Back to Journals » OncoTargets and Therapy » Volume 13
Latest Advances of Long Non-Coding RNA SNHG5 in Human Cancers
Authors Han W ![]() , Shi J, Cao J, Dong B, Guan W
, Shi J, Cao J, Dong B, Guan W
Received 6 March 2020
Accepted for publication 11 June 2020
Published 1 July 2020 Volume 2020:13 Pages 6393—6403
DOI https://doi.org/10.2147/OTT.S252750
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Leo Jen-Liang Su
Wei Han, Jia Shi, Jiachao Cao, Bo Dong, Wei Guan
Department of Neurosurgery, Third Affiliated Hospital of Soochow University, Changzhou, People’s Republic of China
Correspondence: Wei Guan No. 185, Juqian Street, Changzhou, Jiangsu 213003, People’s Republic of China
Email [email protected]
Abstract: Long non-coding RNAs (lncRNAs) have been potent regulators in the initiation and development of human cancers regarding their biological roles in the modulation of dosage compensation effect, epigenetics and cell differentiation. Recently, aberrant expression of lncRNA small nucleolar RNA host gene 5 (SNHG5) has been observed in various solid tumors, which was intently correlated with tumor range, metastasis, pathological stage and prognosis. Additional mechanical investigation disclosed that SNHG5 was involved in multiple cellular activities, including proliferation, migration, invasion, cell-cycle, apoptosis and autophagy, via targeting miRNAs, signaling pathways and other biological molecules or proteins. In this review, we summarized the latest advances made towards understanding the roles of SNHG5 in human cancers and further discussed potential methods that could be adopted for clinical interventions.
Keywords: lncRNA, SNHG5, biomarker, therapeutic target, human cancers
Introduction
The human genome-wide studies have demonstrated that approximately 70% of the human genome is transcribed into RNAs, while the ratio of non-coding RNAs (ncRNAs) is up to 98%.1 The ncRNAs consist of long non-coding RNAs (lncRNAs), microRNAs (miRNAs), small interfering RNAs (siRNAs), transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), small nuclear RNAs (snRNAs), small nucleolar RNAs (snoRNAs) and other types of ncRNAs.2 Though ncRNAs were initially considered transcriptional noises, more researchers have elucidated functional roles of ncRNAs in various biological activities, especially lncRNAs.3 Transcribed by RNA polymerase II, lncRNAs are transcripts that are greater than 200 nucleotides in length without functional open reading frame (ORF).4 In the past decades, accumulating studies illustrated that lncRNAs were pivotal regulatory molecules involved in cellular processes and drug resistance in multiple cancers.5,6 Therefore, it is an urgent need to explore biological function and clinical significance of these lncRNAs for more reliable clinical indicators and targets for cancer treatment.
LncRNA SNHG5, a transcript of small nucleolar RNA host gene 5 on 6q15, has been widely concerned as gathering studies revealed its potential role in human cancers.7,8 For instance, SNHG5 served as an oncogenic factor in glioma via promoting glucose uptake, migration, invasion in vitro and in-vivo tumorigenesis.9 While Zhao et al reported that a suppressive role of SNHG5 was detected in gastric cancer.10 Notably, it was also confirmed that SNHG5 exerted its oncogenic or suppressive effects on other human cancers including glioblastoma,11 endometrial cancer,12 acute myeloid leukemia,13 chronic myeloid leukemia,14 melanoma,15 colorectal cancer16 and osteosarcoma.17 Additional clinical features analysis revealed that SNHG5 had a close correlation with tumor stage and overall survival.18 Further functional experiments unfolded that SNHG5 could act as a sponge for miR-154-5p to elevate expression level of proliferating cell nuclear antigen (PCNA), thus markedly promoting proliferation.19 In addition, Wnt/β-catenin signaling, p38/MAPK signaling, mTOR signaling and ROCK signaling were significantly activated by SNHG5.11,13,17,20 Surprisingly, Zhao et al also verified that several biological molecules or proteins, such as metastasis-associated genes protein 2 (MTA2), spermatogenesis associated serine rich 2 (SPATS2), p27 and TRAIL-R1 (DR4) were involved in the functional process of SNHG5.10,21-23
In this review, we generalized the latest progress in the expression, function and regulation of lncRNA SNHG5 in multiple human cancers (Table 1–2) and further explored its potential role as a clinical biomarker and therapeutic target for cancers (Figure 1–3).
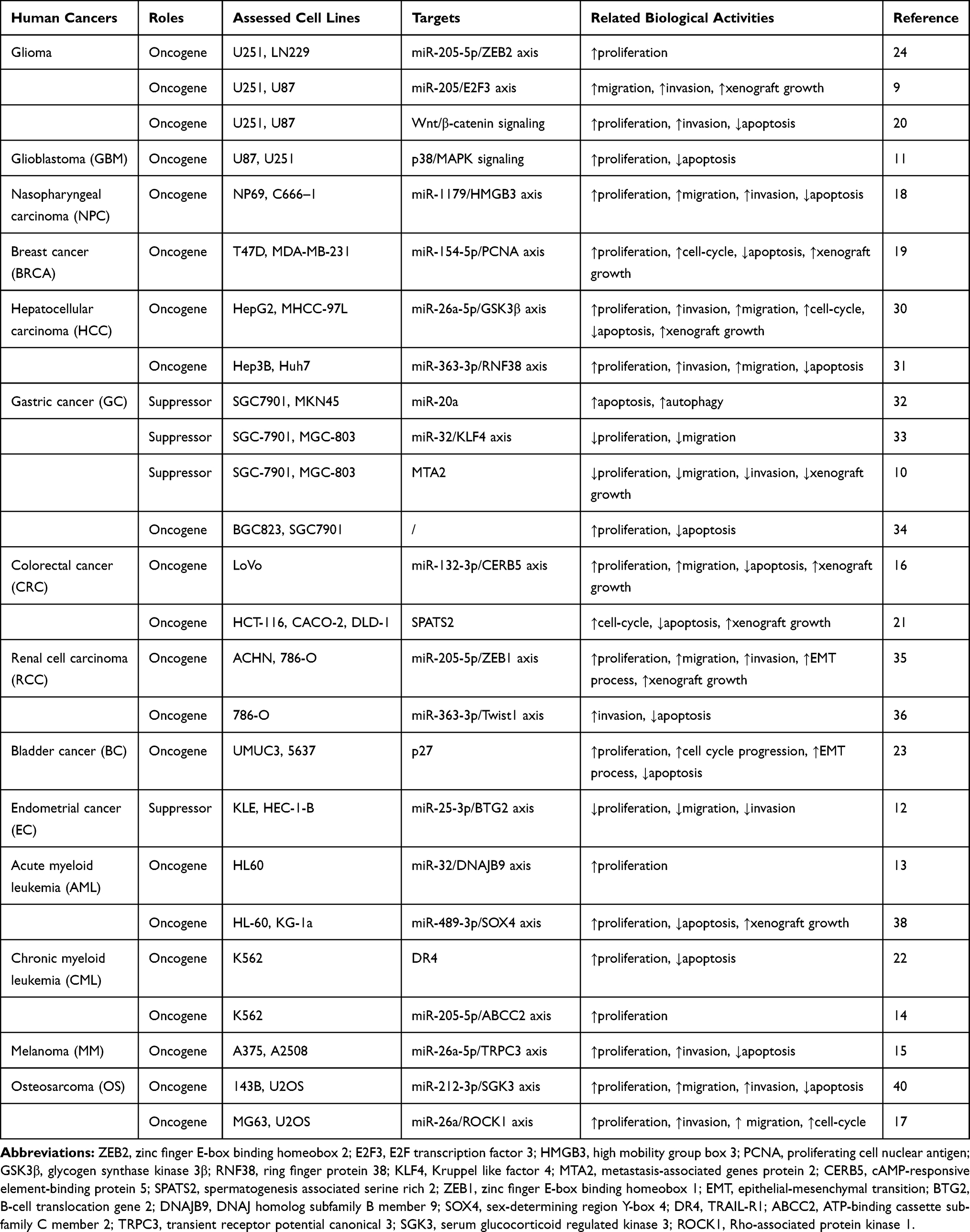 |
Table 1 Oncogenic or Suppressive Roles of lncRNA SNHG5 in Cancers |
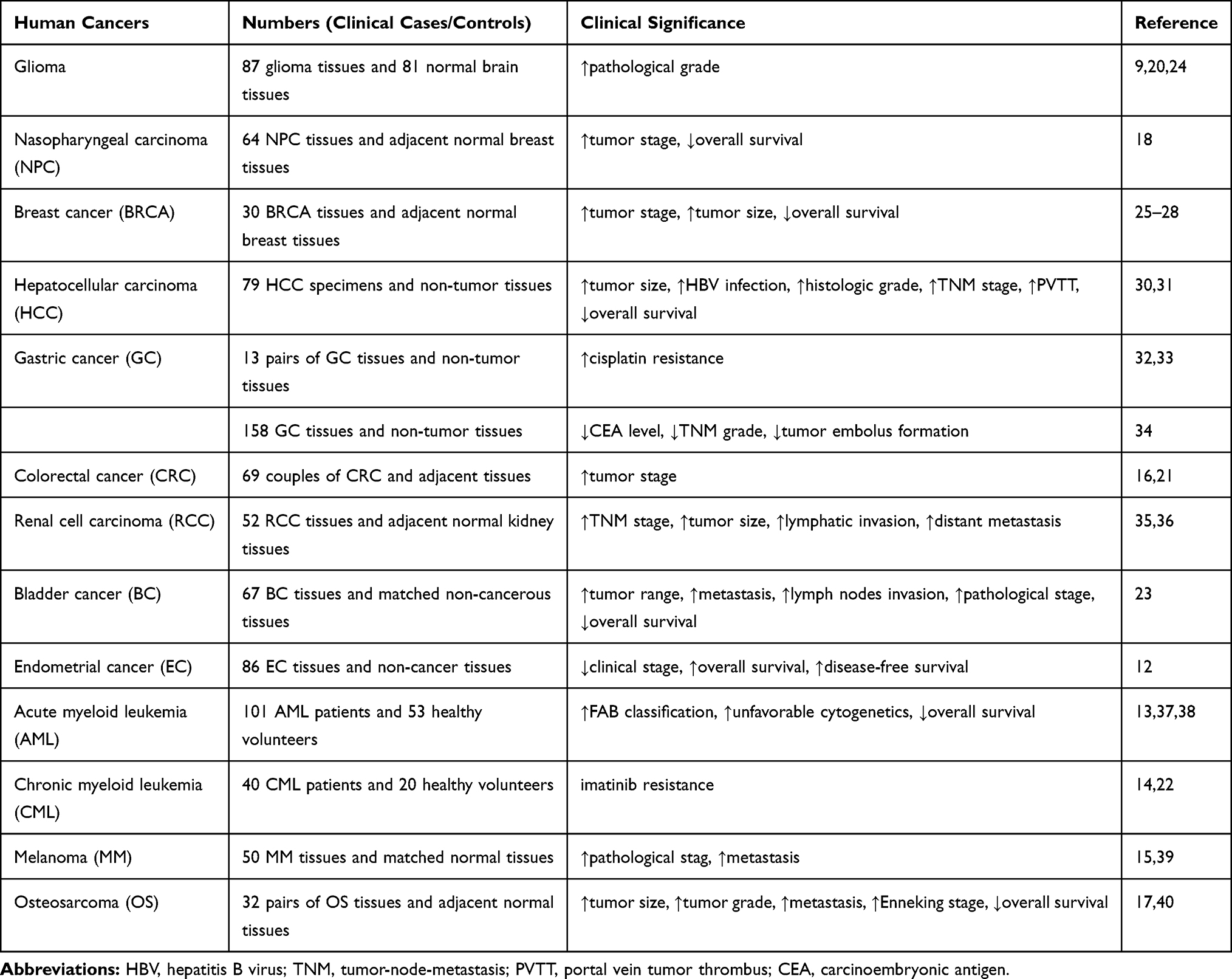 |
Table 2 Clinical Implications of lncRNA SNHG5 in Cancers |
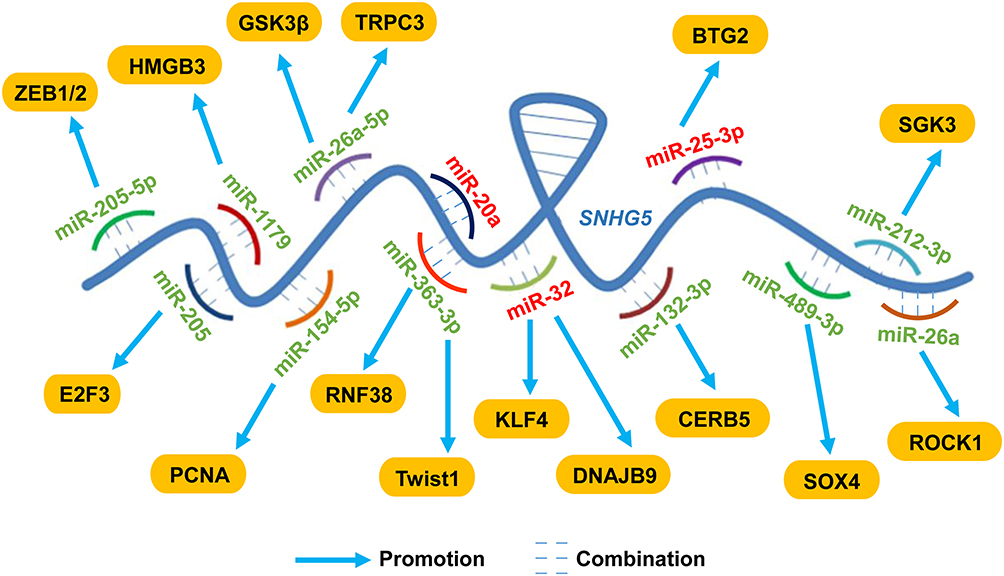 |
Figure 1 LncRNA SNHG5 exerted functional activities via sponging miRNAs in cancers LncRNA SNHG5 served as a ceRNA, directly interacting with miRNAs and regulating the production of mature miRNAs, thus participating in multiple biological activities. SNHG5 sponged miR-205-5p, miR-205, miR-1179, miR-154-5p, miR-26a-5p, miR-363-3p, miR-132-3p, miR-489-3p, miR-212-3p and miR-26a to exert oncogenic activities while sponging miR-20a, miR-32 and miR-25-3p to suppress tumor progression. Abbreviations: ZEB1/2, zinc finger E-box binding homeobox 1/2; E2F3, E2F transcription factor 3; HMGB3, high mobility group box 3; PCNA, proliferating cell nuclear antigen; GSK3β, glycogen synthase kinase 3β; TRPC3, transient receptor potential canonical 3; RNF38, ring finger protein 38; KLF4, Kruppel like factor 4; DNAJB9, DNAJ homolog subfamily B member 9; CERB5, cAMP-responsive element-binding protein 5; BTG2, B-cell translocation gene 2; SOX4, sex-determining region Y-box 4; SGK3, serum glucocorticoid regulated kinase 3; ROCK1, Rho-associated protein kinase 1. |
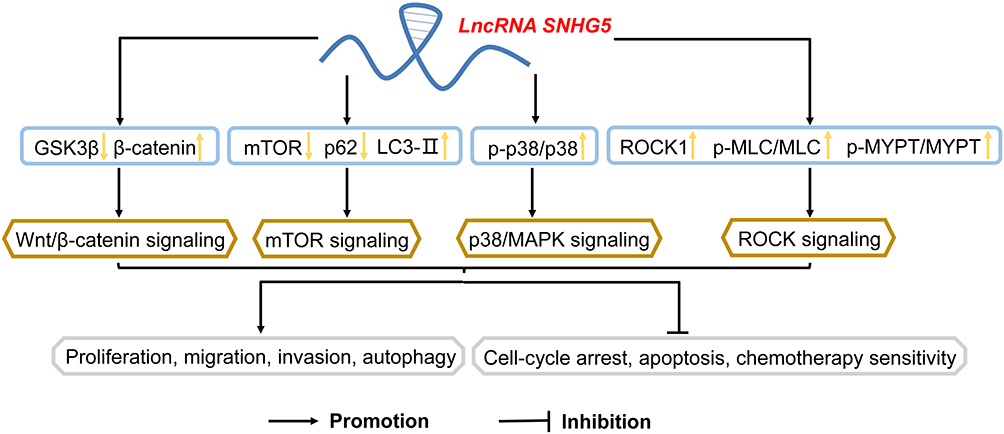 |
Figure 2 Involvement of signaling pathways by lncRNA SNHG5 in cancers LncRNA SNHG5 significantly activated Wnt/β-catenin signaling, p38/MAPK signaling, mTOR signaling and ROCK signaling, involved in various cellular activities including proliferation, migration, invasion, autophagy, cell-cycle, apoptosis and chemotherapy sensitivity. Abbreviations: GSK3β, glycogen synthase kinase 3β; mTOR, mammalian target of rapamycin; MAPK, mitogen-activated protein kinase; ROCK, Rho-associated protein kinase. |
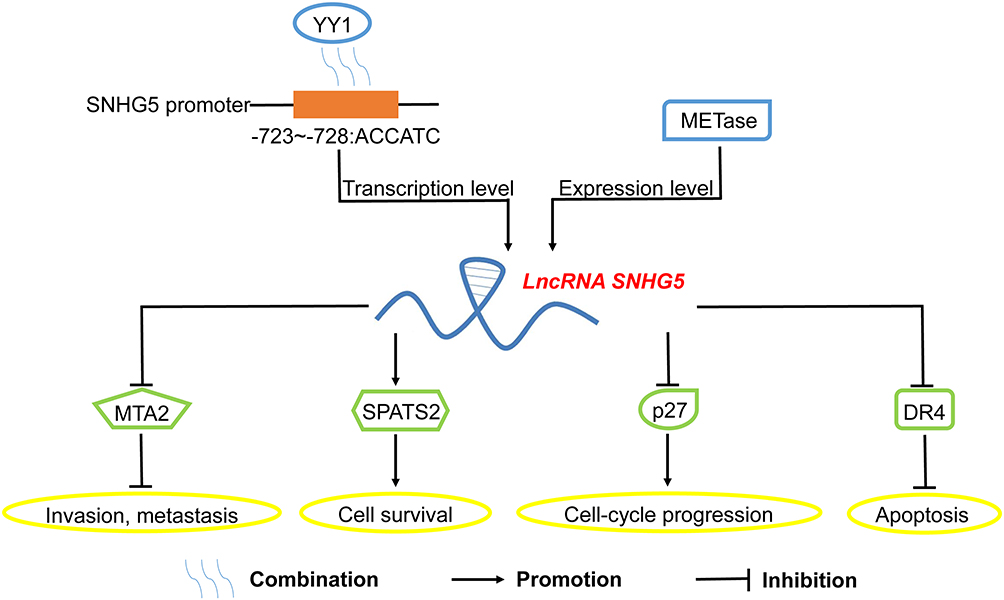 |
Figure 3 Interaction with biological molecules or proteins by SNHG5 in cancers LncRNA SNHG5 could be motivated by YY1 or METase at the transcription level or expression level respectively. Additionally, lncRNA SNHG5 interacted with MTA2, SPATS2, p27 and DR4, thus participating in the modulation of invasion, metastasis, survival, cell-cycle progression and apoptosis of tumor cells. Abbreviations: YY1, Yin-Yang 1; METase, L-methionine-α-deamino-γ-mercaptomethane-lyase; MTA2, metastasis-associated genes protein 2; SPATS2, spermatogenesis associated serine rich 2; DR4, TRAIL-R1. |
Aberrant Expression of lncRNA SNHG5 in Human Cancers
Glioma and Glioblastoma (GBM)
It has been reported that SNHG5 was high-expressed in 87 glioma tissues and four glioma cell lines (LN229, A172, U251 and U87) compared with corresponding normal tissues and NHAs.9,20,24 Subsequent analysis of clinical parameters revealed that high expression level of SNHG5 was positively correlated with advanced pathological grade (p < 0.01), and was independent of other parameters including age and gender of glioma patients (p > 0.01), suggesting that SNHG5 may be involved in tumorigenesis and progression of glioma.20 Accordingly, SNHG5 promoted glioma cell proliferation, migration, invasion and glucose uptake in vitro.9,24 Loss-of-function assay also verified that SNHG5 knockdown repressed malignant cellular phenotypes of glioma cells.20 Further in-vivo experiments disclosed that SNHG5 silencing significantly lowered the volume and weight of the tumors, which illustrated that SNHG5 knockdown efficaciously inhibited tumor growth.9 Mechanically, SNHG5 could serve as a competing endogenous RNA (ceRNA) to modulate the miR-205-5p/zinc finger E-box binding homeobox 2 (ZEB2) axis24 as well as miR-205/E2F transcription factor 3 (E2F3) axis9 in glioma cells. Wnt/β-catenin signaling also participated in the oncogenic activities of SNHG5.20 Interestingly, Chen et al confirmed that SNHG5 overexpression promoted GBM cellular proliferation and inhibited apoptosis via p38/MAPK signaling.11 However, the effects of SNHG5 on apoptotic response of glioma cells and xenograft growth of GBM cells are still uncertain. Taken together, these results have authenticated the correlation between the increased level of SNHG5 and malignant features of glioma, and partially indicated the same effects in GBM, which still needs further exploration.
Nasopharyngeal Carcinoma (NPC)
The gene detection performed by Liu et.al18 in 64 NPC tissues and two NPC cell lines (NP69 and C666–1) indicated that the expression level of SNHG5 was remarkably increased in NPC tissues and cell lines (p < 0.05). Clinically, the relative higher expression of SNHG5 was tightly associated with advanced tumor stage and shorter overall survival. In-vitro experiments unfolded that SNHG5 facilitated proliferation, migration and invasion in addition to suppressing apoptosis, which would be testified by additional in-vivo trials. Further mechanical exploration demonstrated that lncRNA SNHG5 promoted nasopharyngeal carcinoma progression by regulating miR-1179/high mobility group box 3 (HMGB3) axis. In summary, SNHG5 served as an oncogenic factor in the progression of NPC, which could be adopted as a promising predictor and therapeutic target.
Breast Cancer (BRCA)
Relative to normal ones, the expression level of SNHG5 was elevated in 30 BRCA tissues and cell lines including MDA-MB-231, MCF7, SKBR3, T47D, BT474 and BT20.25–27 Zahra et al also confirmed that SNHG5 was involved in cancer stage, overall grade, mitotic rate and tumor size in BRCA patients.28 Notably, SNHG5 was detected to correlate with positive reaction of estrogen receptor, progesterone receptor and Her2/neu expression,28 which were main subtypes of BRCA patients.29 Besides, SNHG5 knockdown inhibited proliferation and induced apoptosis, cell-cycle arrest at G1 phases in BRCA cells.19 Chi et al also investigated that SNHG5 overexpression facilitated BRCA cell growth in vivo.19 Further mechanical investigation uncovered that SNHG5 acted as a sponge for miR-154-5p to weaken the suppressive effect of miR-154-5p on PCNA, thus facilitating cell proliferation.19 In conclusion, SNHG5 could be utilized as an oncogene in BRCA, which may be a promising biomarker for different subtypes of BRCA patients.
Hepatocellular Carcinoma (HCC)
Compared to matched adjacent non-malignant tissues and human immortalized hepatic cell line THLE-2, SNHG5 expression was significantly elevated in 79 HCC tissues and four cell lines (HepG2, MHCC-97L Hep3B, Huh7).30,31 Accumulating clinical studies indicated that various clinicopathological parameters of HCC, such as tumor size, HBV infection, histologic grade, TNM stage and portal vein tumor thrombus (PVTT), were correlated to SNHG5 expression.30 Regarding the prognosis of patients with HCCs, Kaplan–Meier and log-rank analysis revealed that SNHG5 knockdown contributed to longer overall survival and lower recurrence rates of patients.30 In addition, SNHG5 overexpression was certified to promote proliferation, invasion, migration, cell-cycle progression and inhibit apoptosis in vitro, while SNHG5 knockdown had an opposite biological effects.30,31 Further in-vivo investigation showed the oncogenic role of SNHG5 in tumor growth and metastasis.30 Accordingly, SNHG5 boosted the progression of HCC via miR-26a-5p/glycogen synthase kinase-3β (GSK3β) axis30 or miR-363-3p/ring finger protein 38 (RNF38) axis.31 In brief, SNHG5 was of great clinical significance in diagnosis and treatment of patients with HCCs, which forecasted its potential clinical application.
Gastric Cancer (GC)
Compared to normal ones, the expression level of SNHG5 was downregulated in GC tissues and cell lines.32 Zhao et al reported that SNHG5 overexpression reduced cell proliferation, migration and invasion in GC cell lines including SGC-7901 and MGC-803.33 Similarly in vivo, the high expression of SNHG5 inhibited the growth and metastasis of tumors.10 Mechanically, SNHG5, upregulated by L-methionine-α-deamino-γ-mercaptomethane-lyase (METase), facilitated cell proliferation, migration and autophagy via sponging miR-20a and miR-32/Kruppel like factor 4 (KLF4) axis in GC.32,33 Moreover, MTA2, trapped by SNHG5 in the cytosol, was also involved in suppressive activities of GC.10 However, Li et al found that cisplatin-resistance was usually observed in GC patients, which was correlated with the high expression level of SNHG5.34 Clinically, cytotoxicity and apoptosis experiments revealed that lower expression of SNHG5 sensitizes GC cells to cisplatin.34 Taken together, the clinical significance of further investigations for SNHG5 has been demonstrated obviously in GC patients despite the discrepancy of these findings.
Colorectal Cancer (CRC)
It has been acknowledged that SNHG5 was evidently overexpressed in CRC tissues and cells.16 Interestingly, Nkerorema et al also investigated that SNHG5 was significantly upregulated both in adenomas and stage I carcinoma as compared with normal tissues, suggesting that SNHG5 overexpression acted as activator in CRC progression.21 Further in-vitro experiments revealed that the elevated expression of SNHG5 promoted proliferation, migration, metastasis, cell-cycle progression and inhibited apoptosis.16,21 In addition, SNHG5 exerted pro-tumor effects of facilitating the progression and growth of CRCs in vivo.16 Subsequent mechanical studies illustrated that SNHG5 promoted CRC cell survival by alleviating STAU1-induced degradation of SPATS2.21 Notably, Zhang et al also confirmed relevance of miR-132-3p/cAMP-responsive element binding protein 5 (CREB5) axis in the oncogenic process of SNHG5.16 However, the effects of SNHG5 on prognosis, clinical stage progression and overall survival of CRC patients are still obscure. In conclusion, SNHG5 positively affected progression of CRC tissues and cells, which may be an oncogene for CRC patients.
Renal Cell Carcinoma (RCC)
Concluding from the analysis from The Cancer Genome Atlas (TCGA) database and 72 pairs of human RCC tissues and non-tumor tissues, SNHG5 was significantly elevated in RCC tissues, which was also positively correlated with tumor-node-metastasis (TNM) stage, tumor size, lymphatic invasion, and distant metastasis.35,36 Similar high expression of SNHG5 was also detected in RCC cell lines, including ACHN, 786-O, SW13 and Caki-1.36 Subsequent in-vitro experiments disclosed that SNHG5 knockdown suppressed RCC cellular proliferation, migration and invasion.35 More importantly, SNHG5 was involved in the epithelial-mesenchymal transition (EMT) process and apoptotic response.35,36 In xenograft models of RCC cells, SNHG5 laid an obvious suppressive effect on tumor volume, weight and pulmonary metastatic nodules.35 Mechanically, SNHG5 targeted miR-205-5p/zinc finger E-box binding homeobox 1 (ZEB1) axis35 or miR-363-3p-Twist1 interaction36 to affect cellular invasion and apoptosis. In conclusion, SNHG5 served as an oncogene in the progression of RCC cells, which could be utilized as a novel predictor and therapeutic target.
Bladder Cancer (BC)
Ma et al investigated that the expression level of SNHG5 was aberrantly upregulated in 67 BC tissues and four cell lines, such as SW780, UMUC3, 5637 and T-24.23 Gathering clinical data demonstrated that SNHG5 overexpression was closely related to larger tumor range (p = 0.001), metastasis (p = 0.013), lymph nodes invasion (p = 0.001) and advanced pathological stage (p = 0.003).23 For assessment of prognosis, the elevated expression of SNHG5 contributed to shorter overall survival in patients with BCs via Kaplan–Meier method analysis (p < 0.001).23 Ma et al also reported that SNHG5 knockdown inhibited proliferation, EMT process and induced apoptosis, cell-cycle arrest at G1 phase.23 Meanwhile, it is an urgent demand for further investigation on xenograft models. Mechanically, SNHG5 exerted its oncogenic effects on BC via targeting p27.23 In short, these results provided evidence that SNHG5 could serve as a clinical biomarker for BC and could be promising therapeutic target, which still needs further in-vivo experiments.
Endometrial Cancer (EC)
Li et al characterized SNHG5 as a stable cytoplasmic lncRNA with downregulated expression in EC tumor tissues via the public tumor sequencing databases GEPIA and TIMER.12 The elevated expression of SNHG5 was also positively correlated with early clinical stage, improved overall survival and disease-free survival of EC patients.12 Furthermore, SNHG5 knockdown notably facilitated the proliferation, migration and invasion abilities of EC cells in vitro, whereas SNHG5 overexpression had the opposite effect.12 Mechanically, SNHG5 modulated malignant subtypes of EC via the miR-25-3p/B-cell translocation gene 2 (BTG2) axis.12 However, the effect of SNHG5 on tumor growth in vivo is still ambiguous, which needs additional studies. In summary, SNHG5 knockdown is strongly linked to poor prognosis, clinical stage progression and reduced overall survival of EC patients, which may be a diagnostic indicator or promising therapy target for EC patients in the future.
Acute Myeloid Leukemia (AML) and Chronic Myeloid Leukemia (CML)
Compared to 101 healthy controls, the expression level of SNHG5 was significantly upregulated in the bone marrow and plasma of 274 AML patients (p < 0.01).37,38 The elevated level of SNHG5 was more frequently found in AML patients with advanced FAB classification (p < 0.005) and unfavorable cytogenetics (p = 0.001).37 In addition, Kaplan-Meier analysis revealed that the high expression of SNHG5 was related to shorter overall survival for patients with AMLs.37,38 Additional in-vitro trials demonstrated that SNHG5 silencing repressed proliferation while inducing apoptosis.38 In vitro, the cytotoxicity tests have confirmed that the overexpression of SNHG5 promoted adriamycin (ADM) resistance in AML cells, while SNHG5 knockdown had an opposite influence.13 Interestingly, tumor volume and weight of xenograft models were also inhibited by SNHG5 knockdown.38 Mechanically, SNHG5 mainly targeted miR-489-3p/sex determining region Y-box 4 (SOX4) axis or miR-32/DNAJB9 axis to act as an oncogenic factor in AML.13,38 Similarly in CML cells, SNHG5 promoted imatinib resistance through evaluating cell viability.14 Further investigation demonstrated that inhibition of SNHG5 suppressed cell proliferation and induced differentiation, apoptosis of CML cells.22 Subsequent mechanical investigation showed that SNHG5 could serve as a ceRNA sponging miR-205-5p to regulate imatinib resistance.14 Interestingly, DR4, an oncogene in the occurrence and development of tumors, was also involved in functional process of SNHG5.22 In conclusion, these composite findings confirmed the imbalance of SNHG5 in AML and CML, forecasting its potential application in chemosensitivity.
Melanoma (MM)
Gao et al reported that the elevated expression level of SNHG5 was found in 50 MM tissues and cells (A375, SK-MEL-110, M14, MEL-RM, B16, and A2508).15 Clinically, the serum levels of SNHG5 in patients with MMs were upregulated obviously, suggesting that SNHG5 may be involved in genesis and metastasis of melanoma.39 Moreover, SNHG5 knockdown exerted tumor-suppressive function by suppressing cell proliferation, invasion and promoting apoptosis in all MM cell lines, whereas SNHG5 overexpression had an opposite effect.15 Notably, SNHG5 promoted growth and invasion of MM by regulating the miR-26a-5p/transient receptor potential canonical 3 (TRPC3) axis.15 In summary, SNHG5 may be an MM pro-tumor factor that is likely to be a prognostic factor and a potential therapeutic target, which still needs more investigations in vivo to verify.
Osteosarcoma (OS)
The expression level of SNHG5 was significantly higher in the OS tissues and cell lines including hFOB, U2OS, SW1353, HOS, Saos2 and MG63 than the normal ones.17 Clinically, the elevated SNHG5 expression was closely related to enhanced tumor size (p = 0.013), higher tumor grade (p = 0.012), increased metastasis (p = 0.012) and elevated Enneking stage (p = 0.015),17 which was also correlated with a poor diagnosis in patients with OS.40 As for biological function, SNHG5 knockdown suppressed cell proliferation, invasion, migration, and induced apoptosis, cell-cycle arrest at G0/G1 in vitro, whereas SNHG5 overexpression had the opposite influence.17,40 Mechanically, SNHG5 strengthened the tumorigenesis of OS by sponging the miR-212-3p/serum glucocorticoid regulated kinase 3 (SGK3) axis40 as well as miR-26a/Rho-associated protein kinase 1 (ROCK1) axis.17 Above all, these studies testified that SNHG5 acted as an oncogene in OSs and could provide a target for the treatment and diagnosis of OS patients (Table 1–2).
Biological Mechanisms of lncRNA SNHG5 in Human Cancers
Interaction with miRNAs
Accumulating studies have illustrated that lncRNAs could serve as ceRNAs, which may directly interact with miRNAs and regulate the production of mature miRNAs, thus participating in multiple biological activities.41 For example, SNHG5 served as a ceRNA for miR-205-5p to elevate expression of ZEB2 in facilitating glioma progression.24 Li et al also reported that SNHG5 exerted the same effects in glioma via sponging miR-205 to elevate expression of E2F3.9 In nasopharyngeal carcinoma, SNHG5 promoted tumor progression by regulating miR-1179/HMGB3 axis.18 Moreover, SNHG5 targeted miR-154-5p to weaken the suppressive effect of miR-154-5p on PCNA, which was significantly involved in proliferation of breast cancer cells.19 Furthermore, miR-26a-5p, which markedly suppressed GSK3β, was inversely correlated with SNHG5 in hepatocellular carcinoma.30 In hepatocellular carcinoma, SNHG5 acted as an oncogene by regulating miR-363-3p/RNF38 axis.31 Xin et al also found that SNHG5 had a negative correlation with miR-20a to boost cellular autophagy in gastric cancer.32 In gastric cancer cells, SNHG5 suppressed proliferation and migration via regulating KLF4 expression in a miR-32-dependent manner.33 In addition, SNHG5 downregulated miR-132-3p to boost CREB5 expression in colorectal cancer.16
Similar roles of SNHG5 were also investigated in other solid tumors. In renal cell carcinoma, SNHG5 participated in the initiation and development targeting miR-205-5p/ZEB1 axis.35 Similarly, a study by Li et al confirmed that SNHG5 affected the invasion and apoptosis via miR-363-3p-Twist1 interaction in renal cell carcinoma.36 In endometrial cancer cells, SNHG5 played a role as tumor inhibitor through upregulation of B-cell translocation gene 2 (BTG2) by suppressing miR-25-3p.12 Besides, SNHG5 positively regulated DNAJB9 through miR-32 to promote ADM resistance of acute myeloid leukemia cells.13 Interestingly, SNHG5 also regulated SOX4 expression through competitive combination with miR-489-3p in acute myeloid leukemia.38 Similarly in chronic myeloid leukemia, ATP-binding cassette sub-family C member 2 (ABCC2), one of the ATP-binding cassette transporters, was dramatically upregulated by SNHG5 targeting miR-205-5p, thus promoting imatinib resistance.14 Additionally, SNHG5 could facilitate TRPC3 expression via sponging miR-26a-5p in melanoma.15 In osteosarcoma, SNHG5 downregulated miR‑212‑3p to elevate SGK3,40 and activated ROCK1 expression targeting miR-26a17 (Figure 1).
Involvement in Signaling Pathways
To our surprise, SNHG5 has been reported to be relevant to several signaling pathways, which were involved in various aspects during tumorigenesis.42 The elevated expression of SNHG5 reduced the expression of GSK3β and upregulated the expression of β-catenin, thus activating Wnt/β-catenin signaling pathway in glioma.20 Similarly, SNHG5 markedly facilitated GSK3β expression targeting miR-26a-5p through Wnt/β-catenin signaling pathway in hepatocellular carcinoma.30 In glioblastoma, p38/MAPK signaling pathway was activated by the elevation of p-p38/p38, ELK1 and STAT1, which was positively related to SNHG5.11 Wang et al also found that DNAJB9, elevated by SNHG5, resulted in a decrease in mTOR and p62 expression, with increased LC3B expression, via mTOR signaling pathway in acute myeloid leukemia.13 Moreover, SNHG5 promoted ROCK1 expression and further activated the ROCK signaling pathway through upregulation of p-MLC and p-MYPT in osteosarcoma17 (Figure 2).
Other Biological Molecules
In gastric cancer, SNHG5 promoted MTA2 trapped in the cytosol.10 In addition, STAU1 could be alleviated by SNHG5, promoted colorectal cancer cells progression by degradation of SPATS2.21 Ma et al reported that the effect of SNHG5 on bladder cancer cells was partially involved with targeting p27.23 Moreover, the methylation and downregulation of DR4 could also be induced by SNHG522 (Figure 3).
Conclusions and Future Perspectives
Many lncRNAs are certified to exhibit low inter-species homology and high tissue-specific signature, manifesting the significance of lncRNAs in cellular differentiation and development in human cancers.43 Moreover, Abbasifard et al also found that lncRNAs were involved in multiple pathological processes, including extracellular matrix (ECM) degradation, inflammatory responses, apoptosis and angiogenesis.44 Recently, accumulating efforts have been made to investigate the clinical significance of lncRNAs in human cancers.45 All the above findings may provide reliable and comprehensive evidence for clinical application of lncRNAs.
SNHG5, a relatively novel lncRNA, was attached to more and more attention due to its distinct biological and clinical effects in human cancers. Concluding from current clinical investigation results, SNHG5 overexpression was detected in multiple human cancers, including glioma,20,24 acute myeloid leukemia,37 breast cancer,25,28 melanoma,15,39 colorectal cancer,16,21 osteosarcoma,17,40 hepatocellular carcinoma30 and bladder cancer.23 In these cancers, SNHG5 could serve as an oncogene in tumor size, pathological stage, recurrence rate and overall survival. Conversely, SNHG5 exerted an opposite influence in endometrial cancer, of which the overexpression was correlated with early clinical stage, improved overall survival and disease-free survival.12 In addition, the elevated expression of SNHG5 contributed to chemotherapy in acute myeloid leukemia and chronic myeloid leukemia,13,14 which was consistent with cisplatin-resistant patients in gastric cancers.34 Taken together, these results have confirmed the significance of further application for SNHG5 in human cancers.
Meanwhile, there were some overlaps on the role of SNHG5 in particular tumors. For example, miRNA-32,13,33 miR-26a-5p15,30 and miR-363-3p31,36 were involved in functional processes of SNHG5 in two or more cancers, suggesting that the three miRNAs may be specific receptors of SNHG5, which could also be investigated in other tumors. Moreover, Wnt/β-catenin signaling pathway could be activated by SNHG5 directly or indirectly,20,30 indicating its significance in biological activities of SNHG5. However, overlaps were accompanied by some conflicts deserving additional studies. It was reported that in gastric cancer, SNHG5 suppressed proliferation, migration, invasion in vitro and inhibited the tumor growth and metastasis in vivo.10,33 While Li et al found that SNHG5 acted as an oncogene in facilitating proliferation and inhibiting cell apoptosis, thus promoting gastric cancer resistance to cisplatin.34 We suppose that the main reasons for this differential expression of SNHG5 may result from gene copy variation (deletion or amplification), transcription factors, histone modifications or DNA methylation. For example, the heterogeneity that tumors vary in gene expression and genetic alterations could assume the responsibility. Moreover, in the latter experiment, the tumor microenvironment may be challenged by the chemotherapy drug, thus inducing the differential expression. The two hypotheses would provide directions for further exploration of SNHG5 and propose a demand for novel RNA sequencing methods.
To our surprise, some upstream transcriptional regulators of SNHG5 were also detected in recent years. Yin-Yang 1 (YY1), a transcription factor ubiquitously expressed in all human tissues, could induce the overexpression of SNHG5, thus facilitating progression of GBM. Subsequent ChIP assay results disclosed that YY1 bound with specific sites (−723~-728: ACCATC) in the promoter of SNHG5, which might accelerate the transcription rate.11 Xin et al also explored that SNHG5 could be significantly upregulated by METase, while the related functional mechanism was still ambiguous.32 Nevertheless, there are still several limitations with current investigation. Firstly, the role of SNHG5 in some known cancers, including glioblastoma and acute myeloid leukemia, is still blurred, which needs more experiments in vitro and in vivo. Secondly, the functions and mechanisms of SNHG5 in other cancers should be detected. Last but not least, subsequent exploration of upstream transcriptional regulators for SNHG5 is urgently needed.
In conclusion, SNHG5 has been verified its role in cellular activities and clinical practices in human cancers via miRNAs, signaling pathways and biological molecules or proteins, which may be promising clinical biomarker and therapeutic target for cancer treatment.
Abbreviations
lncRNA, long non-coding RNA; SNHG5, small nucleolar RNA host gene 5; ncRNA, non-coding RNA; miRNA, microRNA; siRNA, small interfering RNA; tRNA, transfer RNA; rRNA, ribosomal RNA; snRNA, small nuclear RNA; snoRNA, small nucleolar RNA; ORF, open reading frame; PCNA, proliferating cell nuclear antigen; MTA2, metastasis-associated genes protein 2; SPATS2, spermatogenesis associated serine rich 2; DR4, TRAIL-R1; GBM, glioblastoma; ceRNA, competing endogenous RNA; ZEB2, zinc finger E-box binding homeobox 2; E2F3, E2F transcription factor 3; NPC, nasopharyngeal carcinoma; HMGB3, high mobility group box 3; BRCA, breast cancer; HCC, hepatocellular carcinoma; PVTT, portal vein tumor thrombus; GSK3β, glycogen synthase kinase 3β; RNF38, ring finger protein 38; GC, gastric cancer; METase, L-methionine-α-deamino-γ-mercaptomethane-lyase; KLF4, Kruppel like Factor 4; CRC, colorectal cancer; CREB5, cAMP-responsive element binding protein 5; RCC, renal cell carcinoma; TCGA, The Cancer Genome Atlas; TNM, tumor-node-metastasis; EMT, epithelial-mesenchymal transition; ZEB1, zinc finger E-box binding homeobox 1; BC, bladder cancer; EC, endometrial cancer; BTG2, B-cell translocation gene 2; AML, acute myeloid leukemia; CML, chronic myeloid leukemia; ADM, adriamycin; SOX4, sex determining region Y-box 4; MM, melanoma; TRPC3, transient receptor potential canonical 3; OS, osteosarcoma; SGK3, serum glucocorticoid regulated kinase 3; ROCK1, Rho-associated protein kinase 1; ABCC2, ATP-binding cassette sub-family C member 2; ECM, extracellular matrix; YY1, Yin-Yang 1.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; agree to be accountable for all aspects of the work.
Funding
This study was funded by the Changzhou Health and Family Planning Commission Youth Talent Science and Technology Project (WZ201811).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Hu Q, Tai S, Wang J. Oncogenicity of lncRNA FOXD2-AS1 and its molecular mechanisms in human cancers. Pathol Res Pract. 2019;215(5):843–848. doi:10.1016/j.prp.2019.01.033
2. Xin Y, Li Z, Shen J, Chan MT, Wu WK. CCAT1: a pivotal oncogenic long non-coding RNA in human cancers. Cell Prolif. 2016;49(3):255–260. doi:10.1111/cpr.12252
3. Li Y, Han X, Feng H, Han J. Long noncoding RNA OIP5-AS1 in cancer. Clin Chim Acta. 2019;499:75–80. doi:10.1016/j.cca.2019.08.031
4. Wang DQ, Fu P, Yao C, et al. Long non-coding RNAs, novel culprits, or bodyguards in neurodegenerative diseases. Mol Ther Nucleic Acids. 2018;10:269–276. doi:10.1016/j.omtn.2017.12.011
5. Jin X, Liu X, Zhang Z, Guan Y. lncRNA CCAT1 acts as a MicroRNA-218 sponge to increase gefitinib resistance in NSCLC by targeting HOXA1. Mol Ther Nucleic Acids. 2020;19:1266–1275. doi:10.1016/j.omtn.2020.01.006
6. Ren X, Chen C, Luo Y, et al. lncRNA-PLACT1 sustains activation of NF-kappaB pathway through a positive feedback loop with IkappaBalpha/E2F1 axis in pancreatic cancer. Mol Cancer. 2020;19(1):35. doi:10.1186/s12943-020-01153-1
7. Lin H, Shen L, Lin Q, et al. SNHG5 enhances Paclitaxel sensitivity of ovarian cancer cells through sponging miR-23a. Biomed Pharmacother. 2020;123:109711. doi:10.1016/j.biopha.2019.109711
8. Nakamura Y, Takahashi N, Kakegawa E, et al. The GAS5 (growth arrest-specific transcript 5) gene fuses to BCL6 as a result of t(1;3)(q25;q27) in a patient with B-cell lymphoma. Cancer Genet Cytogenet. 2008;182(2):144–149. doi:10.1016/j.cancergencyto.2008.01.013
9. Li X, Liu L, Luo Y, Cui S, Chen W, Zeng A, Shi Y, Luo L. Long non-coding RNA SNHG5 promotes glioma progression via miR-205/E2F3 axis. Biosci Rep. 2019;39(7):7. doi:10.1042/BSR20190668
10. Zhao L, Guo H, Zhou B, et al. Long non-coding RNA SNHG5 suppresses gastric cancer progression by trapping MTA2 in the cytosol. Oncogene. 2016;35(44):5770–5780. doi:10.1038/onc.2016.110
11. Chen L, Gong X, Huang M. YY1-activated long noncoding RNA SNHG5 promotes glioblastoma cell proliferation through p38/MAPK signaling pathway. Cancer Biother Radiopharm. 2019;34(9):589–596. doi:10.1089/cbr.2019.2779
12. Li S, Shan Y, Li X, et al. lncRNA SNHG5 modulates endometrial cancer progression via the miR-25-3p/BTG2 axis. J Oncol. 2019;2019:7024675. doi:10.1155/2019/7024675
13. Wang D, Zeng T, Lin Z, et al. Long non-coding RNA SNHG5 regulates chemotherapy resistance through the miR-32/DNAJB9 axis in acute myeloid leukemia. Biomed Pharmacother. 2020;123:109802. doi:10.1016/j.biopha.2019.109802
14. He B, Bai Y, Kang W, Zhang X, Jiang X. LncRNA SNHG5 regulates imatinib resistance in chronic myeloid leukemia via acting as a CeRNA against MiR-205-5p. Am J Cancer Res. 2017;7(8):1704–1713.
15. Gao J, Zeng K, Liu Y, Gao L, Liu L. LncRNA SNHG5 promotes growth and invasion in melanoma by regulating the miR-26a-5p/TRPC3 pathway. Onco Targets Ther. 2019;12:169–179. doi:10.2147/OTT.S184078
16. Zhang M, Li Y, Wang H, Yu W, Lin S, Guo J. LncRNA SNHG5 affects cell proliferation, metastasis and migration of colorectal cancer through regulating miR-132-3p/CREB5. Cancer Biol Ther. 2019;20(4):524–536. doi:10.1080/15384047.2018.1537579
17. Wang Z, Wang Z, Liu J, Yang H. Long non-coding RNA SNHG5 sponges miR-26a to promote the tumorigenesis of osteosarcoma by targeting ROCK1. Biomed Pharmacother. 2018;107:598–605. doi:10.1016/j.biopha.2018.08.025
18. Liu D, Wang Y, Zhao Y, Gu X. LncRNA SNHG5 promotes nasopharyngeal carcinoma progression by regulating miR-1179/HMGB3 axis. BMC Cancer. 2020;20(1):178. doi:10.1186/s12885-020-6662-5
19. Chi JR, Yu ZH, Liu BW, Zhang D, Ge J, Yu Y, Cao XC. SNHG5 promotes breast cancer proliferation by sponging the miR-154-5p/PCNA axis. Mol Ther Nucleic Acids. 2019;17:138–149. doi:10.1016/j.omtn.2019.05.013
20. Hu X, Hong Y, Shang C. Knockdown of long non-coding RNA SNHG5 inhibits malignant cellular phenotypes of glioma via Wnt/CTNNB1 signaling pathway. J Cancer. 2019;10(5):1333–1340. doi:10.7150/jca.29517
21. Damas ND, Marcatti M, Come C, et al. SNHG5 promotes colorectal cancer cell survival by counteracting STAU1-mediated mRNA destabilization. Nat Commun. 2016;7:13875. doi:10.1038/ncomms13875
22. Gao B, Li S, Li G. Long noncoding RNA (lncRNA) small nucleolar RNA Host Gene 5 (SNHG5) regulates proliferation, differentiation, and apoptosis of K562 cells in chronic myeloid leukemia. Med Sci Monitor. 2019;25:6812–6819. doi:10.12659/MSM.916661
23. Ma Z, Xue S, Zeng B, Qiu D. LncRNA SNHG5 is associated with poor prognosis of bladder cancer and promotes bladder cancer cell proliferation through targeting p27. Oncol Lett. 2018;15(2):1924–1930. doi:10.3892/ol.2017.7527
24. Meng X, Deng Y, Lv Z, et al. LncRNA SNHG5 promotes proliferation of glioma by regulating miR-205-5p/ZEB2 axis. Onco Targets Ther. 2019;12:11487–11496. doi:10.2147/OTT.S228439
25. Sterbova M, Pazourkova E, Santorova-Pospisilova S, Zednikova I, Tesarova P, Korabecna M. The use of human inflammatory response and autoimmunity RT2 lncRNA PCR array for plasma examination in breast cancer patients prior to therapy. Neoplasma. 2019;66(4):641–646. doi:10.4149/neo_2018_180907N679
26. Lee J, Park HY, Kim WW, Lee SJ, Jeong JH, Kang SH, Jung JH, Chae YS. Biological function of long noncoding RNA snaR in HER2-positive breast cancer cells. Tumour Biol. 2017;39(6):1010428317707374. doi:10.1177/1010428317707374
27. Lee J, Jung JH, Chae YS, et al. Long noncoding RNA snaR regulates proliferation, migration and invasion of triple-negative breast cancer cells. Anticancer Res. 2016;36(12):6289–6295. doi:10.21873/anticanres.11224
28. Taherian-Esfahani Z, Taheri M, Dashti S, Kholghi-Oskooei V, Geranpayeh L, Ghafouri-Fard S. Assessment of the expression pattern of mTOR-associated lncRNAs and their genomic variants in the patients with breast cancer. J Cell Physiol. 2019;234(12):22044–22056. doi:10.1002/jcp.28767
29. Lee E, Choi A, Jun Y, Kim N, Yook JI, Kim SY, Lee S, Kang SW.. Glutathione peroxidase-1 regulates adhesion and metastasis of triple-negative breast cancer cells via FAK signaling. Redox Biol. 2020;29:101391. doi:10.1016/j.redox.2019.101391
30. Li Y, Guo D, Zhao Y, et al. He S and Lu X. Long non-coding RNA SNHG5 promotes human hepatocellular carcinoma progression by regulating miR-26a-5p/GSK3beta signal pathway. Cell Death Dis. 2018;9(9):888. doi:10.1038/s41419-018-0882-5
31. Hu PA, Miao YY, Yu S, Guo N. Long non-coding RNA SNHG5 promotes human hepatocellular carcinoma progression by regulating miR-363-3p/RNF38 axis. Eur Rev Med Pharmacol Sci. 2020;24(7):3592–3604. doi:10.26355/eurrev_202004_20821
32. Xin L, Zhou LQ, Liu L, Yuan YW, Zhang HT, Zeng F. METase promotes cell autophagy via promoting SNHG5 and suppressing miR-20a in gastric cancer. Int J Biol Macromol. 2019;122:1046–1052. doi:10.1016/j.ijbiomac.2018.09.051
33. Zhao L, Han T, Li Y, et al. The lncRNA SNHG5/miR-32 axis regulates gastric cancer cell proliferation and migration by targeting KLF4. FASEB J. 2017;31(3):893–903. doi:10.1096/fj.201600994R
34. Li M, Zhang YY, Shang J, Xu YD. LncRNA SNHG5 promotes cisplatin resistance in gastric cancer via inhibiting cell apoptosis. Eur Rev Med Pharmacol Sci. 2019;23(10):4185–4191. doi:10.26355/eurrev_201905_17921
35. Xiang W, Lv L, Zhou G, Wu W, Yuan J, Zhang C, Jiang G. The lncRNA SNHG5-mediated miR-205-5p downregulation contributes to the progression of clear cell renal cell carcinoma by targeting ZEB1. Cancer Med. 2020. doi:10.1002/cam4.3052
36. Li WZ, Zou Y, Song ZY, Wei ZW, Chen G. Long non-coding RNA SNHG5 affects the invasion and apoptosis of renal cell carcinoma by regulating the miR-363-3p-Twist1 interaction. Am J Transl Res. 2020;12(2):697–707.
37. Li J, Sun CK. Long noncoding RNA SNHG5 is up-regulated and serves as a potential prognostic biomarker in acute myeloid leukemia. Eur Rev Med Pharmacol Sci. 2018;22(11):3342–3347. doi:10.26355/eurrev_201806_15154
38. Ying X, Zhang W, Fang M, Wang C, Han L, Yang C. LncRNA SNHG5 regulates SOX4 expression through competitive binding to miR-489-3p in acute myeloid leukemia. Inflammation Res. 2020;69(6):607–618. doi:10.1007/s00011-020-01345-x
39. Ichigozaki Y, Fukushima S, Jinnin M, et al. Serum long non-coding RNA, snoRNA host gene 5 level as a new tumor marker of malignant melanoma. Exp Dermatol. 2016;25(1):67–69. doi:10.1111/exd.12868
40. Ju C, Zhou R, Sun J, et al. LncRNA SNHG5 promotes the progression of osteosarcoma by sponging the miR-212-3p/SGK3 axis. Cancer Cell Int. 2018;18:141. doi:10.1186/s12935-018-0641-9
41. Wang J, Cao Y, Lu X, et al. Identification of the regulatory role of lncRNA SNHG16 in myasthenia gravis by constructing a competing endogenous RNA network. Mol Ther Nucleic Acids. 2020;19:1123–1133. doi:10.1016/j.omtn.2020.01.005
42. Janghorban M, Xin L, Rosen JM, Zhang XH. Notch signaling as a regulator of the tumor immune response: to target or not to target? Front Immunol. 2018;9:1649. doi:10.3389/fimmu.2018.01649
43. Abildgaard C, Do Canto LM, Steffensen KD, Rogatto SR. Long non-coding RNAs involved in resistance to chemotherapy in ovarian cancer. Front Oncol. 2019;9:1549. doi:10.3389/fonc.2019.01549
44. Abbasifard M, Kamiab Z, Sadeghi I, Bagheri-Hosseinabadi Z. The role and function of long non-coding RNAs in osteoarthritis. Exp Mol Pathol. 2020;104407. doi:10.1016/j.yexmp.2020.104407
45. Meseure D, Drak Alsibai K, Nicolas A, Bieche I, Morillon A. Long noncoding RNAs as new architects in cancer epigenetics, prognostic biomarkers, and potential therapeutic targets. Biomed Res Int. 2015;2015:320214. doi:10.1155/2015/320214
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.