Back to Journals » Cancer Management and Research » Volume 12
LINC00963 Functions as an Oncogene in Bladder Cancer by Regulating the miR-766-3p/MTA1 Axis
Authors Zhou N ![]() , Zhu X
, Zhu X ![]() , Man L
, Man L
Received 15 February 2020
Accepted for publication 19 April 2020
Published 12 May 2020 Volume 2020:12 Pages 3353—3361
DOI https://doi.org/10.2147/CMAR.S249979
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Ning Zhou, Xiaofei Zhu, Libo Man
Department of Urology, Beijing Jishuitan Hospital, Beijing 100035, People’s Republic of China
Correspondence: Libo Man Email [email protected]
Purpose: Long non-coding RNAs have been found to be involved in bladder cancer development. This article studied LINC00963 effects on bladder cancer progression to provide a novel treatment target.
Patients and Methods: Totally 56 bladder cancer patients participated in this research. Bladder cancer cells were transfected. Cell counting kit 8 assay and clone formation experiment were used for cell viability and colony formation detection. Cell migration and invasion were determined by Transwell experiment. LINC00963 distribution was explored by cytoplasmic and nuclear extract isolation and quantitative real-time polymerase chain reaction. Luciferase reporter experiment and RNA pulldown experiment were performed to detect the relationship between these two genes. The cancer genome atlas analysis was used for the detection of metastasis-associated protein 1 (MTA1) expression in bladder cancer.
Results: LINC00963 was seriously up-regulated in bladder cancer patients. High LINC00963 expression indicated high histological grade and low survival. LINC00963 was obviously up-regulated in bladder cancer cells. Knockdown of LINC00963 significantly reduced bladder cancer cells viability, colony formation, migration and invasion. Luciferase reporter experiment and RNA pulldown experiment revealed that LINC00963 promoted MTA1 expression via directly inhibiting miR-766-3p. MTA1 was up-regulated in bladder cancer patients. MTA1 up-regulation reversed the inhibitory effect of LINC00963 knockdown on bladder cancer cell viability, migration and invasion.
Conclusion: LINC00963 functions as an oncogene in bladder cancer by regulating the miR-766-3p/MTA1 axis.
Keywords: bladder cancer, LINC00963, miR-766-3p, MTA1, progression
Introduction
Bladder cancer is the 9th most common human malignant tumor worldwide and about 386,000 new bladder cancer cases are diagnosed worldwide each year.1 High metastasis, recurrence and mortality poses a serious threat not only to patients’ lives but also a heavy economic burden to the society.2 Significant progress has been made in the treatment of human malignancies in recent years, including bladder cancer. However, the prognosis of patients with bladder cancer is far from satisfactory.3 Therefore, the exploration of new therapeutic targets for bladder cancer is urgently needed for the improvement of prognosis.
Long non-coding RNAs (lncRNAs) are a class of RNA with a transcript length of 200nt-100kb that do not have the function of encoding proteins.4 It regulates gene expression at multiple levels, including at transcriptional, post-transcriptional and epigenetic levels.5 LncRNAs are involved in the physiological and pathological processes of the body, and its abnormal expression can result in a variety of diseases.6 Existing evidences have shown that one type of lncRNA can be abnormally expressed in different types of tumors, and multiple lncRNAs dysregulation can also exist in the same type of tumor.7–10 LINC00963 was a novel lncRNA and its function in tumors has attracted researchers’ attention until recent years. The activation of the PI3K/AKT signaling pathway is one of the important reasons for the development of various cancers. However, it has been found that the PI3K/AKT signaling pathway could be activated by LINC00963 in hepatocellular carcinoma.11 Available data indicated that LINC00963 could facilitate ovarian cancer proliferation, epithelial–mesenchymal transition and promote tumorigenesis and radiation resistance of breast cancer.12,13 In cutaneous squamous cell carcinoma and melanoma, LINC00963 was reported to function as a cancer-promoting factor and predicted poor prognosis.14,15 Significantly increased LINC00963 expression was occurred in esophageal squamous cell carcinoma, and knockdown of LINC00963 inhibited the growth of esophageal squamous cell carcinoma in vitro and in vivo.16 Zhou et al17 illustrated that seriously up-regulated LINC00963 in osteosarcoma was correlated with poor prognosis. However, the molecular mechanism of LINC00963 in regulating bladder cancer has not yet been elucidated.
Therefore, the function of LINC00963 in bladder cancer has been studied in detail in this article. More importantly, by investigating the effect of LINC00963 on the miR-766-3p/metastasis-associated protein 1 (MTA1) axis, the potential molecular mechanism of LINC00963 in regulating the development of bladder cancer was further verified. To our knowledge, this article firstly researched the regulation of bladder cancer development by LINC00963. This study will provide a novel potential target for the treatment of bladder cancer.
Patients and Methods
Patients and Tissues
Patients (n = 56) who were initially diagnosed with bladder cancer were enrolled in this research. All patients had no history of cancer-related diseases and no history of cancer-related treatments such as radiotherapy or chemotherapy. Tumor tissues and matched normal tissues (5 cm from bladder tumor margin) of all patients were collected during surgery by partial (39 samples) or radical (17 samples) cystectomy, followed by being immediately stored in liquid nitrogen. All patients volunteered to participate in this study and signed written informed consent. Correlation between LINC00963 expression and clinical features in BC tissues is presented in Table 1. This research has been approved by the ethics committee of Beijing Jishuitan Hospital and is in line with the Helsinki Declaration.
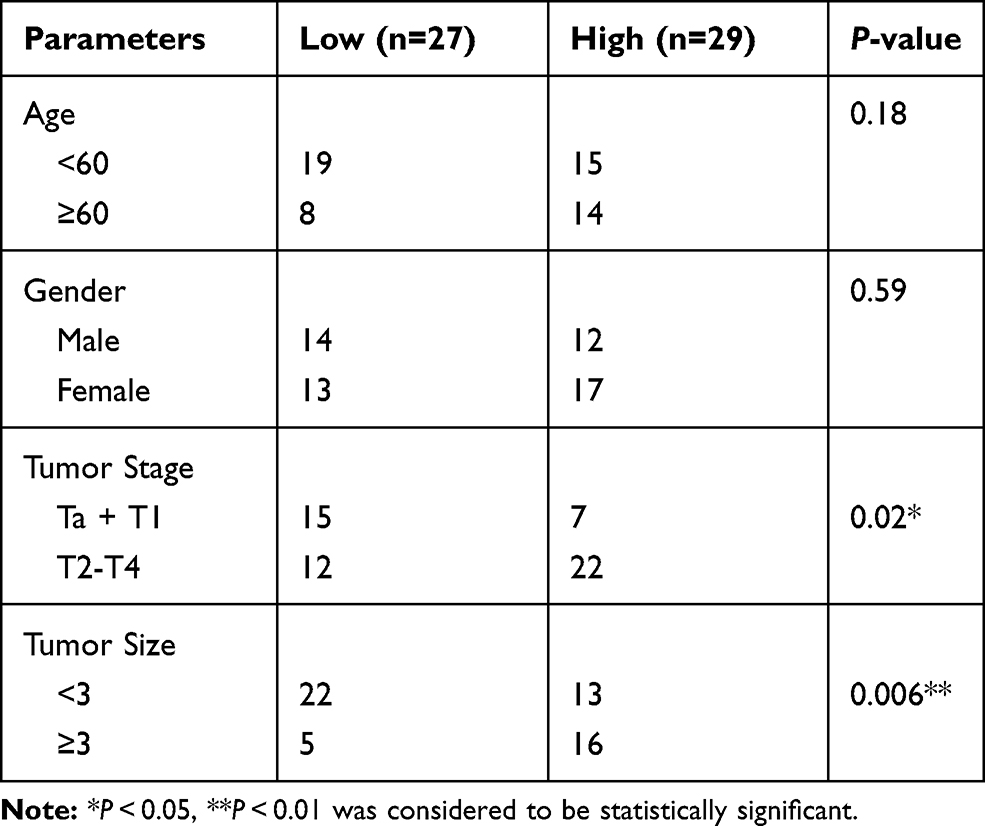 |
Table 1 Correlation Between LINC00963 Expression and Clinical Features in BC Tissues (N = 56) |
Cell Culture
Human bladder epithelial cell line (SV-HUC1) and bladder cancer cell lines (SW780, 5637, RT4, T24 and J82) were all provided by the Cell Bank of the Chinese Academy of Science (Shanghai, China), and maintained in dulbecco’s modified eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) in a humidified incubator at 37°C, 5% CO2.
Transfection
LINC00963 siRNA and relative negative control (NC), miR-766-3p mimics and miR-766-3p inhibitors were purchased from GenePharma (Shanghai, China). MTA1 overexpression sequence was inserted into the pcDNA3.1 vector (Realgene, Shanghai, China) strictly in line with the manufacturer’s instructions. T24 and J82 cells dispersed in serum-free DMEM were transfected by miR-766-3p mimics (miR-766-3p group), LINC00963 siRNA (si-LINC00963 group) and relative negative NC (si-NC group). Lipofectamine 2000 reagent used for transfection was obtained from Thermo Fisher Scientific (Waltham, MA, USA). DMEM containing 10% FBS was used to maintain these cells after 8 h of transfection and the transfection efficiency was researched by qRT-PCR.
Cell Counting Kit 8 (CCK-8) Assay
T24 and J82 cells dispersed in DMEM containing 10% FBS were plated into 96-well plates. After 24, 48 and 72 h incubation at 37°C, 5% CO2, 10 μL CCK-8 solutions was added into each well for 2 h incubation at 37°C. The optical density (OD) value was measured using a multi-well plate reader at 450 nm. Higher OD value meant higher cell viability.
Clone Formation Experiment
T24 and J82 cells dispersed in DMEM containing 10% FBS (500 cells/mL) were seeded and cultured for 12 days. Then, cells attached to the bottom of petri dishes were fixed for 10 min with 4% polyformaldehyde. Crystal violet (0.1%) was then added for 10 min staining at room temperature. Colonies of more than 50 cells were observed and counted under a microscope.
Transwell Experiment
T24 and J82 cells dispersed in serum-free DMEM were added into the upper chamber of 24-well transwell insert. DMEM containing 10% FBS was added into the lower chamber with a volume of 600 μL. After 24 h incubation at 37°C, 5% CO2, cells attached onto the upper membrane were wiped using cotton swab, and those migrated onto the lower membrane was fixed with 4% polyformaldehyde for 10 min. Crystal violet (0.1%) was then used for 10 min staining. The number of migrated cells was counted under a microscope in five random non-overlapping fields of view. The relative cell migration of calculated with the formula of (number of migrated cells/50,000).
The relative cell invasion was also detected using the method as described above. However, it was important to note that, a layer of Matrigel (100 μL) was pre-spread on the upper chamber.
Luciferase Reporter Experiment
The wild type (wt)-LINC00963 and mutant type (mut)-LINC00963 fragments containing the miR-766-3p binding site were synthesized by GenePharma (Shanghai, China) and were inserted into the luciferase reporter according to the instructions. T24 cells transfected by miR-766-3p mimics and NC were further subjected to cotransfection with wt-LINC00963 luciferase reporter and mut-LINC00963 luciferase reporter. After 48 h transfection at 37°C, 5% CO2, cells were harvested to detect the luciferase activity using Dual Luciferase Reporter Assay System. (Promega, USA)
In addition, the relationship between miR-766-3p and MTA1 was also verified by the same operations as described above. The wt-MTA1 and mut-MTA1 fragments containing the miR-766-3p binding site were synthesized by GenePharma (Shanghai, China).
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Total RNA in cells/tissues were extracted via using Trizol reagent (Solarbio, Beijing, China), and reverse transcribed by using PrimeScript RT reagent Kit (TaKaRa, Shiga, Japan). The qPCR was conducted using the iQ5 quantitative PCR system (Bio-Rad, Hercules, CA, USA) according to the following procedures: 95°C for 30 s, followed by 40 cycles at 95°C for 15 s, 60°C for 30 s, and 72°C for 30 s. The primer sequences were listed as follows: LINC00963, forward, 5′-GGTAAATCGAGGCCCAGAGAT-3′, reverse: 5′-ACGTGGATGACAGCGTGTGA-3′. miR-766-3p, forward, 5ʹ-ACA CTCCAGCTGGGACTCCAGCCCCACAG-3ʹ, reverse: 5ʹ-CTCAACTGGTGTCGTGGAGTCGGCAAT TCAGTTGAGGCTGAGGC-3ʹ. U6, forward: 5′-GTGCTCGCTTCGGCAGC-3′, reverse: 5′-CAGTGCAGGGTCCGAGGT-3′. MTA1, forward, 5′-CCAACATGTACAGGGTCGGA-3′, reverse: 5′-GGTCCGGCCTTATAGCAGAC-3′. GAPDH, forward, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′, reverse: 5′-AGCCTTCTCCATGGTGGTGAAGAC-3′. U6 was set as the internal control for LINC00963 and miR-766-3p. GAPDH was used as the internal control for MTA1. The relative expression of genes was calculated by 2−ΔΔCt method.
Statistical Analysis
All data were analyzed by SPSS 19.0 software (SPSS, Inc., Chicago, IL, USA) and presented in the form of mean ± standard deviation (SD). Kaplan-Meier analysis was used to analyze patients′ survival. Student′s t-test was used for the comparison between two groups. The comparison at least three groups was determined by one-way analysis of variance (ANOVA). All experiments involved in this research were repeated independently at least three times. P < 0.05 was set as the threshold.
Results
LINC00963 Was Seriously Up-Regulated in Bladder Cancer
LINC00963 expression in tumor/normal tissues of bladder cancer patients was validated via qRT-PCR. Obviously, the expression of LINC00963 was up-regulated in tumor tissues than that in normal tissues (P < 0.05) (Figure 1A). Analysis of histological grades showed that patients with high histological grade had much higher LINC00963 expression than those with low histological grade (P < 0.05) (Figure 1B). And LINC00963 expression was upregulated in invasive bladder cancer tissues (Figure 1C). Meanwhile, patients with high LINC00963 expression were obviously associated with lower 60 months survival (P < 0.05) (Figure 1D). According to in vitro studies, seriously higher LINC00963 expression was found in bladder cancer cell lines (SW780, 5637, RT4, T24 and J82) when relative to human bladder epithelial cell line (SV-HUC1) (P < 0.05) (Figure 1E).
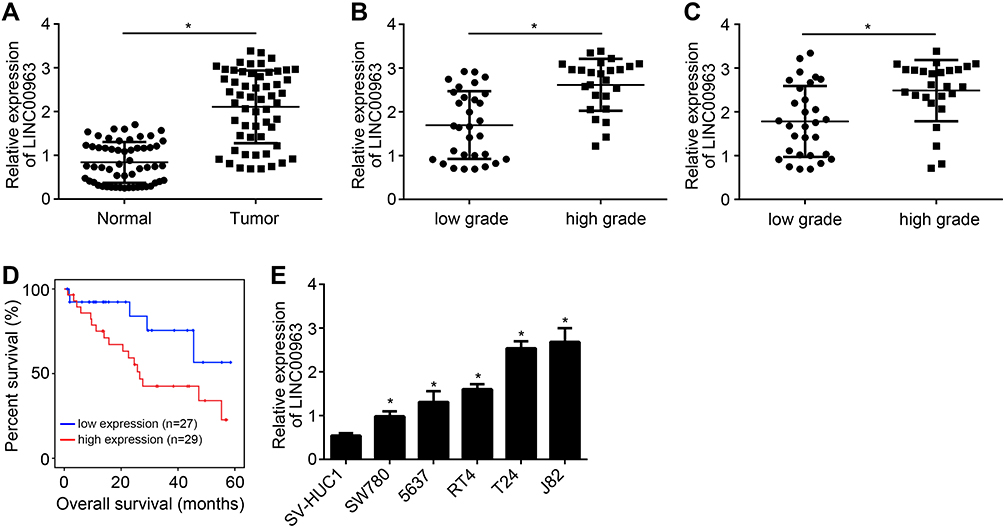 |
Figure 1 LINC00963 was seriously up-regulated in bladder cancer. (A) qRT-PCR showed obviously up-regulated expression of LINC00963 in bladder cancer tissues than that in normal tissues. (B) Patients with high histological grade had much higher LINC00963 expression than those with lower histological grade. (C) Relative expression of LINC00963 in non-muscle invasive bladder cancer and muscle-invasive bladder cancer tissues. (D) Patients with high LINC00963 expression were obviously associated with low survival rate. (E) Seriously higher LINC00963 expression was found in bladder cancer cell lines (SW780, 5637, RT4, T24 and J82) when relative to human bladder epithelial cell line (SV-HUC1). * P < 0.05. |
Knockdown of LINC00963 Inhibited Bladder Cancer Cells Viability, Colony Formation, Migration and Invasion
Transfection efficiency was determined by qRT-PCR. Relative to T24 and J82 cells of si-NC group, those of si-LINC00963 group had prominently lower relative LINC00963 expression (P < 0.05) (Figure 2A). Thus, T24 and J82 cells were successfully transfected. Using CCK-8 assay, T24 and J82 cells of si-LINC00963 group exhibited markedly lower cell viability at 72 h than those of si-NC group (P < 0.05) (Figure 2B and C). Clone formation experiment showed that, relative to si-NC group, T24 and J82 cells of si-LINC00963 group had much lower relative colony formation (P < 0.05) (Figure 2D). Analysis from transwell experiment showed seriously lower relative cell migration and invasion of si-LINC00963 group when compared with si-NC group (P < 0.05) (Figure 2E and F).
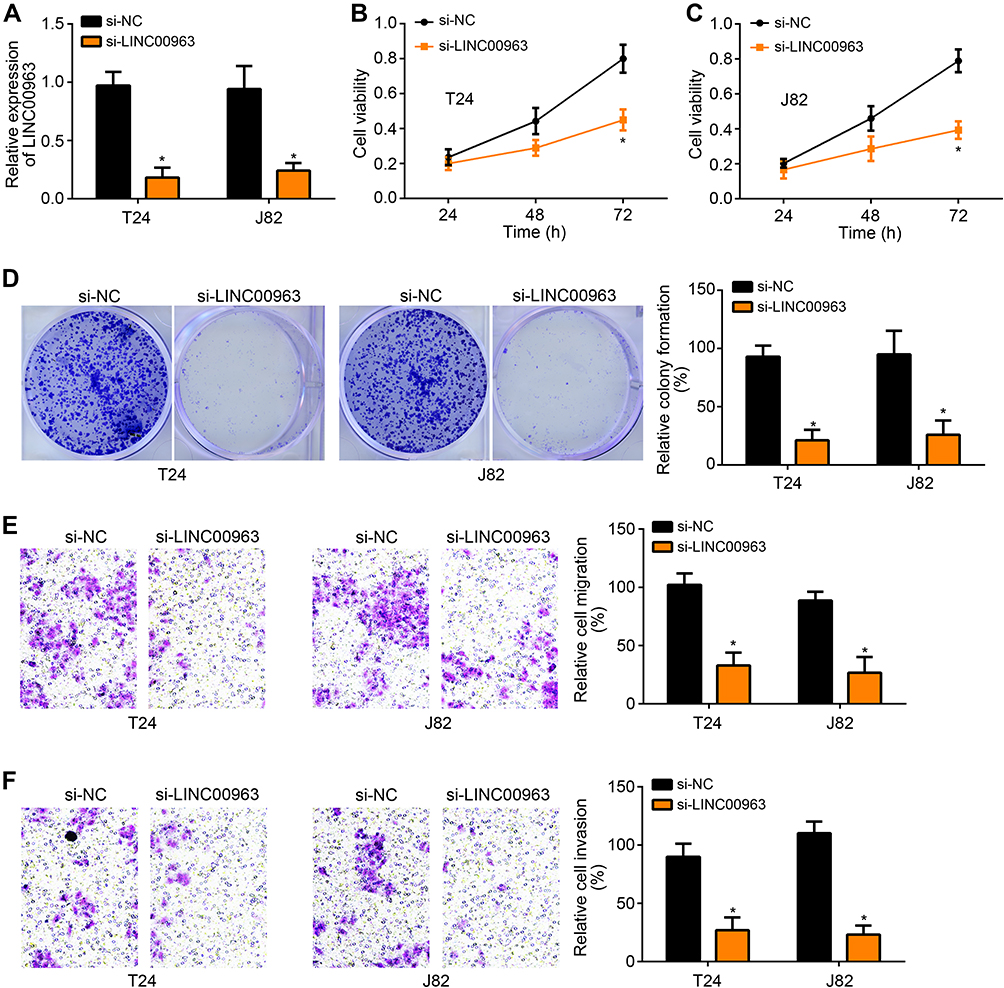 |
Figure 2 Knockdown of LINC00963 inhibited bladder cancer cells viability, colony formation, migration and invasion. (A) Relative to T24 and J82 cells of si-NC group, those of si-LINC00963 group had prominently lower relative LINC00963 expression. (B and C) Using CCK-8 assay, T24 and J82 cells of si-LINC00963 group exhibited markedly lower cell viability at 72 h than those of si-NC group. (D) Colony formation experiment showed that, relative to si-NC group, T24 and J82 cells of si-LINC00963 group had much lower relative colony formation. (E and F) Analysis from transwell experiment showed seriously lower relative cell migration and invasion of si-LINC00963 group when compared with si-NC group. * P < 0.05. |
LINC00963 Promoted MTA1 Expression via Directly Inhibiting miR-766-3p Expression
LINC00963 distribution in bladder cancer cells was detected. As shown in Figure 3A and B, LINC00963 was mainly distributed in the cytoplasm. miRDB and TargetScan predictions showed that LINC00963 possessed the binding site for miR-766-3p and miR-766-3 had the binding site for MTA1 (Figure 3C). Luciferase reporter experiment was further used to validate the relationship between two genes. The results exhibited that, compared with miR-NC group, miR-766-3p obviously reduced the luciferase activity of wt-LINC00963 luciferase reporter (P < 0.05), but had no obvious effect on luciferase activity of mut-LINC00963 luciferase reporter (Figure 3D). Similar results were also observed in effects of miR-766-3p on wt-MTA1 luciferase reporter and mut-MTA1 luciferase reporter (Figure 3E). In addition, RNA pulldown assay showed that, wt-biotin-miR-766-3p significantly elevated LINC00963 enrichment than biotin-NC and mut-biotin-miR-766-3p in T24 and J82 cells (P < 0.05) (Figure 3F). Relative to si-NC group, much higher miR-766-3p expression was found in T24 and J82 cells of si-LINC00963 group (P < 0.05) (Figure 3G). Moreover, prominently higher MTA1 expression was presented in T24 and J82 cells of si-LINC00963 group and miR-766-3p group when compared with si-NC group (P < 0.05). However, relative to si-LINC00963 group and miR-766-3p group, the expression of MTA1 in si-LINC00963 - miR-766-3p inhibitor group was significantly increased (P < 0.05) (Figure 3H).
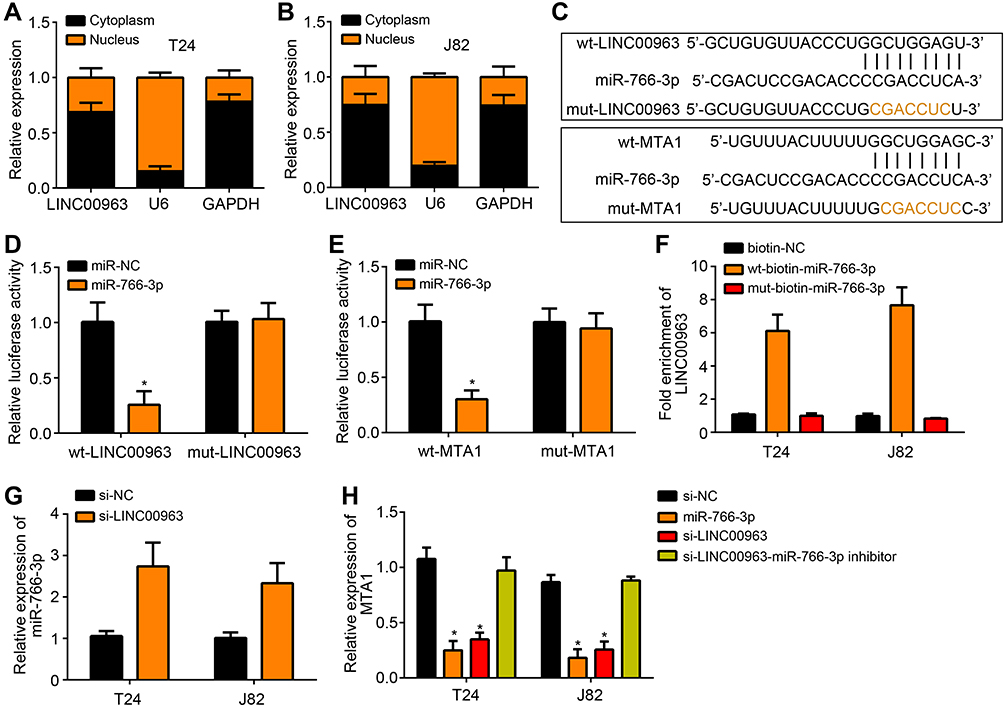 |
Figure 3 LINC00963 promoted MTA1 expression via directly inhibiting miR-766-3p expression. (A and B) LINC00963 distribution detection exhibited that LINC00963 was mainly distributed in the cytoplasm. (C) The binding site between LINC00963 and miR-766-3p or between miR-766-3p and MTA1. (D and E) Luciferase reporter experiment verified that miR-766-3p was a target gene of LINC00963 and MTA1 was a target gene of miR-766-3p. (F) RNA pulldown assay showed that wt-biotin-miR-766-3p had elevated LINC00963 enrichment. (G) Relative to si-NC group, much higher miR-766-3p expression was found in T24 and J82 cells of si-LINC00963 group. (H) LINC00963 promoted MTA1 expression via directly inhibiting miR-766-3p expression. * P < 0.05. |
MTA1 Up-Regulation Reversed The Inhibitory Effect of LINC00963 Knockdown on Bladder Cancer Progression
According to data from TCGA database, MTA1 expression in bladder cancer tissues was remarkably up-regulated than that in normal tissues (P < 0.05) (Figure 4A). Analysis from 56 bladder cancer patients also revealed significantly up-regulated MTA1 expression in tumor tissues than that in normal tissues (P < 0.05) (Figure 4B). Based on CCK-8 assay, it could be noticed that, compared with si-LINC00963 group, T24 and J82 cells of si-LINC00963 + oeMTA1 group had much higher cell viability at 72 h (P < 0.05) (Figure 4C and D). Prominently higher migration and invasion was also observed in si-LINC00963 + oeMTA1 group when relative to si-LINC00963 group (P < 0.05) (Figure 4E and F).
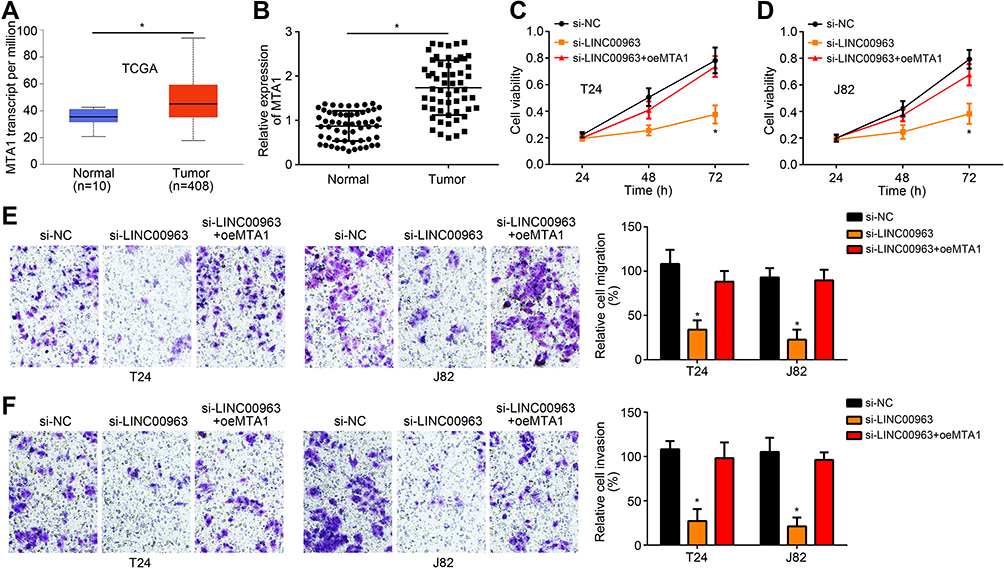 |
Figure 4 MTA1 up-regulation reversed the inhibitory effect of LINC00963 knockdown on bladder cancer progression. (A) According to data from TCGA database, MTA1 expression in bladder cancer tissues was remarkably up-regulated than that in normal tissues. (B) Analysis from 56 bladder cancer patients revealed significantly up-regulated MTA1 expression in tumor tissues than that in normal tissues. (C and D) CCK-8 assay showed that, compared with si-LINC00963 group, T24 and J82 cells of si-LINC00963 + oeMTA1 group had much higher cell viability at 72 h. (E and F) Prominently higher migration and invasion was observed in si-LINC00963 + oeMTA1 group when relative to si-LINC00963 group. * P < 0.05. |
Discussion
In previous researches, many different lncRNAs had been identified in bladder cancer, such as FOXD2-AS1, CAT266, CAT1297, CAT1647, LINC00857 and BANCR.18–21 One of the main ways for lncRNAs to participate in tumor development is to act as an endogenous competitive RNA of microRNAs (miRNAs) and indirectly regulates gene expression post-transcriptionally.5 LINC00963 has been found to be an oncogene in several human malignant tumors, including breast cancer, hepatocellular carcinoma, esophageal squamous cell carcinoma, etc.13,16,22 Similarly, we initially confirmed in this paper that LINC00963 was a cancer-promoting factor in bladder cancer and high LINC00963 expression was prominently associated with poor prognosis of patients. Previous data revealed that LINC00963, as a competing endogenous RNA, could regulate gene expression indirectly via targeting miRNAs in a direct manner, thereby promoting tumor progression.12,17 This research also demonstrated the role of LINC00963 as a competing endogenous RNA in bladder cancer, which facilitate the progression of bladder cancer via regulating miR-766-3p/MTA1 axis.
miRNAs are a kind of small non-coding RNA molecules with 18–25 nucleotides in length.23 miRNAs can inhibit or degrade mRNA translation via complementary base pairing, which leads to post-transcriptional silencing of target genes and thereby regulated cell differentiation, proliferation and apoptosis.24 miR-766-3p, as an important member of miRNAs family, has recently received much attention due to its vital roles in human malignant tumors. It has been found that miR-766-3p expression was aberrantly decreased in hepatocellular carcinoma and was significantly correlated with poor prognosis of patients. Wnt3a was found to be a target gene of miR-766-3p, and in hepatocellular carcinoma, miR-766-3p suppressed the tumor progression via inhibiting Wnt3a expression.25 Moreover, another research also demonstrated the role of miR-766-3p as a tumor suppressor in hepatocellular carcinoma, which could alleviate the development of hepatocellular carcinoma by directly inhibiting metastasis-associated protein 3 (MTA3).26 Chen et al27 declared that highly methylated miR-766-3p promoter was observed in renal cell carcinoma tissues and reduced miR-766-3p expression was seriously related to the poor prognosis of patients. Regarding the mechanism, the repression of miR-766-3p might facilitate renal cell carcinoma cells proliferation through targeting SF2. In this research, we noticed that miR-766-3p was a tumor suppressor in bladder cancer and its expression was directly suppressed by LINC00963. It was also worth noting that miR-766-3p could directly repress the expression of MTA1 in bladder cancer.
Interference or destruction of epigenetic homeostasis causes abnormal gene expression, which lead to the activation of proto-oncogenes, suppression of tumor suppressors and eventually tumorigenesis. This article indicated that LINC00963 facilitated bladder cancer progression by indirectly enhancing MTA1 expression via directly suppressing miR-766-3p. The rescue experiment results illustrated that MTA1 up-regulation reversed the inhibitory effect of LINC00963 knockdown on bladder cancer progression. Thus, MTA1 was a tumor-promoting gene for bladder cancer. MTA1 has been found to be up-regulated in several human malignancies and acted as a promoter of tumor proliferation and metastasis. Nan et al28 discovered that MTA1 facilitated the tumorigenesis and progression of esophageal squamous cell carcinoma might be through the activation of the MEK/ERK/p90RSK signaling pathway. In oral squamous carcinoma, MTA1 was found to induce epithelial–mesenchymal transition through activating the hedgehog signaling pathway.29 MTA1 expression was also up-regulated by lncRNA LUCAT1 in cervical cancer, which exacerbated tumor cells proliferation and invasion.30 However, data on MTA1 affecting bladder cancer development are currently scarcely reported. Our research confirmed the carcinogenic role of MTA1 in bladder cancer. Of course, more researches will be conducted in our future so as to confirm the influence of MTA1 on important cancer-related signaling pathways.
In conclusion, this research verified the function of LINC00963 in bladder cancer and proved the related mechanism. As a result, LINC00963 acted as a competing endogenous RNA, which functioned as an oncogene in bladder cancer by regulating the miR-766-3p/MTA1 axis. Thus, the current paper explored a novel target for the treatment of bladder cancer. However, there are some limitations in our study. The correlation between LINC00963 expression and prognosis in HG non-invasive bladder cancer or CIS alone or associate should be investigated in the future. And whether LINC00963 expression correlates with inflammation status or obesity also requires research. Whether LINC00963 has a role on BC in response to BCG also remains unknown. And, whether LINC00963 plays potential impact in the era of immunotherapy and NAC therapy pre cystectomy also needs to be determined.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zhan Y, Li Y, Guan B, et al. Long non-coding RNA HNF1A-AS1 promotes proliferation and suppresses apoptosis of bladder cancer cells through upregulating Bcl-2. Oncotarget. 2017;8(44):76656–76665. doi:10.18632/oncotarget.20795
2. Chen M, Li J, Zhuang C, et al. Increased lncRNA ABHD11-AS1 represses the malignant phenotypes of bladder cancer. Oncotarget. 2017;8(17):28176–28186. doi:10.18632/oncotarget.14945
3. Teng H, Zhou L, Wang L, et al. CLCA4 inhibits bladder cancer cell proliferation, migration, and invasion by suppressing the PI3K/AKT pathway. Oncotarget. 2017;8(54):93001–93013. doi:10.18632/oncotarget.21724
4. Chen S, Liang H, Yang H, et al. Long non-coding RNAs: the novel diagnostic biomarkers for leukemia. Environ Toxicol Pharmacol. 2017;55:81–86. doi:10.1016/j.etap.2017.08.014
5. Liu W, Rui M, Yuan Y. Post-transcriptional regulation of genes related to biological behaviors of gastric cancer by long noncoding RNAs and MicroRNAs. J Cancer. 2017;8(19):4141–4154. doi:10.7150/jca.22076
6. Liu CL, Deng ZY, Du ER, et al. Long non‑coding RNA BC168687 small interfering RNA reduces high glucose and high free fatty acid‑induced expression of P2X7 receptors in satellite glial cells. Mol Med Rep. 2018;17(4):5851–5859.
7. Arun G, Spector DL. MALAT1 long non-coding RNA and breast cancer. RNA Biol. 2019;16:6. doi:10.1080/15476286.2019.1592072
8. Chang J, Xu W, Du X, et al. MALAT1 silencing suppresses prostate cancer progression by upregulating miR-1 and downregulating KRAS. Onco Targets Ther. 2018;11:3461–3473. doi:10.2147/OTT.S164131
9. Zhuang R, Zhang X, Lu D, et al. lncRNA DRHC inhibits proliferation and invasion in hepatocellular carcinoma via c㎝yb‐regulated MEK/ERK signaling. Mol Carcinog. 2018;58:3.
10. Wei C, Wang H, Xu F, et al. LncRNA SOX21-AS1 is associated with progression of hepatocellular carcinoma and predicts prognosis through epigenetically silencing p21. Biomedicine Pharmacother. 2018;104:137–144.
11. Wu J-H, Tian XY, An QM, et al. LINC00963 promotes hepatocellular carcinoma progression by activating PI3K/AKT pathway. Eur Rev Med Pharmacol Sci.
12. Liu W, Yang YJ, An Q. LINC00963 promotes ovarian cancer proliferation, migration and EMT via the miR-378g/CHI3L1 axis. Cancer Manag Res. 2020;12:463–473. doi:10.2147/CMAR.S229083
13. Zhang N, Zeng X, Sun C, et al. LncRNA LINC00963 promotes tumorigenesis and radioresistance in breast cancer by sponging miR-324-3p and inducing ACK1 expression. Mol Ther Nucleic Acids. 2019;18:871–881. doi:10.1016/j.omtn.2019.09.033
14. Wang J, Li C, Xu L, et al. MiR-1193 was sponged by LINC00963 and inhibited cutaneous squamous cell carcinoma progression by targeting SOX4. Pathol Res Pract. 2019;215(10):152600. doi:10.1016/j.prp.2019.152600
15. Jiao H, Jiang S, Wang H, et al. Upregulation of LINC00963 facilitates melanoma progression through miR-608/NACC1 pathway and predicts poor prognosis. Biochem Biophys Res Commun. 2018;504(1):34–39. doi:10.1016/j.bbrc.2018.08.115
16. Liu HF, Zhen Q, Fan YK. LINC00963 predicts poor prognosis and promotes esophageal cancer cells invasion via targeting miR-214-5p/RAB14 axis. Eur Rev Med Pharmacol Sci. 2020;24(1):164–173. doi:10.26355/eurrev_202001_19907
17. Zhou Y, Yin L, Li H, et al. The LncRNA LINC00963 facilitates osteosarcoma proliferation and invasion by suppressing miR-204-3p/FN1 axis. Cancer Biol Ther. 2019;20(8):1141–1148.
18. An Q, Zhou L, Xu N. Long noncoding RNA FOXD2-AS1 accelerates the gemcitabine-resistance of bladder cancer by sponging miR-143. Biomed Pharmacother. 2018;103:415–420. doi:10.1016/j.biopha.2018.03.138
19. Dudek AM, Boer SJ, Boon N, et al. Identification of long non-coding RNAs that stimulate cell survival in bladder cancer. Oncotarget. 2017;8(21):34442–34452. doi:10.18632/oncotarget.16284
20. Dudek AM, van Kampen JG, Witjes JA, Kiemeney LA, Verhaegh GW. LINC00857 expression predicts and mediates the response to platinum-based chemotherapy in muscle-invasive bladder cancer. Cancer Med. 2018;7(7):3342–3350.
21. He A, Liu Y, Chen Z, et al. Over-expression of long noncoding RNA BANCR inhibits malignant phenotypes of human bladder cancer. J Exp Clin Cancer Res. 2016;35(1):125. doi:10.1186/s13046-016-0397-9
22. Wu J-H, Tian X-Y, An Q-M, et al. LINC00963 promotes hepatocellular carcinoma progression by activating PI3K/AKT pathway. Eur Rev Med Pharmacol Sci. 2018;22(6):1645–1652. doi:10.26355/eurrev_201803_14574
23. Chunqing Wang HH, Wang L, Jiang Y, Youjia X. Reduced miR-144-3p expression in serum and bone mediates osteoporosis pathogenesis by targeting RANK. Biochem Cell Biol. 2018;96:84.
24. Thomson JM. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006;20(16):2202–2207. doi:10.1101/gad.1444406
25. You Y, Que K, Zhou Y, et al. MicroRNA-766-3p Inhibits Tumour Progression by Targeting Wnt3a in Hepatocellular Carcinoma. Mol Cells. 2018;41(9):830–841. doi:10.14348/molcells.2018.0181
26. Liu L, Qi X, Gui Y, et al. Overexpression of circ_0021093 circular RNA forecasts an unfavorable prognosis and facilitates cell progression by targeting the miR-766-3p/MTA3 pathway in hepatocellular carcinoma. Gene. 2019;714:143992. doi:10.1016/j.gene.2019.143992
27. Chen C, Xue S, Zhang J, et al. DNA-methylation-mediated repression of miR-766-3p promotes cell proliferation via targeting SF2 expression in renal cell carcinoma. 2017;141(9):1867–1878.
28. Nan P, Wang T, Li C, et al. MTA1 promotes tumorigenesis and development of esophageal squamous cell carcinoma via activating the MEK/ERK/p90RSK signaling pathway. Carcinogenesis. 2019. doi:10.1093/carcin/bgz200
29. Song Q, Wang B, Liu M, et al. MTA1 promotes the invasion and migration of oral squamous carcinoma by inducing epithelial-mesenchymal transition via the hedgehog signaling pathway. Exp Cell Res. 2019;382(1):111450. doi:10.1016/j.yexcr.2019.05.031
30. Wang AH, Zhao J-M, Du J, et al. Long noncoding RNA LUCAT1 promotes cervical cancer cell proliferation and invasion by upregulating MTA1. Eur Rev Med Pharmacol Sci. 2019;23(16):6824–6829. doi:10.26355/eurrev_201908_18721
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.