Back to Journals » OncoTargets and Therapy » Volume 12
LncSNHG3/miR-139-5p/BMI1 axis regulates proliferation, migration, and invasion in hepatocellular carcinoma
Authors Wu J, Liu L ![]() , Jin H, Li Q, Wang S, Peng B
, Jin H, Li Q, Wang S, Peng B
Received 30 November 2018
Accepted for publication 25 January 2019
Published 19 August 2019 Volume 2019:12 Pages 6623—6638
DOI https://doi.org/10.2147/OTT.S196630
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Federico Perche
Jian Wu,1,* Lingyun Liu,2,* Huilin Jin,1 Qiao Li,1 Shutong Wang,1 Baogang Peng1
1Department of Hepatic Surgery, The First Affiliated Hospital of Sun Yat-sen University, Guangzhou, People’s Republic of China; 2Department of Hepatobiliary and Pancreatic Surgery, Affiliated Hospital of Guilin Medical University, Guilin, People’s Republic of China
Correspondence: Baogang Peng
Department of Hepatic Surgery, The First Affiliated Hospital of Sun Yat-sen University, No.58, Zhongshan 2nd Road, Guangzhou 510080, People’s Republic of China
Tel +860 208 775 5766
Email [email protected]
*These authors contributed equally to this work
Objective: Emerging evidence has revealed that lncRNA small nucleolar RNA host gene 3 (SNHG3) is involved in cell proliferation, migration, and invasion in various tumors. However, the underlying molecular mechanism of SNHG3 in hepatocellular carcinoma (HCC) is still not fully explored.
Methods: Quantitative reverse transcriptase PCR was employed to detect the expression of SNHG3, miR-139-5p, and BMI1. Colony assay and MTT assay were used to detect the proliferation. Transwell assay was introduced to measure the migration and invasion ability. Bioinformatics analysis and luciferase reporter assay were used to confirm the relationship between SNHG3, miR-139-5p, and BMI1. An animal experiment was adopted to detect the function of SNHG3 in vivo.
Results: SNHG3 and BMI1 were upregulated in HCC, while miR-139-5p was downregulated. Knockdown of SNHG3 or BMI1 and overexpression of miR-139-5p could inhibit cell proliferation, migration, and invasion in HCC. miR-139-5p was a target of SNHG3 and BMI1 was a direct target mRNA of miR-139-5p. Silencing SNHG3 could impair the tumor progression in vivo.
Conclusion: The lncRNA SNHG3/miR-139-5p/BMI1 axis plays an important role in cell proliferation, migration, and invasion in HCC.
Keywords: SNHG3, miR-139-5p, BMI1, hepatocellular carcinoma
Introduction
Hepatocellular carcinoma (HCC) is commonly diagnosed and identified as a leading cause of cancer death. According to the cancer statistics in China in 2015,1 the incidence was about 466,100 and mortality was 422,100 annually, which makes it the third most deadly cancer. In the USA, it was estimated that about 41,000 cases and 29,000 deaths were caused by HCC in 2017.2 Although great efforts were made in HCC treatment and they revealed that many genes were involved in the progression, migration, invasion, and metastasis of HCC, the molecular mechanism of HCC progression is still poorly understood. Therefore, it is important to explore the underlying mechanism of HCC progression.
Recent studies have highlighted that long non-coding RNAs (lncRNAs) are related to cancer progression. Long non-coding RNAs are endogenous cellular RNAs which are more than 200 nucleotides in length. LncRNAs may regulate the cell processes by mediating transcription or post-transcription. It was reported that lncRNAs could interact with DNA, RNA, and protein to play the regulatory roles. Their dysregulation may lead to many diseases including cancers. For example, lncRNA p10247 overexpression is correlated with metastasis in breast cancer.3 Downregulated lncRNA-HOTAIR suppressed colorectal cancer proliferation, invasion, and migration.4 LncRNA MIR4435-2HG promoted lung cancer progression.5 LINC00844 regulates cell migration and invasion in prostate cancer.6 LncRNA TBILA may promote the epithelial–mesenchymal transition of non-small cell lung cancer.7 Besides the above-mentioned cancer types, lncRNAs also play roles in HCC. LncRNA FLVCR-AS1,8 HNF1A-AS1,9 HOXA11-AS,10 and lncRNA OGFRP111 were upregulated, while XLOC006926,12 CASC2,13 and lncRNA NKILA14 were downregulated in HCC. However, there are still many lncRNAs involved in HCC tumorigenesis and progression that need to be explored.
It has been underscored that lncRNAs act as competing endogenous RNAs for miRNAs in biological processes. miRNAs are small non-coding RNAs which are about 18–25 nucleotides in length. Previous studies demonstrated that miRNAs may mediate gene expression by binding the 3ʹUTR of specific mRNA15 and showed that miRNAs were involved in many physiological and pathological processes such as tumors. For instance, miR-214 suppressed ovarian cancer,16 serum miR-196a could be a biomarker in gastric cancer,17 miR-34a could inhibit liver cancer cell growth,18 and miRNA-139 downregulation was associated with HCC risk and survival.19 But the function of miRNA in HCC is not fully understood.
BMI1 encodes a ring finger protein which is a major component of the polycomb group complex 1. BMI1 and other components are involved in multiple regulatory mechanisms through chromatin remodeling. Some studies reported that BMI1 is an oncogene and aberrant expression in numerous cancers including non-small cell lung cancer,20 colorectal cancer,21 breast cancer,22 endometrial cancer,23 and HCC.24 Yet, the detail mechanism of how BMI1 was regulated in cancers is unclear.
In the present study, we aimed to explore the biological role of lncRNA small nucleolar RNA host gene 3 (SNHG3) in HCC. We found that SNHG3 could negatively regulate miR-139-5p. Meanwhile, BMI1 was a downstream target mRNA of miR-139-5p. So, we proposed that SNHG3 may regulate proliferation, migration, and invasion via regulating miR-139-5p and BMI1 in HCC.
Materials and methods
Patient samples
The ethical approval was obtained from the Research Ethics Committee of the First Affiliated Hospital of Sun-Yat-sen University (Guangzhou, China). Thirty-one specimens of cancerous tissue and 31 specimens of paracancerous tissue were taken from the HCC patients, and all patients signed the informed consent form, and this study was performed in accordance with the ethical standards detailed in the Declaration of Helsinki. No patients had undergone chemotherapy or radiotherapy before surgery. The samples were stored in liquid nitrogen for subsequent laboratory testing.
Cell culture
HCC cell lines Hep3B and HepG2 and immortalized human liver cell line L-02 were obtained from RiboBio Co. (Guangzhou, China). Cells were cultured in Dulbecco’s Modified Eagle’s Medium DMEM (Gibco, Carlsbad, CA, USA) with 10% FBS (HyClone, Logan, UT, USA) and 1% penicillin and streptomycin (Sigma-Aldrich, St Louis, MO, USA) at 37°C with 5% of CO2. Medium was changed every 2 days.
Cell transfection
For transient transfection, siSNHG3, miR-139-5p, pcDNA-SNHG3, miR-139-5p inhibitor, siBMI1, and the corresponding negative control were purchased from GenePharma Co., Ltd. (Shanghai, China). All oligos and plasmids were transfected into Hep3B or HepG2 cells using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions.
For stable transfection, shRNA targeting lnc SNHG3 and negative control were synthesized by GenePharma Co., Ltd. (Shanghai, China). These fragments were inserted into the lentiviral vector pLVXshRNA2 (Clontech, Mountain view, CA, USA). Then the lentiviral vectors were transfected into HEK293T cells with the Lenti-X HTX packaging system (Clontech) according to the protocol. The medium containing lentivirus was collected at 48 hours after transfection. Then the lentiviruses were added to the medium containing HepG2 cells. After that, the transfected cells were sorted with FITC fluorescence using a BD FACSMelody Cell Sorter (BD Biosciences, San Jose, CA, USA).
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from tissues and cells using TRIzol (Invitrogen, Carlsbad, CA, USA) and RNA concentration was detected by a NanoDropND-1000 spectrophotometer (Thermo Fisher Scientific, Rockford, IL, USA). cDNA was reverse transcribed using the TaqMan miRNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) for miRNA and Reverse Transcription Reagents (Applied Biosystems) for mRNA and lncRNA. SNHG3, miR-139-5p, and BMI1 expression were determined using SYBR® Green (Promega, Madison, WI, USA) according to the manufacturer’s protocol. A U6 small nuclear RNA (snRNA) was used as a reference gene for miR-139-5p, and 18S rRNA was used as a reference gene for SNHG3 and BMI1. Fluorescence was detected in the iQTM5 Multicolor Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). The relative expression of SNHG3, miR-139-5p, and BMI1 was calculated by the 2−ΔΔCt method. Primer sequences: SNHG3, forward 5ʹ-AGTGGTCGCTTCTTCTCCTTG-3ʹ and reverse 5ʹ-GATTGTCAAACCCTCCCTGTTA-3ʹ; miR-139-5p, forward 5ʹ-ACACTCCAGCTGGGTCTACAGTGCACGTG-3ʹ and reverse 5ʹ-CTCAACTGGTGTCGTGGA-3ʹ; BMI1, forward 5ʹ-GGTACTTCATTGATGCCACAAC-3ʹ and reverse 5ʹ-CTGCTGGGCATCGTAAGTATC-3ʹ;18S rRNA, forward 5ʹ-GGCCCTGTAATTGGAATGAGTC-3ʹ and reverse 5ʹ-CCAAGATCCAACTACGAGCTT-3ʹ; U6, forward 5ʹ-GTTGACATCCGTAAAGACC-3ʹ and reverse 5ʹ-GGAGCCAGGGCAGTAA-3ʹ.
Western blot
Cells were washed with Tris-buffered saline (TBS), and then added to RIPA buffer and maintained on ice for 30 minutes, accompanied with vortexing 30 seconds per 10 minutes. They were then centrifuged for 15 minutes at 4°C with 12,000 rpm and the supernatant was transferred to a new tube for next experiments. The protein concentrations were measured using Nanodrop2000 (Thermo Fisher Scientific, San Jose, CA, USA). Then the loading buffer was added to the protein solution and heated to 95°C for 5 minutes. Proteins (10 μg) were separated using SDS-PAGE and then transferred to polyvinylidene fluoride membranes (PVDF, Millipore, Bedford, MA, USA). After being blocked with 5% dried skimmed milk in TBS, the membranes were incubated with specific primary antibodies including anti-BMI1 (1:100 solution, and all of these antibodies were purchased from Santa Cruz Biotechnology Inc., CA, USA) and anti-β actin antibody (1:2,000 dilution, Santa Cruz Biotechnology Inc.) at 4°C overnight. The PVDF membrane was washed five times for 10 minutes each with Tris-buffered saline with Tween-20 (TBST), following incubation with horseradish peroxidase (HRP)-conjugated anti-mouse IgG secondary antibody (1:2,000 dilution, Santa Cruz Biotechnology Inc.) at room temperature for 1 hour. After being washed with TBST five times, the protein signals were detected using PierceTM ECL Western blotting substrate (ThermoFisher Scientific).
Colony formation assay
Transfected cells were trypsinized after 24 hours, counted, and replated at a density of 500 cells/6 cm dish. Ten days later, colonies resulting from the surviving cells were fixed with 3.7% methanol, stained with 0.1% crystal violet, and counted. Colonies containing at least 50 cells were scored. Each assay was performed three times.
MTT assay
Cell viability was measured with MTT assay kit (Sigma-Aldrich) according to the manufacturer’s protocol. In brief, cells (2×103) were seeded into 96-well plates (Corning Costar, Corning, NY, USA). Then 30 µL serum-free media with MTT solution was added into each well and incubated for 4 hours at 37°C. After discarding the media, 150 µL MTT solvent (4 mM HCl, 0.1% NP40 in isopropanol) was added into each well and then incubated for 3 hours at 37°C. Then the absorbance (A) value of each well was measured at OD=450 nm using the Microplate Reader (MG LABTECH, Durham, NC, USA).
Cell invasion and migration
Cell invasion analysis was performed using Transwell chamber with Matrigel (BD Biosciences, San Jose, CA, USA). Cell migration was displayed using a Transwell chamber without Matrigel. 1×105 cells were cultured in non-serum medium in the Transwell upper chamber. The Transwell lower chamber was added in DMEM medium with 10% FBS. After incubated for 24 hours at 37°C, cells on the upper chamber membrane were moved. Then the membrane was fixed with 90% ethyl alcohol and was stained with 0.1% crystal violet for 30 minutes. The number of cell migrations and invasions were counted using a phase contrast microscope.
Animal experiments
Animal experiments were carried out according to the National Institutes of Health guidelines to the Care and Use of Laboratory Animals of the First Affiliated Hospital of Sun-Yat-sen University. shSNHG3 and their negative control (shNC) were stably transfected into HepG2 cells using lentiviruses. Then 5×106 cells were subcutaneously injected in the flank of female nude mice (Beijing Vital River Laboratory Animal Technology Co., Ltd., China). There were five nude mice in each group. Tumor volumes were measured every 7 days until 35 days. All mice were sacrificed to collect tumor xenografts and tumor xenografts were weighted.
Immunohistochemistry
The tumor tissue from mice was fixed with 12 mL 4% paraformaldehyde (PFA) in PBS followed by being embedded with paraffin. Then the tissues were made into 4 μM sections and de-waxed in xylene, rehydrated in ethanol. Peroxidase blocking reagent was used to quench the endogenous peroxidase activity for 10 minutes. The sections were incubated overnight at 4°C with primary antibody (BMI1, ab38295, Abcam, Cambridge, MA, USA). Then washed with PBS and treated with HRP-conjugated secondary antibody at room temperature for 1 hour and developed with liquid diaminobenzidine substrate chromogen system.
Statistical analysis
The data are presented as mean±SD (standard deviation) from three biological replicated experiments. The results of analysis were shown and plotted using GraphPad Prism 7.0 (GraphPad Software, San Diego, CA, USA). All group comparisons were carried out using Student’s t-test. P-values less than 0.05 were regarded as statistically significant.
Results
SNHG3 was upregulated and miR-139-5p was downregulated in HCC
Firstly, we obtained HCC tumor tissues and adjacent tissues from 31 patients without any treatment except surgery. qRT-PCR was used to detect the expression of SNHG3 and the results showed that SNHG3 was upregulated significantly in HCC tissues than in normal tissues (Figure 1A). Then we continued to check SNHG3 expression in HCC cell lines. Consistent with that in tissues, SNHG3 was upregulated in Hep3B and HepG2 cells (Figure 1B). Meanwhile, we also checked miR-139-5p expression in HCC tissues and cell lines. The qRT-PCR results showed that miR-139-5p was downregulated in HCC tissues (Figure 1C), Hep3B and HepG2 cells (Figure 1D). Our finding suggested that upregulation of SNHG3 and downregulation of miR-139-5p might be involved in HCC.
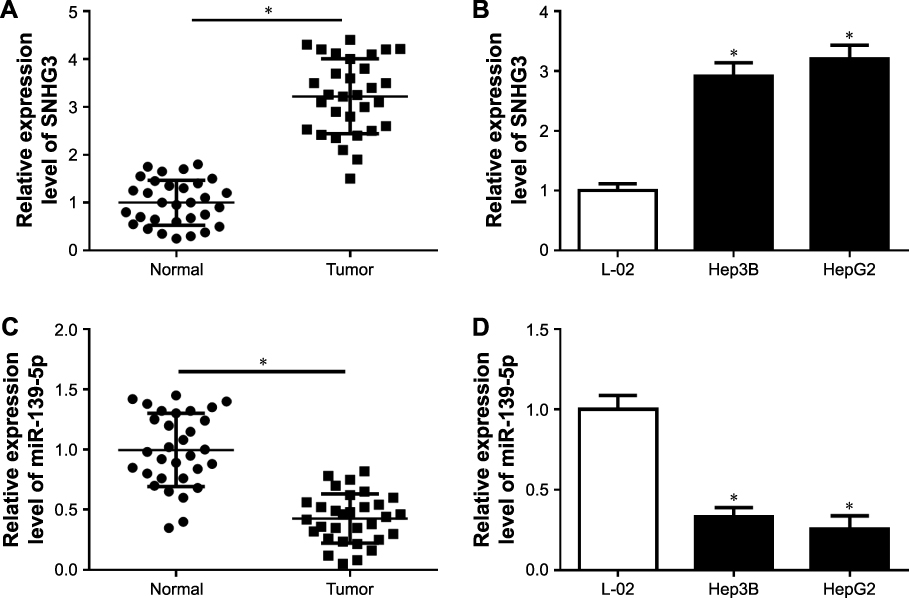 |
Figure 1 Expression of SNHG3 and hsa-mir-139-5p in HCC tissue and cells. (A) SNHG3 expression in normal and HCC tissues. (B) SNHG3 expression in L-02, Hep3B, and HepG2 cells. (C) miR-139-5p expression in normal and HCC tissues. (D) miR-139-5p expression in L-02, Hep3B, and HepG2 cells. *P<0.05. Abbreviations: HCC, hepatocellular carcinoma; SNHG3, small nucleolar RNAhost gene 3. |
Knockdown of SNHG3 inhibited cell proliferation, migration, and invasion in HCC
To better understand the function of SNHG3 in HCC, we downregulated SNHG3 via small interfering RNA in Hep3B and HepG2 cells. After 48 hours of transfection, the knockdown efficiency was checked and SNHG3 was indeed downregulated in Hep3B and HepG2 cells (Figure 2A). Then a colony formation assay was performed to detect the proliferation, and the colony number was significantly decreased after SNHG3 silencing (Figure 2B). Meanwhile, an MTT assay confirmed the inhibitory effect of SNHG3 in cell proliferation in HCC (Figure 2C and D). Next, we checked the function of SNHG3 in migration and invasion in HCC, and Transwell assay suggested that knockdown of SNHG3 significant inhibited the migration (Figure 2E) and invasion (Figure 2F) of Hep3B and Hep2G cells.
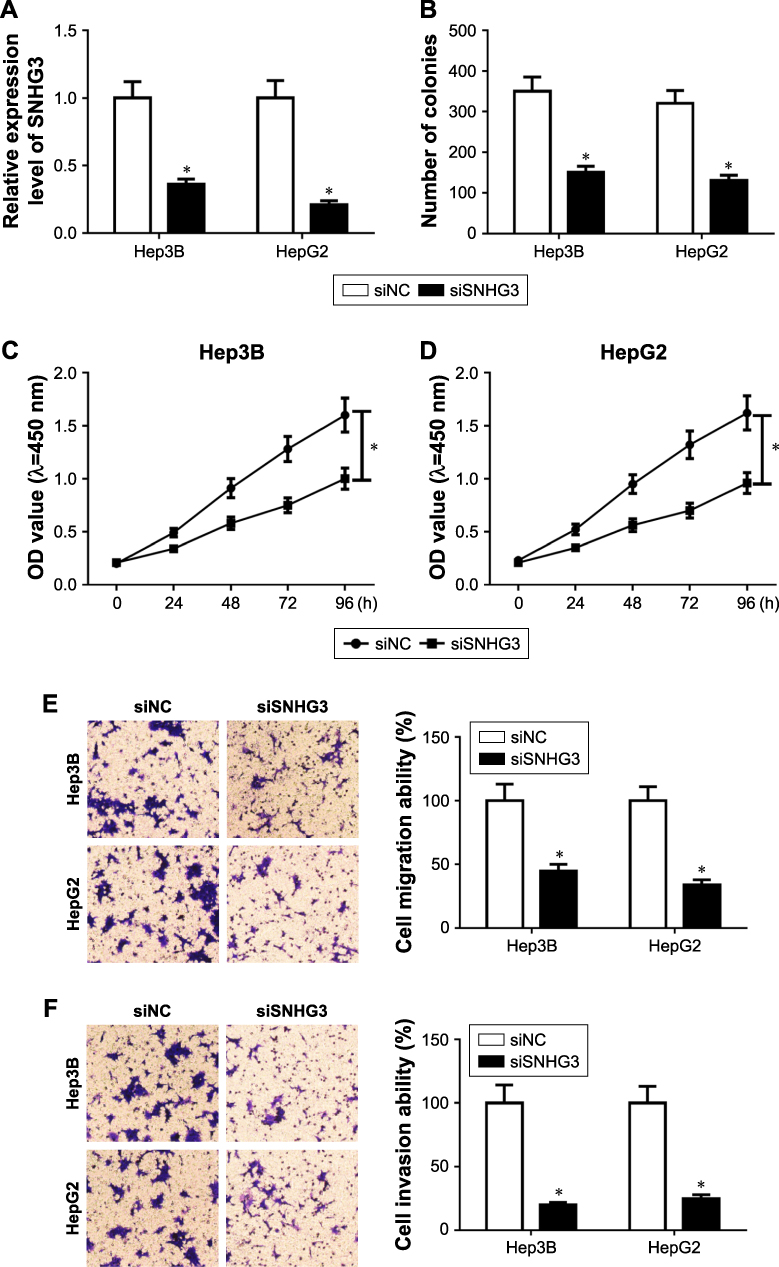 |
Figure 2 Effect of lncSNHG3 on the cell proliferation, migration and invasion in HCC. (A) SNHG3 knockdown in Hep3B and HepG2 cells. (B) Colony number of Hep3B and HepG2 transfected with siSNHG3. (C and D) MTT assay detected the proliferation of Hep3B (C) and HepG2 (D) after SNHG3 knockdown. (E) Cell migration analysis of Hep3B and HepG2 transfected with siNC and siSNHG3. Magnification 200×. (F) Cell invasion analysis of Hep3B and HepG2 transfected with siNC and siSNHG3. Magnification 200×. *P<0.05. Abbreviations: HCC, hepatocellular carcinoma; lnc, long non-coding; SNHG3, small nucleolar RNA host gene 3. |
Overexpression of miR-139-5p impaired cell proliferation, migration, and invasion in HCC
Considering that miR-139-5p was downregulated in HCC tissues and cells, we overexpressed miR-139-5p in Hep3B and HepG2 cells (Figure 3A). Both colony formation assay (Figure 3B) and MTT assay (Figure 3C and D) suggested that miR-139-5p overexpression inhibited the cell proliferation. In addition, the migration (Figure 3E) and invasion (Figure 3F) ability were impaired after miR-139-5p overexpression.
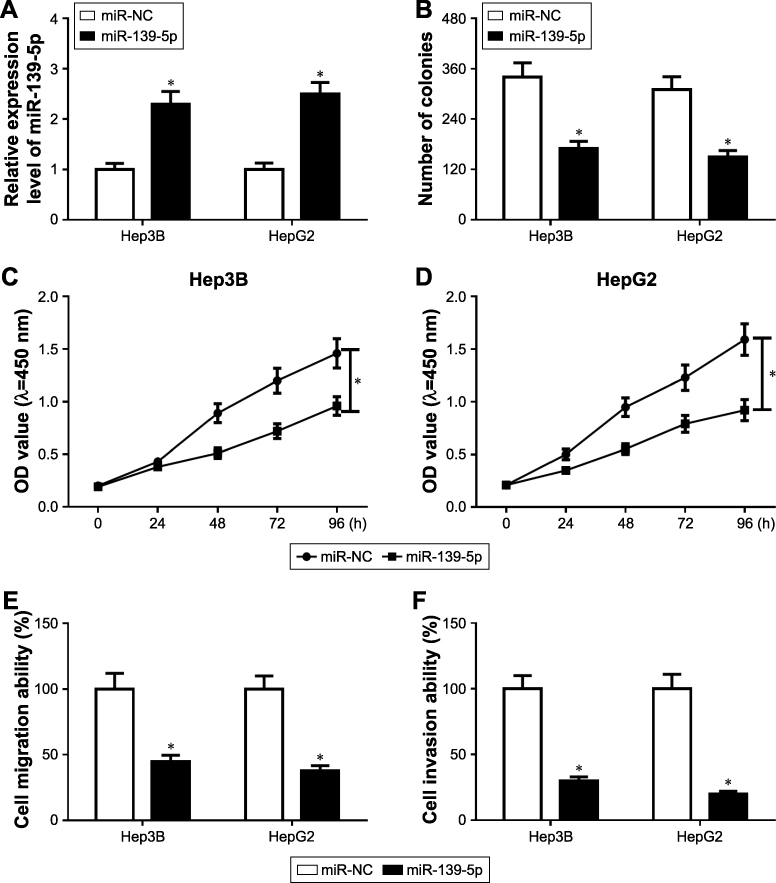 |
Figure 3 Effect of miR-139-5p on the cell proliferation, migration, and invasion in HCC. (A) miR-139-5p overexpression in Hep3B and HepG2 cells. (B) Colony number of Hep3B, and HepG2 transfected with miR-139-5p. (C and D) MTT assay detected the proliferation of Hep3B (C) and HepG2 (D) after miR-139-5p overexpression. (E) Cell migration analysis of Hep3B and HepG2 transfected with miR-NC and miR-139-5p. (F) Cell invasion analysis of Hep3B and HepG2 transfected with miR-NC and miR-139-5p. *P<0.05. Abbreviation: HCC, hepatocellular carcinoma. |
miR-139-5p was a target of SNHG3
Next, starBase was used to predict the relationship between SNHG3 and miR-139-5p. Bioinformatics analysis revealed that there were complementary sequence between lncSNHG3 and miR-139-5p (Figure 4A). Then we constructed luciferase report vector containing wildtype and mutant SNHG3. The luciferase activity was decreased when wildtype SNHG3 and miR-139-5p were transfected into Hep3B (Figure 4B) and HepG2 (Figure 4C) but not with mutant SNHG3. To further confirm the binding between SNGF3 and miR-139-5p, we introduced miR-139-5p inhibitor and the luciferase activity was upregulated about 2-fold when cotransfected with wildtype SNHG3 in Hep3B (Figure 4D) and HepG2 cells (Figure 4E). Subsequently, we overexpressed SNHG3 in Hep3B and HepG2 cells (Figure 4F) and miR-139-5p expression was inhibited after SNHG3 overexpression and increased when SNHG3 was silenced (Figure 4G). The results demonstrated that miR-139-5p was a target of SNHG3.
 |
Figure 4 miR-139-5p was a target of SNHG3. (A) The relationship between SNHG3 and miR-139-5p was predicted by starbase. (B and C) SNHG3-WT or SNHG3-MUT was cotransfected into Hep3B (B) and HepG2 (C) cells with miR-139-3p or their negative controls. (D and E) SNHG3-WT or SNHG3-MUT was cotransfected into Hep3B (D) and HepG2 (E) cells with miR-139-3p inhibitor or their negative controls. (F) SNHG3 overexpression in Hep3B and HepG2 cells. (G) miR-139-5p expression in pcDNA, pcDNA-SNHG3, siNC, and siSNHG3. *P<0.05. Abbreviation: SNHG3, small nucleolar RNA host gene 3. |
BMI1 was a direct target of miR-139-5p
Then, to investigate the mechanism of miR-139-5p in HCC, the starBase database was used to predict the downstream target. Luciferase reporter plasmids of wildtype BMI1 and mutant BMI1 were constructed (Figure 5A). Cotransfection of the luciferase reporter plasmid containing BMI1 3ʹUTR-WT with miR-139-5p in Hep3B and HepG2 resulted in a decreased luciferase activity, but not with mutant BMI1 (Figure 5B and C). The miR-139-5p inhibitor could increase the luciferase activity in Hep3B (Figure 5D) and HepG2 cells (Figure 5E).
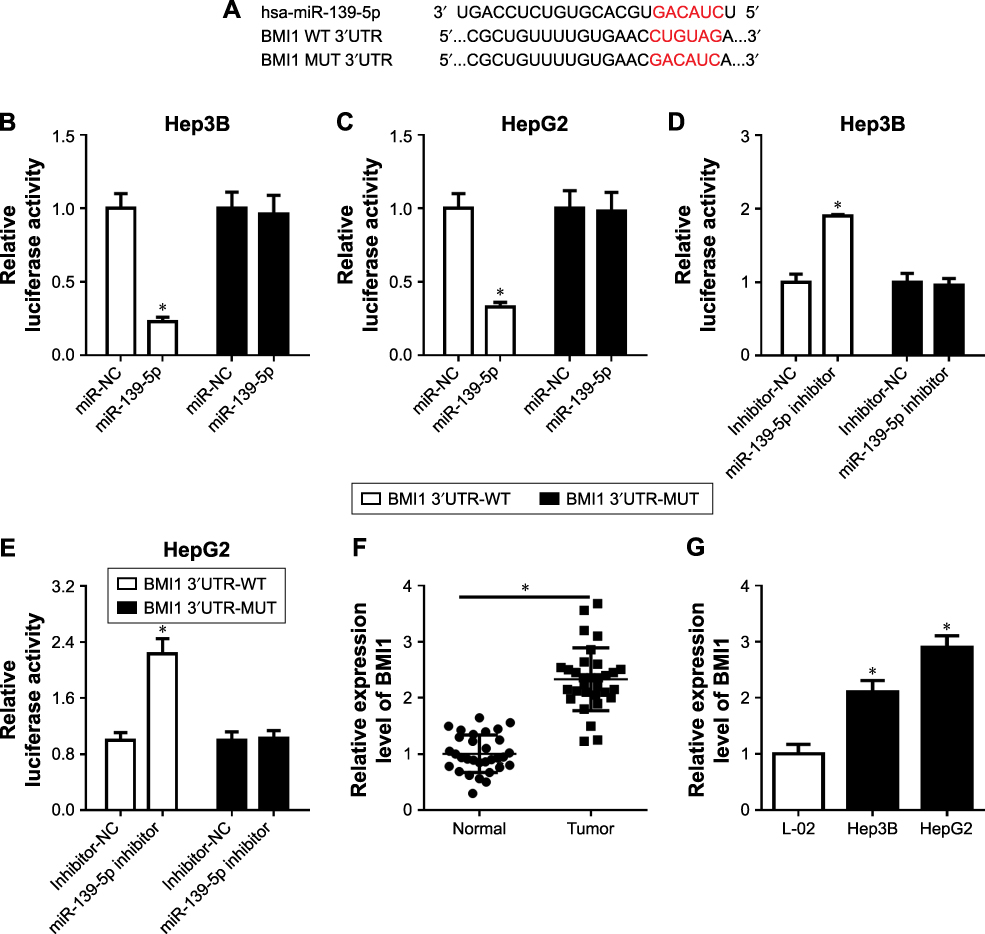 |
Figure 5 (Continued) |
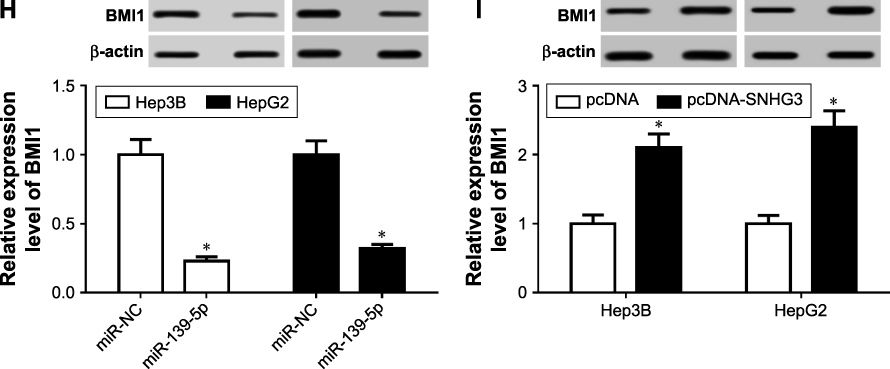 |
Figure 5 BMI1 was a direct target of miR-139-5p. (A) The relationship of miR-139-5p and BMI1 was predicted by starbase. (B and C) WT-BMI1 or MUT-BMI1 was cotransfected into Hep3B (B) and HepG2 (C) cells with miR-139-3p or their negative controls. (D and E) WT-BMI1 or MUT-BMI1 was cotransfected into Hep3B (D) and HepG2 (E) cells with miR-139-3p inhibitor or their negative controls. (F) BMI1 expression in HCC tissues. (G) BMI1 expression in LC-02, Hep3B, and HepG2 cells. (H) BMI1 expression under the condition of miR-139-5p overexpression in Hep3B and HepG2 cells. (I) BMI1 expression under the condition of SNHG3 overexpression in Hep3B and HepG2 cells. *P<0.05. Abbreviations: HCC, hepatocellular carcinoma; SNHG3, small nucleolar RNA host gene 3. |
Next, we checked the BMI1 expression in HCC tissues and cell lines. The mRNA expression level was upregulated in HCC tumor (Figure 5F) and cell lines (Figure 5G). Afterward, we overexpressed miR-139-5p in Hep3B and HepG2 cells and found that BMI1 was decreased (Figure 5H). Meanwhile, BMI1 expression was increased after SNHG3 overexpression (Figure 5I). These findings indicated that BMI1 can serve as a direct target of miR-139-5p.
BMI1 knockdown inhibited cell proliferation, migration, and invasion in HCC
To better understand the function of BMI1 in HCC, we downregulated BMI1 in Hep3B and HepG2 cells (Figure 6A and B). Cell proliferation was detected by colony formation assay (Figure 6C) and MTT assay (Figure 6D and E). The results showed that proliferation was inhibited by siBMI1. Moreover, cell migration (Figure 6F) and invasion (Figure 6G) ability were inhibited by BMI1 silencing in Hep3B and HepG2 cells.
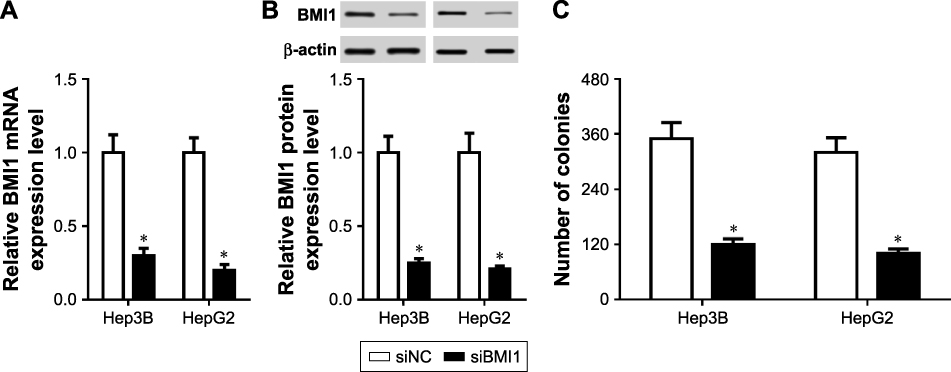 |
Figure 6 (Continued) |
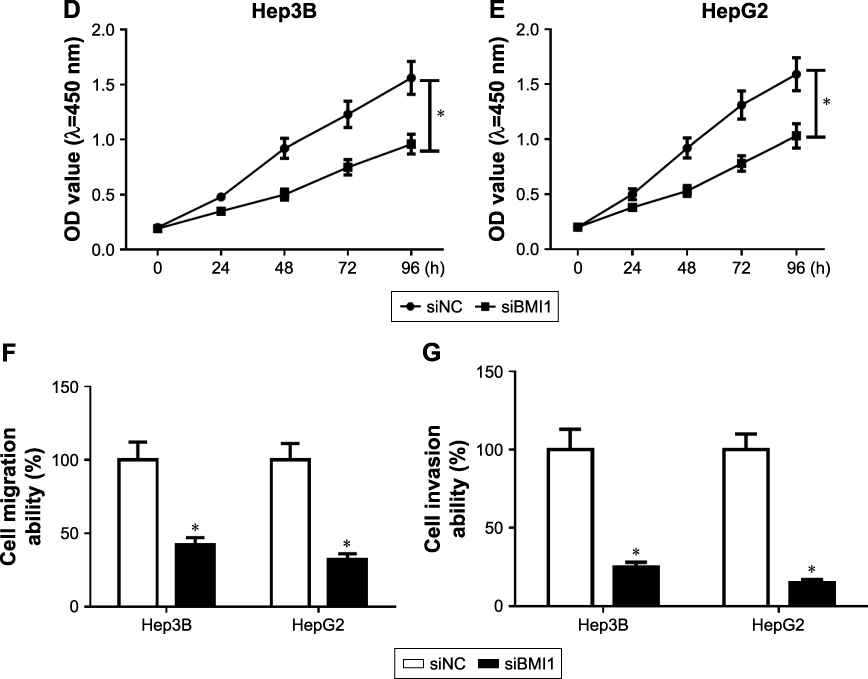 |
Figure 6 Effect of BMI1 on the cell proliferation, migration and invasion in HCC. (A and B) BMI1 knockdown in Hep3B and HepG2 cells. (C) Colonies number of Hep3B and HepG2 transfected with siBMI1. (D and E) MTT assay detected the proliferation of Hep3B (D) and HepG2 (E) after BMI1 knockdown. (F) Cell migration analysis of Hep3B and HepG2 transfected with siNC and siBMI1. (G) Cell invasion analysis of Hep3B and HepG2 transfected with siNC and siBMI1. *P<0.05. Abbreviation: HCC, hepatocellular carcinoma. |
SNHG3 knockdown restrained the tumor progression in vivo
To evaluate whether SNHG3 is involved in HCC progression in vivo, a HepG2 cell nude mouse xenograft model was used. The total nude mice were separated into two groups, one group was stably transfected with siNC and the other with siSNHG3. The tumor size was measured every 7 days until the sacrifice on day 35. Tumor growth was significantly inhibited by SNHG3 knockdown from day 21 (Figure 7A). On day 35, we measured the weight, and SNHG3 silencing obviously inhibited the tumor growth (Figure 7B). Meanwhile, we checked the SNHG3, miR-139-5p, and BMI1 expression in the tumor. qRT-PCR demonstrated that SNHG3 (Figure 7C) were downregulated and miR-139-5p (Figure 7D) was upregulated in the tumor. In addition, immunohistochemical assay revealed that BMI1 protein level was also decreased (Figure 7E). These data suggested that downregulation of SNHG3 could inhibit HCC development in vivo.
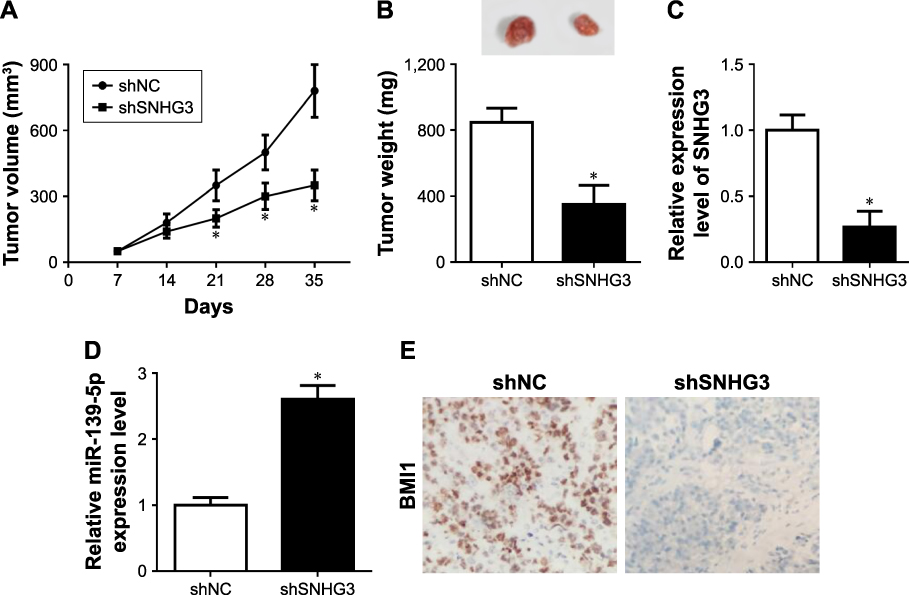 |
Figure 7 Down-regulation of SNHG3 repressed tumor progression in vivo. (A) Tumor volume was measured every 7 days after HepG2 stably transfected with siNC or siSNHG3 xenograft. (B) Tumor weight was measured on day 35. (C–F) SNHG3 (C), miR-139-5p (D), BMI1 mRNA (E), and BMI1 protein, magnification 200×. *P<0.05. Abbreviation: SNHG3, small nucleolar RNA host gene 3. |
Discussion
Over the past few decades, researchers claimed that non-coding transcripts are transcriptional noise.25 However, in recent years, whole genome and transcriptome sequencing suggested that non-coding RNAs (ncRNAs), including long non-coding RNAs (lncRNAs), may be involved in various cell processes,26 for example, epigenetics, alternative splicing, RNA decay, and translation.27–29 Given that massive lncRNAs have been discovered, the detailed function of these lncRNAs needs to be elucidated. Emerging evidence demonstrated that dysregulated lncRNAs were correlated with various cancers,30 for example, anti-NOS2A,31 BC200,32 HULC,33 H19,34 MALAT1,35 HOTAIR,36 etc. Previous reports demonstrated that SNHG3 play an important role in various cancer development. SNHG3 enhanced the malignant process of colorectal cancer.37 SNHG3 promoted proliferation and invasion in breast cancer38 and lung adenocarcinoma.39 In HCC, it was reported that SNHG3 was significantly upregulated and highly correlated with tumor size and relapse.40 In our present work, we found that SNHG3 was upregulated in HCC tissues compared with normal tissues, which was consistent with previous data. Dissimilarly, we knocked down the expression of SNHG3 using RNAi and found that the proliferation, migration, and invasion were inhibited in HCC cell lines in vitro. Meanwhile, we used nude mice and xenograft assay to verify the inhibitory function of SNHG3 in vivo. So we hypothesized that SNHG3 was involved in the progression of HCC. However, another study has demonstrated SNHG3 silencing in HCCLM3, which is a highly metastatic HCC cell line, and overexpression in PLC/PRF/5, which is one lowly metastatic HCC cell line which didn’t affect the cell proliferation.41 As shown in our supplemental material, we knocked down the expression of SNHG3 in HCCLM3, and the cell proliferation was inhibited (Figures S1–S4). We proposed that maybe the culture condition, knockdown efficiency, etc., were different in our two references, which led to distinct results. Besides, in other cancers such as breast cancer and lung cancer,38,39 SNHG3 knockdown inhibited cell proliferation.
Many studies revealed that miR-139-5p is involved in the cancer development, such as breast cancer,42 colorectal cancer, myeloid leukemia,44 etc. We found that miR-139-5p was downregulated in HCC. Through the bioinformatics database, we predicted that miR-139-5p is a potential target of SNHG3. Overexpression of miR-139-5p could impair the proliferation, migration, and invasion of Hep3B and HepG2 cells. Next, we performed bioinformatics analysis and luciferase reporter assay to show that BMI1 is a downstream mRNA target of miR-139-5p, which is consistent with another study.45 However, Luo et al showed the relationship between miR-139-5p and BMI1 in the bladder cancer stem cell.45 In their study, they suggested that BMI1 was a target and regulated by miR-139-5p, but didn’t explain BMI1 biological function in bladder cancer. However, in our study, we silenced BMI1 in HCC cells and found cell proliferation, migration, and invasion were attenuated, which was similar to miR-139-5p overexpression or SNHG3 knockdown.
BMI1 is a member of the polycomb group of proteins. Many studies have shown that BMI1 is upregulated in many cancers, including breast cancer,22 pancreatic cancer,46 and HCC.47 In our study, we found that BMI1 was upregulated in HCC tissues and cell lines. When knocking down the expression of BMI1, the proliferation, migration, and invasion of cells were inhibited.
Our present study revealed that SNHG3 can play an oncogenic role in the HCC development. We also observed that SNHG3 was upregulated in HCC tissues and cells, while miR-139-5p was downregulated. In addition, either SNHG3 knockdown or miR-139-5p overexpression could inhibit the proliferation, migration, and invasion in HCC cells. Bioinformatics analysis suggested that miR-139-5p was a target of SNHG3, and the expression of miR-139-5p was regulated by SNHG3. However, the accurate mechanism of regulatory function of SNHG3 in miR-139-5p expression is still needed to explore in the future study. Next, we predicted that BMI1 was a potential target mRNA of miR-139-5p, and BMI1 was upregulated in HCC tissues. The proliferation, migration, and invasion were impaired by BMI1 silencing. But, we cannot exclude that miR-139-5p may target other mRNAs and regulate HCC progression through unexplored pathways. Hence, exploring other potential mRNA targets will facilitate our understanding of miR-139-5p in HCC. Subsequently, we did the xenograft assay using HepG2 cells stably transfected with siSNHG3, and found that the tumor was significantly inhibited. In conclusion, it was indicated that lncSNHG3/miR-139-5p/BMI1 axis was involved in HCC proliferation, migration, and invasion.
Ethical statement
The ethical approval was obtained from the Research Ethics Committee of the First Affiliated Hospital of Sun-Yat-sen University (Guangzhou, China). Thirty-one specimens of cancerous tissue and 31 specimens of paracancerous tissue were collected from the HCC patients and all patients signed the informed consent form, and this study was performed in accordance with the ethical standards detailed in the Declaration of Helsinki. Animal experiments were conducted according to the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals, and approved by the Institutional Animal Care and Use Committee of the First Affiliated Hospital of Sun-Yat-sen University.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (Grant No 81702416) (to Dr Jian Wu) and the Sun Yat-sen University Clinical Research 5010 Program (Grant No 2015012) (to Dr Baogang Peng).
Author contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
2. Islami F, Miller KD, Siegel RL, Fedewa SA, Ward E, Jemal A. Disparities in liver cancer occurrence in the United States by race/ethnicity and state. CA Cancer J Clin. 2017;67(4):273–289. doi:10.3322/caac.21402
3. Yang YX, Wei L, Zhang YJ, et al. Long non-coding RNA p10247, high expressed in breast cancer (lncRNA-BCHE), is correlated with metastasis. Clin Exp Metastasis. 2018;32:1–13.
4. Lin K, Jiang H, Zhang LL, et al. Down-regulated LncRNA-HOTAIR suppressed colorectal cancer cell proliferation, invasion, and migration by mediating p21. Digest Dis Sci. 2018;63:1–12.
5. Qian H, Chen L, Huang J, et al. The lncRNA MIR4435-2HG promotes lung cancer progression by activating β-catenin signalling. J Mol Med. 2018;96:1–12.
6. Lingadahalli S, Jadhao S, Sung YY, et al. Novel lncRNA LINC00844 regulates prostate cancer cell migration and invasion through AR signaling. Mol Cancer Res. 2018;16(12):1865–1878. doi:10.1158/1541-7786.MCR-18-0087
7. Lu Z, Li Y, Che Y, et al. The TGFβ-induced lncRNA TBILA promotes non-small cell lung cancer progression in vitro and in vivo via cis-regulating HGAL and activating S100A7/JAB1 signaling. Cancer Lett. 2018;432:156–168. doi:10.1016/j.canlet.2018.06.013
8. Zhang K, Zhao Z, Yu J, Chen W, Xu Q, Chen L. LncRNA FLVCR1-AS1 acts as miR-513c sponge to modulate cancer cell proliferation, migration, and invasion in hepatocellular carcinoma. J Cell Biochem. 2018;119(7):6045–6059. doi:10.1002/jcb.26802
9. Ding CH, Yin C, Chen SJ, et al. The HNF1α-regulated lncRNA HNF1A-AS1 reverses the malignancy of hepatocellular carcinoma by enhancing the phosphatase activity of SHP-1. Mol Cancer. 2018;17(1):63. doi:10.1186/s12943-018-0813-1
10. Zhan M, He K, Xiao J, et al. LncRNA HOXA11‐AS promotes hepatocellular carcinoma progression by repressing miR‐214‐3p. J Cell Mol Med. 2018;22(8):3758–3767. doi:10.1111/jcmm.13633
11. Chen W, You J, Zheng Q, Zhu Y. Downregulation of lncRNA OGFRP1 inhibits hepatocellular carcinoma progression by AKT/mTOR and Wnt/β-catenin signaling pathways. Cancer Manag Res. 2018;10:1817–1826. doi:10.2147/CMAR.S164911
12. Fan H, Cheng S, Zhao L, et al. lncRNA XLOC006926 inhibits the proliferation, metastasis and invision of hepatocellular carcinoma. Immunol J. 2018;5:412–420.
13. Fan JC, Zeng F, Le YG, Xin L. LncRNA CASC2 inhibited the viability and induced the apoptosis of hepatocellular carcinoma cells through regulating miR-24-3p. J Cell Biochem. 2018;119(8):6391–6397. doi:10.1002/jcb.26479
14. Yu X, Tang W, Yang Y, et al. Long noncoding RNA NKILA enhances the anti-cancer effects of baicalein in hepatocellular carcinoma via the regulation of NF-κB signaling. Chem Biol Interact. 2018;285:48–58. doi:10.1016/j.cbi.2018.02.027
15. Yates LA, Norbury CJ, Gilbert RJ. The long and short of microRNA. Cell. 2013;153(3):516–519. doi:10.1016/j.cell.2013.04.003
16. Liu Y, Lin J, Zhai S, et al. MicroRNA-214 suppresses ovarian cancer by targeting β-catenin. Cell Physiol Biochem. 2018;45(4):1654–1662. doi:10.1159/000487733
17. Chen TH, Lee C, Chiu CT, et al. Circulating microRNA-196a is an early gastric cancer biomarker. Oncotarget. 2018;9(12):10317–10323. doi:10.18632/oncotarget.23126
18. Zhang HF, Wang YC, Han YD. MicroRNA-34a inhibits liver cancer cell growth by reprogramming glucose metabolism. Mol Med Rep. 2018;17(3):4483–4489. doi:10.3892/mmr.2018.8399
19. Li T, Yin J, Yuan L, et al. Downregulation of microRNA-139 is associated with hepatocellular carcinoma risk and short-term survival. Oncol Rep. 2014;31(4):1699–1706. doi:10.3892/or.2014.3032
20. Vonlanthen S, Heighway J, Altermatt HJ, et al. The bmi-1 oncoprotein is differentially expressed in non-small cell lung cancer and correlates with INK4A-ARF locus expression. Br J Cancer. 2001;84(10):1372–1376. doi:10.1054/bjoc.2001.1791
21. Kim JH, Yoon SY, Kim CN, et al. The Bmi-1 oncoprotein is overexpressed in human colorectal cancer and correlates with the reduced p16INK4a/p14ARF proteins. Cancer Lett. 2004;203(2):217–224.
22. Kim JH, Sun YY, Jeong SH, et al. Overexpression of Bmi-1 oncoprotein correlates with axillary lymph node metastases in invasive ductal breast cancer. Breast. 2004;13(5):383–388. doi:10.1016/j.breast.2004.02.010
23. Dong P, Kaneuchi M, Watari H, et al. MicroRNA-194 inhibits epithelial to mesenchymal transition of endometrial cancer cells by targeting oncogene BMI-1. Mol Cancer. 2011;10(1):99. doi:10.1186/1476-4598-10-99
24. Bartucci M, Hussein MS, Huselid E, et al. Synthesis and characterization of novel BMI1 inhibitors targeting cellular self-renewal in hepatocellular carcinoma. Target Oncol. 2017;12(4):1–14. doi:10.1007/s11523-016-0452-7
25. Van BH, Hughes TR. Establishing legitimacy and function in the new transcriptome. Brief Funct Genomic Proteomic. 2009;8(6):424–436. doi:10.1093/bfgp/elp037
26. Mattick JS. RNA regulation: a new genetics? Nat Rev Genet. 2004;5(5):316–323. doi:10.1038/nrg1321
27. Wapinski O, Chang HY. Long noncoding RNAs and human disease. Trends Cell Biol. 2011;21(6):354–361. doi:10.1016/j.tcb.2011.04.001
28. Noh JH, Kim KM, Mcclusky WG, Abdelmohsen K, Gorospe M. Cytoplasmic functions of long noncoding RNAs. Wiley Interdiscip Rev RNA. 2018;9(3):e1471. doi:10.1002/wrna.1471
29. Dhanasekaran K, Kumari S, Kanduri C. Noncoding RNAs in chromatin organization and transcription regulation: an epigenetic view. Subcell Biochem. 2013;61:343–372. doi:10.1007/978-94-007-4525-4_15
30. Huarte M, Rinn JL. Large non-coding RNAs: missing links in cancer? Human Mol Genet. 2010;19(2):152–161. doi:10.1093/hmg/ddq353
31. Korneev SA, Korneeva EI, Lagarkova MA, Kiselev SL, Critchley G, O’Shea M. Novel noncoding antisense RNA transcribed from human anti-NOS2A locus is differentially regulated during neuronal differentiation of embryonic stem cells. RNA. 2008;14(10):2030–2037. doi:10.1261/rna.1084308
32. Iacoangeli A, Lin YE, Muslimov IA, et al. BC200 RNA in invasive and preinvasive breast cancer. Carcinogenesis. 2004;25(11):2125–2133. doi:10.1093/carcin/bgh228
33. Panzitt K, Tschernatsch MMO, Guelly C, et al. Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology. 2007;132(1):330–342. doi:10.1053/j.gastro.2006.08.026
34. Berteaux N, Lottin S, Adriaenssens E, et al. Hormonal regulation of H19 gene expression in prostate epithelial cells. J Endocrinol. 2004;183(1):69–78. doi:10.1677/joe.1.05696
35. Jeffers LK, Duan K, Ellies LG, et al. Correlation of transcription of MALAT-1, a novel noncoding RNA, with deregulated expression of tumor suppressor p53 in small DNA tumor virus models. J Cancer Ther. 2013;4(4):774–786. doi:10.4236/jct.2013.43094
36. Rinn JL, Kertesz M, Wang JK, et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. doi:10.1016/j.cell.2007.05.022
37. Huang W, Tian Y, Dong S, et al. The long non-coding RNA SNHG3 functions as a competing endogenous RNA to promote malignant development of colorectal cancer. Oncol Rep. 2017;38(3):1402–1410. doi:10.3892/or.2017.5837
38. Chang L, Hu Z, Zhou Z, Zhang H. SNHG3 promotes proliferation and invasion by regulating the miR-101/ZEB1 axis in breast cancer. RSC Adv. 2018;8(27):15229–15240. doi:10.1039/C8RA02090F
39. Liu L, Ni J, He X. Upregulation of the long noncoding RNA SNHG3 promotes lung adenocarcinoma proliferation. Dis Markers. 2018;2018:5736716. doi:10.1155/2018/5736716
40. Zhang T, Cao C, Wu D, Liu L. SNHG3 correlates with malignant status and poor prognosis in hepatocellular carcinoma. Tumor Biol. 2016;37(2):2379–2385. doi:10.1007/s13277-015-4052-4
41. Zhang PF, Wang F, Wu J, et al. LncRNA SNHG3 induces EMT and sorafenib resistance by modulating the miR-128/CD151 pathway in hepatocellular carcinoma. Journal of cellular physiology. 2019;234(3):2788–2794.
42. Krishnan K, Steptoe AL, Martin HC, et al. miR-139-5p is a regulator of metastatic pathways in breast cancer. RNA. 2013;19(12):1767–1780. doi:10.1261/rna.042143.113
43. Song M, Yin Y, Zhang J, et al. MiR-139-5p inhibits migration and invasion of colorectal cancer by downregulating AMFR and NOTCH1. Protein Cell. 2014;5(11):851–861. doi:10.1007/s13238-014-0093-5
44. Emmrich S, Engeland F, Elkhatib M, et al. miR-139-5p controls translation in myeloid leukemia through EIF4G2. Oncogene. 2015;35(14):1822. doi:10.1038/onc.2015.247
45. Luo H, Yang R, Li C, et al. MicroRNA-139-5p inhibits bladder cancer proliferation and self-renewal by targeting the Bmi1 oncogene. Tumour Biol. 2017;39(7):1010428317718414. doi:10.1177/1010428317718414
46. Song W, Tao K, Li H, et al. Bmi-1 is related to proliferation, survival and poor prognosis in pancreatic cancer. Cancer Sci. 2010;101(7):1754–1760. doi:10.1111/j.1349-7006.2010.01577.x
47. Effendi K, Mori T, Komuta M, Masugi Y, Du W, Sakamoto M. Bmi-1 gene is upregulated in early-stage hepatocellular carcinoma and correlates with ATP-binding cassette transporter B1 expression. Cancer Sci. 2010;101(3):666–672. doi:10.1111/j.1349-7006.2009.01431.x
Supplemental materials
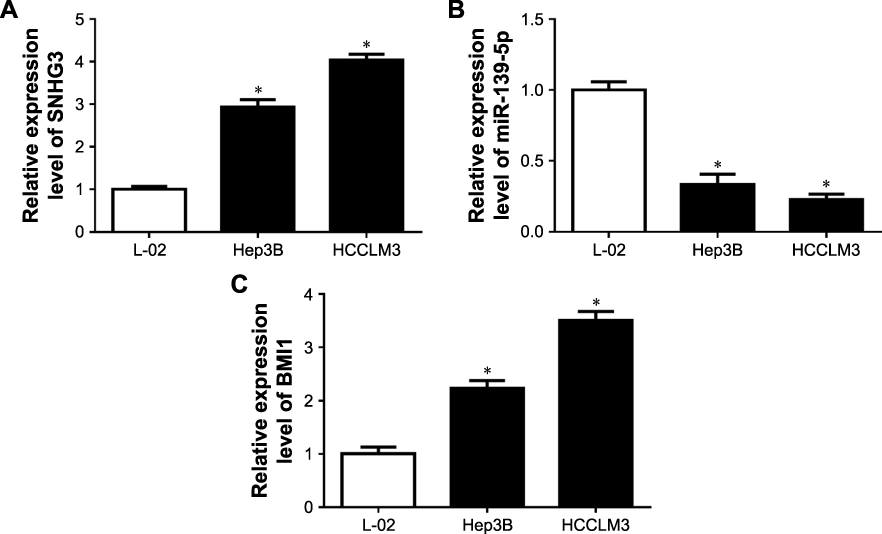 |
Figure S1 The expression of SNHG3, miR-139-5p and BMI1 in normal and HCC cells. (A) SNHG3 expression in L-01, Hep3B and HCCLM3 cells. (B) miR-139-5p expression in L-01, in Hep3B and HCCLM3 cells. (C) BMI1 expression in L-01, Hep3B and BMI1 cells. *P<0.05.Abbreviation: HCC, hepatocellular carcinoma. |
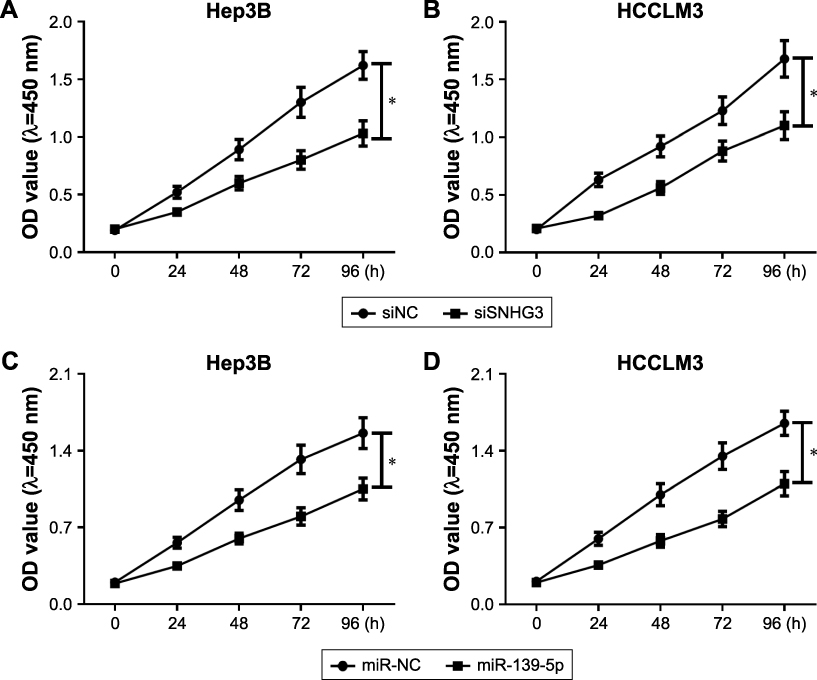 |
Figure S2 (Continued) |
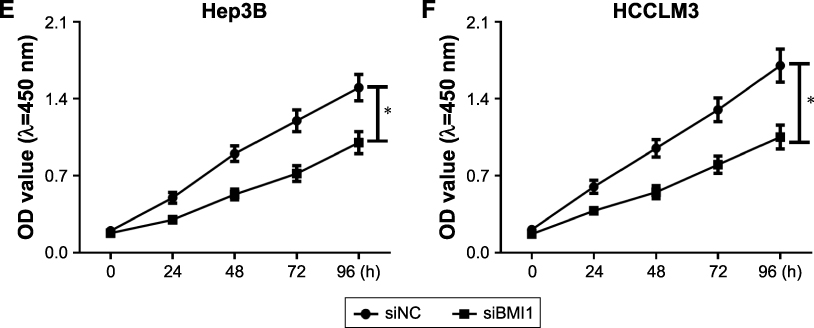 |
Figure S2 Effect of SNHG3, miR-139-5p and BMI1 on cell proliferation in HCC. MTT assay detected the proliferation of Hep3B (A) and HCCLM3 (B) after SNHG3 knockdown. MTT assay measured the proliferation of Hep3B (C) and HCCLM3 (D) after miR-139-5p overexpression. MTT assay evaluated the proliferation of Hep3B (E) and HCCLM3 (F) after BMI1 downregulation. *P<0.05.Abbreviations: HCC, hepatocellular carcinoma; MTT, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide. |
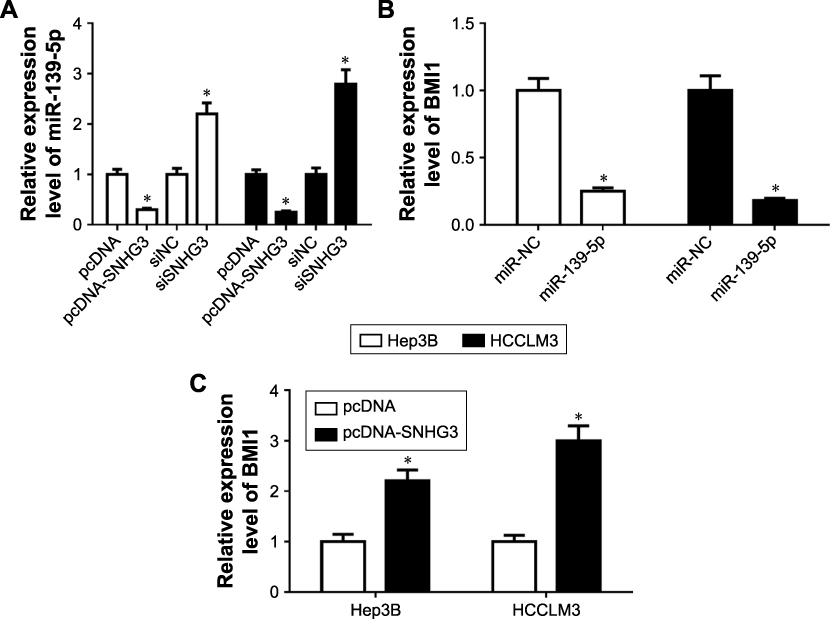 |
Figure S3 The expression of miR-139-5p and BMI1 in HCC cells. (A) miR-139-5p expression in Hep3B and HCCLM3 cells transfected with pcDNA, pcDNASNHG3, si-NC or siSNHG3. (B) BMI1 expression in Hep3B and HCCLM3 cells after miR-139-5p overexpression. (C) BMI1 expression in Hep3B and HCLM3 cells after SNHG3 overexpression. *P<0.05. Abbreviation: HCC, hepatocellular carcinoma. |
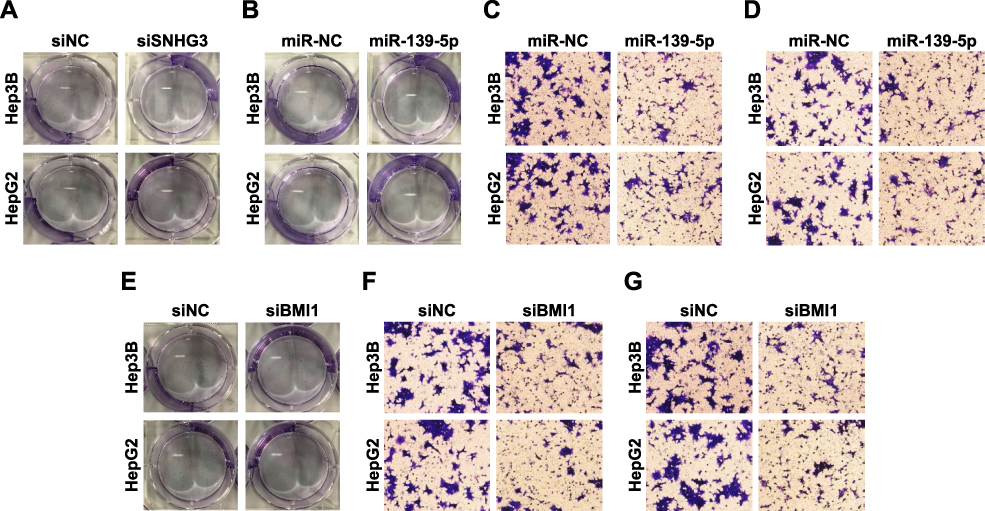 |
Figure S4 SiNHG3 regulated cell migration and invasion by targeting miR-139-5p/BMI1 axis. (A) Colony formation in HepG2 and Hep3B cells transfected with siNC and siSNHG3. (B) Colony formation in HepG2 and Hep3B cells transfected with miR-NC and miR-139-5p. (C, D) Cell migration and invasion of HepG2 and Hep3B cells transfected with miR-NC and miR-139-5p. (E) Colony formation in HepG2 and Hep3B cells transfected with siNC and siBMI1. (F, G) Cell migration and invasion of HepG2 and Hep3B cells transfected with siNC and siBMI1. Magnification 200×.Abbreviation: HCC, hepatocellular carcinoma. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.