Back to Journals » Clinical Interventions in Aging » Volume 14
LncRNA CASC11 is upregulated in postmenopausal osteoporosis and is correlated with TNF-α
Authors Yu H, Zhou W, Yan W, Xu Z, Xie Y, Zhang P ![]()
Received 18 February 2019
Accepted for publication 24 May 2019
Published 20 September 2019 Volume 2019:14 Pages 1663—1669
DOI https://doi.org/10.2147/CIA.S205796
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Richard Walker
Haotao Yu, Wei Zhou, Weiming Yan, Zhongqi Xu, Yinhao Xie, Ping Zhang
Department of Orthopedics, Division 1, The Third Affiliated Hospital of Guangzhou Medical University, Guangzhou City, Guangdong Province 510000, People’s Republic of China
Correspondence: Ping Zhang
Department of Orthopedics, Division 1, The Third Affiliated Hospital of Guangzhou Medical University, No. 63 Duobao Street, Liwan District, Guangzhou City, Guangdong Province 510000, People’s Republic of China
Tel/Fax +86 0 208 134 6935
Email [email protected]
Purpose: In this study, we aimed to investigate the role of lncRNA cancer susceptibility 11 (CASC11) and tumor necrosis factor (TNF-α) in postmenopausal osteoporosis (POP).
Methods and materials: POP patients and healthy controls were included in this study and levels of CASC11 and TNF-α in plasma of those participants were measured by qPCR and Western blot, respectively. ROC curve was used for diagnostic analysis. Patients were followed up for 2 years and the correlations between the levels of CASC11 and TNF-α and disease conditions were analyzed.
Results: We found that CASC11 and TNF-α were both upregulated in plasma of POP patients than in healthy controls. Plasma levels of CASC11 and TNF-α were positively correlated in both POP patients and in healthy controls. Upregulation of CASC11 and TNF-α distinguished POP patients from healthy controls. Treatment and follow-up study showed that high CASC11 levels were significantly correlated with prolonged treatment course and high recurrence rate. Plasma levels of CASC11 and TNF-α decreased after treatment. CASC11 overexpression led to upregulated TNF-α in osteoclasts.
Conclusion: CASC11 is upregulated in POP and is correlated with TNF-α.
Keywords: postmenopausal osteoporosis, CASC11, TNF-α, osteoclasts
Introduction
Only 2% of transcriptions are related to protein-coding genes, and the vast majority of transcripts are transcribed by non-coding RNA (ncRNA) genes.1 Long ncRNAs (lncRNAs) are a group of non-protein coding RNA transcripts composed of more than 200 nucleotides.2,3 Due to the lack of protein-coding ability, lncRNAs participate in both physiological and pathological processes by regulating downstream genes at epigenetic, posttranscriptional and translational levels.4,5 Studies in the last two decades have shown that lncRNAs are critical determinants in human diseases.6 However, function of most lncRNAs remains unknown, which hinders the application of lncRNAs in disease prediction and treatment.
Postmenopausal osteoporosis (POP) is a metabolic and systemic bone disorder resulted from the decreased production of estrogen in ovary after menopause.7 POP affects about half of females in their middle and old age.8 The main cause of POP is the disrupted balance between bone formation by osteoblasts and bone resorption osteoclasts,7 in which lncRNAs have been proven to play critical roles.9 Therefore, functional characterization of lncRNAs in POP may facilitate the development of therapeutic approaches. However, the function of most lncRNAs in this disease remains unknown. CASC11 is a recently identified oncogenic lncRNA in several types of cancer.10–12 Our preliminary microarray data revealed its upregulation in POP patients and its positive correlation with TNF-α (data not shown), which contribute to POP.13 Therefore, exploration of the interactions between CASC11 and TNF-α in POP may provide new insights to the pathogenesis of this disease. In the present study, we investigated the role of CASC 11 in POP and its potential interactions with TNF-α.
Materials and methods
Patients
Our study included 67 patients with POP as well as 66 healthy postmenopausal female controls who were admitted by the Third Affiliated Hospital of Guangzhou Medical University from January 2014 to January 2016. All patients showed decrease in bone mineral density (BMD) is more than 2.5 SD compared with normal population. BMD was measured by dual-energy X-ray absorptiometry. Patients’ inclusion criteria: 1) POP patients who were diagnosed and treated for the first time; 2) patient complete treatment and 2-year follow up. Exclusion criteria: 1) patients who were transferred from other hospital and received therapy before admission; 2) other medical conditions were observed. The 66 healthy controls were enrolled in the physical health center of the Third Affiliated Hospital of Guangzhou Medical University to match the age of patient group. Age of patient group ranged from 47 to 69 years, with a mean age of 56.8±4.9 years. Age of control group ranged from 46 to 68 years, with a mean age of 56.8±4.9 years. This study was approved by Ethics Committee of the Third Affiliated Hospital of Guangzhou Medical University before the admission of patients. All participants signed informed consent. This study was conducted in accordance with the Declaration of Helsinki.
Plasma and cell line
Fasting blood (5 mL) was extracted from 67 patients with POP under fasting conditions before treatment and on the day of discharge. Fasting blood (5 mL) was also extracted from healthy controls on the day of admission. Blood was kept in EDTA treated tubes, followed by centrifugation at 1200 g for 10 mins to remove blood cells and collect plasma.
Treatment and follow-up
Intramuscular injection of elcatonin was performed for all patients every 3 or 4 days. BMD of all patients returned to normal range after maximum of 3.5 months’ treatment. Patients were followed-up for 2 years to record recurrence. The patients failed to complete this study were excluded from this study.
Osteoblasts
Human primary osteoblasts were purchased from Sigma-Aldrich (St. Lous, MO, USA). Cells were cultivated under the conditions recommended by Sigma-Aldrich and collected at passage 5–7 for subsequent experiments.
Total RNA extraction and real-time quantitative PCR (RT-qPCR)
To detect the expression of CASC11, total RNAs were extracted using RNAzol reagent (Sigma-Aldrich (St. Lous, MO, USA), RevertAid RT Reverse Transcription Kit (Thermo Fisher Scientific., Inc.) was used to perform reverse transcription and Luna® Universal One-Step RT-qPCR Kit (NEB, Ipswich, MA, USA) was used to prepare all PCR reaction systems. Applied Biosystems 2720 Thermal Cycler was used to perform all PCR reactions with GAPDH as endogenous control. Primers of CASC11 and GAPDH were from Sangon (Shanghai, People's Republic of China). Expression of CASC11 was normalized to GAPDH using 2−ΔΔCT method.
Enzyme-linked immunosorbent assay (ELISA)
Human TNF-alpha Quantikine ELISA Kit (R&D Systems) was used to measure plasma levels of TNF-α with al operations performed in strict accordance with manufacturer’s instructions. Plasma levels of TNF-α were normalized to pg/ml.
Cell transfection
Vectors and cell transfection
pIRSE2 vector expressing CASC11 and empty pIRSE2 vector from Sangon (Shanghai). Lipofectamine 2000 reagent (Thermo Fisher Scientific. lnc.,) was used to transfect 10 nM vectors into osteoclasts with all operations performed in strict accordance with manufacturers’ instructions. Control cells were un-transfected cells. Negative control cells were cells transfected with empty vectors. Cells were harvested at 24 hrs after cell transfection to perform other experiments.
Western blot
TNF-α in osteoclasts after CASC11 was detected by Western blot. Briefly, Monarch® Total RNA Miniprep Kit (NEB) was used to extract total RNAs from osteoclasts. Following denaturing, gel electrophoresis was performed using 10% SDS-PAGE. After gel transfer to PVDF membranes, membranes were blocked in 5% non-fat milk for 2 hrs at denaturing. Membranes were then incubated with rabbit anti-human TNF-α (1:1200, ab9635, Abcam) or GAPDH (1:1800, ab8245, Abcam) overnight at 4°C. After that, incubation with goat anti-rabbit IgG-HRP secondary antibody (1:1500, MBS435036, MyBioSource) was performed for 2 hrs at room temperature. After ECL (Sigma-Aldrich, USA) method was used for signal development and signals were normalized using Image J software.
Statistical analysis
All experiments were performed in triplicate manner and data were recorded as mean±standard deviation. GraphPad Prism 6 software was used to perform all statistical analyses. Comparisons between patients and controls and comparisons of treatment course were performed unpaired t-test. Comparisons between pre-therapy and post-therapy levels of CASC11 and TNF-α were performed by paired test. Comparisons of TNF-α among cells with different treatments were performed by one-way ANOVA and Tukey test. Comparison of recurrence rate was performed by Chi-square test. Diagnostic values of CASC11 and TNF-α for POP were analyzed by ROC curve analysis with POP patients as true positive cases and healthy controls as true negative cases. Correlation analyses between expression levels of CASC11 and TNF-α were analyzed using Pearson’s correlation coefficient. Differences were statistically significant when p<0.05.
Results
CASC11 and TNF-α were both upregulated in plasma of POP patients
Plasma levels of CASC11 and TNF-α were measured by RT-qPCR and ELISA and compared between POP patients and healthy controls by unpaired t-test. Compared with healthy controls, plasma levels of CASC11 (Figure 1A, 2.32-fold change) and TNF-α (Figure 1B, 1.67-fold change) were significantly upregulated in patients with POP (p<0.05).
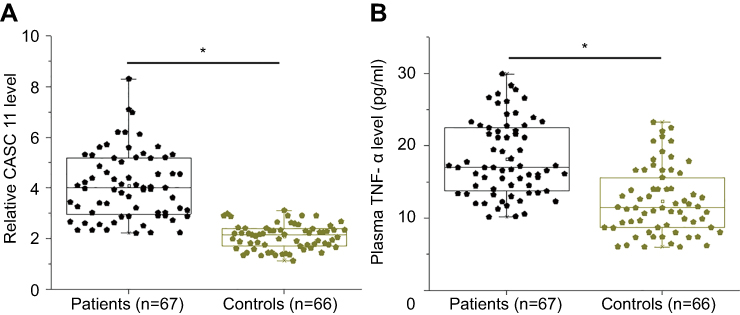 |
Figure 1 CASC11 and TNF-α were both upregulated in plasma of postmenopausal osteoporosis patients. RT-qPCR and ELISA results showed that plasma levels of CASC11 (A) and TNF-α (B) were significantly upregulated in patients with postmenopausal osteoporosis than in healthy controls (*p<0.05). |
Plasma CASC11 and TNF-α have diagnostic potentials for POP
Diagnostic values of CASC11 and TNF-α for POP were analyzed by ROC curve analysis with POP patients as true positive cases and healthy controls as true negative cases. For plasma lncRNA CASC 11, area under the curve was 0.96 (standard error: 0.017, confidence interval: 0.92–0.98, p<0.001, Figure 2A). For plasma TNF-α, area under the curve was 0.80 (standard error: 0.037, confidence interval: 0.73–0.88, p<0.001, Figure 2B).
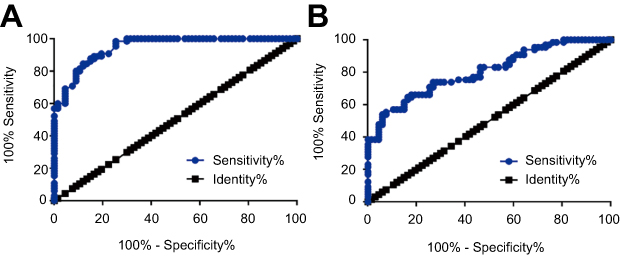 |
Figure 2 Plasma CASC11 and TNF-α have diagnostic potentials for postmenopausal osteoporosis. ROC curve analysis showed that upregulation of CASC11 (A) and TNF-α (B) distinguished postmenopausal osteoporosis patients from healthy controls. |
Plasma levels of CASC11 and TNF-α were positively correlated
Correlation analyses between expression levels of CASC11 and TNF-α were analyzed using Pearson’s correlation coefficient. As shown in Figure 3, a significant and positive correlation between CASC11 and TNF-α were found in both POP patients (Figure 3A, R square=0.79; p<0.01) and healthy controls (Figure 3B, R square=0.74, p<0.01).
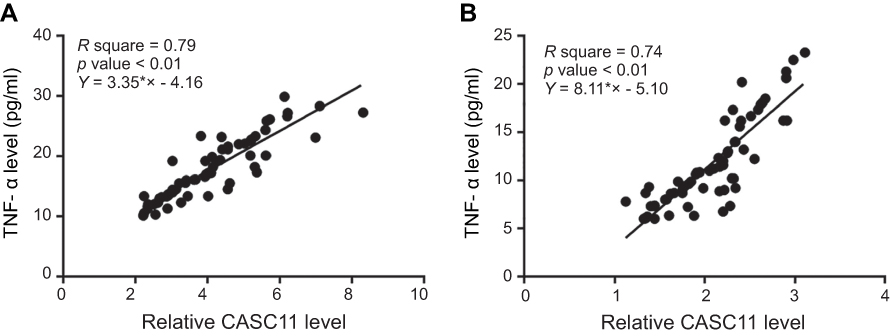 |
Figure 3 Plasma levels of CASC11 and TNF-α were positively correlated. Pearson’s correlation coefficient revealed a significant and positive correlation between CASC11 and TNF-α in both postmenopausal osteoporosis patients (A) and healthy controls (B). |
CASC11 overexpression led to TNF-α upregulation in osteoclasts
CASC11 was overexpressed in osteoclasts to further investigate the interaction between CASC11 and TNF-α. Compared with control (C) and negative control cells, CASC11 overexpression led to upregulated TNF-α in osteoclasts (Figure 4, p<0.05).
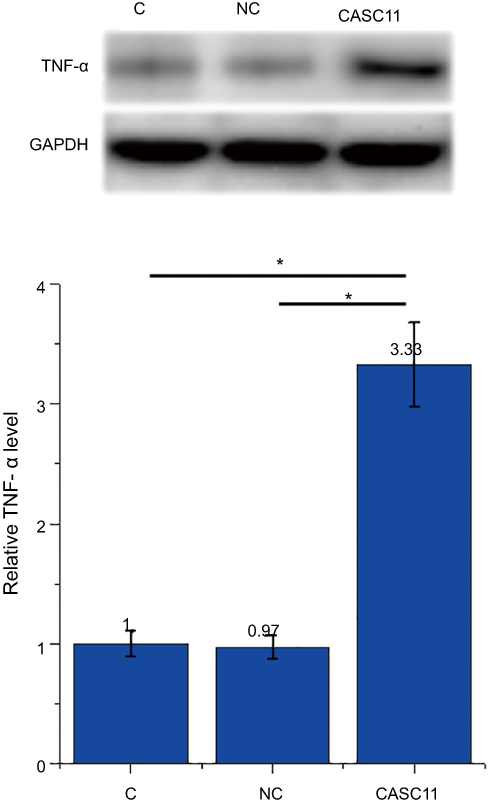 |
Figure 4 CASC11 overexpression led to TNF-α upregulation in osteoclasts. Western blot results showed that CASC11 overexpression led to upregulated TNF-α in osteoclasts (*p<0.05). |
Plasma levels of CASC11 and TNF-α decreased after treatment
Plasma levels of CASC11 and TNF-α in POP patients were also measured on the day of discharge. Compared with pre-therapy levels (paired t-test), plasma levels of CASC11 (Figure 5A) and TNF-α (Figure 5B) decreased after treatment (post-therapy) (p<0.05).
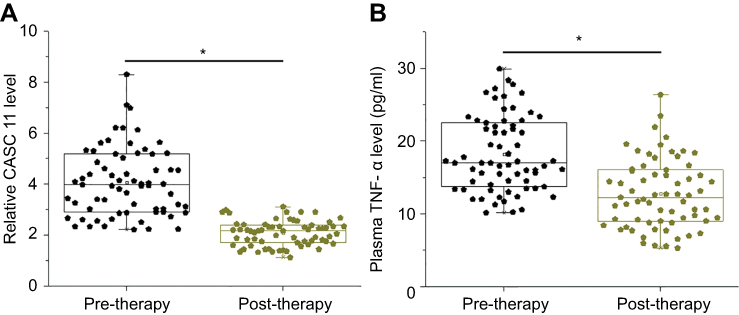 |
Figure 5 Plasma levels of CASC11 and TNF-α decreased after treatment. Compared with pre-therapy levels, plasma levels of CASC11 (A) and TNF-α (B) decreased after treatment (post-therapy) (*p<0.05). |
High CASC11 levels were significantly correlated with prolonged treatment course and high recurrence rate
Based on pre-therapy levels of plasma CASC11, patients were divided into high (n=31) and low (n=36) level CASC11 groups according to Youden’s index. Compared with low-level CASC11 group, treatment course was significantly longer in high-level CASC11 group (Figure 6, p<0.05). Based on post-therapy levels of plasma CASC11, patients were divided into higher (n=30) and low (n=37) level CASC11 groups according to Youden’s index. During follow-up, recurrence occurred in 18 cases of high-level CASC11 group and only 9 cases occurred in low-level CASC11 group, a significant difference was found between 2 groups (Chi-square =8.76, p=0.0031).
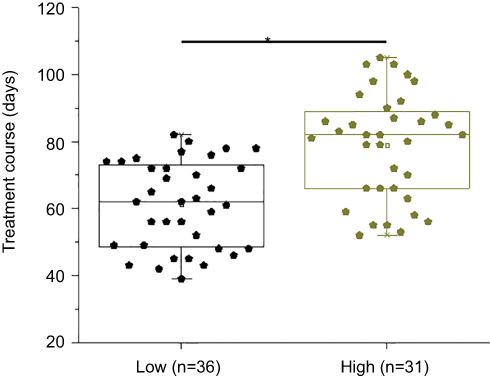 |
Figure 6 High CASC11 levels were significantly correlated with prolonged treatment course. Compared with low-level CASC11 group, treatment course was significantly longer in high-level CASC11 group (*p<0.05). |
Discussion
The development and progression of POP require the involvement of lncRNA, while the function of most lncRNAs in this disease remains unknown. The key finding of the present study is that CASC11 is upregulated in POP and may promote disease by upregulating TNF-α.
TNF-α is a major contributor of POP.13 TNF-α is upregulated in POP and upregulated expression of TNF-α leads to the activation of osteoclasts and increased bone resorption.14 Consistent with previous studies, upregulated TNF-α was observed in POP compared with healthy controls. In effect, upregulation of TNF-α effectively distinguished POP patients from healthy controls. It is known that TNF-α can be regulated by certain lncRNAs, such as HOTAIR.15 In the present study, we proved that CASC11 can upregulate TNF-α in osteoclasts. Therefore, CASC11 may promote bone resorption by upregulating TNF-α. However, molecular mechanism of the regulation of TNF-α by CASC11 is unknown. Our study did not observe significantly changed expression of TNF-α mRNA after CASC11 overexpression. Therefore, CASC11 may regulate the accumulation and/or degradation of TNF-α.
With proper therapy, treatment outcomes of POP patients are generally satisfactory.16 Our study showed that CASC11 can be used as biomarkers to reflect the treatment course of POP. A major challenge in the clinical treatment of POP is the high recurrence rate.17 In the present study, 27 of 67 patients suffered from recurrence within 2 years after discharge, accounting for 40.3%. Therefore, prevention of recurrence is still critical. Our study showed that high plasma level of CASC11 on the day of discharge is significantly correlated with high recurrence rate. Therefore, measurement of plasma levels of CASC11 may provide guidance of the prevention of the recurrence of POP.
It has been reported that CASC11 can promote the expression of MMP7,12 which mediates the release of TNF-α.18 Therefore, MMP7 may mediate the interaction between CASC11 and TNF-α. In addition, CASC11 can also interact with other key players in POP, such as TGF-β1.19 Our future studies will try to analyze the interactions between CASC11 and these factors in POP. It is worth noting that this study is limited by the small sample size. In addition, genetic variants of CASC11 and TNF genes, which may have different functions, were not analyzed in the present study. Our future studies will try to solve these problems.
However, our study lacks in vivo experiments. Our future studies will try to perform animal model experiments to further confirm the functions of CASC11 in POP.
Conclusion
CASC11 is upregulated in POP and may upregulate TNF-α to promote disease development.
Ethics approval and informed consent
The protocol of the present study was approved by the Ethics Review Committee of the Third Affiliated Hospital of Guangzhou Medical University. All patients signed informed consent.
Acknowledgment
We thank the financial support from the Common Guiding Project of 2018 Guangzhou Health and Family Planning Commission (20181A011084) and Public Welfare Reseach and Capacity Building Special Project of 2014 Guangdong Provincial Science and Technology Department (2014A020212367).
Author contributions
Haotao Yu, Wei Zhou, Weiming Yan, Zhongqi Xu, Yinhao Xie and Ping Zhang have made substantial contributions to conception and design, acquisition of data, and analysis and interpretation of data. Haotao Yu and Ping Zhang were involved in drafting the manuscript or revising it critically for important intellectual content. Haotao Yu, Wei Zhou, Weiming Yan, Zhongqi Xu, Yinhao Xie and Ping Zhang gave final approval of the version to be published. Each author agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Mattick JS. Non-coding RNAs: the architects of eukaryotic complexity. EMBO Rep. 2001;2:986–991. doi:10.1093/embo-reports/kve230
2. Fatica A, Bozzoni I. Long non-coding RNAs: new players in cell differentiation and development. Nat Rev Genet. 2014;15:7–21. doi:10.1038/nrg3606
3. Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10:155–159. doi:10.1038/nrg2521
4. Terashima M, Tange S, Ishimura A, et al. MEG3 long noncoding RNA contributes to the epigenetic regulation of epithelial-mesenchymal transition in lung cancer cell lines. J Biol Chem. 2017;292:82–99. doi:10.1074/jbc.M116.750950
5. Rajpathak SN, Vellarikkal SK, Patowary A, et al. Human 45, X fibroblast transcriptome reveals distinct differentially expressed genes including long noncoding RNAs potentially associated with the pathophysiology of Turner syndrome. PLoS One. 2014;9:e1000762014. doi:10.1371/journal.pone.0100076
6. Shi X, Sun M, Liu H, et al. Long non-coding RNAs: a new frontier in the study of human diseases. Cancer Lett. 2013;339:159–166. doi:10.1016/j.canlet.2013.06.013
7. Black DM, Rosen CJ. Postmenopausal osteoporosis. N Engl J Med. 2016;374:254–262. doi:10.1056/NEJMcp1513724
8. Gennari L, Rotatori S, Bianciardi S, et al. Treatment needs and current options for postmenopausal osteoporosis. Expert Opin Pharmacother. 2016;17:1141–1152. doi:10.1080/14656566.2016.1176147
9. Hassan MQ, Tye CE, Stein GS, et al. Non-coding RNAs: epigenetic regulators of bone development and homeostasis. Bone. 2015;81:746–756. doi:10.1016/j.bone.2015.05.026
10. Zhang L, Kang W, Lu X, et al. LncRNA CASC11 promoted gastric cancer cell proliferation, migration and invasion in vitro by regulating cell cycle pathway. Cell Cycle. 2018;17:1886–1900. doi:10.1080/15384101.2018.1502574
11. Han Y, Chen M, Wang A, et al. STAT3-induced upregulation of lncRNA CASC11 promotes the cell migration, invasion and epithelial-mesenchymal transition in hepatocellular carcinoma by epigenetically silencing PTEN and activating PI3K/AKT signaling pathway. Biochem Biophys Res Commun. 2018;32516.
12. Zhang Z, Zhou C, Chang Y, et al. Long non-coding RNA CASC11 interacts with hnRNP-K and activates the WNT/β-catenin pathway to promote growth and metastasis in colorectal cancer. Cancer Letters. 2016;376(1):62–73. doi:10.1016/j.canlet.2016.03.022
13. Zha L, He L, Liang Y, et al. TNF-α contributes to postmenopausal osteoporosis by synergistically promoting RANKL-induced osteoclast formation. Biomed Pharmacother. 2018;102:369–374. doi:10.1016/j.biopha.2018.03.080
14. Zheng SX, Vrindts Y, Lopez M, et al. Increase in cytokine production (IL-1β, IL-6, TNF-α but not IFN-γ, GM-CSF or LIF) by stimulated whole blood cells in postmenopausal osteoporosis. Maturitas. 1997;26:63–71.
15. Wu H, Liu J, Li W, et al. LncRNA-HOTAIR promotes TNF-α production in cardiomyocytes of LPS-induced sepsis mice by activating NF-κB pathway. Biochem Biophys Res Commun. 2016;471:240–246. doi:10.1016/j.bbrc.2016.01.117
16. Tella SH, Gallagher JC. Prevention and treatment of postmenopausal osteoporosis. J Steroid Biochem Mol Biol. 2014;142:155–170. doi:10.1016/j.jsbmb.2013.09.008
17. Maeda SS, Lazaretti-Castro M. An overview on the treatment of postmenopausal osteoporosis. Arq Bras Endocrinol Metabol. 2014;58:162–171.
18. Haro H, Crawford HC, Fingleton B, et al. Matrix metalloproteinase-7–dependent release of tumor necrosis factor-α in a model of herniated disc resorption. J Clin Invest. 2000;105(2):143–150. doi:10.1172/JCI7091
19. Fu Y, Zhang P, Nan H, et al. LncRNA CASC11 promotes TGF-β1, increases cancer cell stemness and predicts postoperative survival in small cell lung cancer. Gene. 2019;704:91–96. doi:10.1016/j.gene.2019.04.019
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.