a vital role in the oncogenicity of clear cell renal cell carcinoma. However, its expression profile and detailed roles in glioblastoma (GBM) remain unknown. In this study, we mainly investigated the expression and roles of LINC01094 in GBM and focused on the mechanism by which LINC01094 regulates the malignant characteristics of GBM.
Patients and Methods: LINC01094 expression in GBM was determined with quantitative reverse transcription polymerase chain reaction. The proliferation, apoptosis, migration, invasion in vitro, and tumor growth in vivo of GBM cells were evaluated using Cell Counting Kit-8 assay, flow cytometry analysis, migration assay, invasion assay, and tumor xenograft models, respectively.
Results: LINC01094 was overexpressed in GBM tissues and cell lines. Moreover, increased LINC01094 expression was associated with adverse clinicopathological parameters in patients with GBM. Loss of LINC01094 inhibited GBM cell proliferation, migration, and invasion; promoted cell apoptosis; and suppressed tumor growth in vivo. Mechanically, LINC01094 functioned as a molecular sponge for microRNA-577 (miR-577) and consequently enhanced the expression of brain-derived neurotrophic factor (BDNF) in GBM cells. Both miR-577 inhibition and BDNF expression enhancement reversed LINC01094 deficiency-mediated inhibition of malignant processes in GBM cells.
Conclusion: Our results verified the involvement of the LINC01094/miR-577/BDNF pathway in GBM cells and its enhancing effects on the aggressive behaviors of GBM cells in vitro and in vivo. This pathway may be a novel and promising focus for the future development of targeted therapies for GBM.
Keywords: long intergenic non-protein coding RNA, glioblastoma, microRNA, brain-derived neurotrophic factor
Introduction
Glioma is the most common and aggressive human malignant tumor of the central nervous system.1 Annually, approximately 200,000 patients worldwide are newly diagnosed with glioma, and this number continues to increase.2 Glioma can be subdivided into four histopathological grades according to the degree of malignancy, i.e., grades I–IV.3 Glioblastoma (GBM), also known as glioblastoma multiforme, is a World Health Organization grade IV glioma. It is the most aggressive subtype and the most lethal malignancy affecting humans.4 Considerable progress has been made in therapeutic techniques for GBM, which range from surgical resection to adjuvant chemotherapy, immunotherapy, and targeted therapies. However, the current clinical oncology strategies are ineffective and the long-term prognosis of most patients remains unsatisfactory.5,6 A large proportion of patients receiving first-line therapies for GBM die within 2 years after confirmed diagnosis.7 The dismal outcome of GBM is mainly due to a characteristic infiltrative growth pattern, which increases the difficulty of radical resection and facilitates relapse.8 In this context, comprehensive elucidation of the mechanisms associated with GBM pathogenesis may significantly enable the development of potential therapeutic targets and guide future anticancer strategies.
Long non-coding RNAs (lncRNAs) are a heterogeneous group of RNA transcripts longer than 200 nucleotides.9 LncRNAs lack an open reading frame and do not encode proteins.10 Although these transcripts were first identified as transcriptional noise, recent evidence has identified the crucial roles of lncRNAs in almost all physiological and pathological processes including cancer genesis and progression.11 Studies on GBM have described differential lncRNA expression and identified certain lncRNAs to be closely linked to oncogenicity.12,13 LncRNAs may exert cancer-inhibiting or cancer-promoting activities in GBM and are implicated in the regulation of a wide array of biological behaviors.14
MicroRNAs (miRNAs) have elicited significant interest from scientific researchers in recent years. These small non-coding RNAs negatively regulate gene expression by directly binding to the 3′-untranslated region (UTR) of the target gene, resulting in mRNA degradation and/or translation inhibition.15 Studies have increasingly revealed the anomalous profiles of miRNAs in GBM and identified modulatory roles of these molecules in oncogenesis and progression.16,18 The competitive endogenous RNA (ceRNA) pathway has become a popular focus of lncRNA research.19 LncRNAs possess miRNA response elements and act as natural miRNA sponges to liberate target genes from bound miRNAs.20,21 Accordingly, exploration of the regulatory actions of the lncRNA/miRNA axis in GBM may reveal attractive cancer diagnostic, prognostic, and therapeutic targets.
Long intergenic non-protein coding RNA 1094 (LINC01094) plays a vital role in the oncogenicity of clear cell renal cell carcinoma.22 In this study, we aimed to investigate the expression patterns and detailed roles of LINC01094 in GBM. We also explored the mechanism by which LINC01094 regulates the malignant characteristics of GBM. MiR-577 is downregulated in GBM, and executes tumor-suppressing actions during cancer progression.23 Upregulation of miR-577 inhibits cell growth and cell viability in GBM by regulating the Wnt Signaling Pathway.23 MiR-577 was predicted as a potential target that may be sponged by LINC01094. After this, a series of experiments were performed to confirm this prediction.
Patients and Methods
Ethical Statement
Written informed consent was provided by all participating subjects. The study was performed under the approval of the Ethics Committee of Weifang People’s Hospital (2015#0514) and carried out in accordance with principles of the Declaration of Helsinki.
Patients and Tissue Specimens
Human GBM tissue samples were acquired from 51 patients with GBM at Weifang People’s Hospital. Normal brain tissue samples were collected from 14 patients with cerebral injuries. Patients who had received radiotherapy, chemotherapy, or other anticancer treatments were excluded from this study. All tissue samples were snap-frozen and stored in liquid nitrogen until further use.
Cell Lines and Culture Conditions
A normal human astrocyte (NHA) cell line was acquired from ScienCell Research Laboratories (Carlsbad, CA, USA). The Shanghai Institutes for Biological Sciences Cell Resource Center (Shanghai, China) provided the U251 and SHG-44 GBM cell lines. The GBM cell lines U138, LN-229, and T98 were purchased from American Type Culture Collection (Manassas, VA, USA).
U251 and LN-229 cells were maintained in Dulbecco’s modified Eagle’s medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), whereas U138 and T98 cells were cultured in Minimum Essential Media (Gibco; Thermo Fisher Scientific, Inc.). The SHG-44 and NHA cell lines were grown in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) and astrocyte medium (ScienCell Research Laboratories), respectively. All basal media were supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin antibiotics (Gibco; Thermo Fisher Scientific, Inc.). Cells were cultivated at 37°C under humidified conditions with 5% CO2.
Cell Transfection
To silence LINC01094, small interfering RNAs (siRNAs) specific for LINC01094 (si-LINC01094#1, si-LINC01094#2, and si-LINC01094#3) and negative control (NC) siRNA (si-NC) were designed and chemically synthesized by GeneChem (Shanghai, China). MiR-577 mimics and miR-577 inhibitor (both from GenePharma; Shanghai, China) were used to increase and reduce miR-577 expression, respectively. NC miRNA mimics (miR-NC) and NC inhibitor (both from GenePharma) were used as scramble controls. The BDNF-overexpressed plasmid pcDNA3.1-BDNF and the empty pcDNA3.1 plasmid vector were purchased from RiboBio (Guangzhou, China). Oligonucleotides and plasmids were transfected into cells using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
Total RNA was extracted using the TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd, Dalian, China) was used to reverse transcribe total RNA into complementary DNA (cDNA). LINC01094 and BDNF mRNA expressions were analyzed via quantitative PCR with the SYBR Premix Ex Taq kit (Takara Biotechnology Co., Ltd). Additionally, cDNA was generated from miRNA using the miScript Reverse Transcription kit (Qiagen GmbH, Hilden, Germany) and subjected to quantitative PCR using the miScript SYBR Green PCR kit (Qiagen GmbH) to quantify miR-577. U6 small nuclear RNA was used as the internal standard for miR-577 quantification, and GAPDH was used as the internal standard for LINC01094 and BDNF mRNA expressions. Relative gene expression was calculated by the 2−ΔΔCt method.
Subcellular Fractionation
Subcellular fractionation of GBM cells was performed using NE-PERTM Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific, Inc.). The relative expression of LINC01094 in the nuclear and cytoplasmic fractions was determined using RT-qPCR. GAPDH and U6 were used as the internal cytoplasmic and nuclear controls, respectively.
Cell Counting Kit-8 (CCK-8) Assay
Transfected cells were harvested after a 24-h culture and resuspended into a single-cell suspension at a density of 2 × 104 cells/mL. A 0.1-mL aliquot of the cell suspension was inoculated into each well of a 96-well plate. The cells were treated with 10 µL CCK-8 solution (Dojindo, Tokyo, Japan) every 24 h until the endpoint of 72 h. After additional 2-h incubation at 37°C in 5% CO2, the optical absorbance at a wavelength of 450 nm was detected using a microplate reader (BioTek Instruments, Winooski, VT, USA).
Flow Cytometry Analysis
At post-transfection 48 h, cells were detached from the culture plates using EDTA-free trypsin (Gibco; Thermo Fisher Scientific, Inc.) and washed twice with phosphate-buffered saline at 4°C. After centrifugation at 1000 rpm for 5 min at 4°C, the supernatant was discarded and the collected cells were subjected to cell apoptosis analysis using the annexin V–fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (BioLegend, San Diego, CA, USA). The transfected cells were resuspended in 100 μL of 1× binding buffer and double stained with 5 μL FITC–annexin V and 5 μL propidium iodide. Apoptotic cells were quantified via flow cytometry using FACScan (BD Biosciences, Franklin Lakes, NJ, USA) after 15-min incubation in the dark.
Migration and Invasion Assays
The migratory abilities of GBM cells were determined using Transwell chambers (8-μm pore size; BD Biosciences). GBM cells that had been transfected with the aforementioned molecular products were digested with Trypsin-EDTA (Gibco; Thermo Fisher Scientific, Inc.), pelleted by centrifugation, and resuspended in FBS-free basal medium. The cell suspension was adjusted to a density of 5 × 105 cells/mL. The upper chambers were loaded with 200 µL of the cell suspension, and the lower chambers were filled with 600 μL of culture medium containing 20% FBS. After 24-h incubation at 37°C with 5% CO2, migratory cells were fixed with 4% paraformaldehyde and stained with 0.5% crystal violet. The stained cells were extensively washed and counted under an inverted microscope (Olympus Corporation, Tokyo, Japan). Invasion assay was performed using the same procedure as migration assay, except that the Transwell chambers were precoated with Matrigel (BD Biosciences).
Tumor Xenograft Model
LINC01094 short hairpin RNA (shRNA; sh-LINC01094) and negative control shRNA (sh-NC) were packaged using a lentiviral packaging kit (GeneChem) and transfected into U251 cells. Puromycin was used to select stably transfected cells.
Male BALB/c nude mice aged 4–6 weeks (Shanghai SLAC Laboratory Animal Co., Ltd; Shanghai, China) were maintained under specific pathogen-free conditions (25°C; 50% humidity; 10-h light/14-h dark cycle). All animal research was approved by the Institutional Animal Care Committee of Weifang People’s Hospital (2018#1204), and conducted in compliance with the Animal Protection Law of the People’s Republic of China-2009 for experimental animals. A total of 5 × 106 U251 cells with stable expression of either sh-LINC01094 or sh-NC were inoculated subcutaneously into the flank of each mouse. The width and length of each subcutaneous tumor was measured every 5 days using a caliper, and tumor volume was calculated using the following formula: 0.5 × length × width2. All mice were euthanized after 30 days, and the subcutaneous tumors were excised, photographed, and weighed. Total RNA and protein samples were also extracted and subjected to RT-qPCR and Western blotting, respectively.
Bioinformatics Prediction Analysis
The Gene Expression Profiling Interactive Analysis (GEPIA) database (http://gepia.cancer-pku.cn/), which includes data from The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx), was used to evaluate the expression of LINC01094 in gliomas and its relationship with overall survival. Starbase 3.0 (http://starbase.sysu.edu.cn/) was used to predict the miRNA(s) that may be sponged by LINC01094. Three bioinformatics databases, Starbase 3.0, miRDB (http://mirdb.org/), and TargetScan (http://www.targetscan.org/vert_71/), were used to identify the putative targets of miR-577.
RNA Immunoprecipitation (RIP) Assay
RIP assay was performed using the Magna RIP RNA-binding protein immunoprecipitation kit (Merck Millipore, Darmstadt, Germany). GBM cell suspensions were lysed using complete RIP lysis buffer (KeyGen BioTECH; Nanjing, China). Whole-cell extract was incubated overnight at 4°C with an equal volume of magnetic beads coupled with human anti-Argonaute 2 (Ago2; Millipore) or anti-immunoglobulin G (IgG; Millipore) antibodies. Then, magnetic beads were harvested and probed with Proteinase K for protein digestion. Immunoprecipitated RNA was subjected to RT-qPCR.
Luciferase Reporter Assay
A LINC01094 fragment containing the predicted wild-type (wt) miR-577 binding site and a mutant (mut) LINC01094 fragment was designed and chemically generated by GenePharma. The fragments were inserted into the pmirGLO luciferase reporter plasmid (Promega Corporation, Madison, WI, USA) to yield LINC01094-wt and LINC01094-mut plasmids, respectively. The same experimental steps were also used to generate BDNF-wt and BDND-mut reporter plasmids.
Cells were cultured in 24-well plates and transfected with either the wt or mut reporter plasmid and miR-577 mimic or miR-NC using Lipofectamine 2000. Transfected cells were incubated at 37°C in a humidified atmosphere with 5% CO2 for 48 h. Luciferase activity was detected using the Dual-Luciferase Reporter Assay System (Promega Corporation). Luciferase activity was normalized against Renilla luciferase activity.
Western Blotting
RIPA buffer (Pierce; Thermo Fisher Scientific, Inc.) was used to isolate total proteins from transfected cells. The concentrations of lysates were determined using a BCA protein assay kit (Beyotime Biotechnology; Shanghai, China). Equal amounts of protein were separated via 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride membranes. After 2-h blocking in 5% defatted milk powder at room temperature, the membranes were reacted overnight at 4°C with primary antibodies against BDNF (ab108319; Abcam, Cambridge, MA, USA) or GAPDH (ab181603; Abcam). Subsequently, the membranes were treated with horseradish peroxidase-labeled secondary antibodies (ab205718; Abcam) for 1 h at room temperature. Finally, signals from labeled proteins were detected using the ECL Detection Kit (GE Healthcare Life Sciences, Chalfont, UK).
Statistical Analysis
All data are expressed as mean ± standard deviation. Comparisons between two groups were performed using Student’s t-test. One-way analysis of variance combined with Tukey’s test was performed to analyze differences among multiple groups. Correlations between the expressions of two genes in GBM tissues were examined using Pearson’s correlation coefficient analysis. The Kaplan–Meier method was performed to generate survival curves, which were compared using the Log rank test. P-values of <0.05 were considered statistically significant.
Results
LINC01094 Expression Is Elevated in GBM Tissues and Cell Lines
Analysis of tissue microarray data from the GEPIA database confirmed an evident increase in LINC01094 expression in glioma tissues (n = 163) compared with that in normal brain tissues (n = 207; Figure 1A). To confirm this finding, RT-qPCR was performed to detect LINC01094 expression in 51 GBM tissues and 14 normal brain tissues. The results revealed that LINC01094 was significantly upregulated in GBM tissues compared with in normal brain tissues (Figure 1B). The same trend of LINC01094 expression was also identified in five GBM cell lines (U251, SHG-44, U138, LN-229, and T98; Figure 1C).
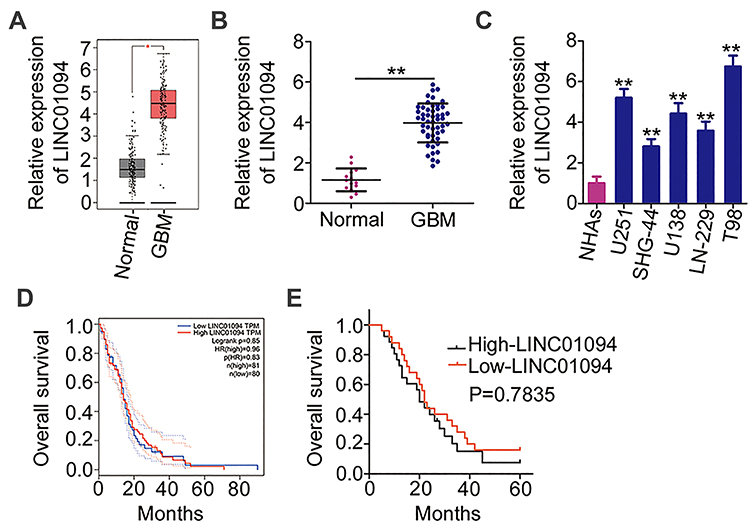 |
Figure 1 Elevated long intergenic non-protein coding RNA 1094 (LINC01094) expression in glioblastoma (GBM) tissues and cell lines (A) The Gene Expression Profiling Interactive Analysis database was used to analyze the expression of LINC01094 in glioma tissues (n = 163) and normal brain tissues (n = 207). (B) LINC01094 expression was evaluated in 51 GBM tissues and 14 normal brain tissues based on quantitative reverse transcription polymerase chain reaction (RT-qPCR). (C) LINC01094 expression in five GBM cell lines (U251, SHG-44, U138, LN-229, and T98) versus normal human astrocytes was evaluated and compared using RT-qPCR. (D) The Gene Expression Profiling Interactive Analysis database was used to analyze the association between LINC01094 expression and overall survival in patients with glioma. (E) Kaplan–Meier analysis was performed to evaluate the correlation between LINC01094 expression and overall survival in patients with GBM (P = 0.7835). **P < 0.01. |
Regarding the clinical relevance of LINC01094 in GBM, overall survival analysis based on the GEPIA database yielded a non-significant correlation between LINC01094 expression and the overall survival of patients with GBM (Figure 1D). Our data yielded the same result (Figure 1E; P = 0.7835). Using the median value of LINC01094 in GBM tissues as the cutoff score, all patients were classified into the high- or low-LINC01094 group. Statistical analysis indicated that high LINC01094 expression was significantly correlated with the mean tumor diameter (P = 0.025) and Karnofsky Performance Scale (KPS) score (P = 0.012; Table 1). Overall, these results suggest that LINC01094 expression is elevated in GBM and is associated with tumor progression.
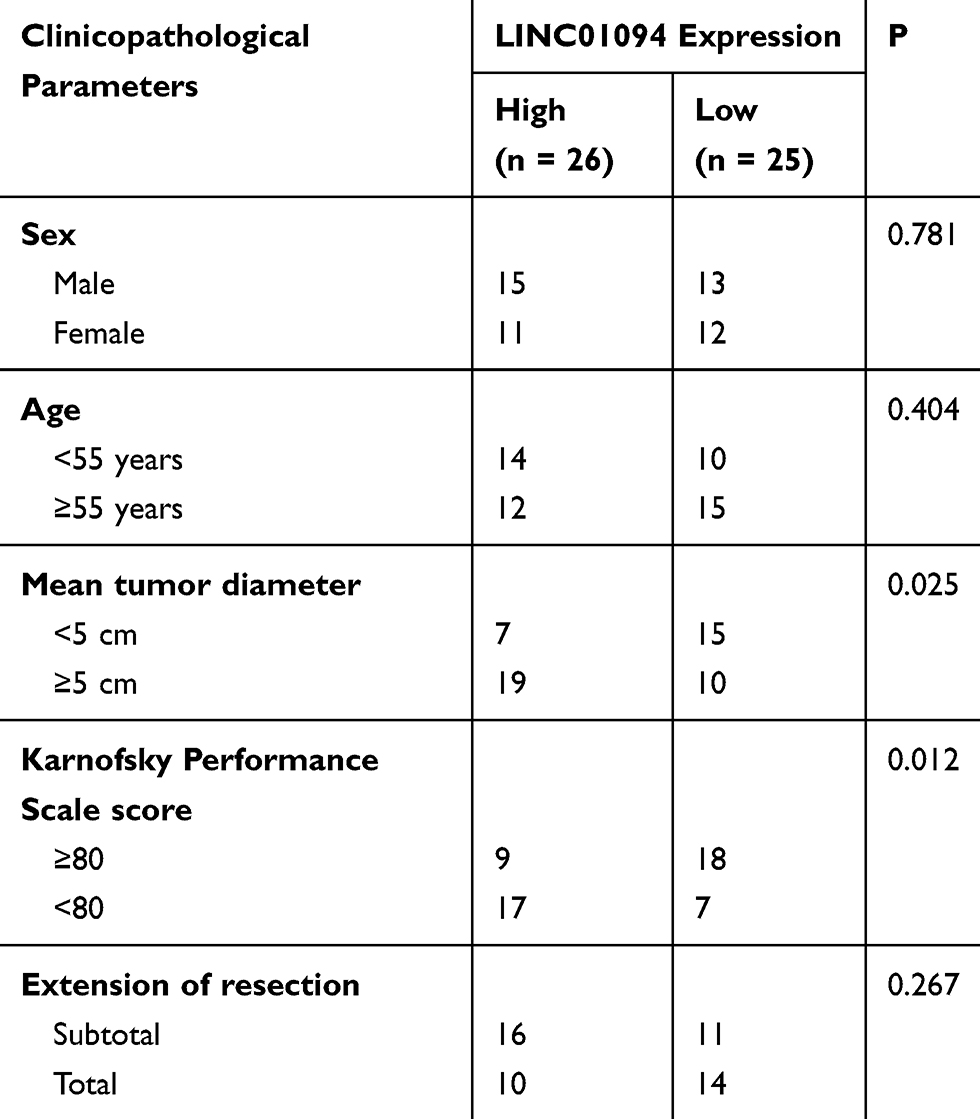 |
Table 1 Association Between Long Intergenic Non-Coding RNA 1094 (LINC01094) and Clinicopathological Parameters in Patients with GBM |
Downregulation of LINC01094 Inhibits the Malignant Processes in GBM Cells
Because the U251 and T98 cell lines exhibited the strongest LINC01094 expression among the five GBM cell lines, these two cell lines were used in the subsequent experiments. To explore the role of LINC01094 as a regulator of malignant processes in GBM cells, siRNAs targeting LINC01094 were designed and evaluated to determine the efficacy of silencing. RT-qPCR verified that si-LINC01094#1 and si-LINC01094#2 achieved optimal LINC01094 knockdown efficacy (Figure 2A). To decrease the risk of off-target effects, si-LINC01094#1 and si-LINC01094#2 were used in the subsequent functional assays. The effect of LINC01094 downregulation on the proliferation of GBM cells was determined using CCK-8 assay. In U251 and T98 cells, LINC01094 knockdown resulted in obvious decreases in cell proliferation (Figure 2B). Additionally, LINC01094 depletion strongly enhanced the apoptosis of U251 and T98 cells, as suggested by flow cytometry analysis (Figure 2C). Migration and invasion assays clearly illustrated that LINC01094 downregulation inhibited the migratory (Figure 2D) and invasive (Figure 2E) abilities of U251 and T98 cells. Altogether, these observations suggest that LINC01094 plays a pro-oncogenic role during GBM genesis and progression.
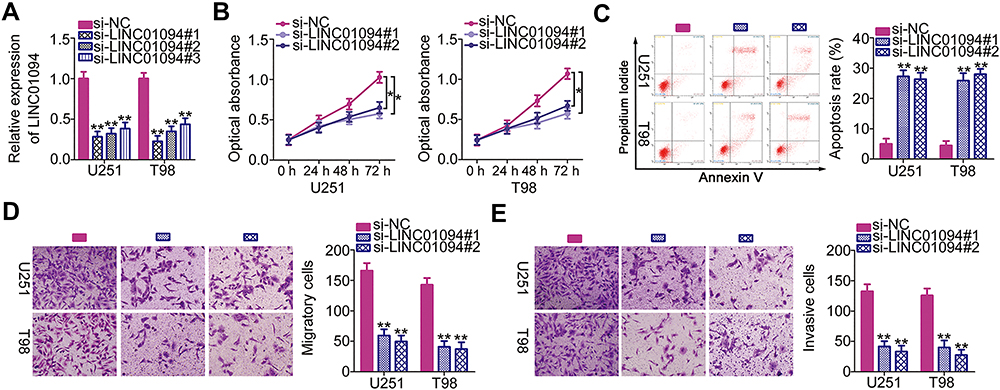 |
Figure 2 Long intergenic non-protein coding RNA 1094 (LINC01094) depletion inhibits the proliferation, migration, and invasion and promotes the apoptosis of U251 and T98 glioblastoma cell lines in vitro. (A) Quantitative reverse transcription polymerase chain reaction (RT-qPCR) was performed to assess the efficiency of transfection with small interfering RNAs specific for LINC01094 (si-LINC01094) or negative control (NC) siRNA (si-NC). (B) Cell Counting Kit-8 assay was performed to analyze the proliferative abilities of U251 and T98 cells after LINC01094 knockdown. (C) Flow cytometry was performed to evaluate the frequency of apoptosis in si-LINC01094-transfected or si-NC-transfected U251 and T98 cells. (D, E) U251 and T98 cells were subjected to migration and invasion assays after transfection with si-LINC01094 or si-NC. *P < 0.05 and **P < 0.01. |
LINC01094 Acts as an Efficient miR-577 Sponge in GBM Cells
After determining that LINC01094 was pro-oncogenic in GBM cells, we next elucidated the mechanisms by which LINC01094 exerted these effects. The mechanisms underlying the functions of lncRNAs are determined primarily by the locations of these molecules.24 Hence, we determined the location of LINC01094 expression in GBM cells using subcellular fractionation. In U251 and T98 cells, LINC01094 was mainly concentrated in the cytoplasm rather than in the nucleus (Figure 3A), suggesting that this lncRNA regulates gene expression as a molecular sponge or ceRNA. We then performed bioinformatics prediction analysis to identify potential target miRNAs. Among the identified candidates, miR-577 (Figure 3B) was exceptional because it was shown to play anti-oncogenic roles in GBM cells23,25 and was selected for further verification.
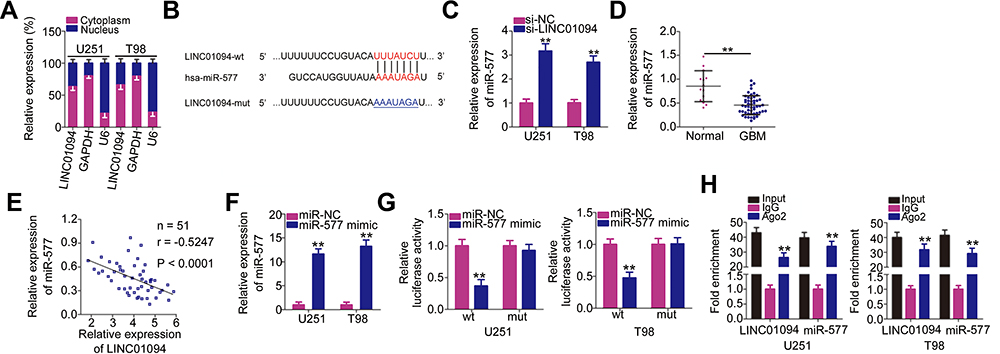 |
Figure 3 Long intergenic non-protein coding RNA 1094 (LINC01094) acts as a molecular sponge for microRNA (miR)-577 in glioblastoma (GBM) cells. (A) Subcellular fractionation was performed to test the localization of LINC01094 expression in U251 and T98 cells. (B) Bioinformatics prediction analysis predicted the site of binding between wild-type LINC01094 and miR-577. The mutant binding sequences are also shown. Hsa represents the meaning of human. (C) miR-577 expression in U251 and T98 cells transfected with si-LINC01094 or si-NC was detected by quantitative reverse transcription polymerase chain reaction (RT-qPCR). (D) miR-577 expression was detected in 51 GBM tissues and 14 normal brain tissues using RT-qPCR. (E) Pearson’s correlation coefficient analysis was performed to determine the association between LINC01094 and miR-577 expressions in 51 GBM tissues (r = −0.5247, P < 0.0001). (F) The efficiency of miR-577 mimic in U251 and T98 cells was quantified using RT-qPCR. (G) U251 and T98 cells were cotransfected with miR-577 mimic or negative control miRNA mimics (miR-NC) and plasmids encoding wild-type (LINC01094-wt) or mutant LINC01094 (LINC01094-mut). After 48-h culture, luciferase activity in the cells was analyzed using the Dual-Luciferase Reporter Assay System. (H) RNA immunoprecipitation assay was performed to verify the interaction between LINC01094 and miR-577 in U251 and T98 cells. **P < 0.01. |
To verify this prediction, we performed RT-qPCR to determine the relationship between LINC01094 and miR-577 in GBM. Notably, miR-577 expression increased in U251 and T98 cells when LINC01094 was knocked down (Figure 3C). In addition, miR-577 expression decreased in GBM tissues compared with that in normal brain tissues (Figure 3D). Importantly, Pearson’s correlation coefficient analysis demonstrated an inverse correlation between the expression levels of LINC01094 and miR-577 in the 51 GBM tissues (Figure 3E; r = −0.5247, P < 0.0001). Next, we performed luciferase reporter assay to determine whether miR-577 could bind directly to LINC01094. First, as illustrated in Figure 3F, RT-qPCR validated that transfection with miR-577 mimic significantly increased miR-577 expression in U251 and T98 cells. This upregulation of miR-577 led to noticeable decreases in the LINC01094-wt-associated luciferase activity in U251 and T98 cells, whereas the luciferase activity of LINC01094-mut was unaffected (Figure 3G). Furthermore, both LINC01094 and miR-577 were preferentially enriched in a precipitation experiment using Ago2 antibody-coupled magnetic beads (Figure 3H). In summary, these findings prove that LINC01094 functions as a molecular sponge for miR-577 in GBM cells.
BDNF Is a Direct Target of miR-577 and Is Positively Regulated by LINC01094 in GBM Cells
Because miR-577 was identified as a cancer-inhibiting miRNA in GBM cells,23,25 we next studied the mechanisms by which miR-577 suppressed the malignant characteristics of these cells. Three databases (Starbase 3.0, miRDB, and TargetScan) were searched to identify miR-577 target genes. We selected BDNF (Figure 4A) based on previous reports describing the tumor-promoting activities of BDNF and the strong expression in GBM.26,27 Luciferase reporter assay was performed to assess whether miR-577 could bind directly to the 3′-UTR of BDNF. The reintroduction of miR-577 clearly reduced the luciferase activity of BDNF-wt, but not of BDNF-mut, in U251 and T98 cells (Figure 4B). Moreover, RT-qPCR and Western blotting revealed that the expressions levels of BDNF mRNA (Figure 4C) and protein (Figure 4D) were distinctly downregulated in U251 and T98 cells treated with the miR-577 mimic. Next, RT-qPCR revealed that BDNF mRNA was overexpressed in GBM tissues compared with that in normal brain tissues (Figure 4E). We then performed Pearson’s correlation coefficient analysis to analyze the correlation between miR-577 and BDNF mRNA expressions in the 51 GBM tissues. Further, BDNF mRNA expression was found to correlate inversely with miR-577 expression in these tissues (Figure 4F; r = −0.5926, P < 0.0001).
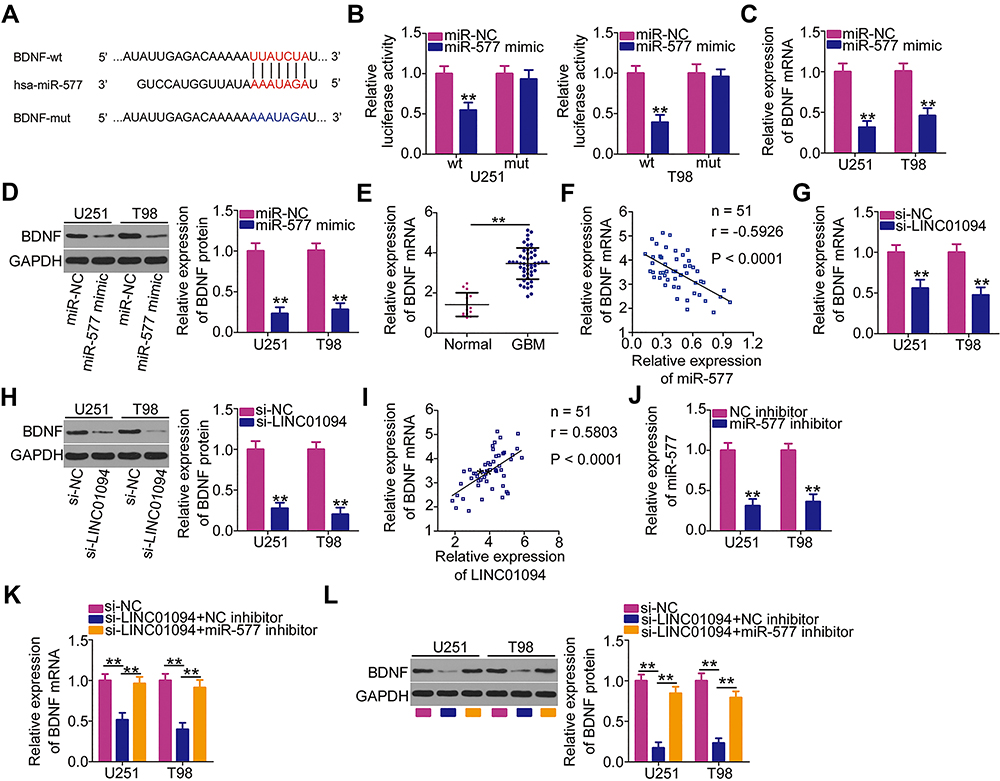 |
Figure 4 Long intergenic non-protein coding RNA 1094 (LINC01094) regulates brain-derived neurotrophic factor (BDNF) expression in U251 and T98 cells by sponging microRNA (miR)-577. (A) The wild-type miR-577 binding site in the 3′-untranslated region of BDNF, as predicted by bioinformatics prediction analysis. Hsa represents the meaning of human. (B) Luciferase activity was measured in U251 and T98 cells after cotransfection with miR-577 mimic or negative control miRNA mimics (miR-NC) and luciferase reporter plasmids encoding wild-type (BDNF-wt) or mutant BDNF (BDNF-mut). (C, D) Detection of BDNF mRNA and protein levels by quantitative reverse transcription polymerase chain reaction (RT-qPCR) and Western blotting, respectively, in miR-577 overexpressing-U251 and T98 cells. (E) BDNF mRNA expression in 51 GBM tissues and 14 normal brain tissues was evaluated using RT-qPCR. (F) Pearson’s correlation coefficient analysis was performed to assess the correlation between miR-577 and BDNF mRNA expressions in 51 GBM tissues (r = −0.5926, P < 0.0001). (G, H) RT-qPCR and Western blotting were performed to measure BDNF mRNA and protein expression levels in LINC01094-deficient U251 and T98 cells. (I) Pearson’s correlation coefficient analysis was performed to detect a positive relationship between LINC01094 and BDNF mRNA expression levels in the 51 GBM tissues (r = 0.5803, P < 0.0001). (J) U251 and T98 cells were treated with miR-577 or negative control (NC) inhibitor. Transfection efficiency was assessed using RT-qPCR. (K, L) U251 and T98 cells were treated simultaneously with small interfering RNA specific for LINC01094 (si-LINC01094) and either miR-577 inhibitor or NC inhibitor. BDNF mRNA and protein expression levels were respectively analyzed using RT-qPCR and Western blotting, respectively. **P < 0.01. |
The ability of lncRNAs to regulate the expressions of target mRNAs via the ceRNA pathway has been widely accepted.20 A series of experiments was performed to determine whether LINC01094 affects BDNF expression in GBM cells. RT-qPCR and Western blotting affirmed that the loss of LINC01094 considerably decreased the expressions levels of BDNF mRNA (Figure 4G) and protein (Figure 4H) in U251 and T98 cells. The correlation between the BDNF and LINC01094 expressions was also analyzed in the 51 GBM tissues using Pearson’s correlation coefficient analysis. The results indicated that LINC01094 expression was correlated positively with BDNF mRNA expression in these tissues (Figure 4I; r = 0.5803, P < 0.0001). Furthermore, rescue experiments were performed to verify whether LINC01094 positively regulated BDNF expression by sponging miR-577. First, the efficiency of miR-577 inhibitor was evaluated using RT-qPCR. Transfection with miR-577 inhibitor markedly suppressed miR-577 expression in U251 and T98 cells (Figure 4J). Subsequently, miR-577 inhibitor or NC inhibitor was cotransfected with si-LINC01094 into U251 and T98 cells. BDNF mRNA and protein expressions in the transfected cells was analyzed using RT-qPCR and Western blotting, respectively. Notably, LINC01094 silencing led to the downregulation of both BDNF mRNA (Figure 4K) and protein (Figure 4L) expressions in U251 and T98 cells. By contrast, the miR-577 inhibitor abrogated these regulatory actions, suggesting that LINC01094 exerts its regulatory effects on BDNF expression by sponging miR-577. These assays suggest that LINC01094 modulates BDNF expression in GBM cells by binding competitively to miR-577.
LINC01094 Exerts Tumor-Promoting Activities in GBM Cells by Upregulating the Output of the miR-577/BDNF Axis
Rescue experiments were performed to determine whether LINC01094 regulates the malignant processes in GBM cells via the miR-577/BDNF axis. U251 and T98 cells were transfected with the miR-577 inhibitor or NC inhibitor in the presence of si-LINC01094. Functional assays demonstrated that the LINC01094 deficiency-induced suppression of U251 and T98 cell proliferation (Figure 5A), promotion of cell apoptosis (Figure 5B), and impairment of cell migration (Figure 5C) and invasion (Figure 5D) were markedly abolished by cotransfection with the miR-577 inhibitor.
 |
Figure 5 Inhibition of microRNA (miR)-577 or reintroduction of brain-derived neurotropic factor (BDNF) reverses the inhibited malignant characteristics of long intergenic non-protein coding RNA 1094 (LINC01094)-deficient glioblastoma (GBM) cells. (A) U251 and T98 GBM cells transfected with small interfering RNA specific for LINC01094 (si-LINC01094) in the presence of miR-577 inhibitor or negative control (NC) inhibitor were subjected to Cell Counting Kit (CCK)-8 assay to evaluate cell proliferation. (B) Flow cytometry analysis was utilized to determine the apoptosis of U251 and T98 cells that were treated as above described. (C, D) Migration and invasion assays were carried out to assess the migratory (C) and invasive (D) capacities of aforementioned cells. (E) BDNF protein expression levels in U251 and T98 cells transfected with plasmid encoding BDNF (pcDNA3.1-BDNF) or empty vector (pcDNA3.1) were detected by Western blotting. (F) U251 and T98 cells were cotransfected with si-LINC01094 and pcDNA3.1-BDNF or pcDNA3.1. The proliferation of transfected cells was detected by CCK-8 assay. (G) Flow cytometry analysis was employed in determining the apoptosis of U251 and T98 cells that were cotransfected with si-LINC01094 and pcDNA3.1-BDNF or pcDNA3.1. (H, I) Migration and invasion assays were used to analyze the migration (H) and invasion (I) of U251 and T98 cells after cotransfection with si-LINC01094 and pcDNA3.1-BDNF or pcDNA3.1. *P < 0.05 and **P < 0.01. |
pcDNA3.1-BDNF was also used in rescue experiments, and its transfection efficiency was evaluated by Western blotting (Figure 5E). pcDNA3.1-BDNF or empty vector was cotransfected with si-LINC01094 into U251 and T98 cells. CCK-8 assay and flow cytometry analysis revealed that LINC010094 downregulation suppressed cell proliferation (Figure 5F) while promoting cell apoptosis (Figure 5G), whereas this pattern was nearly rescued by cotransfection with pcDNA3.1-BDNF. The results of migration and invasion assays also validated this phenomenon (Figure 5H and I). In summary, LINC01094 exerts its pro-oncogenic roles on the malignant processes in GBM cells by regulating the output of the miR-577/BDNF axis.
Depletion of LINC01094 Inhibits GBM Tumor Growth in vivo
A xenograft tumor model was used to further address the biological roles of LINC01094 in the growth of GBM tumors in vivo. Nude mice were injected subcutaneously with U251 cells that had been stably transfected with sh-LINC01094 or sh-NC. Xenograft tumors derived from sh-LINC01094-transfected U251 cells exhibited reductions in size (Figure 6A), volume (Figure 6B), and weight (Figure 6C) compared with the tumors derived from sh-NC-transfected cells. RT-qPCR and Western blotting of xenograft tumors revealed that xenograft tumors derived from sh-LINC01094-treated cells tended to express lower expression levels of LINC01094 (Figure 6D) and BDNF protein (Figure 6E) as well as higher expression level of miR-577 (Figure 6F) than those derived from sh-NC-treated cells. These data suggest that LINC01094 depletion led to GBM tumor growth suppression in vivo by enhancing miR-577 and decreasing BDNF expression.
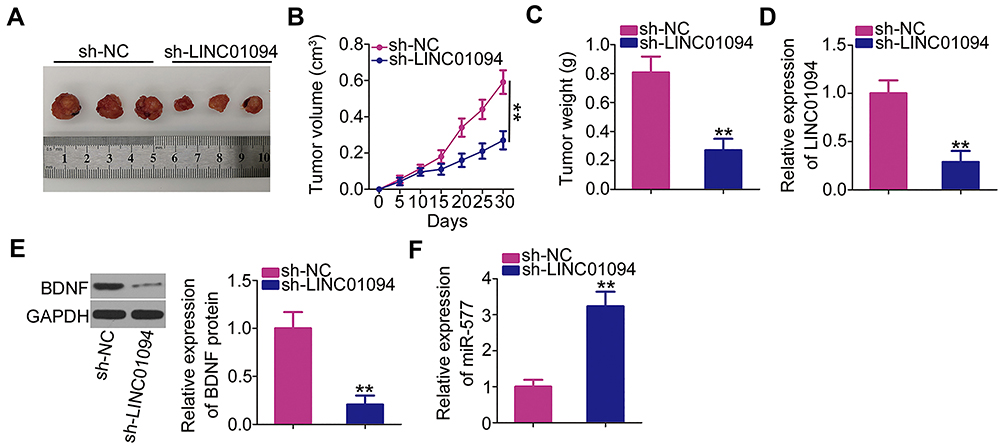 |
Figure 6 Depletion of long intergenic non-protein coding RNA 1094 (LINC01094) inhibits glioblastoma xenograft growth in vivo. (A) Tumor xenografts were excised on day 30 after cell injection and photographed. (B) Tumor volumes were monitored every 5 days after cell injection, as shown in the corresponding growth curves. (C) The weights of the xenograft tumors were measured 30 days after cell injection. (D–F) The expression levels of LINC01094, BDNF protein, and microRNA-577 in the xenograft tumors. **P < 0.01. |
Discussion
Recent research has focused increasingly on lncRNAs, which control many genetic pathways.24,28,29 Studies from the last decade have increasingly highlighted the important roles of lncRNAs in GBM.30,32 Dysregulated lncRNAs can modulate the malignant processes associated with GBM initiation and progression.33 Therefore, lncRNAs may be developed as effective diagnostic biomarkers and therapeutic targets in GBM. Unfortunately, many lncRNAs remain to be thoroughly investigated in GBM. In this study, we mainly evaluated the expression characteristic and clinical relevance of LINC01094 in GBM. More importantly, we explored the biological roles of LINC01094 in GBM cells and the associated molecular mechanism.
LINC01094 is expressed at high levels in clear cell renal cell carcinoma.22 Functionally, LINC01094 depletion restricted cell growth and metastasis in clear cell renal cell carcinoma.22 Nevertheless, the expression profile and detailed roles of LINC01094 in GBM remain unknown. In this study, we used the GEPIA database (including TCGA and GTEx projects) to demonstrate that LINC01094 is strongly expressed in glioma, which is in line with our results. In addition, LINC01094 was upregulated in the five tested GBM cell lines compared with in NHAs. To determine the clinical value, we also analyzed the correlations between LINC01094 expression and clinicopathological parameters in patients with GBM. The increased expression of LINC01094 was significantly correlated with the mean tumor diameter and KPS score in patients with GBM. However, neither the data obtained from the GEPIA database nor our results revealed an association between LINC01094 expression and overall survival in patients with GBM. Biologically, LINC01094 loss led to the suppression of GBM cell proliferation, migration, and invasion but the promotion of cell apoptosis in vitro. In addition, interference with LINC01094 expression impaired the growth of GBM cells in vivo.
LncRNAs have been implicated widely in the regulation of cancer oncogenesis and progression through a range of mechanisms.34 The ceRNA hypothesis is becoming increasingly popular in lncRNA research and provides a network regulatory model that comprises lncRNA, miRNA, and mRNA.20 LncRNAs possess miRNA response elements and act as molecular sponges for specific miRNAs, thereby decreasing the binding of these miRNAs to their target mRNAs.35 To elucidate the possible mechanisms by which LINC01094 contributes to GBM progression, we performed subcellular fractionation and RT-qPCR to determine the localization of LINC01094 expression in GBM cells. As a result, majority of LINC01094 concentrated in GBM cell cytoplasm rather than nucleus, reflecting its probability as a molecular sponge for miRNAs.
Bioinformatics prediction analysis performed in this study predicted complementary base pairing between miR-577 and LINC01094. To prove this hypothesis, we first clarified the correlation between the expressions of LINC01094 and miR-577. LINC01094 knockdown led to notable upregulation of miR-577 in GBM cells. Moreover, miR-577 downregulation was correlated inversely with the expression of LINC01094 in GBM tissues. Furthermore, luciferase reporter and RIP assays were performed to validate the link between LINC01094 and miR-577 in GBM cells. Notably, miR-577 could bind directly to LINC01094 in these cells. We then performed RT-qPCR and Western blotting to demonstrate decreases in the mRNA and protein expression levels of BDNF, a target of miR-577, in GBM cells after LINC01094 knockdown. Further rescue experiments elucidated that LINC01094 silencing caused downregulation of BDNF expression was markedly recovered in GBM cells after miR-577 inhibition. To summarize, our results demonstrated, for the first time, that LINC01094 acts as a ceRNA in GBM cells by competitively binding to miR-577, which rescues the inhibition of BDNF and consequently increases its expression.
MiR-577 is expressed at low levels in GBM tissues and cell lines.23 This miRNA acts as a tumor suppressor in GBM cells, where it affects tumor cell viability, proliferation, migration, and invasion.23,25 However, few studies have explored the downstream targets of miR-577 in GBM cells. The results of this study identified BDNF as a direct target of miR-577 in GBM cells. BDNF is an important neurotrophin in the brain36 and is highly expressed in tumor tissues.26 Functional assays indicated that BDNF acts as an oncogene in GBM by inducing the malignant characteristics of GBM.27 Herein, our data demonstrate that LINC01094 knockdown decreased BDNF expression in GBM cells by sponging miR-577, whereas an increased output of the miR-577/BDNF axis counteracted the LINC01094 deficiency-mediated inhibition of malignant processes in GBM cells. Thus, our study verified the involvement of the LINC01094/miR-577/BDNF pathway in GBM cells and its enhancing effects on the aggressive behaviors of GBM cells in vitro and in vivo.
Conclusion
LINC01094 promoted the initiation and progression of GBM. Mechanically, LINC01094 exerted its oncogenic actions in GBM cells partially by acting as a ceRNA for miR-577 and consequently enhancing BDNF expression. Our results present an attractive, novel, and promising direction for the future development of targeted therapies for GBM.
Disclosure
The authors declare that they have no competing interests.
References
1. Johnson KJ, Schwartzbaum J, Kruchko C, et al. Brain tumor epidemiology in the era of precision medicine: the 2017 brain tumor epidemiology consortium meeting report. Clin Neuropathol. 2017;36(6):255–263. doi:10.5414/NP301066
2. Li M, Deng H, Peng H, Wang Q. Functional nanoparticles in targeting glioma diagnosis and therapies. J Nanosci Nanotechnol. 2014;14(1):415–432. doi:10.1166/jnn.2014.8757
3. Wang Y, Jiang T. Understanding high grade glioma: molecular mechanism, therapy and comprehensive management. Cancer Lett. 2013;331(2):139–146. doi:10.1016/j.canlet.2012.12.024
4. Dolecek TA, Propp JM, Stroup NE, Kruchko C. CBTRUS statistical report: primary brain and central nervous system tumors diagnosed in the United States in 2005–2009. Neuro-Oncology. 2012;14(Suppl 5):v1–v49. doi:10.1093/neuonc/nos218
5. Delgado-Lopez PD, Corrales-Garcia EM. Survival in glioblastoma: a review on the impact of treatment modalities. Clin Transl Oncol. 2016;18(11):1062–1071. doi:10.1007/s12094-016-1497-x
6. Wang J, Su HK, Zhao HF, Chen ZP, To SS. Progress in the application of molecular biomarkers in gliomas. Biochem Biophys Res Commun. 2015;465(1):1–4. doi:10.1016/j.bbrc.2015.07.148
7. Kim BS, Seol HJ, Nam DH, et al. Concurrent chemoradiotherapy with temozolomide followed by adjuvant temozolomide for newly diagnosed glioblastoma patients: a retrospective multicenter observation study in Korea. Cancer Res Treat. 2017;49(1):193–203. doi:10.4143/crt.2015.473
8. Kotliarova S, Fine HA. SnapShot: glioblastoma multiforme. Cancer Cell. 2012;21(5):710–710 e711. doi:10.1016/j.ccr.2012.04.031
9. Park JY, Lee JE, Park JB, Yoo H, Lee SH, Kim JH. Roles of long non-coding RNAs on tumorigenesis and glioma development. Brain Tumor Res Treat. 2014;2(1):1–6. doi:10.14791/btrt.2014.2.1.1
10. Ulitsky I, Bartel DP. lincRNAs: genomics, evolution, and mechanisms. Cell. 2013;154(1):26–46. doi:10.1016/j.cell.2013.06.020
11. Fu Z, Luo W, Wang J, et al. Malat1 activates autophagy and promotes cell proliferation by sponging miR-101 and upregulating STMN1, RAB5A and ATG4D expression in glioma. Biochem Biophys Res Commun. 2017;492(3):480–486. doi:10.1016/j.bbrc.2017.08.070
12. Li J, Zhang M, An G, Ma Q. LncRNA TUG1 acts as a tumor suppressor in human glioma by promoting cell apoptosis. Exp Biol Med. 2016;241(6):644–649. doi:10.1177/1535370215622708
13. Cui B, Li B, Liu Q, Cui Y. lncRNA CCAT1 promotes glioma tumorigenesis by sponging miR-181b. J Cell Biochem. 2017;118(12):4548–4557. doi:10.1002/jcb.26116
14. Zeng T, Li L, Zhou Y, Gao L. Exploring long noncoding RNAs in glioblastoma: regulatory mechanisms and clinical potentials. Int J Genomics. 2018;2018:2895958. doi:10.1155/2018/2895958
15. Acunzo M, Romano G, Wernicke D, Croce CM. MicroRNA and cancer–a brief overview. Adv Biol Regul. 2015;57:1–9. doi:10.1016/j.jbior.2014.09.013
16. Alamdari-Palangi V, Amini R, Karami H. MiRNA-7 enhances erlotinib sensitivity of glioblastoma cells by blocking the IRS-1 and IRS-2 expression. J Pharm Pharmacol. 2020;72(4):531–538. doi:10.1111/jphp.13226
17. Bahreyni-Toossi MT, Dolat E, Khanbabaei H, Zafari N, Azimian H. microRNAs: potential glioblastoma radiosensitizer by targeting radiation-related molecular pathways. Mutat Res. 2019;816–818:111679. doi:10.1016/j.mrfmmm.2019.111679
18. Li Y, Li W, Zeng X, et al. The role of microRNA-148a and downstream DLGAP1 on the molecular regulation and tumor progression on human glioblastoma. Oncogene. 2019;38(47):7234–7248. doi:10.1038/s41388-019-0922-3
19. Shuwen H, Qing Z, Yan Z, Xi Y. Competitive endogenous RNA in colorectal cancer: a systematic review. Gene. 2018;645:157–162. doi:10.1016/j.gene.2017.12.036
20. Wang L, Cho KB, Li Y, Tao G, Xie Z, Guo B. Long noncoding RNA (lncRNA)-mediated competing endogenous RNA networks provide novel potential biomarkers and therapeutic targets for colorectal cancer. Int J Mol Sci. 2019;20(22):5758.
21. Liu J, Li W, Zhang J, Ma Z, Wu X, Tang L. Identification of key genes and long non-coding RNA associated ceRNA networks in hepatocellular carcinoma. PeerJ. 2019;7:e8021. doi:10.7717/peerj.8021
22. Jiang Y, Zhang H, Li W, Yan Y, Yao X, Gu W. FOXM1-activated LINC01094 promotes clear cell renal cell carcinoma development via MicroRNA 224-5p/CHSY1. Mol Cell Biol. 2020;40(3).
23. Zhang W, Shen C, Li C, et al. miR-577 inhibits glioblastoma tumor growth via the Wnt signaling pathway. Mol Carcinog. 2016;55(5):575–585. doi:10.1002/mc.22304
24. Tsagakis I, Douka K, Birds I, Aspden JL. Long non-coding RNAs in development and disease: conservation to mechanisms. J Pathol. 2020;250(5):480–495. doi:10.1002/path.5405
25. Wei N, Wei H, Zhang H. Long non-coding RNA ZEB1-AS1 promotes glioma cell proliferation, migration and invasion through regulating miR-577. Eur Rev Med Pharmacol Sci. 2018;22(10):3085–3093. doi:10.26355/eurrev_201805_15068
26. Yan Q, Yu HL, Li JT. Study on the expression of BDNF in human gliomas. Sichuan Da Xue Xue Bao Yi Xue Ban. 2009;40(3):415–417.
27. Xiong J, Zhou L, Lim Y, et al. Mature BDNF promotes the growth of glioma cells in vitro. Oncol Rep. 2013;30(6):2719–2724. doi:10.3892/or.2013.2746
28. Sahlu BW, Zhao S, Wang X, et al. Long noncoding RNAs: new insights in modulating mammalian spermatogenesis. J Anim Sci Biotechnol. 2020;11(1):16. doi:10.1186/s40104-019-0424-8
29. Ghafouri-Fard S, Esmaeili M, Taheri M. Expression of non-coding RNAs in hematological malignancies. Eur J Pharmacol. 2020;875:172976. doi:10.1016/j.ejphar.2020.172976
30. Hu X, Hong Y, Shang C. Knockdown of long non-coding RNA SNHG5 inhibits malignant cellular phenotypes of glioma via Wnt/CTNNB1 signaling pathway. J Cancer. 2019;10(5):1333–1340. doi:10.7150/jca.29517
31. Gu J, Xu F, Dang Y, Bu X. Long non-coding RNA 001089 is a prognostic marker and inhibits glioma cells proliferation and invasion. Clin Lab. 2019;65:3. doi:10.7754/Clin.Lab.2018.180817
32. Fang J, Huang J. Clinical significance of the expression of long non-coding RNA PVT1 in glioma. Cancer Biomark. 2019;24(4):509–513. doi:10.3233/CBM-182253
33. Li J, Zhu Y, Wang H, Ji X. Targeting long noncoding RNA in glioma: a pathway perspective. Mol Ther Nucleic Acids. 2018;13:431–441. doi:10.1016/j.omtn.2018.09.023
34. Yao F, Wang Q, Wu Q. The prognostic value and mechanisms of lncRNA UCA1 in human cancer. Cancer Manag Res. 2019;11:7685–7696. doi:10.2147/CMAR.S200436
35. Abdollahzadeh R, Daraei A, Mansoori Y, Sepahvand M, Amoli MM, Tavakkoly-Bazzaz J. Competing endogenous RNA (ceRNA) cross talk and language in ceRNA regulatory networks: a new look at hallmarks of breast cancer. J Cell Physiol. 2019;234(7):10080–10100. doi:10.1002/jcp.27941
36. Barde YA, Edgar D, Thoenen H. Purification of a new neurotrophic factor from mammalian brain. EMBO J. 1982;1(5):549–553. doi:10.1002/j.1460-2075.1982.tb01207.x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.