Back to Journals » Journal of Inflammation Research » Volume 17
Limosilactobacillus Fermentun ZS09 Can Improve Antibiotic-Induced Motor Dysfunction in Mice by Regulating the Brain–Gut Functions
Authors Yang Y, Zhao Y, Lei H, Tan X
Received 19 August 2024
Accepted for publication 19 November 2024
Published 30 November 2024 Volume 2024:17 Pages 10093—10106
DOI https://doi.org/10.2147/JIR.S486018
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Yang Yang,1 Yuanji Zhao,2 Huan Lei,1 Xiaohong Tan3
1College of Physical Education, Chengdu Sport University, Chengdu, Sichuan, 610041, People’s Republic of China; 2School of Physical Education, Wuhan Sports University, Wuhan, Hubei, 430079, People’s Republic of China; 3Collaborative Innovation Center for Child Nutrition and Health Development, Chongqing University of Education, Chongqing, 400067, People’s Republic of China
Correspondence: Yuanji Zhao, School of Physical Education, Wuhan Sports University, Wuhan, Hubei, 430079, People’s Republic of China, Email [email protected]
Purpose: When consumed in appropriate quantities, probiotics, which are live microorganisms, are good for health. In this study, a mouse model of antibiotic-induced dyskinesia was established using a sterile mixed antibiotic solution to investigate the preventive impact of Limosilactobacillus fermentum ZS09 (LFZS09) on this condition in mice.
Methods: Following modeling, alterations in the serum and brain tissue of mice were assessed for motor measures such as running and swimming, malondialdehyde (MDA), superoxide dismutase, reduced glutathione, interleukin (IL-6, IL-10), and tumor necrosis factor (TNF)-α. The mouse cecum was used to evaluate the relative mRNA expression levels of the intestinal barrier genes, namely occludin-1, zona occludens-1 (ZO-1), and claudin-1. The relative mRNA expression levels of cAMP-response element binding protein (CREB), extracellular signal-regulated kinase (ERK), and brain-derived neurotrophic factor (BDNF) genes in the mouse brain tissue were also evaluated.
Results: Compared with the model group, LFZS09 considerably increased the swimming and running duration of mice, significantly decreased the levels of the inflammatory factors TNF-α and IL-6 and increased SOD expression in the mouse brain, and decreased MDA accumulation in the mouse brain and serum. Furthermore, LFZS09 upregulated occludin-1 gene expression in the cecal tissue to maintain the intestinal barrier, which in turn maintained the normal physiological function of the body. LFZS09 also enhanced the effect of BDNF and increased the expression of BDNF metabolic pathway-related genes, namely CREB, ERK1/2, and BDNF, in the mouse brain tissue. LFZS09 increased the number of Lactobacillus and Bifidobacterium in the gut of mice with motor dysregulation, and decreased the number of Enterococcus and Clostridium perfringens.
Conclusion: The findings indicate that LFZS09 regulates antibiotic-induced motor impairment in mice, thereby offering a theoretical foundation for future studies and probiotic or parabiotics production aimed at augmenting motor function.
Keywords: Limosilactobacillus fermentum, gut function, fatigue, antibiotic, mouse
Introduction
While antibiotics kill pathogenic bacteria, they also affect the normal bacterial community in the gut. This effect may lead to dysbiosis of the flora, with a decrease in the number of some beneficial bacteria and an increase in the proportion of resistant bacteria or other pathogens. Such dysregulation may affect the normal functions of the intestine, including digestive absorption and metabolic processes.1 There is a close interaction between gut microbiota and the central nervous system, which is called gut-brain axis. Studies have shown that changes in gut microbiota can affect the function of the brain through a variety of mechanisms, such as metabolites and inflammatory responses. These changes may lead to impairments in motor control or affect psychological states, which in turn indirectly affect motor ability.2 Gut microbiota plays an important role in the digestion and absorption of nutrients. The imbalance of bacterial flora caused by antibiotics may lead to the reduction of the synthesis and absorption of some nutrients (such as vitamin B group, short-chain fatty acids, etc)., resulting in insufficient energy obtained by the body. These nutrients and energy are essential for neurological function and muscle health, and their deficiency may lead to movement disorders.3
Following exercise, the body undergoes several changes in energy and substance metabolism along with a constant interchange of substances with the external world. The constant synthesis of ATP guarantees body movement, and stimulation or inhibition of ATP production encourages or impedes movement.4 The body undergoes certain metabolic changes during exercise, including the release of stored carbohydrates, a decline in blood glucose levels, an increase in lactic acid concentrations in the blood and muscle, a drop in pH, and lactic acid accumulation in peripheral skeletal muscle cells, which is then released into the bloodstream. The weariness that an individual develops after exercise is closely associated with these alterations.5 During high-level cognitive functions of the brain, neurons chiefly use lactate as an energy source (instead of glucose) to generate ATP and endure synaptic activity. The brain may use lactate to compensate for the energy required to maintain neuronal activity during high-intensity exercise, as evidenced by the decline in brain glucose absorption with an increase in exercise intensity.6 The brain and peripheral nerves develop more inflammation during endurance training. Thus, lowering the amount of exercise-induced inflammation in the liver, muscles, blood, and peripheral nerves may postpone exercise exhaustion. The brain-derived neurotrophic factor (BDNF) belongs to the neurotrophin family, which is mostly expressed in brain and nerve tissues. BNDF is a component crucial for both nerve cell survival and differentiation.7 Once BDNF binds to TrκB, it activates the downstream serine/threonine kinase pathway, followed by the activation of ERK kinase and ERK, eventually leading to CREB phosphorylation, enhancing learning and memory function, and contributing to an improvement in the motor ability.8
Mice reared for high levels of running and a group of mice reared generally. They were both treated with antibiotics for 10 days, and fecal samples showed that antibiotics eliminated nearly all gut bacteria in the mice. None of the mice showed any outward signs of illness or adverse effects during 10 days of antibiotic treatment. However, the wheel-running ability of the exercise mice decreased by 21%, and these mice did not recover their previous performance ability even 12 days after antibiotic withdrawal. The results showed that antibiotics had a serious negative impact on the exercise ability of the mice, and the specific mechanism was closely related to the destruction of intestinal flora by antibiotics. Therefore, in this study, antibiotics were also used to intervene the gut microbiota of mice to establish dyskinesia.9 Limosilactobacillus fermentum has numerous physiological functions such as lowering cholesterol levels, enhancing nutrition synthesis and metabolism, and controlling the gut immune function.10 The herdsmen of Balaksu Grassland in Xinjiang, China still keep the way of natural grazing. The special geographical environment and unique customs of herdsmen have evolved their unique eating habits. The natural fermented yogurt of local herdsmen maintains the original ecology and is not affected by industrial production. Our team isolated and identified LFZS09 from grassland naturally fermented yogurt, and this study is the first to investigate its biological activity. To offer a theoretical foundation for future research and development of probiotics or parabiotics so as to enhance motor function, a mouse model of dyskinesia was established by intraperitoneally injecting mixed antibiotics into the mouse. Then, the treadmill test and weight-bearing swimming test were considered as parameters and conducted to investigate the regulatory effect of LFZS09 on the motor function of the antibiotic-injected mice.
Material and Methods
Strains
The China General Microbiological Culture Collection Center (Beijing, China) has the LFZS09 strain on file (preservation registration number 18225). Colonies maintained at −80°C were inoculated in 5 mL of MRS liquid medium and incubated at 37°C for 24 h. Following incubation, a passage culture was performed. Then, 1 mL of the bacterial culture medium was transferred to a sterile centrifuge tube and centrifuged at 4000 rpm for 10 min to remove the top layer. Moreover, 1 mL of the bacterial culture was collected in a sterile centrifuge tube, placed in a 100°C water bath for 30 min, and centrifuged at 4000 r/min for 10 min. Similarly, when the bacteria precipitated, they were resuspended in sterile physiological saline after discarding the top clear liquid. This solution was labeled as a heat-inactivated bacterial solution and was later used for gastric lavage.
Animal Models
A laboratory animal production license (SCXK (Hunan) 2019–0004) was obtained from Hunan Slyke Jingda Laboratory Animal Co., Ltd (Changsha, Hunan, China) for 50 male (7 weeks old), specific pathogen-free (SPF) C57BL/6J mice aged 6–8 weeks. The animals were exposed to a light–dark cycle set to 12 h, 60% humidity, and an ambient temperature of 25°C ±1°C. Before the formal trial started, the SPF mice were fed adaptively for 7 days.
The mice were randomized into 5 groups, each with 10 mice, following the results of adaptive feeding: normal group, model group, caffeic acid (CA, Aladdin Reagent (Shanghai) Co., Ltd, Shanghai, China) group, lived group (lived bacteria), and heat-killed (HK, parabiotics) bacteria group. Caffeic acid can promote the oxidation and utilization of fatty acids, increase the uptake and utilization of fat by muscles, thereby improving physical exercise performance.11 At the same time, studies have shown that caffeic acid has a certain regulatory effect on intestinal flora and can promote the proliferation of beneficial microorganisms in the gut,12 so caffeic acid was used as a positive control in this study. The particular course of therapy for each group was as follows: the model group was intraperitoneally injected with a sterile mixed antibiotic solution every other day for 14 days in a row, and each mouse in the normal group was gavaged with 0.2mL distilled water (placebo). For 14 days in a row, the CA group was intraperitoneally injected with a sterile mixed antibiotic solution every other day. At the same time, they received CA (10 mg/kg) via gavage. For 14 days in a row, the lived group was intraperitoneally injected with a sterile mixed antibiotic solution every other day and received concurrent bacterial solution gavages. For 14 days in a row, the HK group was intraperitoneally injected with a sterile mixed antibiotic solution every other day. They also received a gavage of heat-inactivated bacterial solution. The injection and intragastric dosage was 0.1 mL/10 g. A mixture of antibiotics in 0.9% normal saline solution contained 5 mg/mL neomycin, 25 mg/mL vancomycin, 0.1 mg/mL amphotericin, 10 mg/mL ampicillin, and 5 mg/mL metronidazole (Aladdin Reagent (Shanghai) Co., Ltd). After a 12-h fast, the mice were sedated and bearded, and their eyes were promptly enucleated. After the experiment, blood was then drawn from the mice in tubules devoid of endotoxins. The mouse serum samples were labeled and placed in groups. They were centrifuged for 10 min at 4°C and 3000 rpm, and the supernatant was frozen at −80°C to prevent repeated freezing and thawing.13
The mouse serum was swiftly extracted and divided. The whole brain, liver, and cecum of the mice were removed, and the mucosa and serum attached to them were washed away with precooled normal saline. The surface water on the parts washed was rapidly blotted dry using filter paper. The whole brain tissue was cut into three sections. For tissue observation, one portion was submerged in 20 times the volume of a 4% (v/v) poly-formaldehyde solution (made in PBS, Solarbio, Beijing, China). To prevent repeated freezing and thawing, the other portion was rapidly mashed in an electric homogenizer with 9 times pre-cooled 0.9% saline. The portion was then centrifuged at 4°C at 8000 rpm/min for 15 min. The supernatant was collected and frozen at −80°C for the enzyme-linked immunosorbent assay (ELISA). For the fluorescence quantitative measurement, the third portion was rapidly frozen in liquid nitrogen in a tin foil, moved to a sterile 1.5-mL eppendorf tube, and frozen at −80°C. The liver index was calculated by the formula, liver index=liver weight (g)/body weight (g)×100.
Weight-Bearing Swimming and Treadmill Exhaustion Experiments of Mice
The experimental mice were subjected to weight-bearing swimming and treadmill fatigue tests according to the protocols described in the literature.11 The mice were submerged in water at 25°C ±3°C and allowed to swim until they were tired before the final modeling stage. When a mouse was immersed in water for more than 10s and, upon being placed on a level surface, the mouse could not complete the righting reflex, Exhaustion was declared. After being swiftly dried using a blow dryer, the tired mice were placed back in their cages. In each swimming experiment, a glass water cylinder (height, 80 cm; diameter, 50 cm) was used as the container. The water’s depth was 40 cm. Before the official test, the mice were preacclimatized, where they were allowed to swim freely for 3–5 min without having the lead roll fastened to their tails. In the official experiment, a 5% body weight lead block was tied at the base of the mouse tails. The test was conducted after the hair of the mice was thoroughly wet. When the mice were balanced and floating, a glass rod was pushed toward them. The level of tiredness of the mice was assessed, and their weight-bearing swimming times were noted when they could not stay under the water’s surface for more than 8 s.
The running platform (ZH-PT mouse experimental platform, Anhui Zhenghua Biological Instrument Equipment Co., Ltd, Huaibei, Anhui, China) of the mice was used for the exhaustion test following the final modeling. The mice were allowed to freely roam on the track for 1–3 min before the experiment so that they could become acclimated to their surroundings. The running platform’s program was established as follows: 30 min of training, 1 min of rest, 3.0 mA of electric intensity, 4 m/s2 of acceleration, 20° of slope, and a maximum speed of 20 m/min. The mice were considered exhausted and fatigued when they could not run on the track channel, were left at the end of the track or in a forward crouch for an extended period, and neither the electric shock nor human drive could force them to run to the front of the track. The distance and running duration were simultaneously recorded at this point.
Detection of Oxidative Stress and Inflammation Levels in the Serum and Brain Tissue of Mice
Superoxide dismutase (SOD), reduced glutathione (GSH), and malondialdehyde (MDA) kits (Solarbio) were used to measure the amount of oxidative stress in the serum and brain tissue homogenates. Concurrently, ELISA kits were used to ascertain TNF-α, IL-6, and IL-10 (Solarbio) concentrations in the brain tissue homogenate. The test protocol was executed according to the kit’s instructions (Varioskan LUX multifunctional enzyme-linked immunosorbent assay instrument, Thermo Fisher Scientific, Waltham, MA, USA).
Histomorphological Observation of the Mouse Brain
Following harvesting using the coronal technique, brain tissues fixed with the tissue fixative solution were dehydrated in 70%, 80%, 95%, and 100% alcohol. The brain tissues had a thickness of 4 mm. Every dehydration procedure took 30 min. The tissues were first dehydrated, submerged in xylene for 20 min, and finally embedded in paraffin for 30 min. The tissue was subsequently immersed for 30 min in paraffin wax. A microtome was used to cut the tissue into 4-μm slices. Once the tissue flakes gently adhered to the slide, hematoxylin-eosin staining was performed on the flakes, and the flakes were allowed to dry in the oven. Following straining, the histological was examined under a light microscope (BX43, Olympus, Tokyo, Japan).
Detection of CREB, ERK1/2, and BDNF in the Brain Tissue and the Relative mRNA Expression Levels of Intestinal Barrier Genes zo-1, Occludin-1, and Claudin-1 in the Cecal of Mice
RNA was extracted from the brain and cecal tissues frozen at −80°C by using the Trizol (Solarbio) technique. Following purity testing, RNA was stored for usage at −80°C. The extracted RNA was reverse transcribed on ice at 65°C (5min), →42°C (60 min), →70°C (5min), and →4°C (hold) by using the ReverTra Ace® qPCR RT Master Mix kit. The relevant tissue’s cDNA was extracted. Using the TOYOBO SYBR® Green Realtime PCR Master Mix kit, the primers and template cDNA were combined (Thermo Fisher Scientific). The fluorescent dye and high-fidelity enzyme were added according to the kit’s instructions, and the mixture was allowed to sit at 95°C for 5 min. PCR amplification was performed at 40 cycles of 94°C for 30s, 60°C for 40 s, and 72°C for 1 min. β-Actin was used as the reference gene for quantitative real-time PCR (StoponePlus, Thermo Fisher Scientific). Table 1 presents the primer sequences. The comparative CT value approach (2−ΔΔCT) was used for the calculation.14
 |
Table 1 Primer Sequences in This Experiment |
Count of Intestinal Flora in Mice
Mouse feces were collected aseptically, weighed (about 0.5 g), added into 4.5 mL sterile normal saline, fully shaken and mixed, left for 5 min, and the supernatant was diluted by 10 times gradient to 10−6. Diluents were inoculated on Bifidobacterium medium, bile esculin azide agar medium, DeMan, Rogosa and Sharpe (MRS) medium, eosin methylene blue (EMB) medium and sorbitol macconkey agar (SMAC) medium, respectively. Bifidobacterium and Lactobacillus were cultured at 37°C for 48 h, while Enterococcus, Enterobacter and Clostridium perfringens were cultured at 37°C for 24 h. After the end of culture, colony plate count was carried out, the number of each colony in each gram of feces was calculated (CFU/g), for statistical analysis.
Data Processing
One-way analysis of variance with the Turkey test was applied for data analysis. A difference was considered significant when p < 0.05. EXCEL 2019 software was used to plot the data.
Results
Organ Coefficient of the Mouse Liver
The organ index is a crucial metric for assessing toxicity in mice as it depicts the mobility and physiological operation of organs in a stable environment. A low organ index possibly indicates muscle atrophy or harmful effects on the body, whereas a high organ index possibly indicates organ congestion and edema. Figure 1 displays the related findings. The organ index was determined by weighing the mouse livers. The mice receiving antibiotic therapy developed liver congestion and edema, as evidenced by the considerably greater liver organ index of the model group than of the normal group (p < 0.05). This might potentially affect the normal physiological function of the body. The organ index decreased in the CA group, but it remained considerably higher than that in the control group (p < 0.05). After treatment with LFZS09 (both in the lived and HK groups), the liver organ index was significantly lower than that in the model group. The results indicated that LFZS09 was effective in regulating antibiotic-induced liver congestion and edema in the mice and restoring their normal physiological function.
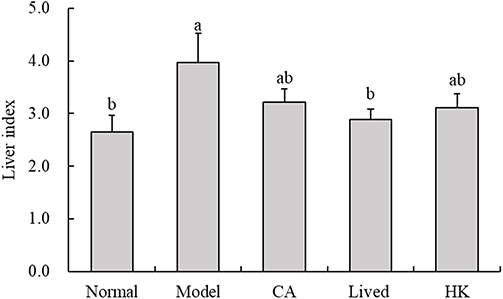 |
Figure 1 The liver index of experimental mice (n=10). a-b The same numerical value indicates insignificant differences, while different letters indicate significant differences p<0.05. Liver index=liver weight (g)/body weight (g)×100. CA, caffeic acid; lived, lived LFZS09 (probiotic); HK, heat-killed LFZS09 (parabiotics). |
Histopathological Observation of the Mouse Brain
The brain tissue shape in the normal group was unaltered, and the cells were densely organized (Figure 2). Inflammatory lesions were observed on the brain surface in the model group. The space between neurons and surrounding tissues near the lesions widened, displaying that the antibiotics caused edematous alterations in the brain tissue. In general, the nerve cells in the tissue were hyperstained, and the nuclei were not distinct. This indicated that the antibiotics produced pyknosis of brain tissue cells, thereby disrupting their normal physiological shape. Some neurons in the cerebral cortex of the bacterial solution-treated mice deteriorated, but inflammatory cell infiltration was not found. In the heat-inactivated bacterial solution-treated group, a high rate of neuronal cell edema, cell swelling, and cytoplasmic vacuolization were noted. Finally, LFZS09 has a protective effect on the brain tissue of the antibiotic-treated mice.
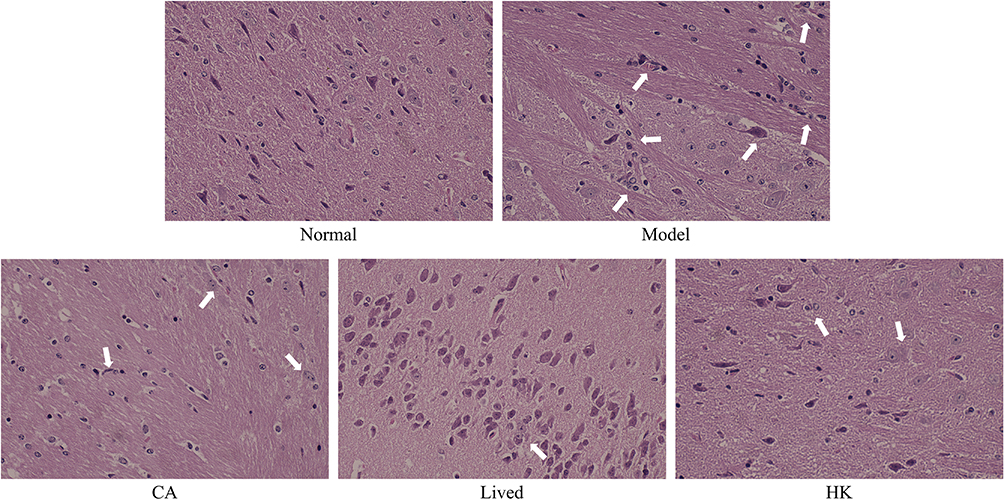 |
Figure 2 The H&E staining histopathological observation of the mouse brain (×100). The position indicated by the arrow is the tissue lesion. CA, caffeic acid; lived, lived LFZS09 (probiotic); HK, heat-killed LFZS09 (parabiotics). |
Changes in the Motor Function of Mice
Each group pf mice underwent the swimming fatigue and treadmill exhaustion trials, and the findings are presented in Table 2. The model group had considerably poorer swimming and running times than the normal group (p < 0.05), demonstrating that antibiotic treatment caused motor impairment in the mice. The exercise duration of the bacterial solution- or inactivated bacterial solution-treated mice was prolonged to varying degrees, suggesting that LFZS09 can ameliorate antibiotic-induced dyskinesia. However, the swimming and running times of each mouse group were significantly lower than those of the normal group. Overall, the group that received bacterial solutions as therapy exhibited greater recovery.
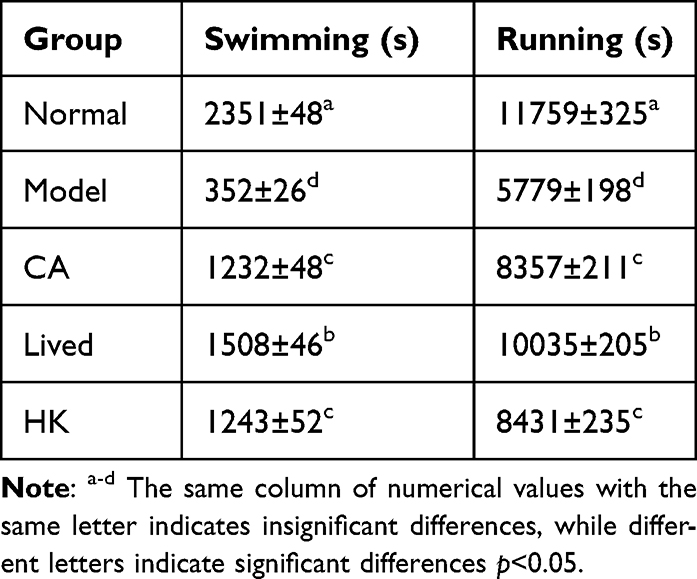 |
Table 2 The Parameters Related to Locomotor Function in Mice |
Changes in Oxidative Stress Levels in Mice
Figure 3 depicts GSH, MDA, and SOD levels in the mouse brain tissue and serum from each group. The antibiotic-treated mice had significantly lower GSH and SOD levels in the serum and brain tissue compared with the normal group (p < 0.05), and MDA levels was raised in the antibiotic-treated mice. This indicated that antibiotics can cause oxidative stress and affect normal physiological function. After the LFZS09 treatment of the mice, the oxidative stress level increased to a point that was considerably greater than that of the model group.
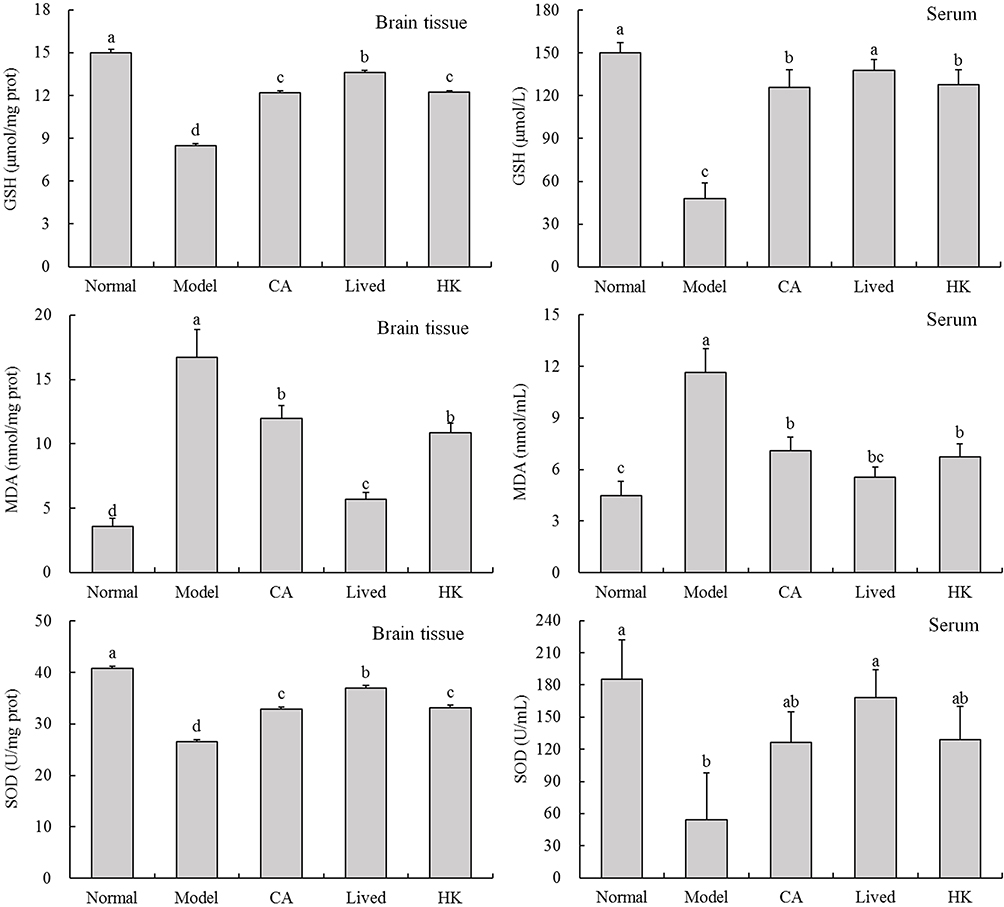 |
Figure 3 The GSH, MDA, and SOD levels in the mouse brain tissue and serum. a-d The same numerical value indicates insignificant differences, while different letters indicate significant differences p<0.05. Abbreviations: CA, caffeic acid; lived, lived LFZS09 (probiotic); HK, heat-killed LFZS09 (parabiotics). |
Changes in Brain Inflammation Levels in Mice
Figure 4 depicts the inflammation levels in the brains of mice from each group. The antibiotic-treated mice had significantly higher IL-6, IL-10, and TNF-α levels in their brains than the normal group (p < 0.05). This suggests that antibiotics can increase the levels of inflammatory factors in the body, thereby affecting normal physiological function. After LFZS09 treatment, the levels of inflammatory factors reduced, which were lower than those in the model group.
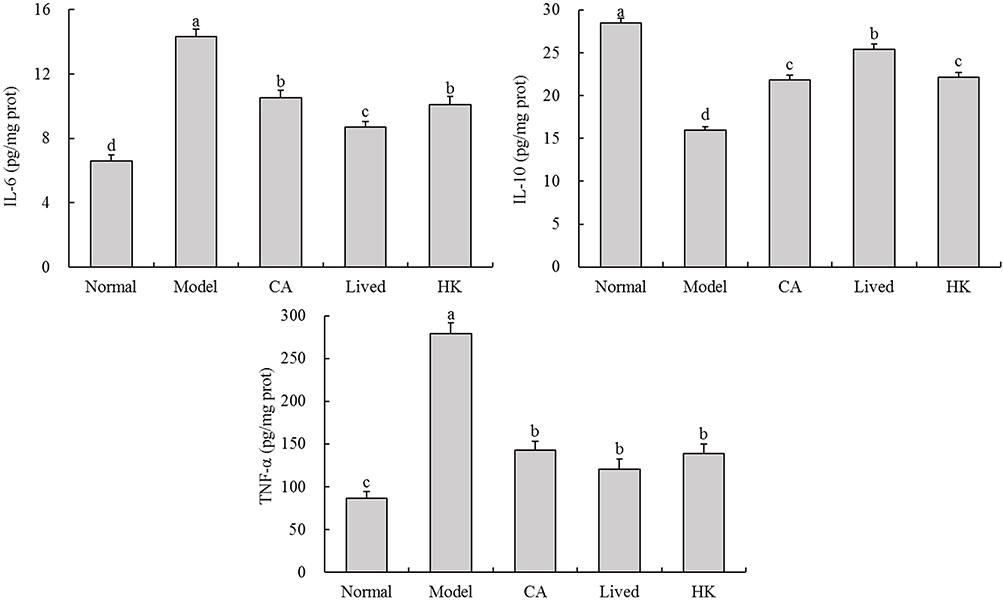 |
Figure 4 The IL-6, IL-10, and TNF-α levels in the mouse brain tissue. a-d The same numerical value indicates insignificant differences, while different letters indicate significant differences p<0.05. Abbreviations: CA, caffeic acid; lived, lived LFZS09 (probiotic); HK, heat-killed LFZS09 (parabiotics). |
Changes in the Relative Expression of BDNF Metabolic Pathway Genes in the Brain of Mice
Figure 5 depicts the relative expression of BDNF metabolic pathway genes in the brains of each mouse group. The antibiotic-treated mice had significantly lower brain expression levels of BDNF, CREB and ERK1/2 genes in the BDNF metabolic pathway than the normal group (p < 0.05). This suggests that antibiotics reduce the expression levels of the BDNF metabolic pathway-related genes in the brain, thereby causing neurotransmitter imbalance. It affects the mice’s ability to maintain normal nervous system function. The LFZS09-treated group exhibited considerably greater expression levels of the three critical genes than the model group (p < 0.05). The expression of the BDNF metabolic pathway-related genes in the mouse brain was generally more significantly affected in the bacterial solution-treated group.
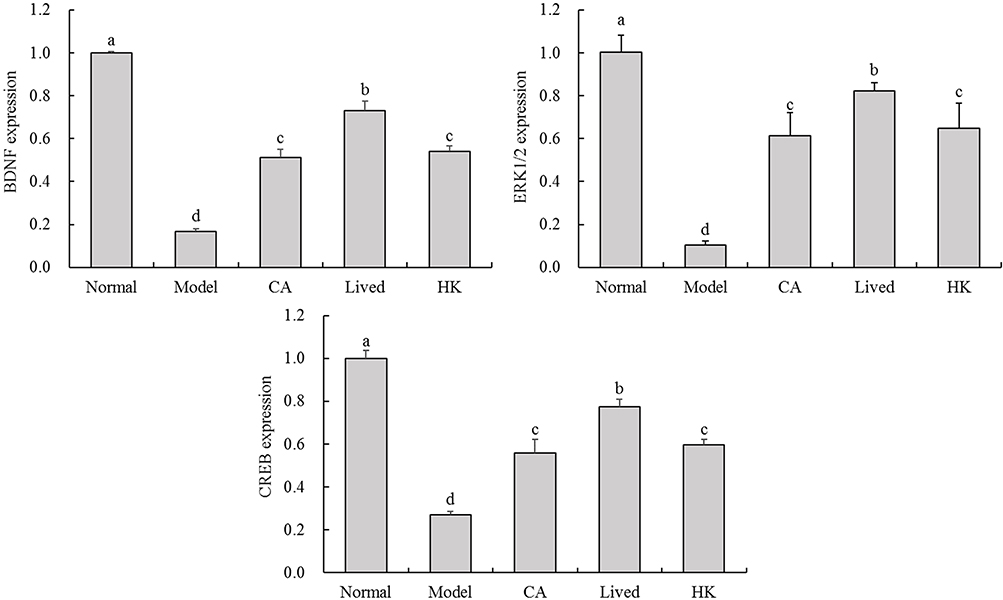 |
Figure 5 The BDNF, CREB and ERK1/2 mRNA expression in the mouse brain tissue. a-d The same numerical value indicates insignificant differences, while different letters indicate significant differences p<0.05. Abbreviations: CA, caffeic acid; lived, lived LFZS09 (probiotic); HK, heat-killed LFZS09 (parabiotics). |
Changes in the Relative Expression of Cecal Intestinal Barrier Genes in Mice
Figure 6 illustrates the variations in the relative expression of genes encoding the cecal intestinal barrier in each mouse group. The administered antibiotics suppressed the expression of intestinal barrier-related genes (Claudin-1, ZO-1 and Occludin-1) in the mouse cecum, ultimately disrupting the intestinal barrier and causing an imbalance in physiological processes. Following the LFZS09 treatment, the relative mRNA expression levels of the three barrier genes increased significantly, thereby surpassing the expression levels of the model group by a significant margin (p < 0.05). This suggests that LFZS09 mitigates the antibiotic-induced damage to the intestinal barrier, with the bacterial solution-treated group exhibiting a more notable protective effect.
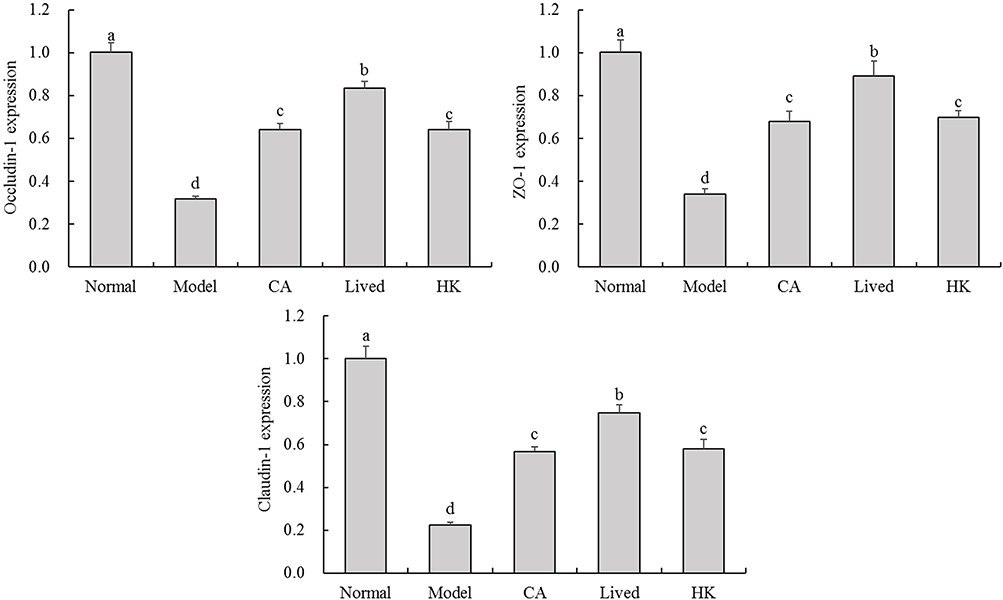 |
Figure 6 The Claudin-1, ZO-1 and Occludin-1 mRNA expression in the mouse cecal intestinal barrier tissue. a-d The same numerical value indicates insignificant differences, while different letters indicate significant differences p<0.05. Abbreviations: CA, caffeic acid; lived, lived LFZS09 (probiotic); HK, heat-killed LFZS09 (parabiotics). |
Changes of Intestinal Flora in Mice
The experimental results showed that compared with the normal group of mice, the number of Lactobacillus and Bifidobacterium in the intestines of mice after antibiotic treatment significantly (p<0.05) decreased (Table 3), while the number of Enterococcus and Clostridium perfringens significantly (p<0.05) increased, while the number of Enterobacter had no significant change. CA can promote Lactobacillus in mice, and can also reduce Enterococcus and Clostridium perfringens. Limosilactobacillus fermentum ZS09 has a significant (p<0.05) regulatory effect on intestinal bacteria, which can reduce the number of Enterococcus and Clostridium perfringens and the number of Lactobacillus and Bifidobacterium, and Enterococcus, Clostridium perfringens, Lactobacillus and Bifidobacterium under live bacteria (Lived group) treatment are significantly (p<0.05) different from other groups.
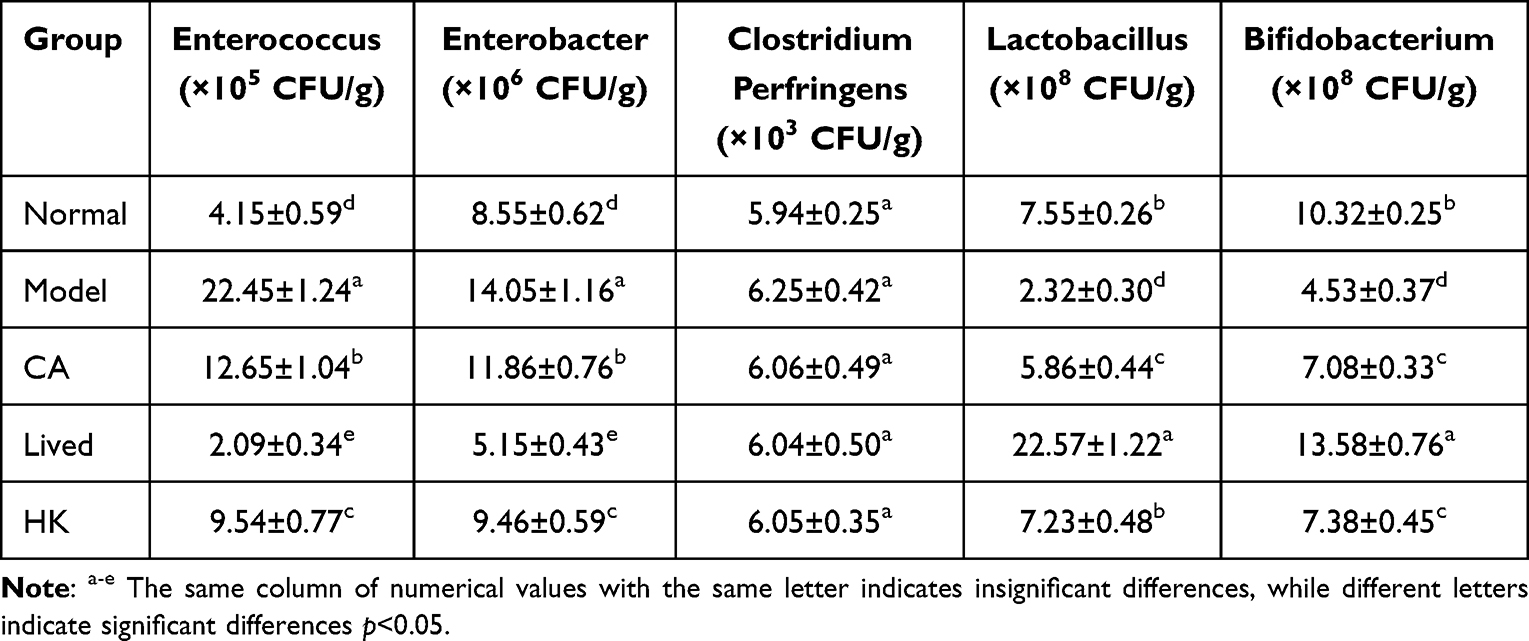 |
Table 3 The Number of Intestinal Bacteria in Mice |
Discussion
Oxidative stress, a redox signal in physiology, is an imbalance between oxidant production and antioxidant defense and may harm biological systems.15 Numerous health problems, including cancer, Alzheimer’s disease, atherosclerosis, and chronic obstructive pulmonary disease, have been linked to oxidative stress. Oxidants can cause cell damage through numerous different mechanisms.16 Oxidative stress causes diseases through two major processes. A primary cause of cell malfunction and mortality under oxidative stress is the generation of reactive molecules, including •OH, ONO−, and HOCl−. These molecules directly damage membrane lipids, structural proteins, enzymes, nucleic acids, and other macromolecular components. Aberrant redox signaling is the second action mechanism of oxidative stress. Second messengers, particularly H2O2, are produced by cells in response to physiological stimuli.17 Nonphysiological H2O2 generation during oxidative stress can cause abnormal REDOX signaling. Because oxidative stress can disrupt multiple signaling pathways and affect various biological processes, including protein modification, inflammation promotion, apoptosis induction, autophagy relaxation, and mitochondrial function impairment, oxidative stress is suggested to play a substantial role in disease and toxicity.18–20 SOD and GSH can efficiently eliminate excess free radicals from the body, which directly and indirectly indicates the body’s antioxidant activity.21 In this study, the brains of the LFZS09-treated mice had considerably greater SOD and GSH levels than those of mice in the model group (p < 0.05), suggesting that LFZS09 controls the antibiotic-induced aberrant oxidation level in the body.
Furthermore, free radicals act upon lipid peroxidation in vivo. The resultant oxidation product, MDA, profoundly affects cellular metabolism and fundamental activities, and the MDA concentration corresponds to the oxidative stress level of the body.22 The Pediococcus pentose-treated mice had significantly higher brain MDA levels than the model group (p < 0.05). This suggests that LFZS09 controls the antibiotic-induced abnormal levels of oxidative stress factors, alleviates the oxidative stress state, and promotes various functions of the body to return to normal.
When environmental factors or genetic mutations trigger abnormal induction, TNF-induced cell death becomes a highly detrimental process during the early stages of various (sterile) inflammatory diseases.23 However, it also aids in establishing an appropriate immune response during microbial infection. Thus, in addition to triggering inflammatory mediator production, TNF indirectly causes inflammation by triggering inflammatory immune responses, inducing cell death, and promoting illness onset. When bodily inflammation occurs, the levels of cytokines such as TNF-α and IL-6 also increase substantially; hence, these cytokines may be used as inflammatory markers.24 In this experiment, IL-6 and IL-10 levels in the model group were markedly greater (p < 0.05) than those in the normal group. After the mice received Limosilactobacillus fermentum treatment, the levels of some inflammatory factors remained higher than those in the normal group but were significantly lower than those in the model group (p < 0.05). This suggests that the inflammatory factor levels increased in the antibiotic-treated mice, potentially resulting in inflammation. Although LFZS09 could affect the normal physiological and biochemical states of the mice, LFZS09 intervention reduced the inflammatory response by delaying inflammatory factor production.
BDNF is among the neurotrophins in the central nervous system that have been studied and described the most. By binding to and activating tyrosine Kinase receptor B (TrkB), a broad Trk receptor family member, BDNF regulates various cellular processes involved in the maintenance and development of the brain’s normal physiological functioning. It induces the activation of downstream serine/threonine kinases, which then stimulate ERK, MAPK/ERK kinase, and CREB phosphorylation, thereby augmenting memory and learning capacities. Glial cells, microglia, and glutamatergic neurons in the brain express BDNF.25,26 Furthermore, BDNF can stimulate the growth of neuromuscular synapses, which indicates that BDNF possibly serves as a target for therapeutic intervention in muscular disorders.27 The study’s findings demonstrated that the relative expression levels of BDNF and ERK1/2 genes in the BDNF metabolic pathway were considerably lower in the brains of the antibiotic-treated mice than in the control group (p < 0.05). Following the LFZS09 treatment, the relative expression levels of the three critical genes increased considerably. Compared with the model group, the relative expression levels of the three vital genes in the brain were considerably higher (p < 0.05). According to these findings, Limosilactobacillus fermentum may successfully restore the antibiotic-induced decrease in neurotransmitter expression, preserve the homeostasis of the nervous system, and thus control the regular physiological processes of the body.
The intestinal epithelial barrier, which is composed of epithelial cells, epithelial glycoproteins, and the mucus layer, inhibits pathogen entry.28 Intercellular junctions, such as tight junctions, gap junctions, adherens junctions, and desmosomes, form various functional complexes and are essential components of the intestinal epithelial barrier.29 The most significant intercellular connections, tight junctions, control intestinal epithelial permeability and preserve the physiological function of the intestinal barrier.30 The three most substantial tight junction proteins are occludin-1, claudin-1, and zo-1. These three proteins have major roles in preserving the intestinal barrier integrity and intestinal cell polarity.31 Because of the intestinal microbiome’s role in intestinal epithelial cell signaling and its effect on intestinal barrier function, tight junction proteins have been demonstrated to play a critical role in the healing of intestinal epithelial injury.32 By preserving the production and distribution of tight junction proteins, which are critical to the intestinal barrier integrity, intestinal bacteria control the intestinal barrier function. Gut bacteria regulate the intestinal barrier function by maintaining the expression and distribution of tight junction proteins.33 In this experiment, the relative mRNA expression of occludin-1 in the mouse cecum in the bacterial solution-treated group was extremely significantly higher than that in the model group (p < 0.05), and the relative mRNA expression of occludin-1 was significantly lower in the model group than in the normal group (p < 0.05). These findings suggested that LFZS09 therapy mitigates the antibiotic-induced intestinal barrier damage and allows the mouse bodies to recover some of their regular physiological processes.
The gut microbiota can suppress the development and colonization of pathogenic microbes by directly fighting with them for nutrition or by producing antimicrobial compounds such as short-chain fatty acids (SCFAs).34 Through numerous methods, gut microorganisms improve the gut barrier function. Beneficial microbes produce SCFAs that enhance tight junctions, lower intestinal permeability, and encourage the growth and differentiation of intestinal epithelial cells.35 Antibiotic overuse can cause an imbalance of gut flora. Disruptions in this intestinal flora balance can induce inflammatory bowel disease, obesity, type 2 diabetes, cardiovascular illnesses, and some neurological disorders. They can also induce intestinal barrier malfunction and increase intestinal permeability.36 Consequently, this disruption impairs how the human body functions and limits its range of motion. Probiotic supplements may efficiently control the gut flora and repair intestinal damage.37 In the present study, LFZS09 performed a similar function. It aided the mice in recuperation and improved their capacity for activity.
Enterococcus is a part of the normal intestinal flora of humans and animals. Studies have confirmed that Enterococcus is an important nosocomial infection pathogen next to staphylococcus. Enterococcus can not only cause urinary tract infection, skin and soft tissue infection, but also cause life-threatening abdominal infection, septicemia, endocarditis and meningitis.38 Clostridium perfringens is widely found in soil, animal and human intestines. Clostridium perfringens can produce a variety of external toxic cords, causing hemolysis, tissue necrosis, vascular endothelial cell damage, and increased vascular permeability. Hyaluronidase and collagenase can destroy the intercellular substance, decompose the collagen fibers of subcutaneous tissue and muscle tissue, and disintegrate tissues, which is conducive to the spread of bacteria and toxins.39 Both Lactobacillus and Bifidobacterium are important beneficial microorganisms in the gut, which play an important role in maintaining intestinal health, promoting absorption, and helping the body resist the invasion of harmful materials.40 LFZS09 plays an obvious regulatory role in the intestinal flora of mice with dyskinesia, increasing the number of Lactobacillus and Bifidobacterium in the intestine, and decreasing the number of Enterococcus and Clostridium perfringens. Thus, by increasing the beneficial bacteria and reducing the harmful bacteria in the intestine, LFZS09 can inhibit brain tissue damage and enhance the exercise ability.
In this work, a mouse motor dysfunction model was established by intraperitoneally administering a mixed antibiotic solution. Antibiotics influence oxidative stress and inflammatory levels in the body, which causes neurotransmitter imbalance and motor impairment in mice. The experimental outcomes revealed that LFZS09 successfully increased the amount of time that mice could swim and run. It also had a certain calming impact on the antibiotic-induced systemic inflammatory response in mice, which could lower TNF-α and IL-6 levels. Additionally, LFZS09 increased SOD levels, decreased MDA accumulation, and boosted antioxidants in the mice. According to the mRNA experiment data, the strain preserved intestinal homeostasis, promoted the expression of intestinal barrier repair-related genes, and preserved the regular physiological processes of the body. Moreover, it preserved the homeostasis of the nervous system, controlled motor function, and dramatically boosted the expression of the neurotransmitter metabolism pathway-related genes.
Conclusion
In conclusion, by influencing the expression of neuroconstitution, LFZS09 reduces inflammation, controls oxidative stress, and regulates the body’s motor performance. Specifically, LFZS09 serves as an intestinal barrier regulator and efficiently heals brain–intestinal injury. Nevertheless, further studies are warranted to fully understand the microbiota and intestinal barrier mechanism. Additionally, the results of clinical studies will support the use of LFZS09 more effectively. This study investigated the effect of LFZS09 intervention on the recovery of antibiotic-induced dyskinesia. It examined the changes of some representative microbes, but lacked a comprehensive analysis of the gut microbial categories and the dynamic changes of gut microbiota over the experimental period. The process microbial composition was measured from the beginning of the experiment to the end of the experiment. In future studies, the detection of changes in gut microbial species and processes is the focus of intensive investigation.
Ethics Committee
The animal experiment was approved by the Animal Experiment Ethics Committee of Collaborative Innovation Center for Child Nutrition and Health Development, Chongqing University of Education (Chongqing, China), with the approval number 2023030021B. The animal experimental process was in accordance with 2010/63/EU directive.
Funding
There is no funding to report.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wu M, Liu J, Li F, et al. Antibiotic-induced dysbiosis of gut microbiota impairs corneal development in postnatal mice by affecting CCR2 negative macrophage distribution. Mucosal Immunol. 2020;13(1):47–63. doi:10.1038/s41385-019-0193-x
2. Scheperjans F, Aho V, Pereira PA, et al. Gut microbiota are related to Parkinson’s disease and clinical phenotype. Mov Disord. 2015;30(3):350–358. doi:10.1002/mds.26069
3. Ferreyra JA, Wu KJ, Hryckowian AJ, et al. Gut microbiota-produced succinate promotes C. difficile infection after antibiotic treatment or motility disturbance. Cell Host Microbe. 2014;16(6):770–777. doi:10.1016/j.chom.2014.11.003
4. Zingman LV, Zhu Z, Sierra A, et al. Exercise-induced expression of cardiac ATP-sensitive potassium channels promotes action potential shortening and energy conservation. J Mol Cell Cardiol. 2011;51(1):72–81. doi:10.1016/j.yjmcc.2011.03.010
5. Peckham PH, Mortimer JT, Marsolais EB. Alteration in the force and fatigability of skeletal muscle in quadriplegic humans following exercise induced by chronic electrical stimulation. Clin Orthop Relat Res. 1976;114:326–333.
6. Bequet F, Peres M, Gomez-Merino D, et al. Simultaneous NMR microdialysis study of brain glucose metabolism in relation to fasting or exercise in the rat. J Appl Physiol. 2020;88(6):1949–1954. doi:10.1152/jappl.2000.88.6.1949
7. Jiménez-Maldonado A, Cerna-Cortés J, Castro-Rodríguez EM, et al. Effects of moderate- and high-intensity chronic exercise on brain-derived neurotrophic factor expression in fast and slow muscles. Muscle Nerve. 2016;53(3):446–451. doi:10.1002/mus.24757
8. Li W, He QZ, Wu CQ, et al. PFOS disturbs BDNF-ERK-CREB signalling in association with increased microRNA-22 in SH-SY5Y cells. Biomed Res Int. 2015;2015:302653. doi:10.1155/2015/302653
9. McNamara MP, Cadney MD, Castro AA, et al. Oral antibiotics reduce voluntary exercise behavior in athletic mice. Behavioural Proc. 2022;199:104650.
10. Simons LA, Amansec SG, Conway P. Effect of lactobacillus fermentum on serum lipids in subjects with elevated serum cholesterol. Nutr, Metab Cardiovasc Dis. 2006;16(8):531–535.
11. Courneya KS. Efficacy, effectiveness, and behavior change trials in exercise research. Int J Behav Nutr Phys Act. 2010;7:81. doi:10.1186/1479-5868-7-81
12. Zhang Z, Wu X, Cao S, et al. Caffeic acid ameliorates colitis in association with increased akkermansia population in the gut microbiota of mice. Oncotarget. 2016;7(22):31790–31799. doi:10.18632/oncotarget.9306
13. Desbonnet L, Clarke G, Traplin A, et al. Gut microbiota depletion from early adolescence in mice: implications for brain and behaviour. Brain Behav Immun. 2015;48:165–173. doi:10.1016/j.bbi.2015.04.004
14. Zhao X, Yi R, Zhou X, et al. Preventive effect of Lactobacillus plantarum KSFY02 isolated from naturally fermented yogurt from Xinjiang, China, on d-galactose–induced oxidative aging in mice. J Dairy Sci. 2019;102(7):5899–5912. doi:10.3168/jds.2018-16033
15. Skipitari M, Kalaitzopoulou E, Papadea P. Titanium dioxide nanoparticle-based hydroxyl and superoxide radical production for oxidative stress biological simulations. J Photochem Photobiol A. 2023;435:114290. doi:10.1016/j.jphotochem.2022.114290
16. Kohli A, Kothiyal P. A review on Alzheimer disease -its treatment, future prospects and the role of Juglans regia in Alzheimer’s disease based on pathogenetic research. Int J Pharm Res Rev. 2015;4(12):6–20.
17. Rodrigues E, Mariutti LRB, Mercadante AZ. Scavenging capacity of marine carotenoids against reactive oxygen and nitrogen species in a membrane-mimicking system. Mar Drugs. 2012;10(8):1784–1798. doi:10.3390/md10081784
18. Sies H. Role of metabolic H2O2 generation: redox signaling and oxidative stress. J Biol Chem. 2014;289:8735–8741. doi:10.1074/jbc.R113.544635
19. Bryan S, Yang G, Wang R, et al. Cystathionine gamma-lyase-deficient smooth muscle cells exhibit redox imbalance and apoptosis under hypoxic stress conditions. Exp Clin Cardiol. 2011;16(4):e36–41.
20. Akki R, Siracusa R, Morabito R, et al. Neuronal-like differentiated SH-SY5Y cells adaptation to a mild and transient H2O2-induced oxidative stress. Cell Biochem Funct. 2018;36:56–64. doi:10.1002/cbf.3317
21. Huang K, Liang Y, Wang K, et al. Influence of circulating nesfatin-1, GSH and SOD on insulin secretion in the development of T2DM. Front Public Health. 2022;10:882686. doi:10.3389/fpubh.2022.882686
22. Tsikas D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges. Anal Biochem. 2017;524:13–30. doi:10.1016/j.ab.2016.10.021
23. Bueter W, Dammann O, Leviton A. Endoplasmic reticulum stress, inflammation, and perinatal brain damage. Pediatr Res. 2009;66:487–494. doi:10.1203/PDR.0b013e3181baa083
24. Singh AK, Vinayak M. Curcumin attenuates CFA induced thermal hyperalgesia by modulation of antioxidant enzymes and down regulation of TNF-α, IL-1β and IL-6. Neurochem Res. 2015;40:463–472. doi:10.1007/s11064-014-1489-6
25. He Z, Jiang J, Kokkinaki M, et al. GDNF induces CREB-1, ATF-1, and CREM-1 phosphorylation and up-regulates c-fos transcription via the Ras/ERK1/2 pathway to promote mouse spermatogonial stem cell proliferation. Stem Cells. 2008;26(1):266–278. doi:10.1634/stemcells.2007-0436
26. Lee WR, Shen SC, Wu PR, et al. CSE1L Links cAMP/PKA and Ras/ERK pathways and regulates the expressions and phosphorylations of ERK1/2, CREB, and MITF in melanoma cells. Mol Carcinog. 2015;55(11):1542–1552. doi:10.1002/mc.22407
27. Neal AP, Guilarte TR. Molecular neurobiology of lead (Pb2+): effects on synaptic function. Mol Neurobiol. 2010;42(3):151–160. doi:10.1007/s12035-010-8146-0
28. Savkovic SD, Koutsouris A, Hecht G. Attachment of a noninvasive enteric pathogen, enteropathogenic Escherichia coli, to cultured human intestinal epithelial monolayers induces transmigration of neutrophils. Infect Immun. 1996;64(1):4480–4487. doi:10.1128/iai.64.11.4480-4487.1996
29. Yukako F. A novel function of botulinum toxin-associated proteins: the HA component disrupts intestinal epithelial intercellular junctions to increase toxin absorption. Toxicon. 2008;51:3. doi:10.1016/j.toxicon.2008.04.009
30. Laukoetter MG, Nava P, Nusrat A. Role of the intestinal barrier in inflammatory bowel disease. World J Gastroenterol. 2008;14(3):401–407. doi:10.3748/wjg.14.401
31. Acharya P, Beckel J, Ruiz WG, et al. Distribution of the tight junction proteins ZO-1, occludin, and claudin-4, −8, and −12 in bladder epithelium. Am J Physiol Renal Physiol. 2004;287(2):305–318. doi:10.1152/ajprenal.00341.2003
32. Tian S, Guo R, Wei S, et al. Curcumin protects against the intestinal ischemia-reperfusion injury: involvement of the tight junction protein ZO-1 and TNF-α related mechanism. Korean J Physiol Pharmacol. 2016;20(2):147–152. doi:10.4196/kjpp.2016.20.2.147
33. Earley ZM, Akhtar S, Green SJ, et al. Burn injury alters the intestinal microbiome and increases gut permeability and bacterial translocation. PLoS One. 2015;10(7):e0129996. doi:10.1371/journal.pone.0129996
34. Liu N, Wang H, Yang Z, et al. The role of functional oligosaccharides as prebiotics in ulcerative colitis. Food Funct. 2022;13(13):6875–6893. doi:10.1039/d2fo00546h
35. Zhou C, Li L, Li T, et al. SCFAs induce autophagy in intestinal epithelial cells and relieve colitis by stabilizing HIF-1 alpha. J Mol Med. 2020;98(8):1189–1202. doi:10.1007/s00109-020-01947-2
36. Limanskaya AY, Voloshyna TV, Baider AK, et al. Restoring the balance of vaginal and intestinal biotopes in preconceptional period after antibiotic therapy. Perinatol I Pediat. 2017;69(1):33–38. doi:10.15574/PP.2017.69.33
37. Prisciandaro LD, Geier MS, Butler RN, et al. Probiotic factors partially improve parameters of 5-fluorouracil-induced intestinal mucositis in rats. Cancer Biol Ther. 2011;11(7):671–677. doi:10.4161/cbt.11.7.14896
38. Lim BSH, Parolia A, Chia MSY, et al. Antimicrobial efficacy of QMix on enterococcus faecalis infected root canals: a systematic review of in vitro studies. Restorat Dentist Endodont. 2020;45(2):e23. doi:10.5395/rde.2020.45.e23
39. Finnie JW. Pathogenesis of brain damage produced in sheep by Clostridium perfringens type D epsilon toxin: a review. Aust Vet J. 2003;81(4):219–221. doi:10.1111/j.1751-0813.2003.tb11474.x
40. Nowak A, Paliwoda A, Błasiak J. Anti-proliferative, pro-apoptotic and anti-oxidative activity of Lactobacillus and Bifidobacterium strains: a review of mechanisms and therapeutic perspectives. Criti Rev Food Sci Nutr. 2019;59(21):3456–3467. doi:10.1080/10408398.2018.1494539
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.