Back to Journals » OncoTargets and Therapy » Volume 13
lncRNA MAFG-AS1 Contributes to Esophageal Squamous-Cell Carcinoma Progression via Regulating miR143/LASP1
Received 14 April 2020
Accepted for publication 23 July 2020
Published 20 August 2020 Volume 2020:13 Pages 8359—8370
DOI https://doi.org/10.2147/OTT.S258157
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
This paper has been retracted.
Yunhui Qu, 1 Jinbo Liu 2
1Department of Clinical Laboratory, First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, People’s Republic of China; 2Department of Colorectal Surgery, First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, People’s Republic of China
Correspondence: Jinbo Liu Email [email protected]
Background: Increasing investigations indicate that long noncoding RNA (lncRNA) is responsible for diverse biological functions during the progression of cancer. However, its functions and underlying mechanisms remain elusive. Here, we investigated the MAFG-AS1- expression profile in esophageal squamous-cell carcinoma (ESCC) patients and explored its biological function and potential molecular mechanisms.
Methods: qRT-PCR and the GEPIA data base were used to evaluate expression levels of MAFG-AS1 in ESCC tissue and cells. WST1-proliferation, -migration, and -invasion assays were performed to define the role of MAFG-AS1 in ESCC. Potential molecular mechanisms of MAFG-AS1 were investigated with online bioinformatic analysis, qRT-PCR, and rescue assays.
Results: MAFG-AS1 was upregulated in 45 ESCC-tissue samples and cell lines compared to that of adjacent nontumor tissue and normal esophageal cells. Higher MAFG-AS1 expressionindicated poor survival. Gain- and loss-of-function experiments suggested that MAFG-AS1 promoted ESCC-cell proliferation, migration, and invasion. Molecular mechanism analysis and rescue assay showed that miR143 inhibitors partly abolished the suppression of MAFG-AS1 knockdown on EC109-cells proliferation. Moreover, we found that LASP1 specifically targeted miR143. Collectively, these data indicated that MAFG-AS1 served as a ceRNA to elevate LASP1 levels by sponging miR143, and played an oncogenic role in ESCC.
Conclusion: Our research findings demonstrate that MAFG-AS1 is a key regulator through a novel MAFG-AS1–miR143–LASP1 axis in ESCC development and progression, which may offer a potential therapeutic target for ESCC.
Keywords: lncRNA, ESCC, MAFG-AS1, miR143, LASP1
Introduction
Esophageal cancer (EC) is one of the leading contributors to high cancer-related mortality.1,2 Esophageal squamous-cell carcinoma (ESCC) and esophageal adenocarcinoma are two major subtypes of EC.3 Despite clinical successes being achieved in ESCC treatment over the past few decades, the low 5-year overall-survival rate and poor prognosis remain major clinical obstacles to achieving successful recovery.4,5 Recently, trends in high-throughput sequencing techniques have led to a proliferation of studies focused on lncRNA,6 which are transcripts >200 nucleotides and have limited protein-coding potential.7 Studies have recognized that lncRNA, as a multitalented transcriptional and posttranscriptional regulator of key cancer pathways, possesses lots of biological functions involved in many types of cancer progression,8 such as hepatocellular carcinoma,9 lung cancer (LC),10 colorectal cancer (CRC),11 and gastric cancer.12 A number of cross-sectional studies have suggested numerous lncRNAs being related to the development of ESCC. It has been reported that NORAD was correlated with tumor size and overall survival in ESCC. SPINT1-AS1 was found to be an independent unfavorable prognostic indicator in ESCC.13 GAS5 has been reported to be an oncogene, evidenced by enhancement of proliferative, migrative, and invasive abilities in ESCC.14
MAFG-AS1, also known as MAFG-DT, is located at 17q25.3, whose transcript contains approximately 1,895 bp. Studies have shown that MAFG-AS1 is upregulated in non–small cell LC (NSCLC)15 and that MAFG-AS1 promotes the aggression of breast cancer via regulating the miR339-5p–MMP15 axis.16 However, few studies have provided quantitative evidence of the effects of MAFG-AS1 on ESCC progression. In this study, we first found that MAFG-AS1 was upregulated in ESCC tissue and cells, and high expression of MAFG1-AS1 was associated with ESCC lymph-node metastasis. Furthermore, functional assays indicated that MAFG-AS1 promoted ESCC cell proliferation, migration, and invasion. The underlying mechanisms of MAFG-AS1 were also explored. We revealed that MAFG-AS1 accelerates ESCC development via regulating themiR143–LASP1 axis. In all, this paper contributes to the understanding of ESCC development.
Methods
Patient Samples
A total of 45 pairs of ESCC fresh-tissue and adjacent nontumor-tissue samples were obtained from the First Affiliated Hospital of Zhengzhou University from April 2016 to May 2017. After resection, the samples were collected immediately and then frozen at −80°C till RNA extraction. Every specimen was evaluated as ESCC by at least three experienced pathologists. The protocol got the approval of the First Affiliated Hospital of Zhengzhou University, and informed consent was obtained from all patients.
Cell Lines and Cell Culture
The human normal esophageal cell line HET1A and ESCC cell lines EC1 and EC109 were purchased from the Shanghai Institutes of Life Science Cell Bank Center (Shanghai, China) andRPMI 1640; HyClone, UT, USA) supplemented with 10% FBS, and 1% penicillin–streptomycin. Cell lines were incubated in a humidified atmosphere at 37°C with 5% CO2.
RNA Extraction and qRT-PCR Analysis
Total RNA was extracted from tissue and cells with Trizol reagent (Takara, Dalian, China) according to the manufacturer’s protocol. Then, cDNA was synthesized with a PrimeScript RT reagent kit with gDNA Eraser (Takara). Then, qRT-PCR of MAFG-AS1, miR143, and LASP1 were examined with the kit specifications of SYBR Premix Ex Taq II (Takara) on a LightCycler 480 II real-time PCR system (Roche, Basel, Switzerland). Relative expression levels of mRNA or miRNA were normalized to the GAPDH gene or U6 gene.
Cell Transfection
Plasmid vectors (GV219-MAFG-AS1 and GV219-NC) were obtained from GeneChem (Shanghai, China) and transfected into EC1 and EC109 cells using Lipo8000 (Beyotime Biotechnology, Shanghai, China) according to the manufacturer’s protocol. Small interfering RNA targeting MAFG-AS1 (siMAFG-AS1) and scrambled siRNA of MAFG-AS1 (siRNA control) were designed by RiboBio (Guangzhou, China) and transfected using RFect siRNA-transfection reagent (Changzhou Biogenerating Biotechnologies). Then, stably MAFG-AS1–silenced transfected GV219-MAFG-AS1 cells were screened out for further study after transfection for 48 hours. miR143 inhibitors and negative control were synthesized by RiboBio.
Cell-Proliferation Assays
Cell proliferation were measured with a WST1-assay kit (Beyotime Biotechnology) as described before.3 In brief, cells transfected with siRNAs or plasmid vectors and negative controls were inoculated in 96-well plates at a density of 2,000 cells per well. Then, cell-proliferation percentages were measured at 24, 48, 72, and 96 hours using a SpectraMax M5 (Molecular Devices, San Francisco, USA).
Cell-Migration and -Invasion Assays
To investigate the migration and invasion potential of ESCC cells, transwell assays were conducted using the transwell 24-well chambers with or without Matrigel as described before.17 In the invasion assay, transfected ESCC-cell suspension (105) was placed in the upper chamber, covered with Matrigel (BD Biosciences) in 200 µL serum-free RPMI 1640 accompanied by 600 µL medium containing 15% FBS in the bottom chamber. For the migration experiment, the top chamber was covered with an uncoated membrane. After incubation for 24 hours at 37°C, cells remaining in the top chamber were cleared away. Then, cells were fixed with 4% paraformaldehyde and stained with crystal violet. Finally, cell numbers were counted and imaged by microscopy.
Statistical Analysis
All statistical analysis was performed using IBM SPSS 23 and GraphPad Prism 8 (GraphPad Software, La Jolla, CA, USA). Student’s t-test, Fisher’s exact test, χ2-test, Pearson’s correlation analysis, and one-way ANOVA were used to detect differences between groups. Kaplan–Meier survival curves were analyzed with the log-rank test. P<0.05 was considered statistically significant.
Results
MAFG-AS1 Upregulated in ESCC Tissue and Cell Lines
To explore the biological function of MAFG-AS1 in ESCC, qRT-PCR was performed to investigate the MAFG-AS1 expression in 45 paired ESCC-tissue and non–ESCC tissue samples firstly. MAFG-AS1 was significantly elevated in ESCC tissue compared with paired adjacent nontumor tissue (Figure 1A). Furthermore, the association between the expression of MAFG-AS1 and clinicopathological features of ESCC patients was analyzed. Results showed that those patients with higher MAFG-AS1-expression levels had lymphatic metastasis, while there was no statistical relation between MAFG-AS1 expression and other clinical characteristics (Table 1, **P<0.01). Moreover, as shown, MAFG-AS1 expression was higher in the lymphatic metastasis group than in the nonmetastasis group (Figure 1B). Bioinformatic data from GEPIA http://gepia2.cancer-pku.cn) supported these findings (Figure 1C and D).
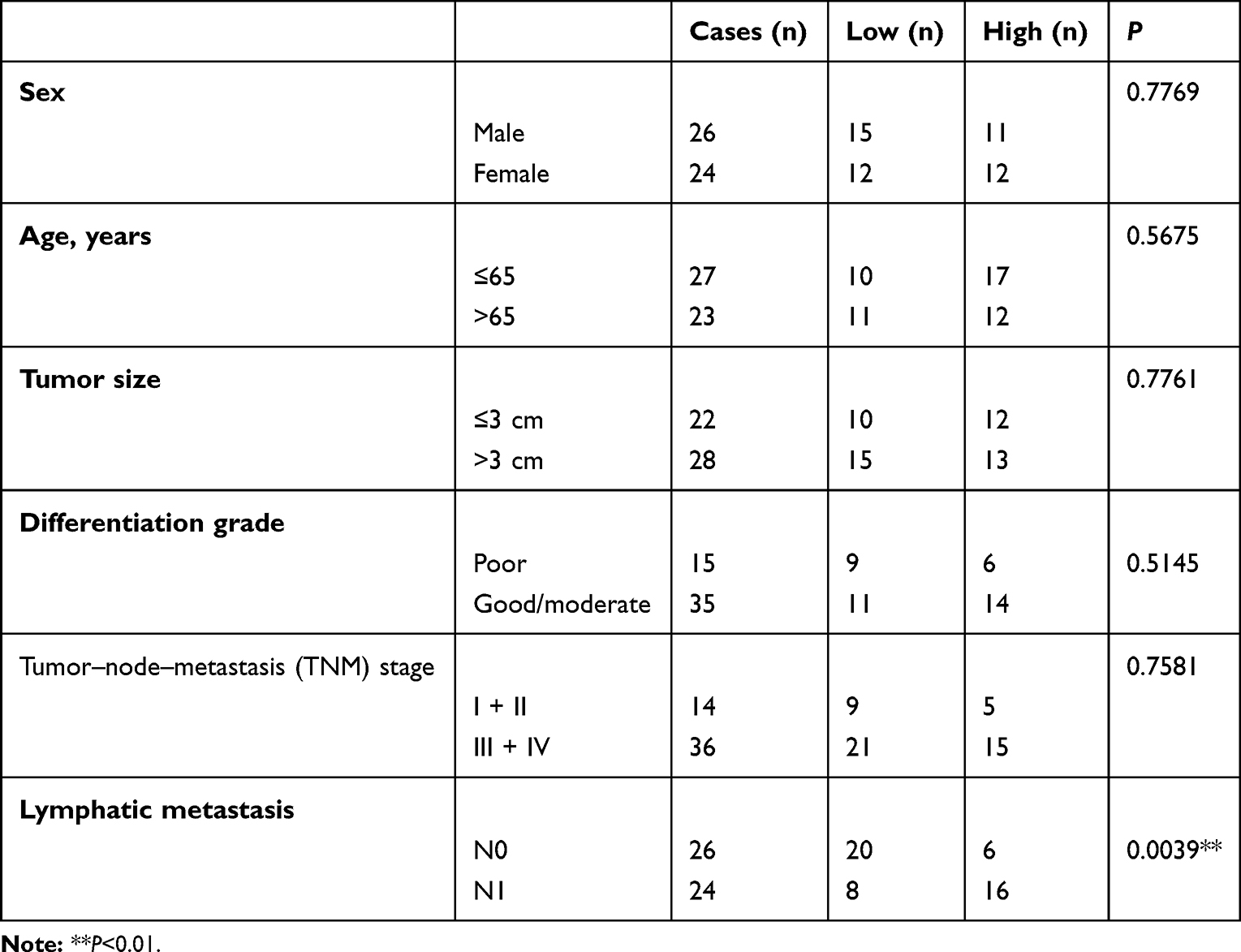 |
Table 1 Correlation Between MAFG-AS1 and Clinicopathological Parameters of Esophageal Squamous-Cell Carcinoma |
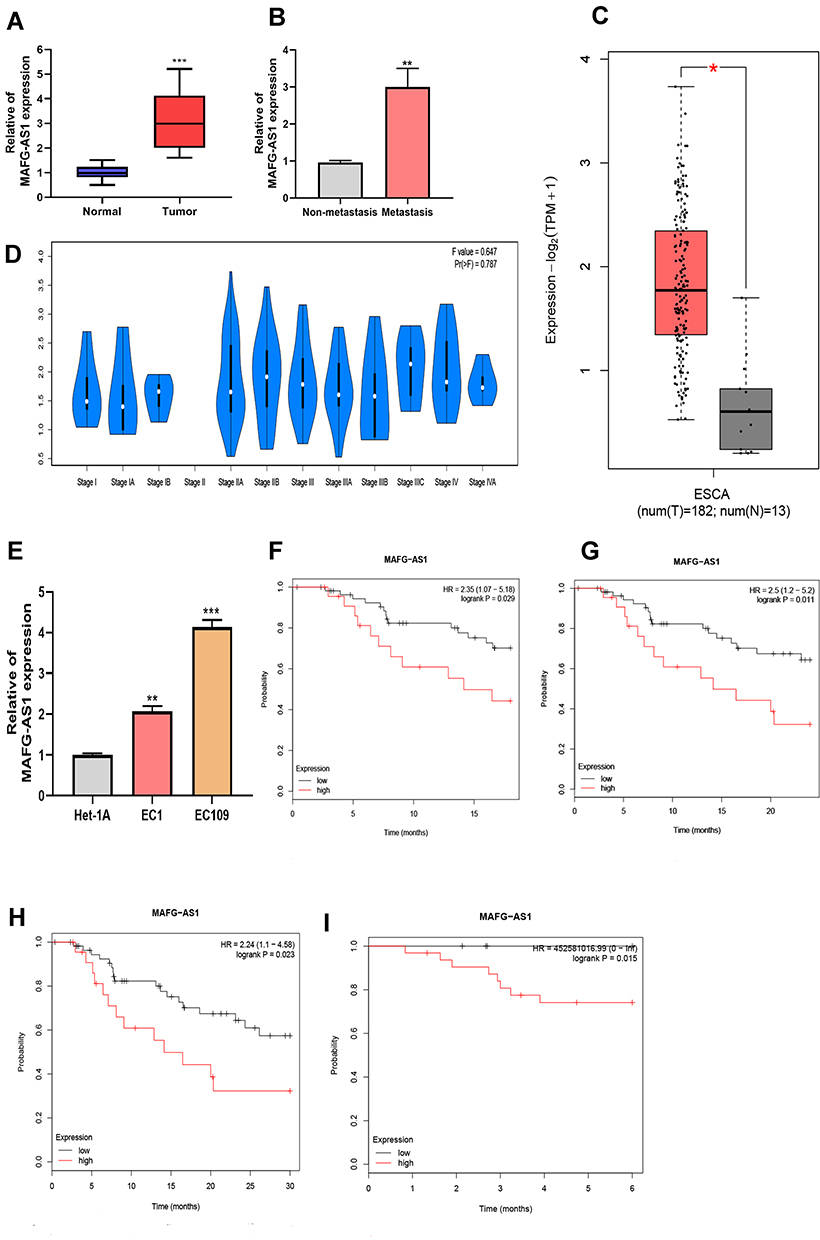 |
Figure 1 The upregulation of MAFG-AS1 in esophageal squamous-cell carcinoma (ESCC) tissue and cells. (A) Expression of MAFG-AS1 was increased in 45 paired ESCC-tissue samples compared to the normal group. (B) Patients with lymphatic metastasis had a higher of MAFG-AS1 expression than those without. (C) Data analysis based on GEPIA revealed high expression of MAFG-AS1 in esophageal carcinoma (ESCA). (D) Bioinformatic analysis from GEPIA showed a correlation between expression of MAFG-AS1 and differentiation stage of ESCA. (E) Increased expression levels of MAFG-AS1 in EC1 and EC109 cells. (F–H) Kaplan–Meier plotter database analysis showed that there was significant meaning when the follow-up threshold of esophageal adenocarcinoma patients was defined as 18months, 24months, and 30months. (I) ESCC patients with lower MAFG-AS1 expression had high disease-free survival. *P<0.05; **P<0.01; ***P<0.001. |
Our results also showed that MAFG-AS1 had higher expression in ESCC cells (EC1 and EC109) than normal esophageal epithelial cells (HET1A), which was consistent with the results for ESCC tissue (Figure 1E). We then investigated the association between MAFG-AS1 expression and patients’ survival rates. Based on Kaplan–Meier Plotter website (https://kmplot.com), overall survival analysis suggested that esophageal adenocarcinoma patients with higher MAFG-AS1 expression possessed lower survival rates when the follow-up threshold was defined as 18 months, 24 months, and 30 months (Figure 1F–H). Meanwhile, relapse-free survival analysis identified ESCC patients with higher MAFG-AS1 expression had lower disease-free survival only when the follow-up threshold was defined as 6 months (Figure 1I). Collectively, these data demonstrated that MAFG-AS1 may serve as a biomarker for ESCC prognosis.
MAFG-AS1 Accelerated ESCC-Cell Proliferation
Noting the dramatically high expression of MAFG-AS1 in ESCC cells, we decided to explore the effect of MAFG-AS1 on ESCC cells. We ectopically silenced and overexpressed MAFG-AS1 in EC1 and EC109 cell lines effectively (Figure 2A and B). WST assays were conducted to assess the impact of MAFG-AS1 on ESCC-cell proliferation. Our findings showed that MAFG-AS1 depletion remarkably decreased the proliferation potential of EC1 and EC109 cells compared to respective controls (Figure 2C and D). Overexpression of MAFG-AS1 produced the opposite effect (Figure 2E and F).
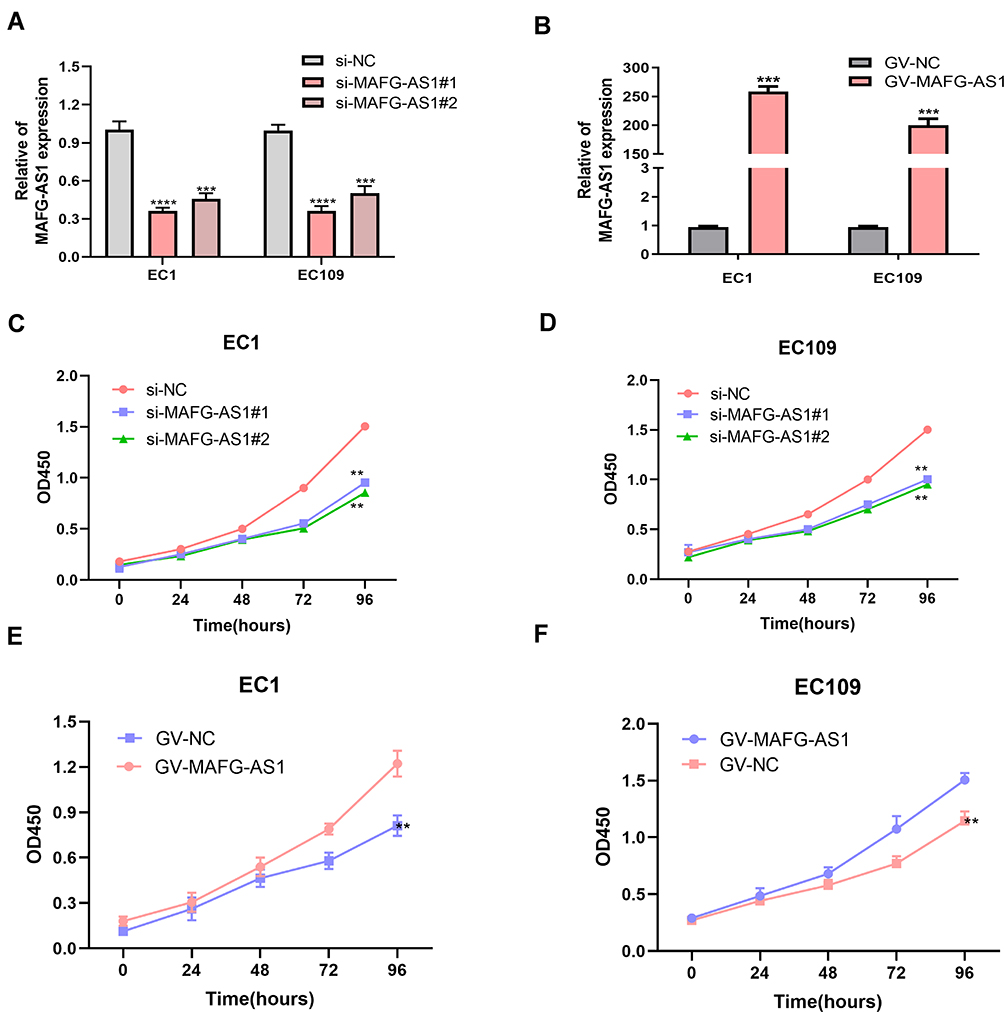 |
Figure 2 MAFG-AS1 promoted esophageal squamous-cell carcinoma (ESCC)-cell proliferation. (A, B) The knock-down and overexpression of MAFG-AS1 in ESCC cells. (C, D) MAFG-AS1 silence inhibited cell proliferation. (E, F) Overexpression of MAFG-AS1 accelerated cell proliferation. **P<0.01; ***P<0.001; ****P<0.0001. |
MAFG-AS1 Promoted ESCC-Cell Migration and Invasion
For further study, we performed transwell assays to clarify the role of MAFG-AS1 in ESCC-cell migration and invasion. Statistical analysis identified that the migration and invasive abilities of EC1 and EC109 cells were significantly impaired following the knockdown of MAFG-AS1 (Figure 3A–D). The experiment on the upregulation of MAFG-AS1 yielded inverse results (Figure 3E–H). Taken together, these research findings indicated that MAFG-AS1 played a pivotal role in controlling malignant ESCC behavior.
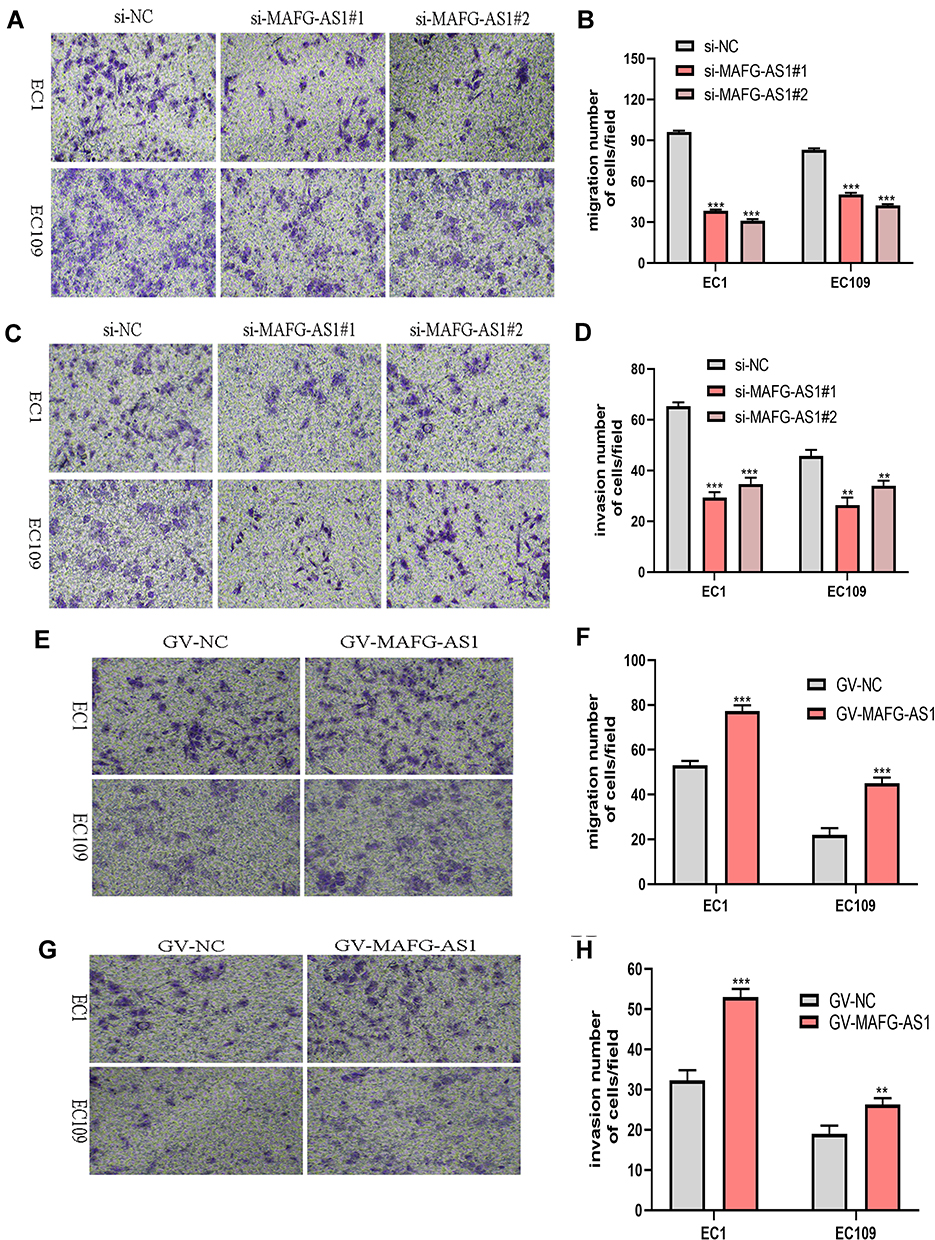 |
Figure 3 MAFG-AS1 contributed to esophageal squamous-cell carcinoma (ESCC)-cell migration and invasion. (A–D) Transwell migration and invasion assays were performed to evaluate the invasive potential of ESCC cells after transfection with si-MAFG-AS1 or negative control. The results are shown in microscopy images (A, C) and statistical graphs (B, D). (E–F) Overexpression of MAFG-AS1 in EC1 and EC109 cells enhanced cell migration. (G–H) Transwell invasion assays produced a similar tendency. **P<0.01; ***P<0.001. |
MAFG-AS1 Regulated Expression of miR143 In Vitro
A considerable amount of literature has been published on ceRNA.18,20 We found that MAFG-AS1 contains a putative miR143-binding area (Figure 4A). In addition, it has been established that miR143 can act as a tumor suppressor in ESCC development and inhibits cell proliferation, migration, and invasion.21,23 To investigate whether MIR143 is a direct target gene of MAFG-AS1, firstly qRT-PCR was used to detect the expression level of miR143 in 45 paired ESCC tissue samples. Results indicated that expression of miR143 was lower in ESCC tissue than adjacent nontumor tissue (Figure 4B). miR143 was also downregulated in ESCC cells (Figure 4C). Then, relationship analysis revealed that miR143 and MAFG-AS1 were negatively correlated (Figure 4D, R2=0.6083, P<0.05). Data from cells also indicated that expression levels of miR143 in EC1 and EC109 cells were upregulated after silencing MAFG-AS1 (Figure 4E). Luciferase-reporter assays also indicated that MAFG-AS1 boundnd with miR143 (Figure 4F). These findings demonstrated that MAFG-AS1 can regulate miR143 expression.
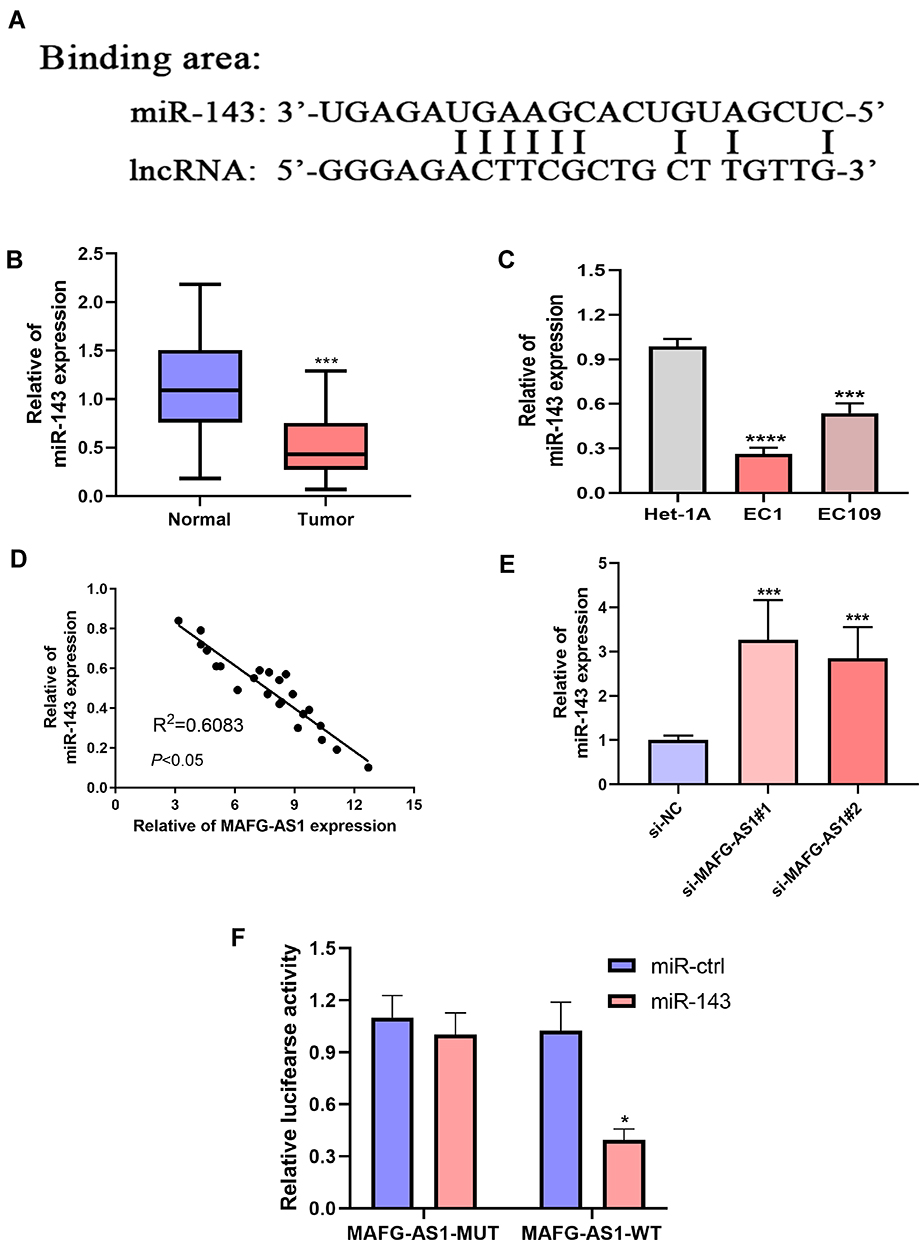 |
Figure 4 MAFG-AS1 negatively regulated miR143 expression. (A) Potential binding sites between MAFG-AS1 and miR143. (B, C) miR143 was downregulated in 45 paired esophageal squamous cell carcinoma (ESCC)-tissue samples and EC1 and EC109 cells. (D) miR143 Expression was negatively linked with MAFG-AS1 expression in 45 paired ESCC tissue samples. (E) There was growth of miR143 in EC1 and EC109 cells transfected with si-MAFG-AS1 or negative-control sequences. (F) Luciferase-reporter assays indicated MAFG-AS1 bound with miR143. *P<0.05; ***P<0.001; ****P<0.0001. |
MAFG-AS1 Promoted ESCC Proliferation by Downregulating miR143
It has been confirmed that miR143 has a tumor-suppressive effect on ESCC. We then designed miR143 inhibitors to achieve knockdown of miR143 in EC109 cells. As shown, those cells transfected with miR143 inhibitors had decrease miR143 expression compared with the NC group (Figure 5A). WST assays suggested that cell-proliferation ability was enhanced after transfection with miR143 inhibitors (Figure 5B). These findings were in keeping with previous reports. For further study, a rescue experiment was performed. We found that knockdown of MAFG-AS1 inhibited cell proliferation, while silencing miR143 abolish suppression partly in EC109 cells (Figure 5C). All these results indicated that MAFG-AS1 controlled ESCC-cell proliferation by regulating miR143.
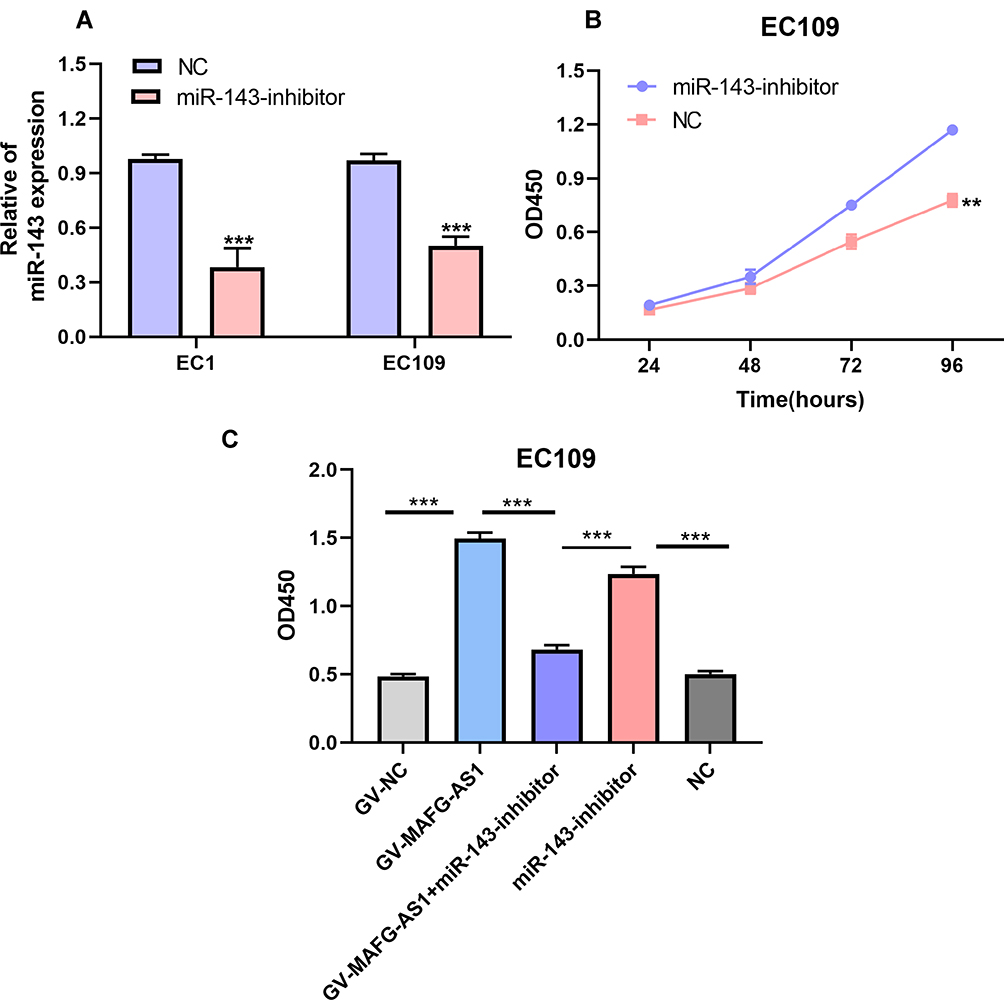 |
Figure 5 Inhibiting miR143 abolished partial increase in cell proliferation induced by MAFG-AS1. (A) miR143 was knocked down effectively in esophageal squamous-cell carcinoma (ESCC) cells. (B) EC109 cell–proliferation ability enhanced after transfection with miR143 inhibitors. (C) WST1 assays indicated that proliferation rates of EC109 cells transfected with GV-MAFG-AS1 decreased after transfection with miR143 inhibitors. **P<0.01; ***P<0.001. |
MAFG-AS1 Regulated LASP1 Expression via miR143
Recent publications24,26 have established that miRNA can induce gene silence by binding to its sequences (most frequently in the 3ʹUTR). To explore more deeply the underlying mechanism of the MAFG-AS1–miR143 axis on ESCC, the TargetScan (http://www.targetscan.org/vert_72), StarBase (http://starbase.sysu.edu.cn), and miRDB (http://mirdb.org) online tools all identified that LASP1 may be a target gene of miR143(binding sites in Figure 6A). It has also been demonstrated that miR143 can serve as a tumor suppressor by targeting LASP1 in ESCC,21 and also conclusivelythat LASP1 acts as an oncogene to accelerate ESCC progression.27 Bioinformatic data from GEPIA indicated that LASP1 was upregulated in ESCA tissue compared to normal esophageal tissue (Figure 6B). Based on evidence, we detected LASP1-expression levels in 45 paired ESCC-tissue and -cell samples with qRT-PCR. Results indicated that LASP1 exhibited high expression in both ESCC tissue and cells (Figure 6C and D). Further research showed LASP1 was upregulated in cells with overexpression of MAFG-AS1 and transfected with miR143 inhibitors (Figure 6E and F). Overall, these findings illustrated MAFG-AS1 controlled LASP1 expression by targeting miR143.
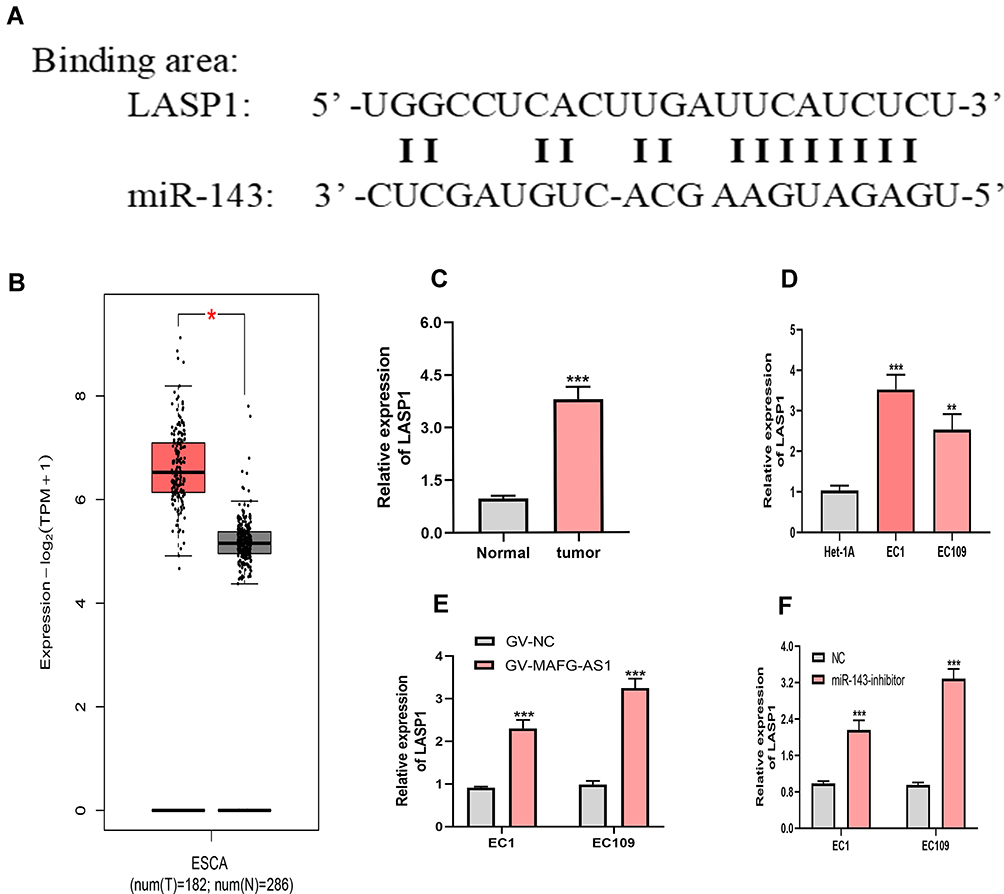 |
Figure 6 MAFG-AS1 regulated LASP1 expression via miR143. (A) Potential binding sites between LASP1 and miR143. (B) Bioinformatic analysis of GEPIA suggested LASP1 was upregulated in ESCA. (C, D) qRT-PCR assays showed LASP1 expression was higher in esophageal squamous-cell carcinoma (ESCC) tissue and cells than normal tissue and cells. (E) There was an increase in LASP1 in ESCC cells transfected with GV-MAFG-AS1 compared with the control group. (F) LASP1 was upregulated in cells transfected with miR143 inhibitors compared to the control group. *P<0.05, **P<0.01, ***P<0.001. |
Discussion
Accumulating evidence has illustrated that lncRNA exerts various roles in cancer progress.28,29 For instance, aberrant expression of lncRNA-HOST2 is closely related to cell proliferation and other malignant biological behavior in hepatocellular carcinoma.30 In breast cancer, lncRNA SNHG20 plays a pivotal role in regulating proliferation, invasion, and migration of cells. Liu et al found that lncRNA-PAGBC was an independent prognostic marker in gallbladder cancer. Prognosis-associated gallbladder cancer lncRNA contributes to tumorigenesis and activates the Akt–mTOR pathway in gallbladder cancer cells by binding to miR133b and miR511.31
Studies have reported that MAFG-AS1 serves as an oncogene in cancer. Cui et al32 proved that MAFG-AS1 was upregulated in CRC tissue and cells. MAFG-AS1 expression is related to tumor–node–metastasis staging of CRC cases. Also, MAFG-AS1 contributes to CRC cell proliferation, the cell cycle, and invasion of biological programs by sponging miR147b and activation of NDUFA4. As an important oncogene, MAFG-AS1 regulates migration, invasion, and boosts epithelial–mesenchymal transition of NSCLC cells.15 MAFG-AS1 exerts carcinogenic influences on lung adenocarcinoma–cell growth and migration through regulating miR744-5p.33 Consistently with previous research, our analysis demonstrated that MAFG-AS1 was distinctly upregulated in both ESCC tissue and cells. MAFG-AS1 expression was linked to lymphatic metastasis in ESCC cases. We also proved that MAFG-AS1 silence alleviated cell proliferation, inhibiting cell migration and invasion. Overexpression of MAFG-AS1 produced inverse results.
A great of deal of literature established that lncRNA serves as a ceRNA to spongemiRNA to exert its functions.18,34 It is a remarkable fact that MAFG-AS1 interacts with miRNA to govern cell functions.15,16 To elucidate the underlying molecular mechanism of MAFG-AS1 in ESCC progression, we used StarBase35 to predict candidate miRNAs binding to MAFG-AS1. We found that miR143 may be a target of MAFG-AS1. miR143 was selected as an optimal biomarker for ESCC, with great diagnostic and prognostic values. miR143 inhibits cell proliferation, invasion, and epithelial–mesenchymal transition in ESCC.23 Our investigations indicated that MAFG-AS1 and miR143 were negatively correlated in ESCC tissue. miR143 expression was increasing after silencing MAFG-AS1 in ESCC cells. miR143 suppressed cell proliferation. This confirmed that miR143 is a tumor-suppressor in ESCC. Also, we observed that knockdown of MAFG-AS1 inhibited cell proliferation, while silencing miR143 abolished the suppression partly. As such, we concluded that MAFG-AS1 promotes ESCC proliferation by downregulating miR143.
LASP1, LIM, and SH3 protein 1, an actin-binding protein, have been reported to be involved in cancer progression.36 LASP1 functions as the direct target of miR143 to accelerate cell proliferation, invasion, and migration in ESCC according to previous research.21,27 In the current study, we also proved that LASP1 mRNA was upregulated in ESCC tissue and cells. In addition, there was a remarkable increase in LASP1 expression after knockdown of MAFG-AS1 in ESCC cells. Depleting miR143 could reverse this trend. Our data showed that MAFG-AS1 regulates LASP1 expression by targeting miR143. However, more biological function assays are required to confirm this finding.
Conclusion
This research demonstrated that MAFG-AS1 is upregulated in ESCC tissue, ans high expression indicates poor prognosis. MAFG-AS1 promoted cell proliferation, migration, and invasion. Moreover, we defined the underlying molecular mechanism of MAFG-AS1 in ESCC. MAFG-AS1 promoted cell proliferation, migration, and invasion by regulating the miR143–LASP1 axis. Finally, these investigations offer some important insights into how lncRNA participates in ESCC development.
Acknowledgment
This project was supported by the Key Scientific Research Project Plan of Colleges and Universities in Henan Province (18A320073).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Torre LA, Bray F, Siegel RL, et al. Global cancer statistics, 2012[J]. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
2. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015[J]. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
3. Chu J, Li H, Xing Y, et al. LncRNA MNX1-AS1 promotes progression of esophageal squamous cell carcinoma by regulating miR-34a/SIRT1 axis[J]. Biomed Pharmacother. 2019;116:109029. doi:10.1016/j.biopha.2019.109029
4. Bao J, Zhou CJ, Zhang JQ, et al. Upregulation of the long noncoding RNA FOXD2-AS1 predicts poor prognosis in esophageal squamous cell carcinoma[J]. Cancer Biomarkers. 2018;21(3):527–533. doi:10.3233/CBM-170260
5. Wu YY, Chen XD, Liang Y, et al. Overexpression of long non-coding RNA SOX2OT promotes esophageal squamous cell carcinoma growth[J]. Cancer Cell Int. 2018;18(1):8. doi:10.1186/s12935-018-0570-7
6. Venkatesh T, Suresh PS, Tsutsumi R. Non-coding RNAs: functions and applications in endocrine-related cancer[J]. Mol Cell Endocrinol. 2015;416:88–96. doi:10.1016/j.mce.2015.08.026
7. Chen HY, Pei HL, Hu WT, et al. Long non-coding RNA CRYBG3 regulates glycolysis of lung cancer cells by interacting with lactate dehydrogenase A[J]. J Cancer. 2018;9(14):2580–2588. doi:10.7150/jca.24896
8. Mao WD, Guo ZY, Dai YC, et al. LNC CRYBG3 inhibits tumor growth by inducing M phase arrest[J]. J Cancer. 2019;10(12):2764–2770. doi:10.7150/jca.31703
9. Liu F, Yuan JH, Huang JF, et al. Long noncoding RNA FTX inhibits hepatocellular carcinoma proliferation and metastasis by binding MCM2 and miR-374a[J]. Oncogene. 2016;35(41):5422–5434. doi:10.1038/onc.2016.80
10. Cheng DZ, Bao CC, Zhang XX, et al. LncRNA PRNCR1 interacts with HEY2 to abolish miR-448-mediated growth inhibition in non-small cell lung cancer[J]. Biomed Pharmacother. 2018;107:1540–1547. doi:10.1016/j.biopha.2018.08.105
11. Gu LQ, Xing XL, Cai H, et al. Long non-coding RNA DILC suppresses cell proliferation and metastasis in colorectal cancer[J]. Gene. 2018;666:18–26. doi:10.1016/j.gene.2018.03.100
12. Liu LP, Shuai TK, Li B, et al. Long non-coding RNA lnc-GNAT1-1 inhibits gastric cancer cell proliferation and invasion through the Wnt/-catenin pathway in Helicobacter pylori infection[J]. Mol Med Rep. 2018;18(4):4009–4015. doi:10.3892/mmr.2018.9405
13. Shen FF, Pan Y, Yang HJ, et al. Decreased expression of SPINT1-AS1 and SPINT1 mRNA might be independent unfavorable prognostic indicators in esophageal squamous cell carcinoma[J]. Onco Targets Ther. 2019;12:4755–4763. doi:10.2147/OTT.S206448
14. Huang JB, Li Y, Lu ZL, et al. Long non-coding RNA GAS5 is induced by interferons and plays an antitumor role in esophageal squamous cell carcinoma[J]. Cancer Med. 2018;7(7):3157–3167. doi:10.1002/cam4.1524
15. Jia YC, Wang JY, Liu YY, et al. LncRNA MAFG-AS1 facilitates the migration and invasion of NSCLC cell via sponging miR-339-5p from MMP15[J]. Cell Biol Int. 2019;43(4):384–393. doi:10.1002/cbin.11092
16. Li H, Zhang GY, Pan CH, et al. LncRNA MAFG-AS1 promotes the aggressiveness of breast carcinoma through regulating miR-339-5p/MMP15[J]. Eur Rev Med Pharmacol Sci. 2019;23(7):2838–2846. doi:10.26355/eurrev_201904_17561
17. Yang L, Ye Y, Chu J, et al. Long noncoding RNA FEZF1-AS1 promotes the motility of esophageal squamous cell carcinoma through Wnt/beta-catenin pathway[J]. Cancer Manag Res. 2019;11:4425–4435. doi:10.2147/CMAR.S196004
18. Zhao CC, Jiao Y, Zhang YY, et al. Lnc SMAD5-AS1 as ceRNA inhibit proliferation of diffuse large B cell lymphoma via Wnt/beta-catenin pathway by sponging miR-135b-5p to elevate expression of APC[J]. Cell Death Dis. 2019;10(4):15. doi:10.1038/s41419-019-1479-3
19. Zhang B, Zhou M, Zou L, et al. Long non-coding RNA LOXL1-AS1 acts as a ceRNA for miR-324-3p to contribute to cholangiocarcinoma progression via modulation of ATP-binding cassette transporter A1[J]. Biochem Biophys Res Commun. 2019;513(4):827–833. doi:10.1016/j.bbrc.2019.04.089
20. Li B, Shi C, Zhao JM, et al. Long noncoding RNA CCAT1 functions as a ceRNA to antagonize the effect of miR-410 on the down-regulation of ITPKB in human HCT-116 and HCT-8 cells[J]. Oncotarget. 2017;8(54):92855–92863. doi:10.18632/oncotarget.21612
21. Liu HF, Zheng ML, Zhao YC, et al. miR-143 inhibits migration and invasion through regulating LASP1 in human esophageal cancer[J]. Int J Clin Exp Pathol. 2019;12(2):466–476.
22. Ni Y, Meng L, Wang L, et al. MicroRNA-143 functions as a tumor suppressor in human esophageal squamous cell carcinoma[J]. Gene. 2013;517(2):197–204. doi:10.1016/j.gene.2012.12.031
23. He Z, Yi J, Liu X, et al. MiR-143-3p functions as a tumor suppressor by regulating cell proliferation, invasion and epithelial-mesenchymal transition by targeting QKI-5 in esophageal squamous cell carcinoma[J]. Mol Cancer. 2016;15(1):51. doi:10.1186/s12943-016-0533-3
24. Shi HJ, Shen HM, Xu J, et al. MiR-143-3p suppresses the progression of ovarian cancer[J]. Am J Transl Res. 2018;10(3):866–874.
25. Tokumaru Y, Tajirika T, Sugito N, et al. Synthetic miR-143 Inhibits Growth of HER2-Positive Gastric Cancer Cells by Suppressing KRAS Networks Including DDX6 RNA Helicase[J]. Int J Mol Sci. 2019;20(7):13. doi:10.3390/ijms20071697
26. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function[J]. Cell. 2004;116(2):281–297. doi:10.1016/S0092-8674(04)00045-5
27. He B, Yin B, Wang B, et al. Overexpression of LASP1 is associated with proliferation, migration and invasion in esophageal squamous cell carcinoma[J]. Oncol Rep. 2013;29(3):1115–1123. doi:10.3892/or.2012.2199
28. Sun BC, Dang Y, Zhang F, et al. Long non-coding RNA RP1-163G9.1 is downregulated in gastric adenocarcinoma and is associated with a poor prognosis[J]. Oncol Rep. 2019;41(6):3575–3585. doi:10.3892/or.2019.7127
29. Li ZH, Jiang P, Li J, et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma[J]. Oncogene. 2018;37(28):3822–3838. doi:10.1038/s41388-018-0237-9
30. Liu RT, Cao JL, Yan CQ, et al. Effects of LncRNA-HOST2 on cell proliferation, migration, invasion and apoptosis of human hepatocellular carcinoma cell line SMMC-7721[J]. Biosci Rep. 2017;37(2):12. doi:10.1042/BSR20160532
31. Wu X-S, Wang F, Li H-F, et al. LncRNA-PAGBC acts as a microRNA sponge and promotes gallbladder tumorigenesis[J]. EMBO Rep. 2017;18(10):1837–1853. doi:10.15252/embr.201744147
32. Cui S, Yang X, Zhang L, et al. LncRNA MAFG-AS1 promotes the progression of colorectal cancer by sponging miR-147b and activation of NDUFA4[J]. Biochem Biophys Res Commun. 2018;506(1):251–258. doi:10.1016/j.bbrc.2018.10.112
33. Sui Y, Lin G, Zheng Y, et al. LncRNA MAFG-AS1 boosts the proliferation of lung adenocarcinoma cells via regulating miR-744-5p/MAFG axis[J]. Eur J Pharmacol. 2019;859:172465. doi:10.1016/j.ejphar.2019.172465
34. Bai T, Liu Y, Li B. LncRNA LOXL1-AS1/miR-let-7a-5p/EGFR-related pathway regulates the doxorubicin resistance of prostate cancer DU-145 cells. IUBMB Life. 2019;71(10):1537–1551. doi:10.1002/iub.2075
35. Li J-H, Liu S, Zhou H, et al. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data`J. Nucleic Acids Res. 2014;42(Database issue):D92–D97. doi:10.1093/nar/gkt1248
36. Gao Q, Tang L, Wu L, et al. LASP1 promotes nasopharyngeal carcinoma progression through negatively regulation of the tumor suppressor PTEN[J]. Cell Death Dis. 2018;9(3):393. doi:10.1038/s41419-018-0443-y
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.