Back to Journals » Journal of Inflammation Research » Volume 12
Linagliptin Inhibits Lipopolysaccharide-Induced Inflammation Concentration-Dependently And -Independently
Authors Sato N, Nakamura Y ![]() , Yamadera S, Inagaki M, Kenmotsu S, Saito H
, Yamadera S, Inagaki M, Kenmotsu S, Saito H ![]() , Oguchi T, Tsuji M, Chokki H, Ohsawa I, Gotoh H, Iwai S
, Oguchi T, Tsuji M, Chokki H, Ohsawa I, Gotoh H, Iwai S ![]() , Kiuchi Y
, Kiuchi Y ![]()
Received 9 July 2019
Accepted for publication 5 September 2019
Published 21 October 2019 Volume 2019:12 Pages 285—291
DOI https://doi.org/10.2147/JIR.S221761
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Naoki Sato,1,2 Yuya Nakamura,1,3 Shiho Yamadera,4 Masahiro Inagaki,1,5 Sachiyo Kenmotsu,5 Hiroshi Saito,4,6 Tatsunori Oguchi,1 Mayumi Tsuji,1 Hirokazu Chokki,1 Isao Ohsawa,3 Hiromichi Gotoh,3 Shinichi Iwai,6 Yuji Kiuchi1
1Department of Pharmacology, Showa University School of Medicine, Shinagawa-ku, Tokyo, Japan; 2Department of Research Center, Tanabe Pharmacy Inc., Tokyo, Japan; 3Department of Nephrology, Saiyu Soka Hospital, Soka City, Saitama-ken, Japan; 4Department of Hospital Pharmaceutics, Showa University School of Pharmacy, Shinagawa-ku, Tokyo, Japan; 5Fuculty of Arts and Sciences at Fujiyoshida, Showa University, Fujiyoshida City, Yamanashi-ken, Japan; 6Department of Healthcare and Regulatory Sciences, Showa University School of Pharmacy, Shinagawa-ku, Tokyo, Japan
Correspondence: Yuya Nakamura
Department of Pharmacology, Showa University School of Medicine, 1-5-8 Hatanodai, Shinagawa-ku, Tokyo, Japan
Tel +81-3-3784-8125
Fax +81-3-3787-4790
Email [email protected]
Purpose: Dipeptidyl peptidase-4 inhibitors, including linagliptin, prevent inflammation. However, the in vitro effects of linagliptin are unclear. Moreover, although linagliptin inhibits lipopolysaccharide (LPS)-induced inflammation, the anti-inflammatory effects of linagliptin in this context are not concentration-dependent. In the absence of LPS-binding protein (LBP), the pro-inflammatory effects of LPS involve pathways other than the Toll-like receptor (TLR) 4 pathway. Here, we aimed to determine the anti-inflammatory mechanisms of linagliptin in an experimental model in which LBP was added to the medium.
Methods: Human U937 monocytes were cultured at 1 × 106 cells/mL in Roswell Park Memorial Institute medium and differentiated into macrophages using phorbol myristate acetate. All processes were carried out in medium containing 10% fetal bovine serum (FBS). After 48 hrs of culture, we replaced the medium and pretreated the cells with 100, 250, 500, or 2500 nM linagliptin for 1 hr. We exchanged the medium again, and the cells were treated with 1 ng/mL LPS with or without 100, 250, 500, or 2500 nM linagliptin. Interleukin (IL)-6 and LBP in the supernatant, nuclear factor (NF)-κB/p65 in the nucleus, and reactive oxygen species (ROS) in the cells, as important markers of the mechanism of inflammation induction by LPS, were measured using enzyme-linked immunosorbent assay kits.
Results: Linagliptin significantly prevented LPS-stimulated IL-6 production and intranuclear NF-κB/p65 levels in a concentration-dependent manner. LPS-induced intracellular ROS levels were significantly decreased by linagliptin at all concentrations. LBP levels were markedly higher in FBS-containing medium than in medium without FBS. However, LBP levels did not change following administration of linagliptin and/or LPS.
Conclusion: Concentration-dependent and -independent inflammatory suppression was observed following linagliptin treatment in the context of LPS-induced pro-inflammatory responses. Thus, our findings suggested that linagliptin induced two different mechanisms to repress inflammation, i.e., TLR4-dependent and -independent mechanisms.
Keywords: linagliptin, lipopolysaccharide-binding protein, interleukin-6, intranuclear subunit of nuclear factor-κB/p65, reactive oxygen species
Plain Language Summary
Linagliptin is a dipeptidyl peptidase-4 inhibitor that suppresses inflammation. We previously reported that linagliptin prevents inflammation in human umbilical vein endothelial cells and human U937 monocytes in vitro. However, the mechanisms mediating the anti-inflammatory effects of linagliptin are unclear. In this study, we aimed to determine the anti-inflammatory mechanisms of linagliptin in an experimental model in which Lipopolysaccharide (LPS)-binding protein (LBP) was added to the medium. Our results showed that linagliptin significantly prevented LPS-stimulated Interleukin-6 production and intranuclear factor-κB/p65 levels in a concentration-dependent manner. In contrast, the effects of linagliptin on LPS-induced intracellular reactive oxygen species levels were not concentration-dependent. Moreover, LBP levels did not change following administration of linagliptin and/or LPS. Overall, these findings suggested that linagliptin induced two different mechanisms repressing inflammation, i.e., Toll-like receptor 4-dependent and -independent mechanisms. Our findings provide insights into the mechanisms of action of the inhibitor linagliptin and may establish a basis for future studies on the applications of this compound.
Introduction
Dipeptidyl peptidase-4 (DPP-4) inhibitors, which activate glucagon-like peptide-1, are antidiabetic drugs.1 Linagliptin is a unique DPP-4 inhibitor that is excreted in the bile. Despite decreased renal function, decreasing the dose of linagliptin is typically unnecessary.2–4 We previously reported that linagliptin prevents inflammation in human umbilical vein endothelial cells and human U937 monocytes in vitro. However, the inhibitory effects of inflammation by linagliptin are not concentration-dependent.5,6
Lipopolysaccharide (LPS) is a common molecule used to induce inflammation.5–7 LPS binds to the complex of Toll-like receptor (TLR) 4 and LPS-binding protein (LBP) and binds to receptors other than TLR4, even without LBP.8–11 Therefore, the pro-inflammatory mechanisms induced by LPS change in the presence or absence of LBP in vitro. Moreover, because LBP synthesized in the liver exists in the serum, these mechanisms may also change depending on the presence or absence of fetal bovine serum (FBS) in vitro. In our previous study, the pro-inflammatory effects of LPS were induced via a mechanism other than the binding to TLR4 in the absence of FBS; thus, linagliptin may have suppressed inflammation via a mechanism other than its binding to TLR4.7 However, no experimental models including FBS in the medium have been used to report the effects of linagliptin on inflammation induced by LPS through TLR4. It is important to examine anti-inflammatory effects of linagliptin from the perspective of LPS-induced TRL4-dependent and -independent. Nuclear factor (NF)-κB/p65 and intracellular reactive oxygen species (ROS) are important markers of the LPS-induced TRL4-dependent inflammation.12,13 Therefore, examination of the anti-inflammatory effects of linagliptin on inflammation induction from TRL4 using NF-κB/p65 and ROS as markers is important to elucidate the anti-inflammatory mechanisms of linagliptin.
In this study, we aimed to elucidate the anti-inflammatory mechanisms of linagliptin in an experimental model of LBP in human U937 monocytes. Furthermore, we compared the anti-inflammatory effects of linagliptin with those of sitagliptin (another DPP-4 inhibitor) and loxoprofen sodium salt dihydrate (Loxo; a representative anti-inflammatory drug).
Materials And Methods
Human U937 monocytes (EC85011440) were commercially purchased from the European Collection of Animal Cell Culture, UK. Moreover, we used linagliptin compounds, FBS, LPS, phorbol 12-myristate 13-acetate, and Roswell Park Memorial Institute medium (RPMI 1640), as described in our previous study.7 Sitagliptin was obtained from Santa Cruz Biotechnology, Inc. (Bergheimer, Heidelberg, Germany), and Loxo was obtained from Wako Pure Chemical Industries Ltd. (Osaka, Japan).
Briefly, cells were cultured in the presence of RPMI 1640 containing 10% FBS, as previously described.7 The passage number of U937 in this study ranged from two to five. Cells were evaluated in duplicate, and each experiment was repeated three times.
Drug Treatment
After 48 hrs of culture, we replaced the medium, and cells were then left untreated or were pretreated with 100, 250, 500, or 2500 nM linagliptin; 500 nM sitagliptin; or 570 μM Loxo for 1 hr. The medium was replaced again, and the cells were left untreated or treated with 1 ng/mL LPS only or in combination with 100, 250, 500, or 2500 nM linagliptin; 500 nM sitagliptin; or 570 μM Loxo. Inflammatory responses in U937 cells were sufficiently induced by 1 ng/mL LPS. The medium was collected and used to examine interleukin (IL)-6 and LBP levels at 5 hrs after drug treatment, NF-κB/p65 levels at 2 hrs after drug treatment, and ROS at 15 mins after drug treatment.
Measurement Of IL-6 In The Medium
IL-6 levels were measured using a Human IL-6 Quantikine enzyme-linked immunosorbent assay (ELISA) kit (Bio-Techne-R&D Systems Inc., Minneapolis, MN, USA) employing a sandwich-type immunoassay, as previously described.7 After pipetting the reference standard and samples into the wells of the microplate, which was precoated with monoclonal antibodies for human IL-6, the immobilized antibody bound to any IL-6 present. Any unbound substances were washed away, and enzyme-linked polyclonal antibodies were added. After washing, a substrate solution was added to detect the amount of IL-6 bound. After adding the stop solution, the color intensity was measured, and the absorbance was examined at 450 nm using a Spectra Max 340 pc (Molecular Devices Co., CA, USA). The results were reported as the amount of IL-6 protein in 1 mL supernatant.
Measurement Of Intranuclear NF-κB/p65
Two hours after drug treatment, we harvested the cells and extracted the nuclei using a Nuclear/Cytosol Fractionation Kit (BioVision Inc., Milpitas, CA, USA). We used an NF-κB p65 (Total) Human InstantOne ELISA Kit (Thermo Fisher Scientific, Waltham, MA, USA) with a sandwich ELISA protocol for quantitative determination of intranuclear p65 levels. Free p65 binding with the anti-p65 primary antibody-coated plate was determined using horseradish peroxidase-conjugated secondary p65 antibodies, and colorimetric detection of the amount of bound p65 was performed at 450 nm (Spectramax, CA, USA). Intranuclear NF-κB/p65 levels were expressed as the absorbance in transcription factor activity assays of NF-κB/p65 per 1 μg protein in nuclear extracts.
Measurement Of Intracellular ROS
We evaluated intracellular ROS formation using 5-(and-6)-chloromethyl-2ʹ,7ʹ-dichlorodihydrofluorescein diacetate (CM-H2DCF-DA; Molecular Probes, Eugene, OR, USA). After uptake in cells, intracellular esterases modify CM-H2DCF-DA to 2ʹ,7ʹ-dichlorofluorescein and intracellular ROS and then oxidize fluorescent 2ʹ,7ʹ-dichlorofluorescein. After dissolving CM-H2DCF-DA in dimethyl sulfoxide, the cells were incubated with 7 μM CM-H2DCF-DA for 15 mins and washed with heated phosphate-buffered isotonic sodium. The intensity of fluorescence was measured on a plate reader (Berthold Technologies; excitation wavelength: 488 nm, measuring wavelength: 525 nm). Intracellular ROS was expressed as the fluorescence intensity per 1 μg intracellular protein.
Measurement Of LBP In The Medium
For analysis of LBP levels in the medium, we harvested cells and quantified LBP levels in the extracted cell medium. We measured LBP levels using a solid-phase ELISA kit (HK503; Hycult Biotech Inc.). We incubated samples and standards in microtiter wells coated with anti-LBP antibodies. After binding of the captured LBP with a biotinylated tracer and binding the biotinylated tracer with a streptavidin–peroxidase conjugate, the reaction of the streptavidin–peroxidase conjugate and the substrate tetramethylbenzidine was stopped by the addition of oxalic acid. The absorbance was detected at 450 nm (Spectramax).
Statistical Analysis
JMP (version 10) statistical software (SAS Institute, Cary, NC, USA) was used. All data were reported as means ± standard errors. Results for various treatments were compared with those for the untreated control and LPS-only groups using one-way analysis of variance and Dunnett’s tests. Differences with P values of less than 0.05 were considered significant.
Results
Effects Of Linagliptin On LPS-Induced IL-6 Levels
IL-6 levels did not differ compared with those in the untreated control (6.0 ± 1.0 pg/mL) after treatment with various concentrations of linagliptin. In addition, following treatment with LPS, IL-6 levels were significantly increased (56.8 ± 5.1 pg/mL) compared with that in the control (P < 0.0001). Importantly, linagliptin suppressed LPS-induced IL-6 levels in a concentration-dependent manner (250 nM: 39.8 ± 9.0 pg/mL, P < 0.05; 500 nM: 30.6 ± 4.8 pg/mL, P < 0.01; 2500 nM: 12.2 ± 2.7 pg/mL; Figure 1).
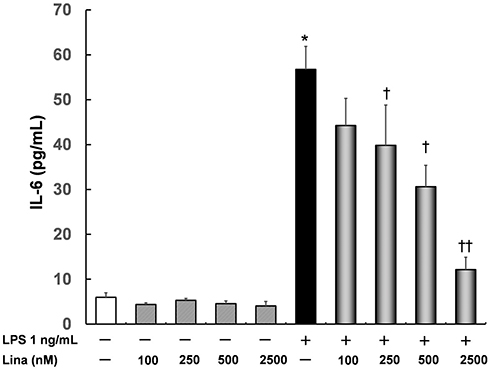 |
Figure 1 Effects of linagliptin on interleukin (IL)-6 induced by lipopolysaccharide (LPS). Human U937 monocytes were treated with LPS and/or linagliptin, and IL-6 levels in the supernatant were evaluated. *p < 0.0001 versus the control; †p < 0.05 versus LPS; ††p < 0.01 versus LPS. Abbreviation: Lina, linagliptin. |
Effects Of Linagliptin, Sitagliptin, And Loxo On LPS-Induced IL-6 Levels
The maximum blood concentrations of linagliptin and Loxo in the protein binding state (500 nM and 570 μM, respectively) were chosen for these experiments. The concentration of sitagliptin was adjusted to be the same as that of linagliptin.
Linagliptin, sitagliptin, and Loxo prevented LPS-induced IL-6 upregulation, and IL-6 levels were significantly decreased by linagliptin (30.6 ± 4.8 pg/mL; P < 0.05) and Loxo (2.0 ± 0.5 pg/mL; P < 0.01; Figure 2).
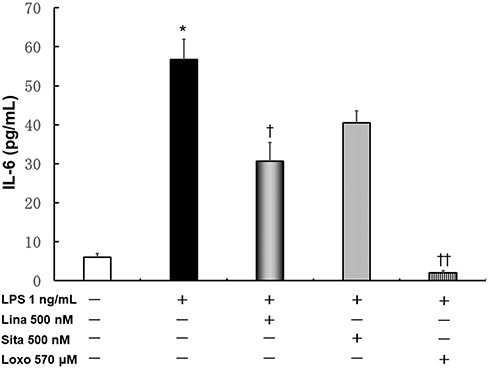 |
Figure 2 Effects of linagliptin, sitagliptin, and loxoprofen sodium salt dehydrate (Loxo) treatment on LPS-induced IL-6 production. Human U937 monocytes were treated with LPS and/or linagliptin, sitagliptin, or Loxo, and IL-6 levels in the supernatant were evaluated. *p < 0.0001 versus the control; †p < 0.05 versus LPS; ††p < 0.01 versus LPS. Abbreviations: Lina, linagliptin; Sita, sitagliptin; Loxo, loxoprofen sodium salt dehydrate. |
Effects Of Linagliptin On LPS-Induced Intranuclear NF-κB/p65 Levels
Intranuclear NF-κB/p65 levels were increased by treatment with LPS alone (175.5 ± 13.2 optical density/μg protein) compared with those in control cells (63.8 ± 9.7 optical density/μg protein; P < 0.001). Linagliptin significantly inhibited the LPS-stimulated increase in intranuclear NF-κB/p65 levels in a concentration-dependent manner (100 nM: 122.5 ± 11.0 optical density/μg protein, P < 0.01; 250 nM: 96.3 ± 10.8 optical density/μg protein, P < 0.01; 500 nM: 89.3 ± 21.4 optical density/μg protein, P < 0.01; 2500 nM: 49.9 ± 9.3 optical density/μg protein, P < 0.01; Figure 3).
 |
Figure 3 Effects of linagliptin on LPS-induced intranuclear nuclear factor-κB (NF-κB)/p65 levels. Human U937 monocytes were treated with LPS and/or linagliptin, and intranuclear NF-κB/p65 levels were determined. *p < 0.001 versus the control; †p < 0.01 versus LPS. Abbreviation: Lina, linagliptin. |
Effects Of Linagliptin On Intracellular ROS Levels Induced By LPS
Intracellular ROS levels were significantly increased by LPS alone (1.5 ± 0.2 fluorescence intensity/μg protein) compared with those in control cells (2.6 ± 0.3 fluorescence intensity/μg protein). At all concentrations, linagliptin inhibited LPS-induced intracellular ROS levels in a concentration-independent manner (100 nM: 1.5 ± 1.2 fluorescence intensity/μg protein, P < 0.05; 250 nM: 1.3 ± 0.2 fluorescence intensity/μg protein, P < 0.05; 500 nM: 1.4 ± 0.1 fluorescence intensity/μg protein, P < 0.05; 2500 nM: 1.5 ± 0.2 fluorescence intensity/μg protein, P < 0.05; Figure 4).
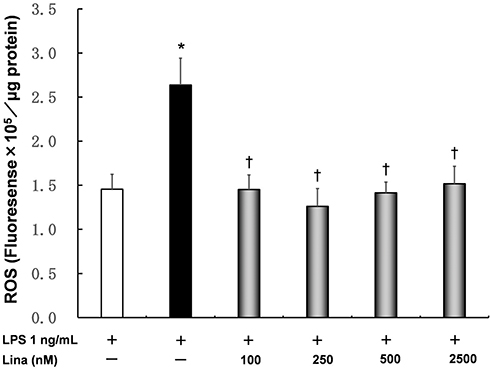 |
Figure 4 Effects of linagliptin on intracellular reactive oxygen species (ROS) induced by LPS. Human U937 monocytes were treated with LPS and/or linagliptin, and intracellular ROS levels were determined. *p < 0.05 versus the control; †p < 0.05 versus LPS. Abbreviation: Lina, linagliptin. |
Effects Of Linagliptin On LBP
LBP levels were higher in medium containing FBS (73.5 ± 0.8 ng/mL) than in medium without FBS (0 ng/mL). LBP levels did not change, regardless of administration of linagliptin and/or LPS (Figure 5).
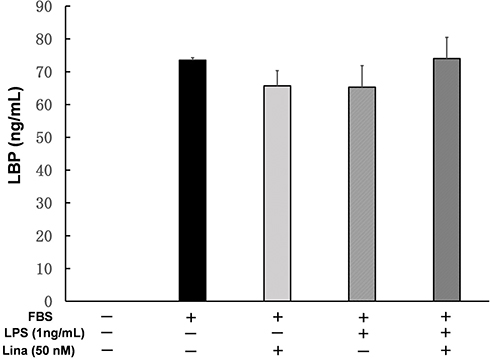 |
Figure 5 Effects of linagliptin on LPS-binding protein (LBP). Human U937 monocytes were treated with fetal bovine serum (FBS), LPS, and/or linagliptin, and LBP levels in medium were measured. Abbreviation: Lina, linagliptin. |
Discussion
In this study, induction of IL-6 and intranuclear p65 by LPS was significantly suppressed by linagliptin in a concentration-dependent manner in the presence of FBS in human U937 monocytes. Several concentrations treated by linagliptin had the first-order correlation against LPS mono on IL-6 levels in order of concentration of linagliptin (correlation coefficient; 0.6476, P<0.0001). In contrast, linagliptin significantly inhibited LPS-induced intracellular ROS levels in a concentration-independent manner, similar to the results of IL-6 or tumor necrosis factor-α in the absence of FBS in our previous research.5,7 These results demonstrating the presence of concentration-dependent and -independent pathways suggest that linagliptin may induce two different anti-inflammatory mechanisms.
During the induction of inflammation by LPS, the mechanism induced by binding of LPS and TLR4 in the presence of LBP has been extensively studied. Additionally, the mechanism through which LPS binds to receptors other than TLR4 has also been reported.8–11 Therefore, the mechanisms through which LPS induces inflammation may change according to the presence or absence of LBP, which may affect the anti-inflammatory effects of linagliptin. In this study, we confirmed the effects of LBP alone in the presence of FBS. Our findings suggested that the concentration dependence of the anti-inflammatory effects is related to differences in the mechanisms induced in the presence or absence of LBP. Indeed, the results of intracellular ROS sampled at an earlier time after administration of LPS and linagliptin did not show concentration dependence; moreover, these findings were observed rapidly and did not require 2–5 hrs, as is required for induction of the LPS/TLR4-dependent inflammatory mechanism (data not shown).
The maximum linagliptin concentrations in the serum of healthy and diabetic patients with no renal dysfunction were 7.32 and 16.7 nM, respectively, after single application of 5 mg linagliptin (Boehringer Ingelheim Pharmaceuticals, Inc. Tradjenta [linagliptin] tablets).14 Moreover, repeated application of 5 mg linagliptin in diabetic patients with a creatinine clearance of less than or equal to 30 mL/min increased the concentration to 22.6 nM.14 Therefore, the maximum linagliptin concentration in the serum in patients is estimated to be 5–50 nM. The protein binding rate of linagliptin is high, for instance up to 80%, even in the presence of a high maximum blood concentration.14 Therefore, we used 100–2500 nM linagliptin to measure changes in IL-6 and intranuclear NF-κB/p65 levels.
In our previous study, pretreatment of human umbilical endothelial cells with sitagliptin and Loxo inhibited LPS-induced inflammation.5,15 Additionally, in this study, sitagliptin suppressed inflammation in monocytes (although the difference was not significant). These experimental results indicated that sitagliptin had anti-inflammatory effects, even if the cell line was changed. Because both linagliptin and sitagliptin prevented inflammation, DPP-4 inhibitors may also exhibit anti-inflammatory effects because these inhibitors are in the same class of compounds. Notably, the anti-inflammatory effects of linagliptin were weaker than those of nonsteroidal anti-inflammatory drugs, a representative class of anti-inflammatory drugs.
As a limitation of this study, we could not examine whether increased IL-6 levels are due to production or secretion or whether increased intranuclear NF-κB/p65 levels are due to expression or nuclear translocation. Hereafter, further studies investigating quantitative polymerase chain reaction of Il-6 expression, intranuclear NFkB/p65 expression, and time-dependent ROS formation are necessary to elucidate the anti-inflammatory effect of linagliptin on TLR4-mediated or -unmediated inflammation. As the other limitation, we cannot investigate the anti-inflammatory mechanism of linagluptin by decreased prostaglandin E2 via suppression of p38 mitogen-activated protein kinase phosphorylation. In our previous studies, linagliptin inhibited lipopolysaccharide-stimulated p38 mitogen-activated protein kinase phosphorylation in human umbilical vein endothelial cells and decreased prostaglandin E2 in hemodialysis patients.5,16 Therefore, linagliptin may have anti-inflammatory mechanism by decreased prostaglandin E2 via suppression of p38 mitogen-activated protein kinase phosphorylation. Moreover, it is important to examine by distinguishing between membrane-bound and soluble. Soluble DPP4 is more important in immune regulation than membrane-bound DPP4 and the soluble DPP4 exert reducing inflammatory manner.17 In this in vitro study, culture medium are changed twice before linagliptin treatment. Therefore in this study, soluble DPP4 may be less involved compared to in vivo study.
Based on our findings in this study, we propose that linagliptin may induce two different anti-inflammatory mechanisms, although the specific molecular mechanisms of these effects are still not clear. In our previous study, we found that linagliptin inhibited inflammation via p38 mitogen-activated protein-kinase-dependent mechanisms stimulated by LPS.5 In this study, we investigated the anti-inflammatory mechanisms further upstream of linagliptin. The detailed mechanisms could not be fully elucidated. LPS binds to the complex of TLR4 and LBP. Our results here showed that LBP levels did not change, regardless of administration of linagliptin and/or LPS. Therefore, in the anti-inflammatory mechanism of linagliptin, it is unlikely that linagliptin inhibits the formation of the LBP, TLR4, and CD14 complex.
Conclusion
Linagliptin inhibited LPS-induced inflammation, even in an experimental model including LBP in human U937 monocytes. Two mechanisms of inflammatory suppression were observed, i.e., concentration-dependent and -independent mechanisms. Overall, our findings provided important insights into the anti-inflammatory mechanisms of linagliptin.
Abbreviations
LPS, lipopolysaccharide; LBP, LPS-binding protein; TLR, Toll-like receptor; FBS, fetal bovine serum; IL, Interleukin; NF, nuclear factor; ROS, reactive oxygen species; DPP-4, Dipeptidyl peptidase-4; Loxo, loxoprofen sodium salt dihydrate.
Author Contributions
Naoki Sato, Yuya Nakamura, and Masahiro Inagaki devised the study concept and design. Shiho Yamadera, Sachiyo Kenmotsu, and Hiroshi Saito performed the experiments and collected the data. Naoki Sato, Yuya Nakamura, Masahiro Inagaki, and Shiho Yamadera analyzed the data. Yuya Nakamura interpreted the data. Yuya Nakamura searched the literature. Naoki Sato, Yuya Nakamura, and Shiho Yamadera drafted the manuscript. Naoki Sato, Yuya Nakamura, Shiho Yamadera, Masahiro Inagaki, Sachiyo Kenmotsu, Hiroshi Saito, Tatsunori Oguchi, Mayumi Tsuji, Hirokazu Chokki, Isao Ohsawa, Hiromichi Gotoh, Shinichi Iwai, and Yuji Kiuchi contributed to substantial revisions to the manuscript. Finally, all authors made substantial contributions to conception and design, acquisition of data, and analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Drucker DJ. Therapeutic potential of dipeptidyl peptidase IV inhibitors for the treatment of type 2 diabetes. Expert Opin Investig Drugs. 2003;12:87–100. doi:10.1517/13543784.12.1.87
2. Blech S, Ludwig-Schwellinger E, Gräfe-Mody EU, Withopf B, Wagner K. The metabolism and disposition of the oral dipeptidyl peptidase-4 inhibitor, linagliptin, in humans. Drug Metab Dispos. 2010;38:667–678. doi:10.1124/dmd.109.031476
3. Hüttner S, Graefe-Mody EU, Withopf B, Ring A, Dugi KA. Safety, tolerability, pharmacokinetics, and pharmacodynamics of single oral doses of BI 1356, an inhibitor of dipeptidyl peptidase 4, in healthy male volunteers. J Clin Pharmacol. 2008;48:1171–1178. doi:10.1177/0091270008323753
4. Gallwitz B. Management of patients with type 2 diabetes and mild/moderate renal impairment: profile of linagliptin. Ther Clin Risk Manag. 2015;11:799–805. doi:10.2147/TCRM.S67076
5. Nakamura Y, Hasegawa H, Tsuji M, et al. Linagliptin inhibits lipopolysaccharide-stimulated interleukin-6 production, intranuclear p65 expression, and p38 mitogen-activated protein kinase phosphorylation in human umbilical vein endothelial cells. Ren Replace The. 2016;2:17. doi:10.1186/s41100-016-0030-6
6. Nakamura Y, Inagaki M, Tsuji M, et al. Linagliptin has wide-ranging anti-inflammatory points of action in human umbilical vein endothelial cells. Jpn Clin Med. 2016;7:27–32. doi:10.4137/JCM.S39317
7. Yamadera S, Nakamura Y, Inagaki M, et al. Linagliptin inhibits lipopolysaccharide induced inflammation in human U937 monocytes. Inflamm Regen. 2018;38:13. doi:10.1186/s41232-018-0071-z
8. Wright SD, Ramos RA, Tobias PS, et al. CD14, a receptor for complexes of lipopolysaccharide (LPS) and LPS binding protein. Science. 1990;249:1431–1433. doi:10.1126/science.2271062
9. Nishijima M, Hara-Kuge S, Takasuka N, et al. Identification of a biochemical lesion, and characteristic response to lipopolysaccharide (LPS) of a cultured macrophage-like cell mutant with defective LPS-binding. J Biochem. 1994;116:1082–1087. doi:10.1093/oxfordjournals.jbchem.a124631
10. Weinstein SL, June CH, DeFranco AL. Lipopolysaccharide-induced protein tyrosine phosphorylation in human macrophages is mediated by CD14. J Immunol. 1993;151:3829–3838.
11. Yamamoto Y, Harashima A, Saito H, et al. Septic shock is associated with receptor for advanced glycation end products (RAGE) ligation of LPS. J Immunol. 2011;186:3248–3257. doi:10.4049/jimmunol.1002253
12. Gonzalez-Rey E, Chorny A, Delgado M. Regulation of immune tolerance by anti-inflammatory neuropeptides. Nat Rev Immunol. 2007;7:52–63. doi:10.1038/nri1984
13. Morgan MJ, Liu ZG. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011;21:103–115. doi:10.1038/cr.2010.178
14. Boehringer Ingelheim Pharmaceuticals, Inc. Tradjenta (linagliptin) tablets. Available from: http://bidocs.boehringer-ingelheim.com/BIWebAccess/ViewServlet.ser?docBase=renetnt&folderPath=/Prescribing+Information/PIs/Tradjenta/Tradjenta.pdf.
15. Hasegawa H, Nakamura Y, Tsuji M, et al. Sitagliptin inhibits the lipopolysaccharide-induced inflammation. J Pharm Drug Deliv Res. 2016;5:2. doi:10.4172/2325-9604.1000148
16. Nakamura Y, Tsuji M, Hasegawa H, et al. Anti-inflammatory effects of linagliptin in hemodialysis patients with diabetes. Hemodial Int. 2014;18:433–442. doi:10.1111/hdi.12127
17. Varin EM, Mulvihill EE, Beaudry JL, et al. Circulating levels of soluble dipeptidyl peptidase-4 are dissociated from inflammation and induced by enzymatic DPP4 inhibition. Cell Metab. 2019;29:320–334.e5. doi:10.1016/j.cmet.2018.10.001
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.