Back to Journals » Journal of Inflammation Research » Volume 18
LncRNA NONHSAT248596.1 Promotes Osteoarthritis via miR-146a-5p/CXCR4 Axis by Inducing Chondrocyte Apoptosis and Extracellular Matrix Degradation
Authors Yang X, Yang G, Yang T, Song E, Xiang Y, Shi Z, Yu Y, Wang G, Li Y ![]()
Received 6 August 2025
Accepted for publication 3 December 2025
Published 10 December 2025 Volume 2025:18 Pages 17235—17252
DOI https://doi.org/10.2147/JIR.S558704
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ujjwol Risal
Xianguang Yang,1 Guang Yang,2 Tengyun Yang,3 En Song,1 Yaoyu Xiang,1 Zhengliang Shi,1 Yue Yu,1 Guoliang Wang,1 Yanlin Li1
1Department of Sports Medicine, The First Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China; 2Department of Joint and Sports Medicine, The First People’s Hospital of Qujing City, Qujing, Yunnan, People’s Republic of China; 3Orthopedic Department, The Second Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China
Correspondence: Yanlin Li, Department of Sports Medicine, The First Affiliated Hospital of Kunming Medical University, No. 295 Xichang Road, Kunming, Yunnan, 650032, People’s Republic of China, Email [email protected] Guoliang Wang, Department of Sports Medicine, The First Affiliated Hospital of Kunming Medical University, No. 295 Xichang Road, Kunming, Yunnan, 650032, People’s Republic of China, Email [email protected]
Objective: Osteoarthritis (OA) is characterized by articular cartilage degeneration, osteophyte formation, subchondral bone remodeling, synovitis, and joint capsule fibrosis, yet its underlying molecular mechanisms and the functional roles of long non-coding RNAs (lncRNAs) remain poorly understood. This study aimed to investigate the role of lncRNA NONHSAT248596.1 in the pathogenesis of OA through the miR-146a-5p/CXCR4 axis.
Materials and Methods: The experimental systems included human articular cartilage samples, a rabbit model of OA, and chondrocytes treated with 100 ng/mL SDF-1. DIANA software predicted lncRNAs targeting the miR-146a-5p/CXCR4 axis. Dual-luciferase reporter assay verified the molecular interactions. The expression levels of target genes and proteins (CXCR4, miR-146a-5p, NONHSAT248596.1, TNF-α, IL-1β, MMP-13, collagen II, and aggrecan) were analyzed using qRT-PCR, Western blotting, and ELISA both in vitro and in vivo. Cell viability and apoptosis were measured by CCK-8 assay and flow cytometry, respectively. Cartilage pathology in the rabbit models was assessed via safranin O and hematoxylin and eosin staining at 4, 8, and 12 weeks.
Results: LncRNA NONHSAT248596.1 was significantly upregulated in OA patients and in SDF-1-induced chondrocytes. The dual-luciferase assays confirmed that NONHSAT248596.1 interacted with miR-146a-5p, and miR-146a-5p interacted with CXCR4 in OA. Overexpression of NONHSAT248596.1 promoted chondrocyte apoptosis (p< 0.0001, n=3), upregulated MMP-13 (p< 0.0001, n=3), and downregulated collagen II (p< 0.0001, n=3) and aggrecan (p< 0.01, n=3), while exacerbating cartilage degradation (p< 0.05, n=3/group/time point). Silencing NONHSAT248596.1 or overexpressing miR-146a-5p reduced chondrocyte apoptosis and extracellular matrix degradation in vitro, which also ameliorated cartilage degradation (Mankin score: 5.9± 1.7 VS 2.4± 0.59, p< 0.0001, n=3/group/time point) in rabbit OA models.
Conclusion: LncRNA NONHSAT248596.1 promotes OA via miR-146a-5p/CXCR4 axis by inducing chondrocyte apoptosis and extracellular matrix degradation. These results identify NONHSAT248596.1 as a potential therapeutic target for OA intervention and underscore the miR-146a-5p/CXCR4 axis as a critical regulatory pathway for cartilage protection.
Keywords: osteoarthritis, competing endogenous RNA, lncRNA NONHSAT248596.1, CXCR4, miR-146a-5p, chondrocyte apoptosis, extracellular matrix degradation
Introduction
Osteoarthritis (OA) is a highly prevalent chronic disorder, affecting over 300 million individuals globally and posing a substantial burden on public health and healthcare systems worldwide.1 Primary risk factors for OA include age, body size, ethnicity, and genetic predisposition.2 The disease is characterized by articular cartilage degeneration, osteophyte formation at joint margins, subchondral bone remodeling, synovitis, and thickening of the joint capsule.3 Despite its high prevalence, the precise molecular mechanisms underlying OA pathogenesis remain incompletely elucidated.4
Long non-coding RNAs (lncRNAs) are a class of non-coding RNA molecules with transcripts longer than 200 nucleotides that lack protein-coding capacity but possess specialized molecular and cellular functions.5 Accumulating evidence implicates lncRNAs in the pathogenesis of various diseases,6,7 including OA, where multiple lncRNAs have been shown to regulate disease progression.8 These transcripts participate in epigenetic regulation and modulate transcriptional and post-transcriptional processes, playing pivotal roles in cartilage development, chondrocyte differentiation, apoptosis, and metabolic homeostasis.9 Their functional significance extends to the pathological alterations characteristic of OA joints.10 In our previous study,11 we employed sequencing technology to analyze SDF-1-induced OA chondrocyte models and identified numerous differentially expressed lncRNAs associated with the disease. Stromal cell-derived factor-1 (SDF-1) functions as a potent inducer of cartilage matrix degradation,12 with its expression markedly elevated in OA tissues and correlated with disease severity.13 Inhibiting the SDF-1/CXCR4 axis-mediated crosstalk between cartilage and subchondral bone has been shown to prevent OA progression.14 Our previous work demonstrated that miR-146a-5p blocks the SDF-1/CXCR4 signaling pathway by targeting CXCR4, thereby delaying cartilage degeneration.15 While targeting SDF-1 activity represents a promising therapeutic strategy for OA, the underlying mechanisms of OA pathogenesis require further elucidation. The precise genomic annotation and functional significance of lncRNAs in OA remain largely unclear.
In this study, we hypothesized that lncRNA NONHSAT248596.1 functions as a competing endogenous RNA (ceRNA) by sponging miR-146a-5p, leading to the upregulation of CXCR4 and thereby promoting chondrocyte apoptosis and extracellular matrix (ECM) degradation in OA. We identified NONHSAT248596.1 as a lncRNA that is significantly upregulated in SDF-1-induced OA models. By analyzing previous next-generation sequencing data, we investigated the expression patterns and potential ceRNA network mechanisms of NONHSAT248596.1. These findings may contribute substantially to the diagnosis and prognostic prediction of OA, while also facilitating related drug discovery efforts.
Materials and Methods
Patients and Cartilage Tissue Samples
Cartilage samples, consisting solely of complete cartilage without subchondral bone from defective areas, were collected from 10 patients undergoing total knee replacement surgery for OA at the First Affiliated Hospital of Kunming Medical University.16 Inclusion criteria were: (1) age between 40 and 70 years; (2) clinical diagnosis of unilateral knee OA requiring total knee arthroplasty; (3) combined femoral condyle chondral injury, including ICRS grades I to III; and (4) provision of signed informed consent (for elderly participants or minors, consent was provided by legal guardians). Patients with a history of knee ligament reconstruction, total meniscectomy, joint infection, or rheumatoid arthritis were excluded. Additionally, healthy cartilage samples were obtained from an unloaded region of the femoral condyle from each of the same 10 OA patients.
The study protocol was approved by the Ethics Committee of the First Affiliated Hospital of Kunming Medical University (Approval No.: (2018) Ethical Review L No. 21; Date: July 25, 2018). All participants provided written informed consent. The investigation was conducted in accordance with the principles of the Declaration of Helsinki.
Chondrocyte Culture, Identification, and Treatment
The C28/I2 human chondrocyte cell line (Beina Chuanglian Biotechnology Institute, China; RRID: CVCL_0187) was cultured in Dulbecco’s Modified Eagle Medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, USA) and 1% penicillin-streptomycin (Gibco, USA) at 37°C in a humidified atmosphere of 5% CO2. All experiments utilized chondrocytes at passages 3 to 4. Cell line identity was confirmed by short tandem repeat profiling. To establish an in vitro OA model, chondrocytes were treated with 100 ng/mL stromal cell-derived factor-1 (SDF-1; PeproTech, USA) for 24 hours.16
LncRNA Overexpression and Transfection
LncRNA NONHSAT248596.1 (18 kb; NONCODE) exceeds conventional viral vector capacity (<10 kb). Therefore, a dCas9-sgRNA dual-target CRISPR activation (CRISPRa) system was used to activate its endogenous promoter, mimicking its upregulation in OA. Key procedures included:
sgRNA Design for NONHSAT248596.1 Promoter
The target region was the core promoter of NONHSAT248596.1 (chr1: 152,340,000–152,341,000; hg38, UCSC Genome Browser), which contains RNA polymerase II-binding elements. Two sgRNA pairs (09668–1 and 09669–1) were designed using the Harvard Zhang Lab tool (http://sam.genome-engineering.org/), prioritizing sequences with high predicted on-target efficiency (off-target score <5). Each sgRNA included a 20-nt sequence complementary to the promoter and 5′-CACC/3′ overhangs for cloning (Table 1). Using dual sgRNA pairs enhanced dCas9 binding and transcriptional activation.
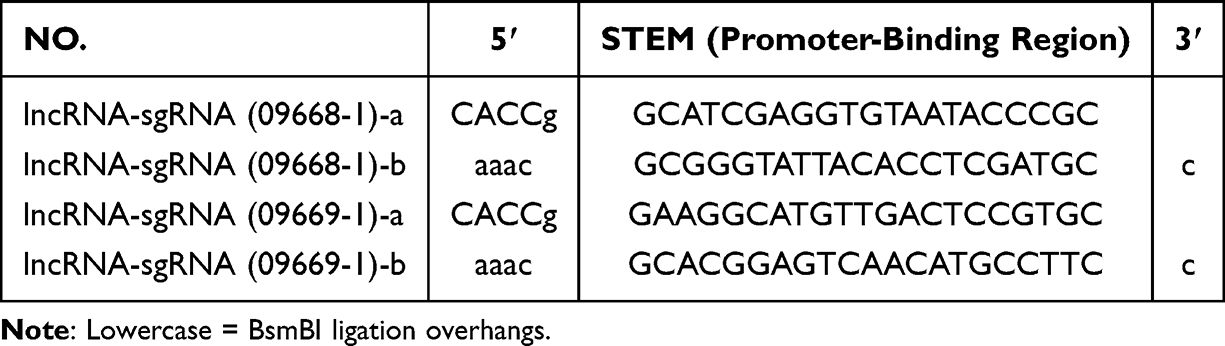 |
Table 1 LncRNA NONHSAT248596.1 siRNA Sequences |
ssDNA Oligo Synthesis & Annealing
sgRNA sequences (Table 1) were synthesized as single-stranded DNA (ssDNA) oligos (Shanghai GenePharma Co) and purified by PAGE. The oligos were dissolved in RNase-free water to a concentration of 100 μM for storage at −20°C. For annealing, complementary ssDNA pairs were combined in a 20-μL reaction containing 1 μL of each forward and reverse ssDNA (100 μM), 2 μL of 10× annealing buffer, and 16 μL of RNase-free water. The thermal cycling program was: 95°C for 5 minutes, followed by cooling to 25°C at a rate of 0.5°C per minute. This process generated double-stranded DNA (dsDNA) fragments with 4-nt sticky ends suitable for vector ligation.
Lentiviral Vector Construction
The dsDNA fragment was cloned into the BsmBI-digested LV-sgRNA-MS2-P65-HSF1-Neon vector (Addgene #61425), which encodes an MS2-P65-HSF1 transcriptional activator and a neomycin resistance gene. The vector was digested with BsmBI (New England Biolabs) at 37°C for 2 hours and purified using a Qiagen gel extraction kit. The purified vector (50 ng) was ligated with 10 ng of dsDNA using 1 μL of T4 DNA ligase at 16°C overnight. A 10 μL aliquot of the ligation product was added to 100 μL of competent E. coli DH5α cells (TransGen). The mixture was incubated on ice for 30 minutes, subjected to a 42°C heat shock for 90 seconds, and then recovered at 37°C for 1 hour in LB medium without antibiotics. Transformed cells were plated on LB-agar plates containing 50 μg/mL neomycin and incubated overnight at 37°C. Single colonies were selected and cultured in LB medium with neomycin for 8 hours. Colony PCR was performed using vector-specific primers (F: 5′-GGTGGAATCATACTGGAACA-3′, R: 5′-TTAACTCTGGCAAAGTGGAT-3′) to verify insert presence, yielding a 150-bp product that was visualized on a 1.5% agarose gel. Positive clones were confirmed by Sanger sequencing (Shanghai Sunny). Validated clones served as the sgRNA lentiviral plasmids for subsequent experiments.
NONHSAT248596.1 siRNA Synthesis
Small interfering RNA (siRNA) targeting NONHSAT248596.1 and a negative control (NC) siRNA were designed and synthesized by Shanghai GeneChem Co., Ltd., based on a dCas9-sgRNA dual-target strategy consistent with the promoter-targeting logic used for sgRNA design. The siRNAs were synthesized as 21-nt double-stranded RNA molecules with 3′ dTdT overhangs and purified by high-performance liquid chromatography to >99% purity. Final products were adjusted to 20 μM in RNase-free water and stored at −80°C. The SAM dual-vector lentiviral system was used for transcriptional activation. For standard transduction, target cells were seeded into plates one day prior to transduction. On the day of transduction, dCAS9-VP64 lentivirus was added according to the experimental design. After 3 days, transduced cells were selected with puromycin. Following stable cell growth recovery, cells were transduced with one or more sgRNA-MS2-P65-HSF1-expressing lentiviruses and subsequently selected with G418 before harvesting for overexpression verification. Separately, NONHSAT248596.1 overexpression vectors were constructed by cloning its sequence into the pcDNA3.1 vector (Thermo Fisher Scientific).17 Additionally, 17 individual siRNAs targeting NONHSAT248596.1, siRNA negative control, miR-146a-5p mimic, miR-NC, miR-146a-5p inhibitor, and inhibitor negative control (anti-miR-NC) were synthesized by Gene Pharm (Shanghai, China).
For transfection, C28/I2 cells were seeded into 6-well plates and cultured until they reached approximately 60% confluence. The cells were then transfected with 40 nM of the respective oligonucleotides using Lipofectamine 2000 (Thermo Fisher Scientific) according to the manufacturer’s protocol, followed by a 24-hour incubation period.
Luciferase Reporter Assay
The potential binding sites between miR-146a-5p and NONHSAT248596.1 were predicted using LncBase Predicted v.2. The wild-type and mutant miR-146a-5p binding sequences of NONHSAT248596.1 were then cloned into the pmirGLO vector (Promega, Madison, WI, USA) to generate the corresponding luciferase reporter constructs. These constructs were co-transfected with either miR-146a-5p mimic or a negative control (miR-NC) into chondrocytes. Using a similar approach, the wild-type and mutant CXCR4 binding sequences of miR-146a-5p were cloned into pmirGLO vector (Promega) to generate corresponding luciferase reporter constructs, which were co-transfected with miR-146a-5p mimic or negative control (miR-NC) into chondrocytes. After 48 hours, luciferase activity was measured using the dual-luciferase reporter assay system (Promega).
Cell Counting Kit-8 Assay
Cell viability was assessed using a Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto, Japan) according to the manufacturer’s protocol. Briefly, transfected and non-transfected OA chondrocytes were seeded in triplicate into a 96-well plate. Following the addition of CCK-8 solution and incubation at 37°C for 2 hours, the absorbance at 450 nm was measured with a microplate reader (Thermo Fisher Scientific, USA). Cell viability was normalized to the respective control groups.
Flow Cytometry
Each experimental group was analyzed in triplicate. Cells were rinsed, resuspended in binding buffer, and stained with Annexin V-FITC and propidium iodide (PI) for 15 minutes in the dark. Apoptosis was then analyzed using a flow cytometer (BD FACSCelesta, USA) to distinguish early and late apoptotic cell populations.
Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from C28/I2 cells using TRIzol reagent (Life Technologies, USA). RNA concentration and purity were quantified with a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA). Complementary DNA (cDNA) was synthesized from the extracted RNA using a commercial cDNA synthesis kit (Servicebio, China). Quantitative real-time PCR was performed using the BIO-RAD CFX96 Touch system (Bio-Rad Laboratories, USA) with SYBR Green qPCR Master Mix (Genecopoeia, China). All primers were designed with Beacon Designer 7.90 software; sequences are provided in Table 2. Gene expression levels were calculated using the 2−ΔΔCt method based on obtained threshold cycle (Ct) values.
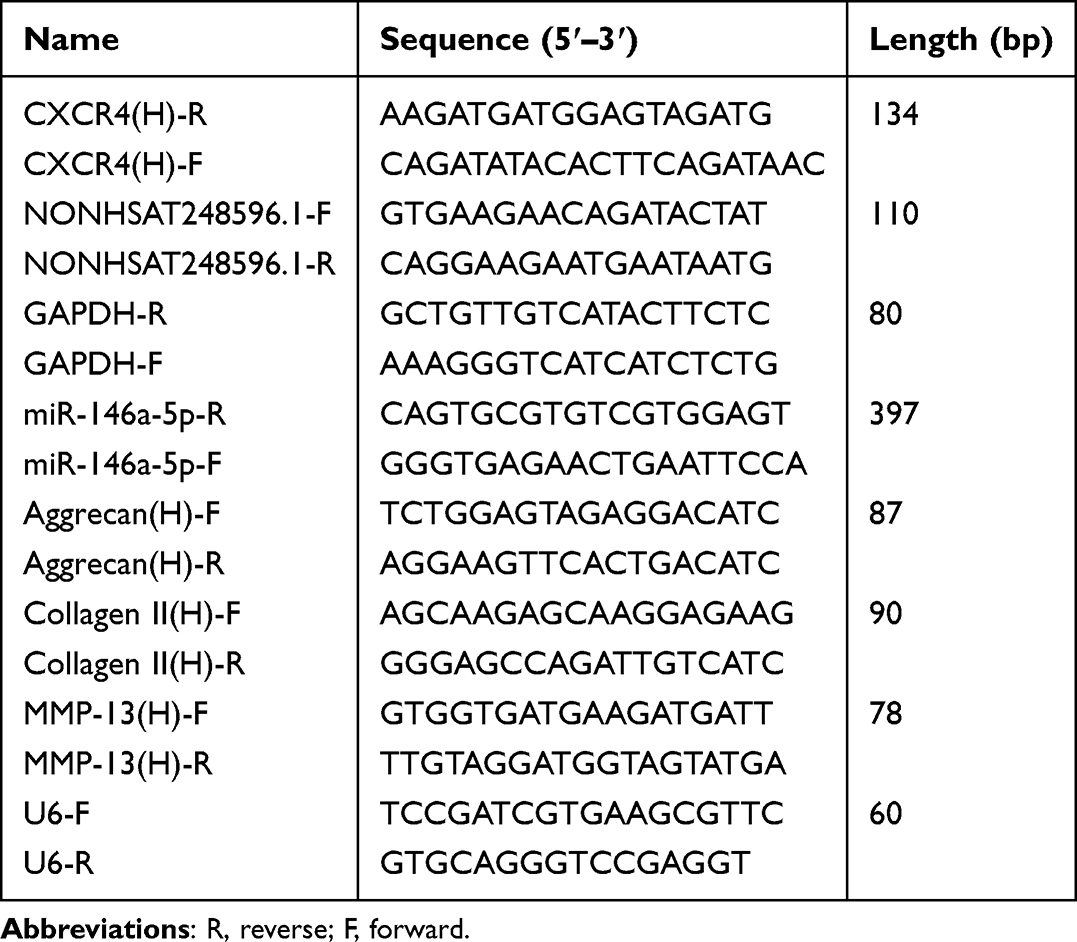 |
Table 2 Primer Sequences for qRT-PCR in vitro |
Enzyme-Linked Immunosorbent Assay (ELISA)
Levels of extracellular matrix-degrading factors in chondrocyte cultures were quantified using a commercial ELISA kit (Thermo Fisher Scientific, USA). Briefly, chondrocytes were seeded at a density of 105 cells per well. The culture supernatant was subsequently collected and analyzed for matrix metalloproteinase-13 (MMP-13) concentration according to the manufacturer’s instructions.
Rabbit Model of OA
Japanese white rabbits (3 months old, 2–2.5 kg) were obtained from the Laboratory Animal Center of Kunming Medical University. All animals were housed under controlled conditions: temperature 22 ± 1°C, relative humidity 50 ± 1%, and a 12-hour light/dark cycle, with ad libitum access to food and water. The experimental protocol was approved by the Experimental Animal Ethics Committee of Kunming Medical University (Approval No.: kmmu2020416; Date: April 1, 2020) and conducted in compliance with the national standard “Laboratory Animal - Guideline for Ethical Review of Animal Welfare” (GB/T 35892–2018, China).
Prior to the study, all rabbits underwent a one-week acclimatization period. OA was induced via intra-articular injection of SDF-1 solution (40 µg/kg in 200 µL; Beijing Yiqiao Shenzhou Technology Co., China)18 into the right posterior knee joint. Following the initial injection, two additional injections were administered on days 3 and 5, completing the three-injection modeling protocol.10 Thirty-six OA rabbits were randomly assigned to groups (n=9 per group, with 18 females and 18 males in total) using a random number table:
- lncRNA group: 10 μL of lentiviral vector overexpressing lncRNA (NONHSAT248596.1) and 10 μL of empty lentiviral vector without miR-146a-5p expression were injected into the right posterior knee.
- miRNA group: 10 μL of lentiviral vector overexpressing miR-146a-5p and 10 μL of empty lentiviral vector without lncRNA (NONHSAT248596.1) expression were injected into the right posterior knee.
- ceRNA group: 10 μL of lentiviral vector overexpressing both miR-146a-5p and lncRNA (NONHSAT248596.1) was injected into the right posterior knee.
- Control group: 10 μL of empty lentivirus vector without lncRNA (NONHSAT248596.1) expression and 10 μL of empty lentivirus vector without miR-146a-5p expression were injected into the right posterior knee.
Lentivirus vectors for gene overexpressing were purchased from Jiman Biotechnology Co. Ltd. (Shanghai, China) and administered via intra-articular injection. Humane endpoint criteria were defined as body weight loss below 1 kg or the onset of uncontrollable joint infection. No significant body weight changes were observed in the study. At 4, 8, and 12 weeks following the final injection, three rabbits per group were euthanized. Histological evaluation was performed on samples of the synovial membrane, the anterior half of the medial femoral condyle cartilage, and the underlying subchondral bone.
Cartilage Histology Staining
Following euthanasia by exsanguination under anesthesia, the right knee joints of the rabbits were dissected to expose the femoral condyles, which were visually examined and documented. Cartilage samples were harvested from the medial femoral condyle. The samples were then decalcified, dehydrated, cleared, and embedded in paraffin. Sections were stained with hematoxylin and eosin (H&E) and safranin O–fast green.
Modified Safranin O-Solid Green Cartilage Staining
Cartilage tissue samples from rabbits were routinely dewaxed and rehydrated. Sections were stained with Weigert’s hematoxylin for 5 minutes, followed by differentiation in an acid-alcohol solution for 15 seconds. After rinsing with distilled water, the sections were stained with a solid green solution for 5 minutes and then with a safranin O solution for an additional 5 minutes. Following whole-slide scanning, the stained sections were dehydrated, cleared, and mounted with a coverslip for observation.
ELISA of Rabbits OA
Synovial tissue samples from OA rabbits were collected and stored in 1 mL centrifuge tubes at 4°C. TNF-α, IL-1β, and SDF-1 levels in the synovial tissue were quantified using ELISA (Minnesota, USA). Absorbance at 450 nm was measured with a SPECTRA MAX 190 microplate reader (Molecular Devices, USA). A standard curve was generated using ELISA Calc software based on the known concentrations and optical densities of the standards, from which a regression equation was derived. The concentrations of TNF-α, SDF-1, and IL-1β in the samples were then determined using this regression equation.
qRT-PCR Analysis of Rabbits OA
Cartilage tissue (20 mg) was pulverized under liquid nitrogen and homogenized. Total RNA was isolated through a series of separation and purification steps, including a wash with pre-chilled 75% ethanol prepared with DEPC-treated water. After discarding the supernatant, the RNA pellet was dissolved in 20 µL of DEPC-treated water. cDNA was synthesized from the extracted RNA using a reverse transcription kit. The resulting cDNA was used to quantify the expression levels of NONHSAT248596.1, miR-146a-5p, MMP-13, CXCR4, aggrecan, and collagen II. QRT-PCR primers were designed with Beacon Designer 7.90 (sequences in Table 3), with GAPDH serving as the internal reference control. The amplification conditions were: initial denaturation at 95°C for 30 seconds; 40 cycles of denaturation at 95°C for 10 seconds, annealing at 60°C for 30 seconds; followed by melt curve analysis: 95°C for 15 seconds, 60°C for 60 seconds, and 95°C for 15 seconds. Gene expression was calculated using the 2−ΔΔCt method based on the Ct values.
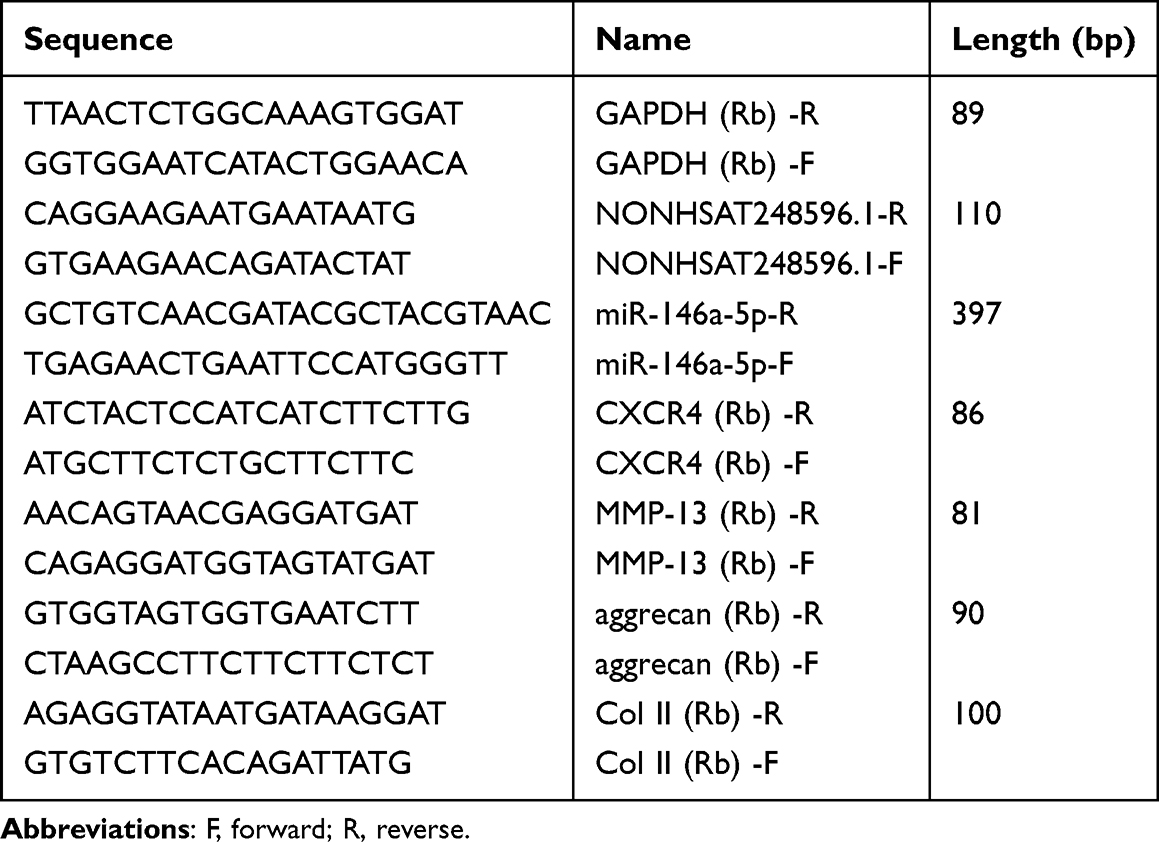 |
Table 3 Primer Sequences for qRT-PCR in vivo |
Western Blot Analysis
Proteins were extracted from chondrocytes or cartilage tissues using RIPA lysis buffer (Beyotime Biotech, China). Protein samples (20 µg per lane) were separated by 10% SDS-PAGE and transferred onto PVDF membranes (Millipore Sigma, USA). After blocking with 5% skim milk for 1 hour at room temperature, the membranes were incubated overnight at 4°C with the following primary antibodies: anti-aggrecan (1:1000, ab34861, Abcam, UK), anti-collagen II (1:800, ab36712, Abcam, UK), and anti-GAPDH (1:1000, P30008, Abmart, China). Subsequently, membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (1:2000, #7074, Cell Signaling Technology, USA) for 2 hours at room temperature. Protein bands were visualized using ECL detection reagents (Millipore Sigma, USA) and quantified with ImageJ software (version 1.5.0).
Statistical Analysis
All experiments were performed in triplicate using cells or animals from three independent preparations. Differences between two groups were assessed using Student’s t-test or Mann–Whitney U-test, as appropriate. Comparisons among multiple groups were analyzed by one-way ANOVA followed by Tukey’s post hoc test. Statistical analyses were conducted using GraphPad Prism software (version 10.2; GraphPad Software, La Jolla, CA, USA). A p-value of less than 0.05 was considered statistically significant.
Results
A Novel lncRNA NONHSAT248596.1 is Overexpressed in SDF-1 Induced OA
Our previous study employed sequencing technology to analyze SDF-1-induced OA in cell models.15 Bioinformatics prediction identified CXCR4 as a primary target of miR-146a-5p. In the present study, we utilized DIANA software to identify lncRNAs potentially targeting the miR-146a-5p/CXCR4 axis, which yielded 37 specific lncRNAs, including NONHSAT248596.1 (Figure 1A). Furthermore, by cross-referencing the top 15 differentially expressed and upregulated lncRNAs from our previous SDF-1-induced OA cell model, we identified four lncRNAs as putative targets of miR-146a-5p (Figure 1B). Subsequent qRT-PCR validation confirmed that the predicted targeting relationship was consistent for lncRNA NONHSAT248596.1 (Figure 1C). The significant differential expression of lncRNA NONHSAT248596.1 suggests its potential importance in SDF-1-induced OA, potentially mediated through the miR-146a-5p/CXCR4 axis.
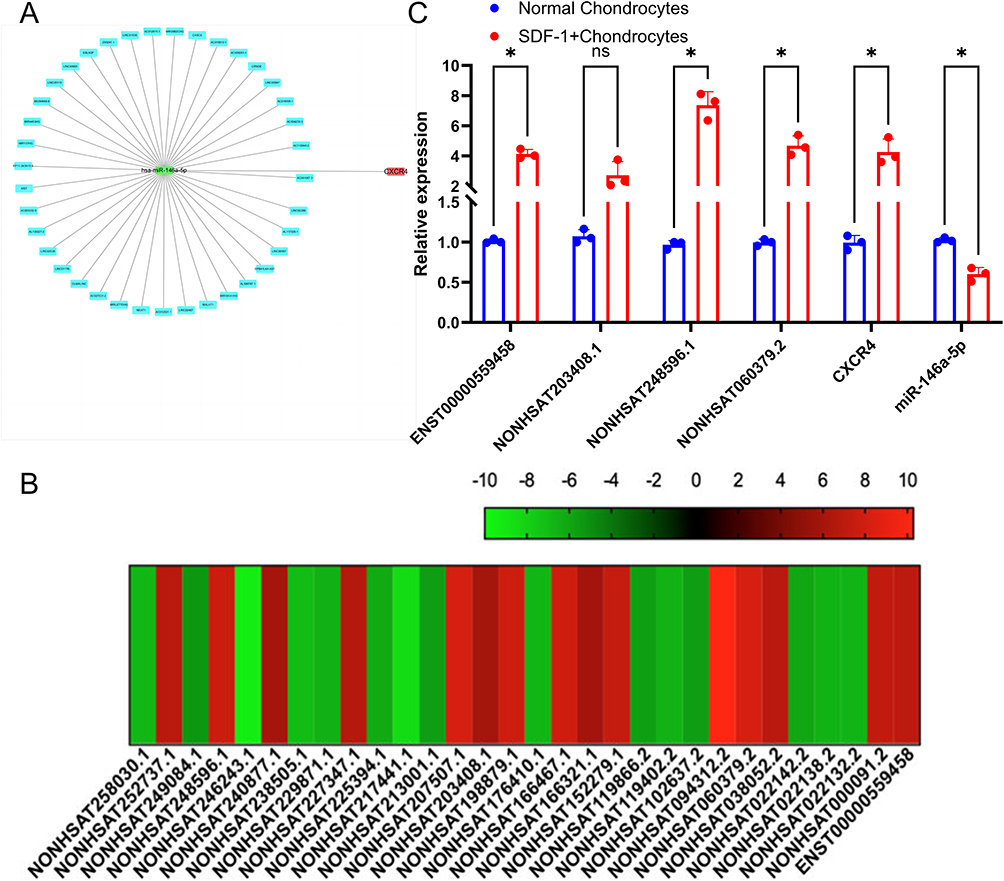 |
Figure 1 The novel lncRNA NONHSAT248596.1 is overexpressed in SDF-1-induced OA. (A) Construction of CXCR4-hsa-miR-146a-5p-lncRNA subnetworks. (B) High‑throughput sequencing results. NONHSAT248596.1, NONHSAT060379.2, and ENST00000559458 were identified as putative targets of miR‑146a‑5p. (C) Expression levels of miR‑146a‑5p, NONHSAT248596.1, and CXCR4. miR‑146a‑5p was significantly down‑regulated in the SDF‑1 group, while NONHSAT248596.1 and CXCR4 were significantly up‑regulated. (n=3, *p<0.05). |
NONHSAT248596.1 Interacts with miR‑146a-5p and miR‑146a-5p Interacts with CXCR4 in OA
LncRNAs can function as miRNA sponges.10 To investigate whether NONHSAT248596.1 interacts with miR-146a-5p in OA. First, qRT-PCR was used to verify the effectiveness of SDF-1 in inducing the OA chondrocyte model. The results showed that mRNA expression of both IL-1β and IL-6 was significantly upregulated (p < 0.05) in the OA-Chondrocyte group, indicating the successful establishment of the inflammatory OA model (Figure 2A). Then, we constructed wild-type NONHSAT248596.1, mutant NONHSAT248596.1, and miR-146a-5p plasmids for dual-luciferase reporter assays. Luciferase activity was measured in all experimental and control groups. A significant reduction in relative luciferase activity and fluorescence intensity was observed specifically in OA chondrocytes co-transfected with the wild-type NONHSAT248596.1 plasmid and the miR-146a-5p mimic (p < 0.0001). In contrast, no such changes were detected with the mutant plasmid or in other experimental groups (Figure 2B). These results indicate a specific binding interaction between miR-146a-5p and NONHSAT248596.1, suggesting that NONHSAT248596.1 functions through direct interaction with miR-146a-5p. Meanwhile, we constructed wild-type and mutant CXCR4, and miR-146a-5p plasmids for dual-luciferase reporter assays. Luciferase activity was measured in all groups, with significant reductions in relative luciferase activity and fluorescence intensity specifically in OA chondrocytes co-transfected with wild-type CXCR4 plasmid and miR-146a-5p mimic (p < 0.0001); no such changes were seen with the mutant plasmid (Figure 2C). These results also indicate a specific binding interaction between miR-146a-5p and CXCR4, suggesting that miR-146a-5p functions through direct interaction with CXCR4.
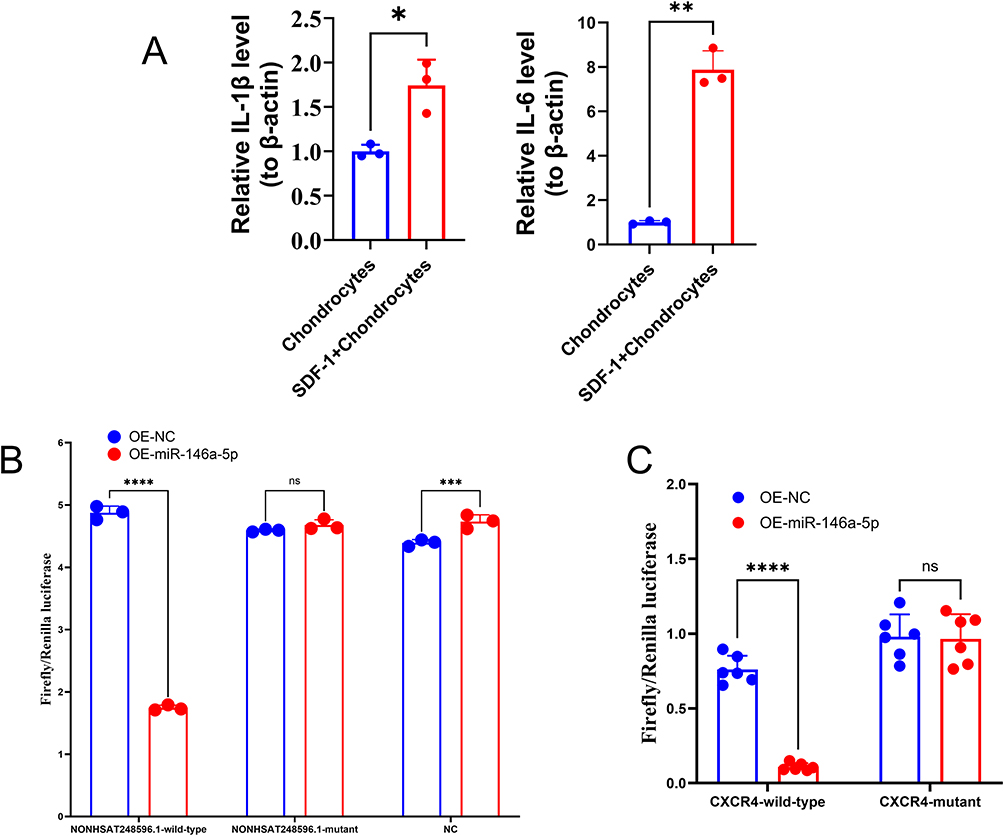 |
Figure 2 miR-146a-5p is downregulated in OA chondrocytes and directly targets lncRNA NONHSAT248596.1 and CXCR4. (A) Relative expression level of IL-1β and IL-6 Levels in normal chondrocytes and SDF-1-induced OA chondrocytes was detected by qRT-PCR. (B) Dual-luciferase reporter gene assay was performed to verify the binding relationship between miR-146a-5p and lncRNA NONHSAT248596.1. (C) Dual-luciferase reporter gene assay was conducted to confirm the targeting interaction between miR-146a-5p and CXCR4. (n=3/6, *p<0.05, **p<0.01, ***p<0.001, ****p < 0.0001). Abbreviation: ns, no significance. |
NONHSAT248596.1 Regulates CXCR4 by Acting as ceRNA Sponge for miR-146a-5p
To validate the NONHSAT248596.1/miR-146a-5p/CXCR4 ceRNA network, chondrocytes were transfected with either a NONHSAT248596.1 overexpression plasmid or a NONHSAT248596.1-specific interfering plasmid, followed by SDF-1 treatment to mimic OA conditions. qRT-PCR analysis confirmed that NONHSAT248596.1 expression was significantly increased by the overexpression plasmid, while the interfering plasmid effectively attenuated the SDF-1-induced upregulation (Figure 3A). Furthermore, NONHSAT248596.1 overexpression increased CXCR4 expression in OA chondrocytes, whereas NONHSAT248596.1 knockdown reduced CXCR4 levels (Figure 3B). In contrast, miR-146a-5p expression showed opposite trends to both NONHSAT248596.1 and CXCR4 across experimental groups (Figure 3C). In rescue experiments, both overexpression and knockdown of miR-146a-5p substantially abolished the regulatory effects of NONHSAT248596.1 on CXCR4 expression (Figure 3). These findings strongly suggest that NONHSAT248596.1 regulates CXCR4 expression through a miR-146a-5p-dependent mechanism.
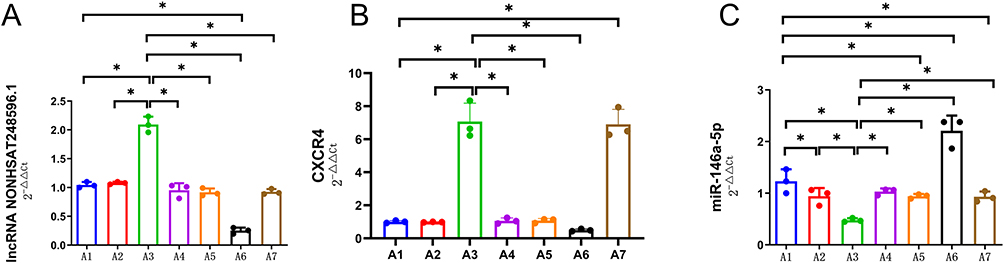 |
Figure 3 Expression of lncRNA NONHSAT248596.1 (A), CXCR4 (B), and miR-146a-5p (C) in chondrocytes across experimental groups. (A) NONHSAT248596.1 levels were significantly increased by its overexpression plasmid and decreased by its interference plasmid following SDF-1 induction. (B) CXCR4 expression was upregulated by NONHSAT248596.1 overexpression and downregulated by NONHSAT248596.1 knockdown. (C) miR-146a-5p expression showed an inverse correlation with NONHSAT248596.1 and CXCR4 levels in all groups. Note: A1: SDF-1-induced OA chondrocytes for 48h. A2: OA chondrocytes were exposed to a control plasmid overexpressing NONHSAT248596.1. A3: OA chondrocytes were exposed to a plasmid overexpressing NONHSAT248596.1. A4: OA chondrocytes were exposed to a plasmid overexpressing NONHSAT248596.1 and miR-146a-5p mimic. A5: OA chondrocytes were exposed to a control NONHSAT248596.1 interference plasmid. A6: OA chondrocytes were exposed to a NONHSAT248596.1 interference plasmid. A7: OA chondrocytes were exposed to a NONHSAT248596.1 interference plasmid and a miR-146a-5p inhibitor. (n=3, *p < 0.05). |
The NONHSAT248596.1/miR-146a-5p/CXCR4 ceRNA Network Promotes Apoptosis and ECM Degradation in vitro
Chondrocytes were transfected separately with plasmids overexpressing NONHSAT248596.1, a NONHSAT248596.1 interference plasmid, a miR-146a-5p mimic, or a miR-146a-5p inhibitor, followed by exposure to SDF-1. Cell viability was assessed by CCK-8 assay, and apoptosis was quantified by flow cytometry. Treatment with SDF-1 alone significantly reduced cell viability (Figure 4A) and increased apoptosis (Figure 4B and C). Overexpression of NONHSAT248596.1 enhanced these SDF-1-induced effects, whereas silencing of NONHSAT248596.1 significantly attenuated them (Figure 4A–C). In rescue experiments, both overexpression and inhibition of miR-146a-5p counteracted the effects on cell viability and apoptosis induced by altering NONHSAT248596.1 expression (Figure 4A–C). These results indicate that NONHSAT248596.1 promotes SDF-1-induced chondrocyte apoptosis, and that this effect can be rescued by modulating miR-146a-5p expression.
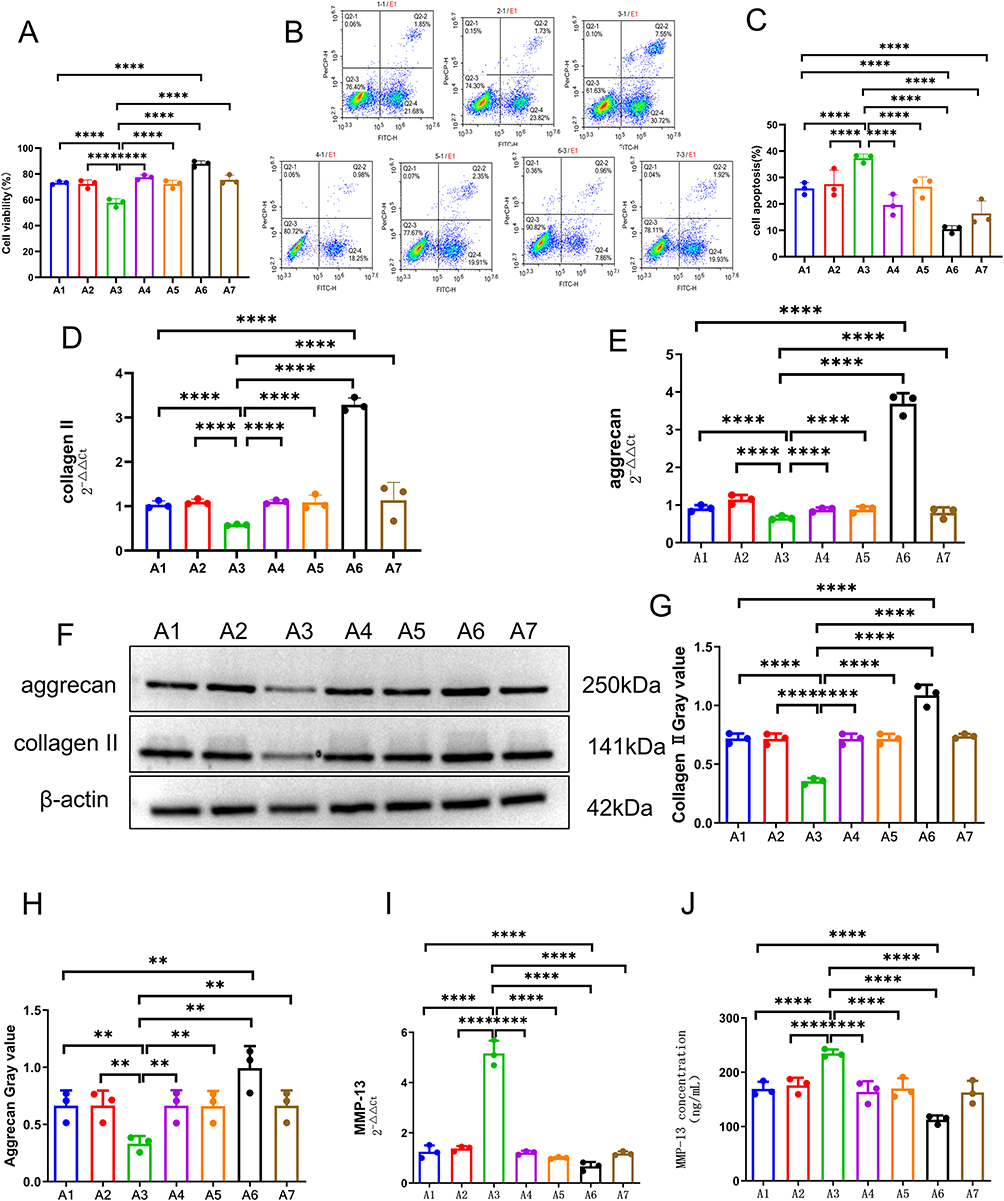 |
Figure 4 lncRNA NONHSAT248596.1 regulates human OA chondrocyte degeneration by sponging miR-146a-5p to modulate the SDF-1/CXCR4 axis. (A) Chondrocyte proliferation was assessed by CCK-8 assay. (B and C) Chondrocyte apoptosis was quantified by flow cytometry. (D–H) Expression of collagen II and aggrecan was analyzed by qRT-PCR and Western blot. (I and J) Expression and secretion of MMP-13 were quantified by qRT-PCR and ELISA, respectively. Note: A1: SDF-1-induced OA chondrocytes for 48h. A2: OA chondrocytes were exposed to a control plasmid overexpressing NONHSAT248596.1. A3: OA chondrocytes were exposed to a plasmid overexpressing NONHSAT248596.1. A4: OA chondrocytes were exposed to a plasmid overexpressing NONHSAT248596.1 and miR-146a-5p mimic. A5: OA chondrocytes were exposed to a control NONHSAT248596.1 interference plasmid. A6: OA chondrocytes were exposed to a NONHSAT248596.1 interference plasmid. A7: OA chondrocytes were exposed to a NONHSAT248596.1 interference plasmid and a miR-146a-5p inhibitor. (n=3, **p<0.01, ****p<0.0001). |
Furthermore, qRT-PCR and Western blot analysis revealed that SDF-1 stimulation downregulated collagen II and aggrecan expression in chondrocytes (Figure 4D–H, Supplementary File S1). Concurrently, qRT-PCR and ELISA results showed that SDF-1 upregulated MMP-13 expression (Figure 4I and J). These effects were enhanced by NONHSAT248596.1 overexpression but reversed by NONHSAT248596.1 interference (Figure 4D–I), indicating that NONHSAT248596.1 promotes SDF-1-induced ECM degradation in chondrocytes. Both overexpression and inhibition of miR-146a-5p attenuated the ECM degradation resulting from altered NONHSAT248596.1 expression (Figure 4). These findings demonstrate that the NONHSAT248596.1/miR-146a-5p/CXCR4 ceRNA network regulates the progression of SDF-1-induced OA.
NONHSAT248596.1 Promotes OA Progression via the miR-146a-5p/CXCR4 Axis in a Rabbit Model
The function of the NONHSAT248596.1/miR-146a-5p/CXCR4 axis in OA was further investigated in a rabbit model. Gross morphology revealed distinct pathological features across experimental groups: the lncRNA group exhibited a dry, granular cartilage surface with reduced glossiness, large-area defects, osteophyte formation, and joint deformity; the miRNA group showed a smooth cartilage surface without defects, slightly reduced glossiness, and subchondral hyperemia of the femoral condyle; the ceRNA group displayed a slightly dry surface with reduced glossiness, partial cartilage defects, and obvious inflammatory reaction; the control group demonstrated a slightly dry and rough cartilage surface with reduced gloss, but without large-area cartilage loss (Figure 5A). NONHSAT248596.1 overexpression exacerbated SDF-1-induced morphological damage in rabbit cartilage (OARSI score: 4.67±2.1), while miR-146a-5p overexpression attenuated this effect (OARSI score: 1.56±0.48, p<0.0001, n=3/group/time point) at 12 week (Figure 5B).
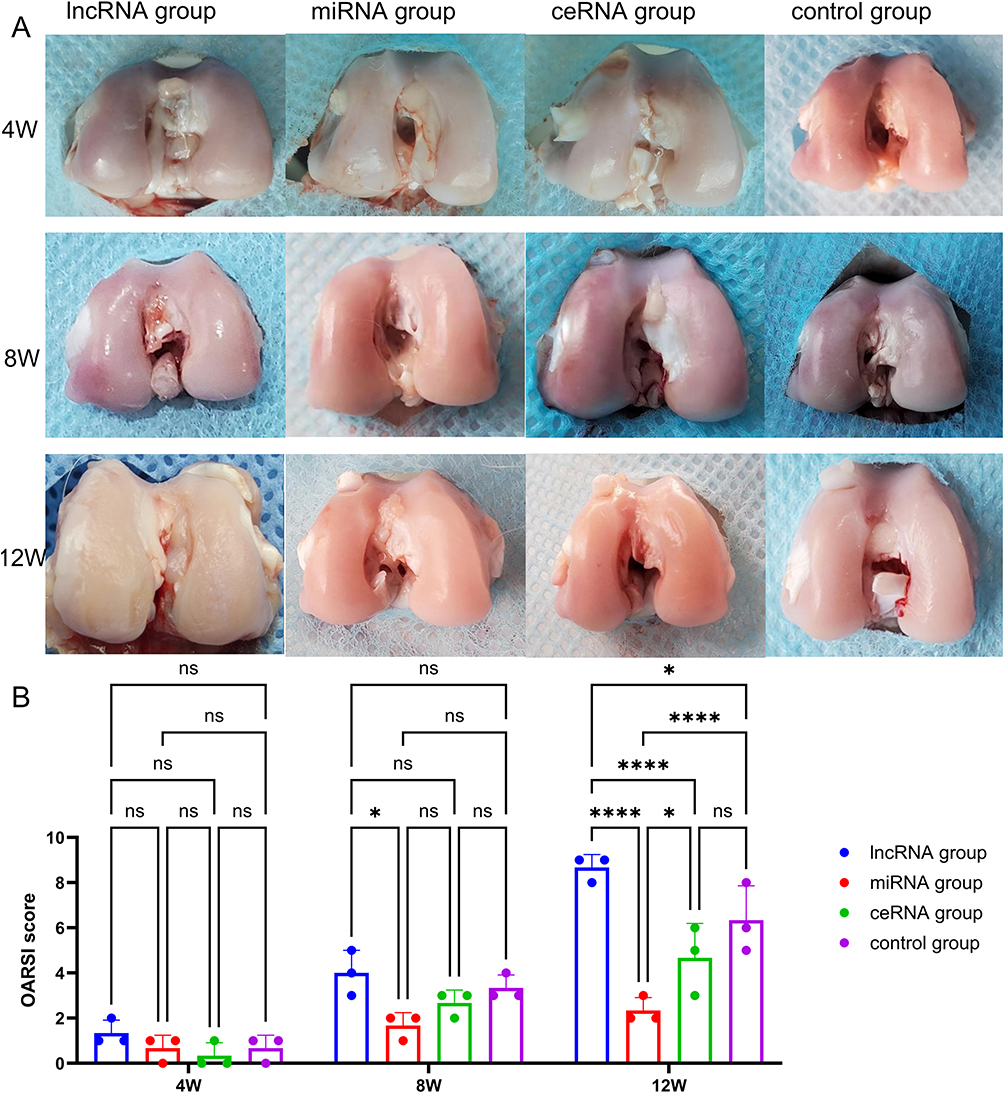 |
Figure 5 Gross morphology (A) and OARSI macroscopic score statistical results (B) of the OA rabbit model assessed at 4, 8, and 12 weeks. LncRNA group: 10 μL lentiviral vector overexpressing lncRNA NONHSAT248596.1 and 10 μL empty lentiviral vector without miR-146a-5p expression were injected into the right posterior knee; miRNA group: 10 μL lentiviral vector overexpressing miR-146a-5p and 10 μL empty lentiviral vector without lncRNA NONHSAT248596.1 expression were injected into the right posterior knee; ceRNA group: 10 μL lentiviral vector overexpressing both miR-146a-5p and lncRNA NONHSAT248596.1 was injected into the right posterior knee; control group: 10 μL empty lentivirus vector without lncRNA NONHSAT248596.1 expression and 10 μL empty lentivirus vector without miR-146a-5p expression were injected into the right posterior knee. (n=3, *p<0.05, ****p < 0.0001). Abbreviation: ns, no significance. |
Histological staining demonstrated that NONHSAT248596.1 overexpression exacerbated SDF-1-induced cartilage tissue destruction and proteoglycan loss, effects that were attenuated by miR-146a-5p overexpression (Figure 6A and B). Detailed histological analysis revealed distinct pathological characteristics: In the lncRNA group, cartilage was substantially thinner with markedly reduced chondrocytes; cell polarity was lost with disordered arrangement; significant cell swelling and large fissures near the subchondral bone were observed. In the miRNA group, the cartilage surface remained uniform with only mild cell reduction; chondrocytes maintained normal polarity and arrangement with no apparent swelling. In the ceRNA group, cartilage thinning was evident with disordered cell arrangement, loss of polarity, cell swelling, disappearance of cartilage lacunae, and reduced chondrocytes in the middle layers. In the control group, cartilage showed thinning, loss of cell polarity, disordered arrangement, obvious cell swelling, significant chondrocyte reduction, disappearance of cartilage lacunae, and fissure formation near the bone (Figure 6A and B). These results indicate that while the control group progressed to advanced OA after 12 weeks of NONHSAT248596.1 and SDF-1 stimulation, the miR-146a-5p overexpression group remained in the early to middle stages of OA progression (Mankin score: 5.9±1.7 VS 2.4±0.59, p<0.0001, n=3/group/time point) (Figure 6C).
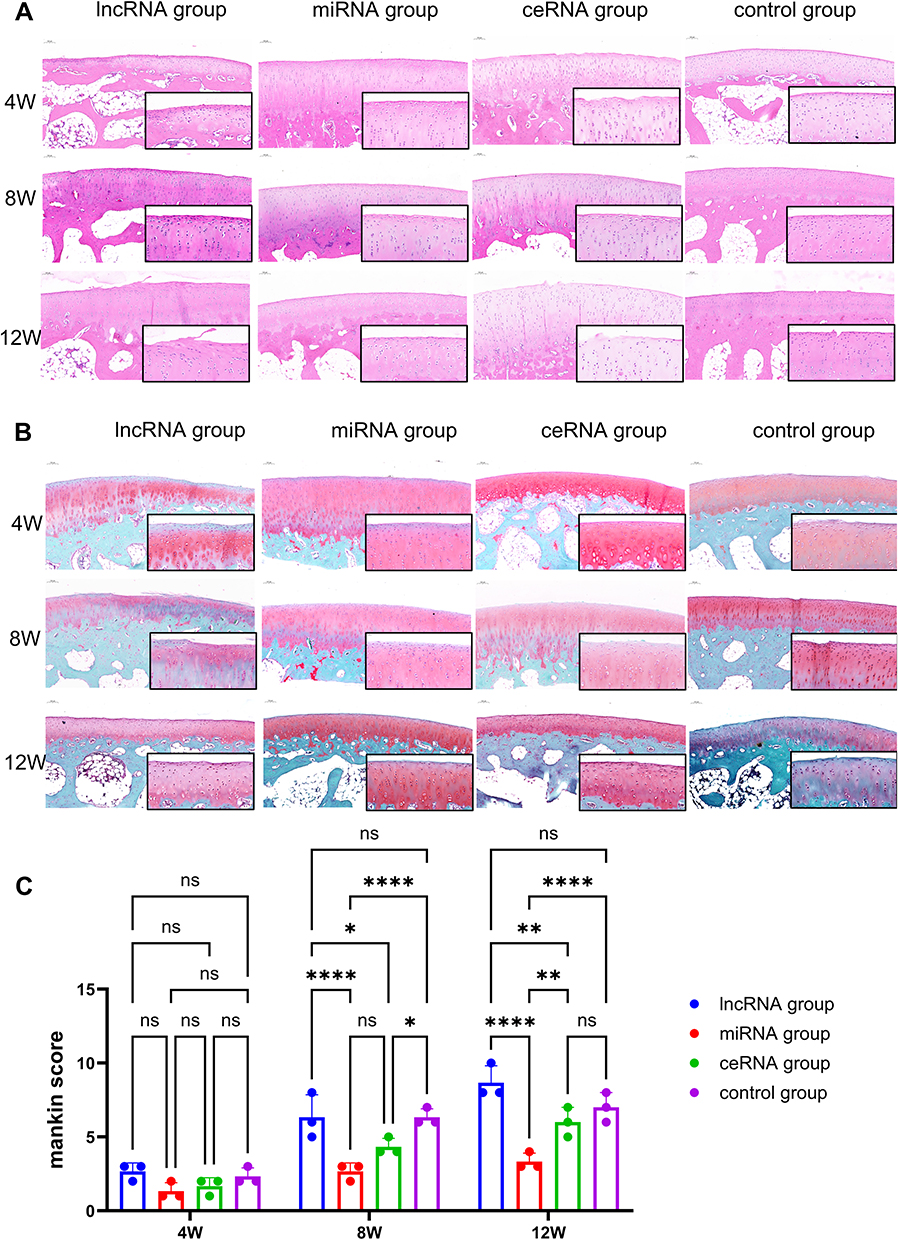 |
Figure 6 HE staining (A), Safranin O-fast green staining (B) and Mankin scores (C) of cartilage tissues from each group at 4, 8, and 12 weeks after OA modeling. Scale bars = 100/25 μm. Histological evaluation included the cartilage surface, chondrocyte number, cell arrangement, and ECM structure. The control group progressed to advanced-stage OA following 12 weeks of lncRNA NONHSAT248596.1 stimulation, whereas the miRNA group remained at an intermediate stage of OA progression. (n=3/group/time point, *p<0.05, **p<0.01, ****p < 0.0001). Abbreviation: ns, no significance. |
Western blot analysis further demonstrated that NONHSAT248596.1 reduced aggrecan and collagen II protein expression in cartilage tissue (Figure 7A–C, Supplementary File S2). qRT-PCR analysis revealed increased MMP-13 mRNA expression in cartilage, while ELISA detected elevated SDF-1, IL-1β, and TNF-α protein levels in synovial tissue (Figure 7D–G). At the 12-week time point, qRT-PCR results showed that NONHSAT248596.1 significantly upregulated CXCR4 and MMP-13 mRNA expression while downregulating miR-146a-5p, aggrecan, and collagen II (Figure 7H). Critically, intra-articular injection of miR-146a-5p abolished all the effects induced by NONHSAT248596.1 (Figure 7). Collectively, these results confirm that NONHSAT248596.1 promotes SDF-1-induced OA progression in vivo through the miR-146a-5p/CXCR4 axis.
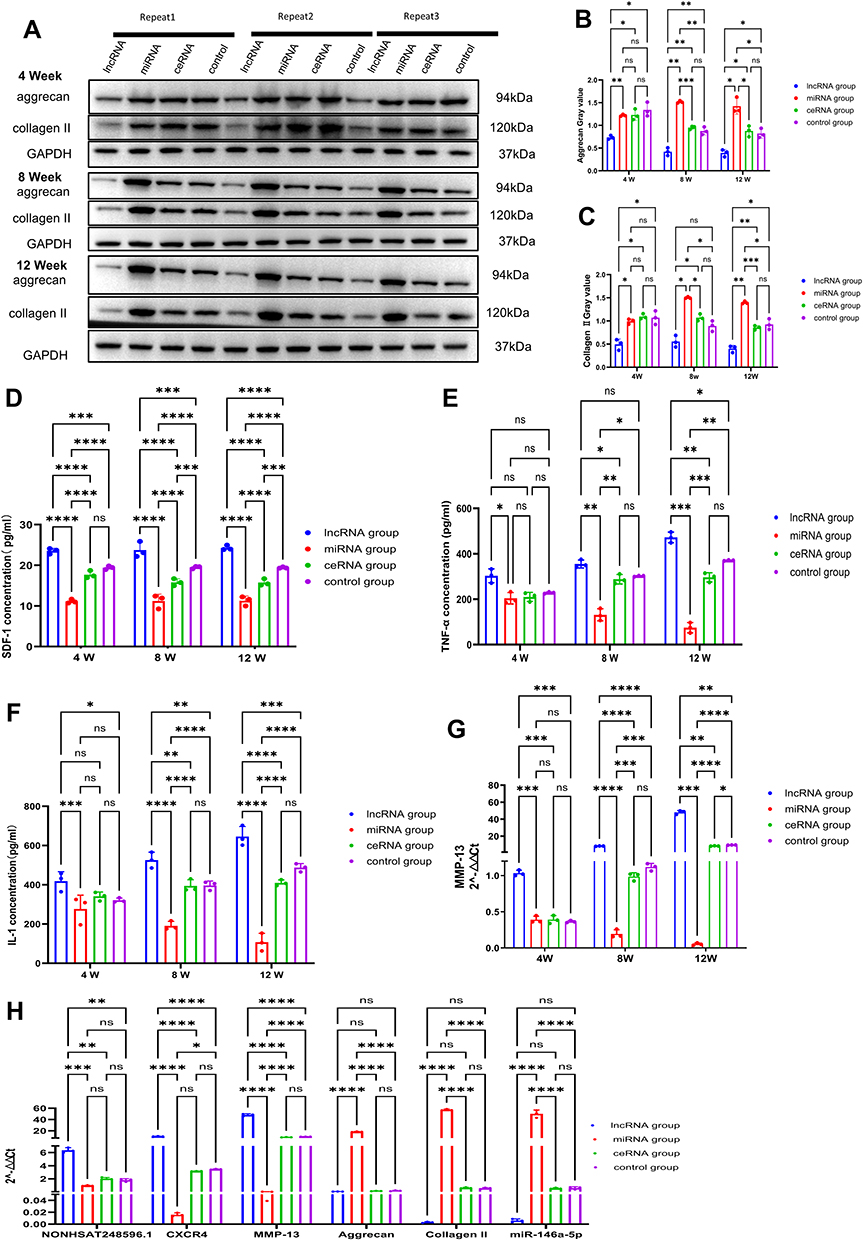 |
Figure 7 Functional role of the NONHSAT248596.1/miR-146a-5p/CXCR4 axis in rabbit OA. (A–C) Protein expression of aggrecan and collagen II in cartilage tissue analyzed by Western blot. (D–G) Expression levels of SDF-1 (D), TNF-α (E), IL-1β (F), and MMP-13 (G) measured by ELISA or qRT-PCR. (H) qRT-PCR analysis at 12 weeks showing NONHSAT248596.1-induced upregulation of CXCR4 and MMP-13, and downregulation of miR-146a-5p, aggrecan, and collagen II in cartilage tissue. (n=3, *p<0.05, **p<0.01, ***p<0.001, ****p < 0.0001). |
Discussion
Accumulating evidence indicates that dysregulated lncRNAs are intricately associated with OA pathogenesis,19–21 yet the functional roles of most lncRNAs identified via human transcriptome sequencing in OA remain unclear.22 Our previous sequencing analysis of SDF-1-induced OA cell models identified CXCR4 as a key target of miR-146a-5p15,23 and revealed several differentially expressed lncRNAs, including NONHSAT248596.1. The present study demonstrates that NONHSAT248596.1 functions as a ceRNA for miR-146a-5p to mediate SDF-1-induced ECM degradation and chondrocyte apoptosis in vitro.
miR-146a-5p is an immunomodulatory miRNA with recognized tumor suppressor functions in various cancers24,25 and has been shown to attenuate inflammatory responses in OA.26 Our previous work established that miR-146a-5p inhibits chondrocyte apoptosis and ECM degeneration in OA through regulation of the CXCR4 axis,16 a finding consistent with other reports of miR-146a-5p-mediated CXCR4 regulation.27 Nevertheless, the precise mechanistic role of the miR-146a-5p-CXCR4 axis in OA requires further elucidation.
NONHSAT248596.1 is a cytoplasmic long non-coding RNA that functions as a ceRNA.11 Our findings indicate that NONHSAT248596.1 promotes SDF-1-induced OA in chondrocytes by regulating the miR-146a-5p/CXCR4 axis. Analysis of OA cartilage tissue revealed a positive correlation between NONHSAT248596.1 and CXCR4 expression, and a negative correlation between miR-146a-5p and CXCR4. Importantly, overexpression of miR-146a-5p reversed the growth inhibition, cell cycle arrest, and apoptosis induced by NONHSAT248596.1 in OA chondrocytes.
Recent clinical studies demonstrate that carboxymethyl-chitosan (CM-C) injections improve pain and functional outcomes in advanced knee OA.28 Given that CM-C modulates cartilage homeostasis, it may interact with the NONHSAT248596.1/miR-146a-5p/CXCR4 axis. Future studies could investigate whether combining CM-C therapy with NONHSAT248596.1 silencing produces enhanced therapeutic effects for OA. NONHSAT248596.1 is upregulated in OA patients’ cartilage tissue, suggesting it may correlate with histological severity, future studies will also explore its potential as a diagnostic biomarker.
This study has several limitations. First, although our preliminary data confirmed that CXCR4 protein levels were significantly upregulated in OA chondrocytes following miR-146a-5p inhibitor transfection15 and dual luciferase assays preliminarily established physical binding between miR-146a-5p and CXCR4, we did not pursue further validation through methods such as RNA immunoprecipitation. Second, chondrocyte autophagy plays complex roles in OA pathogenesis, and its activity at different disease stages requires systematic investigation across multiple time points. Third, a formal power analysis was not performed prior to the study. Although our sample sizes (10 human cartilage specimens, n=9 rabbits per group) are consistent with typical preclinical OA research, they may limit the detection of small effect sizes. Fourth, the potential involvement of the NONHSAT248596.1/miR-146a-5p/CXCR4 axis in regulating chondrocyte autophagy merits further investigation to fully elucidate its regulatory mechanisms.
Conclusions
This study demonstrates that lncRNA NONHSAT248596.1 promotes chondrocyte apoptosis and ECM degradation in OA by functioning as a ceRNA for miR-146a-5p, thereby regulating CXCR4 expression. These findings identify NONHSAT248596.1 as a potential therapeutic target for OA intervention.
Abbreviations
OA, osteoarthritis; lncRNAs, long non-coding RNAs; CeRNA, competing endogenous RNA; SDF-1, stromal cell-derived factor-1; CXCL12, CXC motif chemokine ligand 12; CXCR4, C-X-C chemokine receptor type 4; ELISA: Enzyme-linked immunosorbent assay; MMP-13: Matrix metallopeptidase 13; ECM, extracellular matrix.
Data Sharing Statement
Sequencing data have been uploaded to Sequence Read Archive (SRA) database (https://www.ncbi.nlm.nih.gov/sra/SRX8499525), and the BioProject ID is PRJNA638147.The other datasets are available from the corresponding author Yanlin Li upon reasonable request.
Ethics Approval and Consent to Participate
The investigation was approved by the Ethics Committee of Kunming Medical University First Affiliated Hospital (approval number: (2018) Ethical Review L No. 21; approval date: 2018-07-25), and all patients consented in writing. The investigation was conducted in accordance with the principles of the Declaration of Helsinki.
All animal experiments were approved by the Experimental Animal Ethics Committee of Kunming Medical University (Approval No.: kmmu2020416; Date: April 1, 2020) and conducted in compliance with the national standard “Laboratory Animal - Guideline for Ethical Review of Animal Welfare” (GB/T 35892-2018, China).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Yunnan Fundamental Research Projects (grant No. 202501AT070114), Yunnan Fundamental Research Kunming Medical University Projects (grant No. 202401AY070001-007, 202401AY070001-242) and Yunnan Province Major Science and Technology Special Project (grant No. 202403AC100025). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Disclosure
The authors declare no conflict of interest in this work.
References
1. Ratneswaran A, Kapoor M. Osteoarthritis year in review: genetics, genomics, epigenetics. Osteoarthr Cartilage. 2021;29(2):151–160. doi:10.1016/j.joca.2021.11.004
2. Chen L, Yu Y. Exercise and osteoarthritis. Adv Exp Med Biol. 2020;1228(16):219–231. doi:10.1007/978-981-15-1792-1_15
3. Li Z, Huang Z, Bai L. Cell Interplay in Osteoarthritis. Front Cell Develop Biol. 2021;9:720477. doi:10.3389/fcell.2021.720477
4. Wilkinson JM, Zeggini E. The genetic epidemiology of joint shape and the development of osteoarthritis. Calcified Tissue Int. 2021;109(3):257–276. doi:10.1007/s00223-020-00702-6
5. Yuan Y, Tang Y, Fang Z, Wen J, Wicha MS, Luo M. Long non-coding RNAs: key regulators of tumor epithelial/mesenchymal plasticity and cancer stemness. Cells. 2025;14(3). doi:10.3390/cells14030227
6. Zhang Y, Liu H, Niu M, et al. Roles of long noncoding RNAs in human inflammatory diseases. Cell Death Discovery. 2024;10(1):235. doi:10.1038/s41420-024-02002-6
7. Pan Y, Jiao Q, Wei W, Zheng T, Yang X, Xin W. Emerging role of LncRNAs in ischemic stroke-novel insights into the regulation of inflammation. J Inflamm Res. 2021;14:4467–4483. doi:10.2147/jir.S327291
8. Ghafouri-Fard S, Poulet C, Malaise M, et al. The emerging role of non-coding RNAs in osteoarthritis. Front Immunol. 2021;12:773171. doi:10.3389/fimmu.2021.773171
9. Shi D, Mei Y, Hao W, Li J, Liu S, Lin X. Biological functions and applications of LncRNAs in the regulation of the extracellular matrix in osteoarthritis. Front Cell Dev Biol. 2023;11:1330624. doi:10.3389/fcell.2023.1330624
10. Wang R, Shiu HT, Lee WYW. Emerging role of lncRNAs in osteoarthritis: an updated review. Front Immunol. 2022;13(10):982773. doi:10.3389/fimmu.2022.982773
11. Wang G, He L, Xiang Y, Jia D, Li Y. Long noncoding and micro-RNA expression in a model of articular chondrocyte degeneration induced by stromal cell-derived factor-1. Asian Biomed (Res Rev News). 2022;16(4):169–179. doi:10.2478/abm-2022-0021
12. Biyani S, Patil A, Swami V. The influence of SDF-1 (CXCL12) gene in health and disease: a review of literature. Biophys Rev. 2025;17(1):127–138. doi:10.1007/s12551-024-01230-5
13. Wang S, Mobasheri A, Zhang Y, Wang Y, Dai T, Zhang Z. Exogenous stromal cell-derived factor-1 (SDF-1) suppresses the NLRP3 inflammasome and inhibits pyroptosis in synoviocytes from osteoarthritic joints via activation of the AMPK signaling pathway. Inflammopharmacology. 2021;29(3):695–704. doi:10.1007/s10787-021-00814-x
14. Wang Z, Zhu P, Li H, et al. Sodium hyaluronate-PDGF repairs cartilage and subchondral bone microenvironment via HIF-1α-VEGF-notch and SDF-1-CXCR4 inhibition in osteoarthritis. J Cell Mol Med. 2025;29(7):e70515. doi:10.1111/jcmm.70515
15. Jia D, Li Y, Han R, et al. miR‑146a‑5p expression is upregulated by the CXCR4 antagonist TN14003 and attenuates SDF‑1‑induced cartilage degradation. Mol med rep. 2019;19(5):4388–4400. doi:10.3892/mmr.2019.10076
16. Yang T, Li C, Li Y, et al. MicroRNA-146a-5p alleviates the pathogenesis of osteoarthritis by inhibiting SDF-1/CXCR4-induced chondrocyte autophagy. Int Immunopharmacol. 2023;117(2):109938. doi:10.1016/j.intimp.2023.109938
17. Konermann S, Brigham MD, Trevino AE, et al. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature. 2015:517(7536):583–588. doi:10.1038/nature14136
18. Li C, He Y, Li Y, et al. A novel method to establish the rabbit model of knee osteoarthritis: intra-articular injection of SDF-1 induces OA. BMC Musculoskelet Disord. 2021;22(1):329–339. doi:10.1186/s12891-021-04188-7
19. Jiang M, Liu J, Luo T, Chen Q, Lu M, Meng D. LncRNA PACER is down-regulated in osteoarthritis and regulates chondrocyte apoptosis and lncRNA HOTAIR expression. Biosci Rep. 2019;39(6):BSR20190404. doi:10.1042/BSR20190404
20. Kong H, Sun ML, Zhang XA, Wang XQ. Crosstalk among circRNA/lncRNA, miRNA, and mRNA in osteoarthritis. Front Cell Develop Biol. 2021;9(8):774370. doi:10.3389/fcell.2021.774370
21. Chen Y, Zhang L, Li E, et al. Long-chain non-coding RNA HOTAIR promotes the progression of osteoarthritis via sponging miR-20b/PTEN axis. Life Sci. 2020;253:117685. doi:10.1016/j.lfs.2020.117685
22. Wu X, Bian B, Lin Z, et al. Identification of exosomal mRNA, lncRNA and circRNA signatures in an osteoarthritis synovial fluid-exosomal study. Exp Cell Res. 2022;410(1):112881. doi:10.1016/j.yexcr.2021.112881
23. Yang T, Yang G, Wang G, et al. Bioinformatics identification and integrative analysis of ferroptosis-related key lncRNAs in patients with osteoarthritis. Biosci Rep. 2023;43(9):1–17. doi:10.1042/BSR20230255
24. Liu X, Liu B, Li R, et al. miR-146a-5p plays an oncogenic role in NSCLC via suppression of TRAF6. Front Cell Develop Biol. 2020;8(36):847–864. doi:10.3389/fcell.2020.00847
25. Cabello P, Torres-Ruiz S, Adam-Artigues A, et al. miR-146a-5p promotes angiogenesis and confers trastuzumab resistance in HER2+ breast cancer. Cancers. 2023;15(7):2138. doi:10.3390/cancers15072138
26. Li MW, Gao L, Dang YW, et al. Protective potential of miR-146a-5p and its underlying molecular mechanism in diverse cancers: a comprehensive meta-analysis and bioinformatics analysis. Cancer Cell Int. 2019;19(08):167–188. doi:10.1186/s12935-019-0886-y
27. Wang D, Wang X, Song Y, et al. Exosomal miR-146a-5p and miR-155-5p promote CXCL12/CXCR7-induced metastasis of colorectal cancer by crosstalk with cancer-associated fibroblasts. Cell Death Dis. 2022;13(4):380–395. doi:10.1038/s41419-022-04825-6
28. Manocchio N, Pirri C, Ljoka C, et al. Long-term efficacy of carboxymethyl-chitosan in advanced knee osteoarthritis: a twelve-month follow-up study on non-responders to hyaluronic acid. Biomedicines. 2025;13(2). doi:10.3390/biomedicines13020270
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.