Back to Journals » OncoTargets and Therapy » Volume 13
LncRNA GA-Binding Protein Transcription Factor Subunit Beta-1 Antisense RNA 1 Inhibits Renal Carcinoma Growth Through an MiR-1246/Phosphoenolpyruvate Carboxykinase 1 Pathway
Authors Gao S, Zhang F, Sun H ![]() , Yang X
, Yang X
Received 7 April 2020
Accepted for publication 1 July 2020
Published 12 July 2020 Volume 2020:13 Pages 6827—6836
DOI https://doi.org/10.2147/OTT.S257275
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Carlos E Vigil
This paper has been retracted.
Shuang Gao,1,2 Feng Zhang,3 Hanxue Sun,1 Xianghong Yang1
1Department of Pathology, Shengjing Hospital of China Medical University, Shenyang 110004, People’s Republic of China; 2Department of Pathology, People’s Hospital of Liaoning Province, Shenyang 110016, People’s Republic of China; 3Department of Urology, Shengjing Hospital of China Medical University, Shenyang 110004, People’s Republic of China
Correspondence: Xianghong Yang Email [email protected]
Objective: To clarify the role and mechanism of GABPB1-AS1 in renal cell carcinoma.
Methods: We collected 48 pairs of tumor and adjacent normal tissues from patients with clear cell renal cell carcinoma (ccRCC). Both 786-o and caki-1 ccRCC cell lines were transfected with GA-binding protein transcription factor subunit beta-1 antisense RNA 1 (GABPB1-AS1), miR-1246, or small interfering RNA phosphoenolpyruvate carboxykinase 1 (siPCK1) vectors. RNA expression was examined by quantitative reverse transcription-PCR and protein expression by Western blot. Cell proliferation was measured by Cell Counting Kit-8 assays. Cell migration and invasion were measured by transwell assays. Targeting relationships between genes were tested by luciferase reporter gene assays.
Results: Lower GABPB1-AS1 expression was found in ccRCC cells and tissues. GABPB1-AS1 expression was inversely associated with tumor size, TNM stage, and Furhman stage. High GABPB1-AS1 expression was associated with a better prognosis. GABPB1-AS1 overexpression significantly inhibited proliferation, migration, and invasion by 786-o and caki-1 cells. GABPB1-AS1 overexpression reduced tumor weights in xenograft experiments. Luciferase reporter assays showed that miR-1246 overexpression significantly inhibited the luciferase activity of 786-o and caki-1 cells transfected with wild-type (WT)-GABPB1-AS1 or WT-PCK1. Knockdown of PCK1 weakened the inhibition of proliferation, migration, and invasion induced by GABPB1-AS1 overexpression in 786-o and caki-1 cells.
Conclusion: GABPB1-AS1 inhibits ccRCC growth and plays a tumor suppressor role through an miR-1246/PCK1 axis.
Keywords: renal cell carcinoma, GABPB1-AS1, miR-1246, PCK1
Introduction
Renal cancer is a common malignant tumor of the urogenital system that accounts for about 4% of all new cancers.1 Renal cancer usually originates from renal epithelium, with clear cell renal cell carcinoma (ccRCC) the most common histological subtype.2 Clear cell renal cell carcinoma is insensitive to radiotherapy and chemotherapy, with treatment becoming very difficult when reaching an advanced stage.3 Although targeted therapy and immunotherapy have a certain therapeutic effect on advanced renal cell carcinoma, the majority of patients will die of tumor metastasis.4 Although great progress has been made in elucidating renal cell carcinoma at the gene level, the treatment of metastatic ccRCC still remains a challenge.5
Long noncoding RNA (lncRNA) is a noncoding RNA with more than 200 nucleotides; most have an mRNA-like structure but do not encode protein. LncRNAs can interact with chromosomes, proteins or RNA to regulate genes. Duan et al reported that lncRNA long intergenic non-protein coding RNA, p53-induced transcript (LINC-PINT) predicts a poor prognosis and promotes proliferation by the enhancer of zeste homolog 2 (EZH2) in ccRCC.6 LncRNA GA-binding protein transcription factor subunit beta-1 antisense RNA 1 (GABPB1-AS1) is the antisense RNA of GABPB1 mRNA, which is located in the cytoplasm and has a total length of 4139nt.7 The expression level of GABPB1-AS1 changes in response to various stimuli such as stress.8 GABPB1-AS1 acts as a tumor suppressor gene in hepatocellular carcinoma (HCC).9 However, the role of GABPB1-AS1 in renal cell carcinoma has not been elucidated. MicroRNA (miRNA) is a small single-strand non-coding molecule that can participate in many biological behaviors, such as cell differentiation, proliferation, and apoptosis, by regulating the translation or transcription of target genes.10 MicroRNA-1246 is widely studied as being oncogene-like in non-small cell lung cancer (NSCLC),11 HCC,12 and cervical cancer.13 Phosphoenolpyruvate carboxykinase (PCK1) is the rate-limiting enzyme of gluconeogenesis in the liver and kidney and was the first isoenzyme to be located in the cytoplasm.14 The overexpression of PCK1 induces renal cancer cell death by promoting gluconeogenesis.15 Tuo et al reported PCK1 is a tumor suppressor gene in HCC.16,17
In the present study, the effect of GABPB1-AS1 on ccRCC cells and the association between GABPB1-AS1, miRNA-1246, and PCK1 was assessed. This article reports on the effect of GABPB1-AS1 on the inhibition of ccRCC growth via the miR-1246/PCK1 axis. GABPB1-AS1 may potentially be a useful biomarker for the diagnosis of ccRCC and may be an effective target for treatment.
Methods
Patients and Tissues
Forty-eight pairs of ccRCC and adjacent tissues were collected from patients who had undergone radical nephrectomy between 2010 January and 2015 January at the Shengjing Hospital of China Medical University. The study was approved by the Ethics Committee of Shengjing Hospital, which abided by the guidelines of the Declaration of Helsinki. All patients signed informed consent. The clinicopathological characteristics of patients are outlined in Table 1.
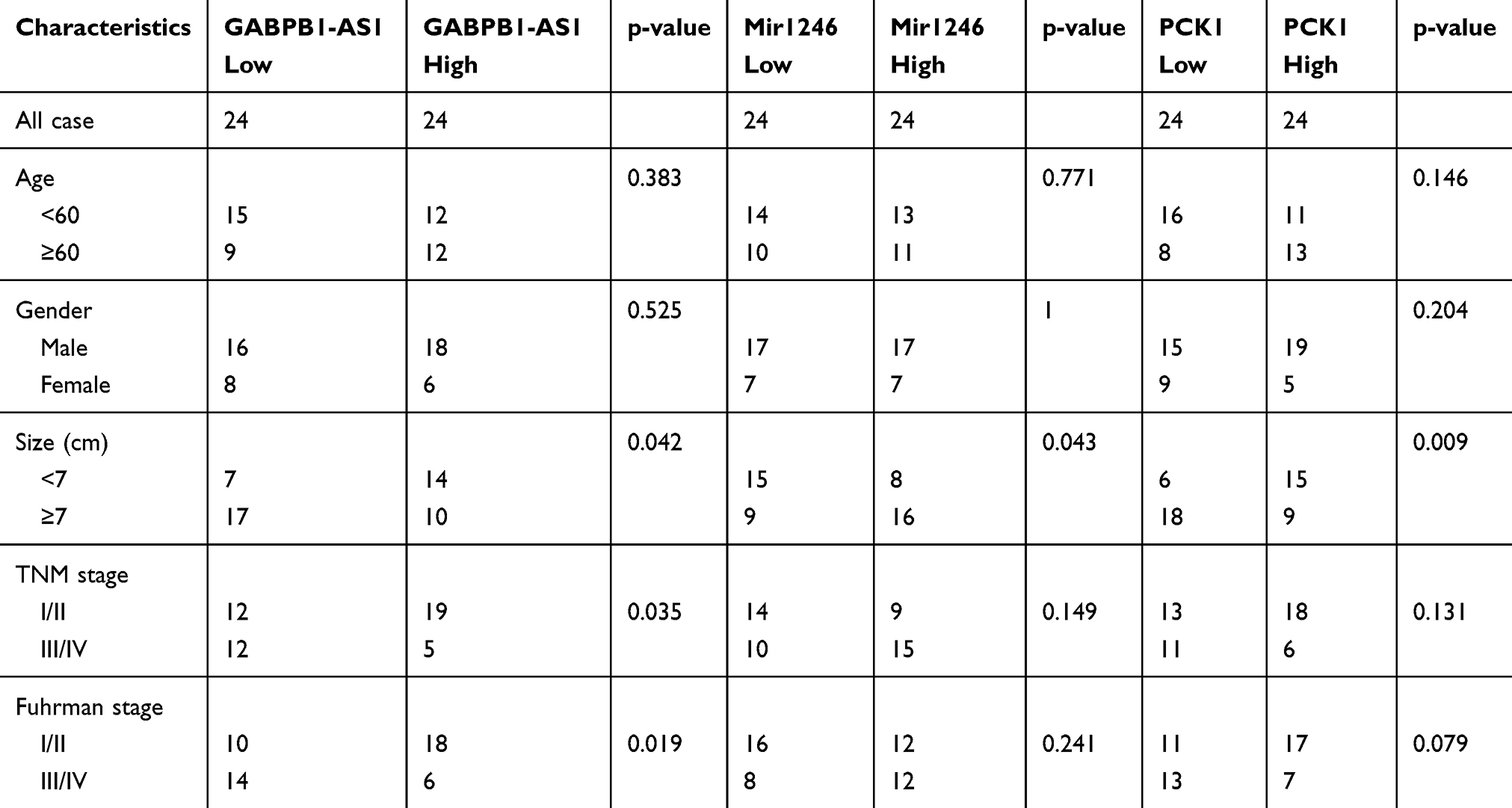 |
Table 1 Clinicopathological Characteristics of High and Low Expression of GABPB1-AS1, Mir1246 and PCK1 |
Cell Lines and Transfection
The ccRCC cell lines, 786-o and caki-1, and an HK-2 cell line were purchased from Shanghai Biological Technology. Cell lines were cultured in RPMI-1640 with 10% fetal bovine serum (FBS) in an incubator at 37°C, 5% CO2. Vectors containing full-length pcDNA3.1- GABPB1-AS1, miR-1246 mimics, and siPCK1 were purchased from GeneCopoeia (Guangzhou, China). Cells (786-o and caki-1) were transfected with GABPB1-AS1 vector, miR-1246 mimics, siPCK1 or blank controls using Lipofectamine 2000 (Thermo Fisher Scientific, Waltham, MA, USA) in accordance with the instructions of the manufacturer.
qRT-PCR
The total RNA of tissues or cells was extracted using TRIzol reagent. Total RNA was reverse-transcribed. Quantitative reverse transcription (qRT)-PCR was performed with a Roche PCR system in accordance with reaction procedures. Relative expression was normalized to GAPDH and the fold-change was calculated using the 2−ΔΔCt method.
CCK8 Assays
Cells were seeded in a 96-well plate at 5×103 cells per well. After 24, 48, and 72 h, 10 μL of CCK8 agent was added to each well. Cells were incubated for another 2 h. The optical density of each well was measured using a microplate reader at a wavelength of 450 nm.
Transwell Invasion and Migration Assays
A transwell system and Matrigel (BD Biosciences, New York, USA) were used according to the manufacturers’ instructions for invasion assays. Cells were added to the upper chamber, which was pre-coated with Matrigel, and continuously cultured in serum-free RPMI 1640. The lower chamber was filled with RPMI 1640 and 10% FBS. Cells were incubated for 48 h, and the cells in the upper chamber were removed. The cells in the bottom chamber were fixed using 4% paraformaldehyde and stained with 0.5% crystal violet. The number of cells was determined by a microscope. For the migration assay, cells were added to the upper chamber, which did not have a Matrigel coating. The other steps were performed as described for the invasion assay.
Luciferase Reporter Assay
Cells (786-o and caki-1) were placed into a 24-well plate. Cells were transfected with wild-type (WT), mut (mutated) GABPB1-AS1 or PCK1 reporter plasmids synthesized by GenePharma (Shanghai, China) using miR-1246 mimics or controls. Cells were cultured for 48 h, and relative luciferase activity was tested by luciferase reporter assay system (Promega, Madison, WI, USA).
Tumor Growth in vivo Assay
Animal experiments were approved by the Ethics Committee of Shengjing Hospital of China Medical University (No. 2017PS278K). This study was carried out in accordance with the Guidelines for the Care and Use of Laboratory Animals (Ministry of Science and Technology of China, 2006). To evaluate in vivo tumor growth, 5×106 786-o cells transfected with overexpressed GABPB1-AS1 vector or normal control (NC)-vector were injected into the subcutaneous flanks of 6-week-old BALB/c nude mice. All mice were raised under the same conditions for 42 days to allow the formation of solid tumors. Tumor volumes were determined every week by measuring the length and width of each tumor. Volumes of tumors were calculated according to the formula: (width2 × length)/2. Mice were then sacrificed and tumors were weighed after removal.
Western Blot
Protein concentrations were measured using a bicinchoninic protein assay kit (Pierce) after total protein was extracted by radioimmunoprecipitation assay lysis buffer. Proteins (30 μg per lane) were loaded on a 10% gel, resolved using SDS–polyacrylamide gel electrophoresis and transferred to polyvinylidene (PVDF) membranes. Membranes were blocked using 5% skimmed milk, and subsequently incubated overnight with primary antibodies against target proteins at 4°C, and then secondary antibody for 2 h at 20°C. PVDF membranes were incubated with chemiluminescence reagent. GAPDH was used as an internal reference. Protein bands were measured using ImageJ software.
Immunohistochemical Staining
Formalin-fixed paraffin-embedded tissue samples were cut into 4 μm-thick sections, deparaffinized, and rehydrated in an alcohol gradient starting from high-percentage ethanol to distilled water. For quenching endogenous peroxidase activity, sections on glass slides were immersed in 3% hydrogen peroxide at room temperature for 15 min. Antigen retrieval involved boiling slides in 10 mM sodium citrate buffer (pH 6.0) for 3 min, followed by cooling to room temperature. Sections were incubated with an anti-PCK1 rabbit IgG antibody (diluted 1:100; Proteintech, Rosemont, IL, USA) at 4°C overnight. After washing with phosphate-buffered saline (PBS), slides were incubated with a horseradish peroxidase-linked antibody against the anti-PCK1 rabbit IgG antibody (Proteintech) for 30 min at room temperature, followed by washing in PBS and 3,3ʹ-diaminobenzidine tetrahydrochloride staining. The sections were counterstained with hematoxylin.
The immunostaining was evaluated by two pathologists who were blinded to patients’ information. A semiquantitative immunohistochemistry score on a scale of 0–300 was estimated for each sample by multiplying the staining intensity (0, no staining; 1, weak; 2, moderate; and 3, strong) and the percentage of cells (0–100%) at each intensity level. We selected a cut-off score (60) to divide the low and high expression of PCK1.
Statistical Analysis
All data are presented as mean ± standard (SD). Student’s t-test or one-way ANOVA analysis was used for comparisons between groups. P < 0.05 was considered statistically significant. All the experiments were repeated three times.
Results
GABPB1-AS1 Expression Was Downregulated in RCC Tissues
GABPB1-AS1 expression was measured by qRT-PCR. We found that the expression of GABPB1-AS1 was markedly downregulated in ccRCC tissues in comparison with adjacent tissues (Figure 1A). We subsequently found that GABPB1-AS1 expression was significantly downregulated in 786-o and caki-1 RCC cell lines compared to the normal human renal cell line, HK-2 (Figure 1B). The association between GABPB1-AS1 expression and clinicopathological features of RCC patients is shown in Table 1. GABPB1-AS1 expression was inversely associated with tumor size, TNM stage, and Fuhrman stage. Kaplan–Meier analysis revealed that RCC patients with higher GABPB1-AS1 expression had a better survival rate (Figure 1C).
 |
Figure 1 GABPB1-AS1 expression was downregulated in RCC tissues. (A) Expression of GABPB1-AS1 in 48 pairs of RCC tissues compared with adjacent normal tissues by qRT-PCR. (B) Expression of GABPB1-AS1 in cell lines by qRT-PCR. (C) RCC patients’ OS rate was measured by Kaplan–Meier curve analysis according to GABPB1-AS1 expression. The data show the mean ± SD. *p < 0.05. Abbreviations: GABPB1-AS1, GA-binding protein transcription factor subunit beta-1 antisense RNA 1; OS, overall survival; qRT-PCR, quantitative reverse transcription PCR; RCC, renal cell carcinoma; SD, standard deviation. |
GABPB1-AS1 Overexpression Inhibits Growth of RCC Cells
To investigate the effects of GABPB-1-AS1 on RCC cells, the overexpressed GABPB1-AS1 vector was transfected into 786-o and caki-1 cells. GABPB1-AS1 was found to be upregulated in 786-o and caki-1 cells after transfection (Figure 2A). We showed that GABPB1-AS1 overexpression significantly inhibited the proliferation of 786-o and caki-1 cells as shown by CCK8 assay (Figure 2B). We found that GABPB1-AS1 overexpression markedly suppressed the migration and invasion of 786-o and caki-1 cells (Figure 2C and D). We also found that GABPB1-AS1 overexpression led to reduced tumor weights in xenograft experiments (Figure 2E and F). This suggests that overexpression of GABPB1-AS1 suppressed RCC cell proliferation in vivo. Above all, the results show that GABPB1-AS1 is a tumor suppressor gene.
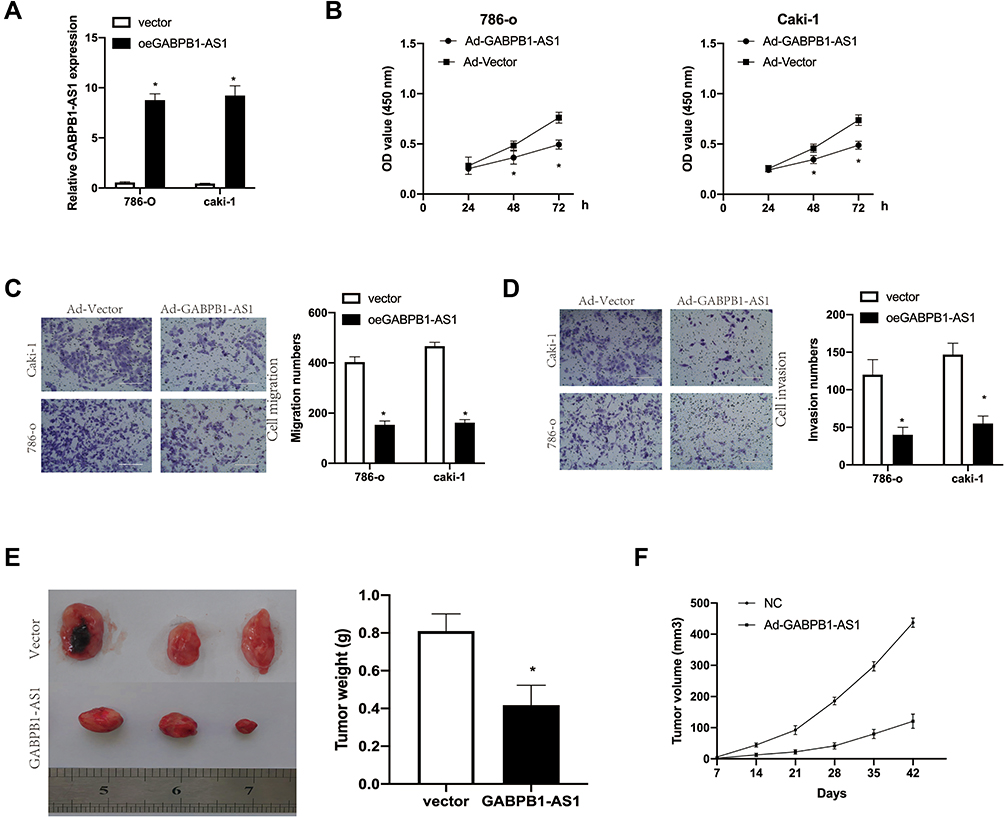 |
Figure 2 (A) GABPB1-AS1 overexpression inhibits growth of RCC cells. Relative expression of GABPB1-AS1 in 786-o and caki-1 cells transfected with GABPB1-AS1 vector or a blank control. (B) CCK8 assays to measure cell proliferation in 786-o and caki-1 cells transfected with GABPB1-AS1 vector or an empty vector control. (C and D) Cell migration and invasion in 786-o and caki-1 cells transfected with GABPB1-AS1 vector or an empty vector control were evaluated by transwell assays. (E) Tumors were weighed after a xenograft experiment. (F) Tumor volumes were measured. The data show the mean ± SD. *p < 0.05. Abbreviations: CCK8, Cell Counting Kit-8; GABPB1-AS1, GA-binding protein transcription factor subunit beta-1 antisense RNA 1; RCC, renal cell carcinoma; SD, standard deviation. |
GABPB1-AS1 Stimulated PCK1 Expression by Sponging miR-1246 in RCC
We first found that miR-1246 was a potential target of GABPB1-AS1 through the biological website, DIANA-LncBasev2.18 We then predicted that PCK1 may be a possible combined target of miR-1246 through TargetScan.19 The association between miR-1246 or PCK1 expression and clinicopathological features of RCC patients is also shown in Table 1. We established luciferase reporter plasmids that contain wild-type or mutant binding sites of miR-1246 to evaluate our prediction (Figure 3A). Subsequent luciferase reporter assays showed that miR-1246 overexpression significantly inhibited the luciferase activity of 786-o and caki-1 cells transfected with WT-GABPB1-AS1 or WT-PCK1-3ʹ untranslated region (UTR), but the luciferase activity of the mutant binding sites of miR-1246 was not inhibited (Figure 3B and C). Overexpression of GABPB1-AS1 restrained miR-1246 expression in 786-o and caki-1 cells (Figure 3D). Overexpression of miR-1246 inhibited both the mRNA and protein PCK1 expression of 786-o and caki-1 cells. But overexpression of GABPB1-AS1 at the same time restored the effect of miR-1246 overexpression, lowering PCK1 expression in 786-o and caki-1 cells (Figure 3E and F). Finally, we found that the miR-1246 expression negatively correlated with GABPB1-AS1 or PCK1 in RCC tissues (Figure 3G).
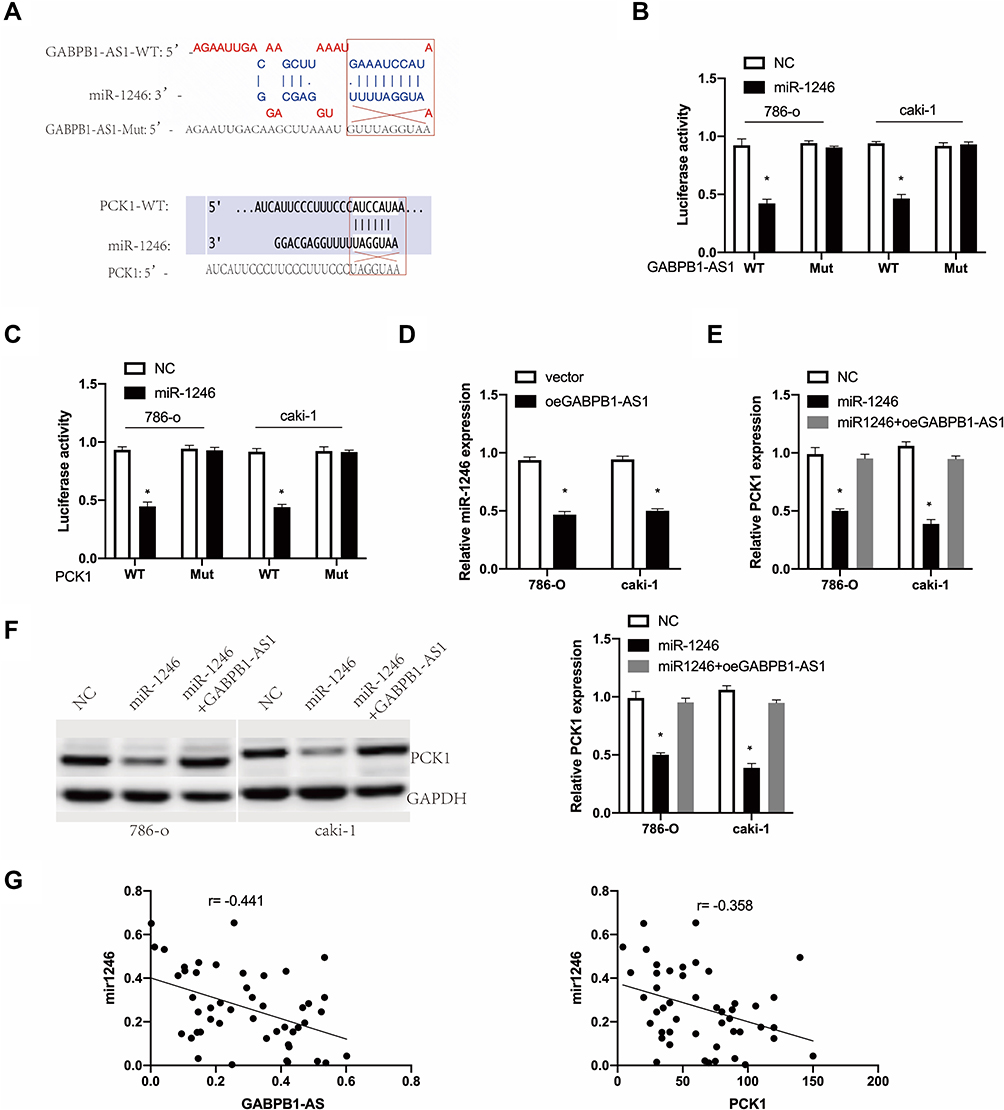 |
Figure 3 GABPB1-AS1 stimulated PCK1 by sponging miR-1246 in RCC. (A) Potential binding sites of miR-1246 with GABPB1-AS1 and PCK1 mRNA in wild type (WT) and mutant type (mut). (B) The luciferase activity of LUC-GABPB1-AS1 or GABPB1-AS1-mut in 786-o and caki-1 cells transfected with miR-1246. (C) The luciferase activity of LUC-PCK1 or LUC-PCK1-mut in 786-o and caki-1 cells transfected with miR-1246. (D) MiR-1246 expression in 786-o and caki-1 cells transfected with GABPB1-AS1 vector or a blank control. (E) PCK1 mRNA expression in 786-o and caki-1 cells co-transfected with miR-1246 or miR-1246 and GABPB1-AS1 vector. (F) PCK1 protein expression in 786-o and caki-1 cells transfected with miR-1246 or miR-1246 with GABPB1-AS1 vector. (G) Correlation of miR-1246 with GABPB1-AS1 or PCK1 in RCC tissues. The data show the mean ± SD. *p < 0.05. Abbreviations: GABPB1-AS1, GA-binding protein transcription factor subunit beta-1 antisense RNA 1; miRNA, microRNA; PKC1, phosphoenolpyruvate carboxykinase 1; RCC, renal cell carcinoma; SD, standard deviation. |
GABPB1-AS1 Inhibited RCC Cell Proliferation, Migration, and Invasion by PCK1
We found that PCK1 was significantly downregulated in RCC tissues compared to adjacent normal tissues in GSE40435 datasets (Figure 4A). We also found that PCK1 was expressed at a lower level compared to adjacent tissues in RCC using immunohistochemistry (Figure 4B). We transfected 786-o and caki-1 cells with GABPB1-AS1 vector and siPCK1 or siNC, and performed CCK8 and transwell assays. We found that the knockdown of PCK1 weakened the inhibition of proliferation, migration, and invasion induced by GABPB1-AS1 overexpression (Figure 4C-E).
 |
Figure 4 GABPB1-AS1 inhibited RCC cell proliferation, migration, and invasion induced by PCK1. (A) Expression of PCK1 in RCC tissues and adjacent normal tissues of GSE40435 datasets. (B) PCK1 expression of RCC tissues compared to adjacent tissues shown by immunohistochemistry. (C) Cell proliferation was measured by CCK8 assays in 786-o and caki-1 cells transfected with the indicated plasmids. (D and E) Cell migration and invasion were measured by transwell assays in 786-o and caki-1 cells, which were transfected with the labeled plasmids. The data show the mean ± SD. *p < 0.05. Abbreviations: CCK8, Cell Counting Kit-8; GABPB1-AS1, GA-binding protein transcription factor subunit beta-1 antisense RNA 1; PKC1, phosphoenolpyruvate carboxykinase 1; RCC, renal cell carcinoma; SD, standard deviation. |
Discussion
GA-binding protein, also called nuclear respiration factor-2 (NRF2), is a transcription activator that affects cellular antioxidant capacity by regulating the expression of genes encoding peroxidases.9 Salem et al showed that NFR2 promotes cancer development by inhibiting autophagy in breast cancer cells.20 NRF2 has alpha, beta, and gamma subunits, while GABPB1 encodes the beta subunit of NRF2. The activation of GABPB1 promotes the development of several cancers.21 Chen et al reported that the knockdown of GABPB1 inhibits cell proliferation via p21 induction in renal cell carcinoma.22 Antisense RNA is one of the most common forms of lncRNA, with GABPB1-AS1 the antisense RNA of GABPB1. Generally, the function of antisense lncRNA is the opposite of its gene, which usually interferes with the normal translation of the coding gene. Under oxidative stress, the accumulation of GABPB1-AS1 was induced.8 Qi et al found that GAPBPB1-AS1 inhibited the antioxidant ability of cancer cells and cell proliferation by inhibiting the expression of GABPB1 and peroxiredoxin 5 (PRDX5).9 To our knowledge, we are the first to find that the expression of GABPB1-AS1 was low in renal cell carcinoma and that this correlated with clinicopathological features.
In recent years, intense research has focused on the transcriptome whereby lncRNA regulates the expression and function of miRNA through a mechanism of competing endogenous RNA to regulate the expression level of target genes.23 This theory has also been widely shown in renal cancer. Yang et al described how homeobox A11 antisense lncRNA (HOXA11-AS) sponged miR-146b-5p and how upregulated matrix metalloproteinase 16 expression stimulated growth and invasion by RCC cells.24 Hong et al reported that HOTAIR regulated hypoxia-inducible factor (HIF)-1alpha/AXL signaling through the inhibition of miR-217 in renal cell carcinoma.25 Qu et al discovered that the upregulation of myocardial infarction associated transcript (MIAT) regulated LOX-like protein (LOXL) 2 expression by competitively binding miR-29c in ccRCC.26 We speculated as to whether GABPB1-AS1 also works through this mechanism and used the biological website, DIANA-LncBasev2,18 to predict the binding site between GABPB1-AS1 and miR-1246. We performed a luciferase reporter assay to investigate the interaction between these factors and found a negative correlation between GABPB1-AS1 and miR-1246 in RCC.
More recently, miR-1246 has been studied as an oncogene in several tumors. MiR-1246 on the circulating miRNA landscape was highlighted as a promising diagnostic biomarker in high-grade serous ovarian carcinoma.27 Colorectal cancer cell–derived microvesicles containing miR-1246 promoted angiogenesis by activating Smad 1/5/8 signaling elicited by PML downregulation in endothelial cells.28 MiR-1246 enhanced migration and invasion through cell adhesion molecule (CADM)1 in HCC.12 The current evidence suggests that miR-1246 has a role as an oncogene, which is consistent with our findings.
We predict that PCK1 may be a target gene of miR-1246 using TargetScan.19 Two PCK genes exist encoding for cytoplasmic (PCPEK-C or PCK1) and mitochondrial (PEPCK-M or PCK2) isoforms in mammalian cells.29 Liu et al reported the expression of both PCK1 and PCK2 genes were downregulated in HCC. The overexpression of PCK1 inhibits cell survival and promotes tricarboxylic acid cycle cataplerosis, oxidative stress and apoptosis.30 Tuo et al reported that PCK1 played a role as a tumor suppressor gene in liver cancer.16 Not only did the reprogramming of glucose metabolism occur but also that of fatty acid metabolism, which are tumor characteristics. The accumulation of abnormal lipid metabolism is a typical feature of ccRCC. We verified that PCK1 was expressed at a low level in RCC, which was associated with survival and prognosis. Recently, Shi et al reported PCK1 regulates glycolysis and tumor progression in ccRCC through LDHA.31 We performed a luciferase reporter assay to investigate the interaction between PCK1 and miR-1246. After the overexpression of miR-1246, PCK1 expression decreased significantly, which indicated that miR-1246 could negatively regulate the transcription or translation of PCK1. When overexpressing miR-1246 and GABPB1-AS1 together, we found that PCK1 expression was higher than in the miR-1246 alone overexpression group. These results suggest that GABPB1-AS1 reversed the inhibiting action of miR-1246 on PCK1. We also found that PCK1 knockdown led to the recovery of cell proliferation, migration, and invasion in RCC, which were reduced by GABPB1-AS1 overexpression.
Conclusion
In summary, our results reveal that GABPB1-AS1 inhibits RCC growth and plays a tumor suppressor role through an miR-1246/PCK1 pathway axis.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
2. Ljungberg B, Bensalah K, Canfield S, et al. EAU guidelines on renal cell carcinoma: 2014 update. Eur Urol. 2015;67(5):913–924. doi:10.1016/j.eururo.2015.01.005
3. Linehan WM, Srinivasan R, Schmidt LS. The genetic basis of kidney cancer: a metabolic disease. Nat Rev Urol. 2010;7(5):277–285. doi:10.1038/nrurol.2010.47
4. Wei EY, Hsieh JJ. A river model to map convergent cancer evolution and guide therapy in RCC. Nat Rev Urol. 2015;12:706. doi:10.1038/nrurol.2015.260
5. Braga EA, Fridman MV, Loginov VI, Dmitriev AA, Morozov SG. Molecular mechanisms in clear cell renal cell carcinoma: role of miRNAs and hypermethylated miRNA genes in crucial oncogenic pathways and processes. Front Genet. 2019;10:320. doi:10.3389/fgene.2019.00320
6. Duan J, Ma X, Shi J, et al. Long noncoding RNA LINC-PINT promotes proliferation through EZH2 and predicts poor prognosis in clear cell renal cell carcinoma. Onco Targets Ther. 2019;12:4729–4740. doi:10.2147/OTT.S202938
7. Tani H, Torimura M. Identification of short-lived long non-coding RNAs as surrogate indicators for chemical stress response. Biochem Biophys Res Commun. 2013;439(4):547–551. doi:10.1016/j.bbrc.2013.09.006
8. Tani H, Onuma Y, Ito Y, Torimura M. Long non-coding RNAs as surrogate indicators for chemical stress responses in human-induced pluripotent stem cells. PLoS One. 2014;9(8):e106282. doi:10.1371/journal.pone.0106282
9. Qi W, Li Z, Xia L, et al. LncRNA GABPB1-AS1 and GABPB1 regulate oxidative stress during erastin-induced ferroptosis in HepG2 hepatocellular carcinoma cells. Sci Rep. 2019;9(1):16185. doi:10.1038/s41598-019-52837-8
10. Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120(1):15–20. doi:10.1016/j.cell.2004.12.035
11. Huang W, Li H, Luo R. The microRNA-1246 promotes metastasis in non-small cell lung cancer by targeting cytoplasmic polyadenylation element-binding protein 4. Diagn Pathol. 2015;10(1):127. doi:10.1186/s13000-015-0366-1
12. Sun Z, Meng C, Wang S, et al. MicroRNA-1246 enhances migration and invasion through CADM1 in hepatocellular carcinoma. BMC Cancer. 2014;14:616. doi:10.1186/1471-2407-14-616
13. Du P, Lai YH, Yao DS, Chen JY, Ding N. Downregulation of microRNA-1246 inhibits tumor growth and promotes apoptosis of cervical cancer cells by targeting thrombospondin-2. Oncol Lett. 2019;18(3):2491–2499. doi:10.3892/ol.2019.10571
14. Montal ED, Dewi R, Bhalla K, et al. PEPCK coordinates the regulation of central carbon metabolism to promote cancer cell growth. Mol Cell. 2015;60(4):571–583. doi:10.1016/j.molcel.2015.09.025
15. Khan MW, Biswas D, Ghosh M, Mandloi S, Chakrabarti S, Chakrabarti P. mTORC2 controls cancer cell survival by modulating gluconeogenesis. Cell Death Discov. 2015;1:15016.
16. Tuo L, Xiang J, Pan X, et al. PCK1 negatively regulates cell cycle progression and hepatoma cell proliferation via the AMPK/p27(Kip1) axis. J Exp Clin Cancer Res. 2019;38(1):50. doi:10.1186/s13046-019-1029-y
17. Tuo L, Xiang J, Pan X, et al. PCK1 downregulation promotes TXNRD1 expression and hepatoma cell growth via the Nrf2/Keap1 pathway. Front Oncol. 2018;8:611. doi:10.3389/fonc.2018.00611
18. Paraskevopoulou MD, Vlachos IS, Karagkouni D, et al. DIANA-LncBase v2: indexing microRNA targets on non-coding transcripts. Nucleic Acids Res. 2016;44(D1):D231–238. doi:10.1093/nar/gkv1270
19. Agarwal V, Bell GW, Nam JW, Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. Elife. 2015;4.
20. Salem AF, Whitaker-Menezes D, Howell A, Sotgia F, Lisanti MP. Mitochondrial biogenesis in epithelial cancer cells promotes breast cancer tumor growth and confers autophagy resistance. Cell Cycle. 2012;11(22):4174–4180. doi:10.4161/cc.22376
21. Bell RJ, Rube HT, Kreig A, et al. Cancer. The transcription factor GABP selectively binds and activates the mutant TERT promoter in cancer. Science. 2015;348(6238):1036–1039. doi:10.1126/science.aab0015
22. Chen SC, Yen MC, Chen FW, et al. Knockdown of GA-binding protein subunit beta1 inhibits cell proliferation via p21 induction in renal cell carcinoma. Int J Oncol. 2018;53(2):886–894. doi:10.3892/ijo.2018.4411
23. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
24. Yang FQ, Zhang JQ, Jin JJ, et al. HOXA11-AS promotes the growth and invasion of renal cancer by sponging miR-146b-5p to upregulate MMP16 expression. J Cell Physiol. 2018;233(12):9611–9619. doi:10.1002/jcp.26864
25. Hong Q, Li O, Zheng W, et al. LncRNA HOTAIR regulates HIF-1alpha/AXL signaling through inhibition of miR-217 in renal cell carcinoma. Cell Death Dis. 2017;8(5):e2772. doi:10.1038/cddis.2017.181
26. Qu Y, Xiao H, Xiao W, et al. Upregulation of MIAT regulates LOXL2 expression by competitively binding MiR-29c in clear cell renal cell carcinoma. Cell Physiol Biochem. 2018;48(3):1075–1087. doi:10.1159/000491974
27. Todeschini P, Salviato E, Paracchini L, et al. Circulating miRNA landscape identifies miR-1246 as promising diagnostic biomarker in high-grade serous ovarian carcinoma: a validation across two independent cohorts. Cancer Lett. 2017;388:320–327. doi:10.1016/j.canlet.2016.12.017
28. Yamada N, Tsujimura N, Kumazaki M, et al. Colorectal cancer cell-derived microvesicles containing microRNA-1246 promote angiogenesis by activating Smad 1/5/8 signaling elicited by PML down-regulation in endothelial cells. Biochim Biophys Acta. 2014;1839(11):1256–1272. doi:10.1016/j.bbagrm.2014.09.002
29. Yang J, Kalhan SC, Hanson RW. What is the metabolic role of phosphoenolpyruvate carboxykinase? J Biol Chem. 2009;284(40):27025–27029. doi:10.1074/jbc.R109.040543
30. Liu MX, Jin L, Sun SJ, et al. Metabolic reprogramming by PCK1 promotes TCA cataplerosis, oxidative stress and apoptosis in liver cancer cells and suppresses hepatocellular carcinoma. Oncogene. 2018;37(12):1637–1653. doi:10.1038/s41388-017-0070-6
31. Shi L, An S, Liu Y, Liu J, Wang F. PCK1 regulates glycolysis and tumor progression in clear cell renal cell carcinoma through LDHA. Onco Targets Ther. 2020;13:2613–2627. doi:10.2147/OTT.S241717
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.