Back to Journals » Biologics: Targets and Therapy » Volume 18
Lactobacillus Protects Against Chronic Suppurative Otitis Media via Modulating RFTN1/ Lipid Raft /TLR4-Mediated Inflammation
Authors Liu Z, Zhang F, Jia F, Li S, Jiang C, Ruan B, Long R
Received 5 September 2024
Accepted for publication 10 December 2024
Published 27 December 2024 Volume 2024:18 Pages 453—468
DOI https://doi.org/10.2147/BTT.S484410
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shein-Chung Chow
Zhuohui Liu,* Fan Zhang,* Fengfeng Jia, Shuling Li, Chaowu Jiang, Biao Ruan, Ruiqing Long
Department of Otolaryngology, The First Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ruiqing Long, Department of Otolaryngology, The First Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, People’s Republic of China, Email [email protected]
Purpose: Chronic suppurative otitis media (CSOM) is a prominent contributor to preventable hearing loss globally. Probiotic therapy has attracted research interest in human infectious and inflammatory disease. As the most prevalent probiotic, the role of Lactobacillus in CSOM remains poorly defined. This study aimed to investigate the antipathogenic effects and underlying mechanism of Lactobacillus on CSOM.
Methods: RNA sequencing of granulation of middle ear cavity from CSOM patients and lavage fluid of middle ear from normal volunteer was conducted. Human middle ear epithelial cells (HMEEC) and rats infected with Bacillus cereus (B. cereus) and Staphylococcus aureus (S. aureus) were used for CSOM constructing. Western blot, qPCR and Vybrant™ Alexa Fluor™ 488 lipid raft labeling were performed to explore the possible molecular mechanism by which lipid raft linker (RFTN1) regulates lipid raft/toll-like receptor 4 (TLR4). ELISA and HE staining was utilized to evaluate the effect of Lactobacillus on the progression of CSOM in vivo.
Results: Based on RNA Sequence analysis, a total of 3646 differentially expressed genes (1620 up-regulated and 2026 down-regulated) were identified in CSOM. RFTN1 was highly expressed in CSOM. Inhibition of RFTN1 not only reduced the inflammatory response of CSOM but also suppressed the formation of lipid rafts. Further investigation revealed that RFTN1 inhibition could reduce the expression of TLR4, which also localizes to the lipid rafts. TLR4 responds to RFTN1-mediated inflammatory responses in CSOM. We treated the CSOM model with Lactobacillus, which has great potential for alleviating the inflammatory response, and found that Lactobacillus attenuated the development of CSOM by reducing RFTN1 and TLR4 expression.
Conclusion: In conclusion, these findings suggest a crucial role for Lactobacillus in alleviating CSOM progression and uncovered the molecular mechanism involving Lactobacillus-regulated inhibition of the RFTN1-lipid raft-TLR4 signaling pathway under CSOM conditions.
Keywords: CSOM, Lactobacillus, RFTN1, TLR4, S. aureus, B. cereus
Introduction
Chronic suppurative otitis media (CSOM) is a prominent contributor to preventable hearing loss globally. Once it sets in, patients present with suppurative lesions in the mucosa and submucosa of the middle ear, and in severe cases, may even involve the bone, resulting in tympanic membrane perforation and long-term purulent symptoms in the ear. Characterized by a prolonged disease course and persistence of chronic illness, CSOM exerts a severe impact on patients’ daily lives and poses a threat to their health, consistently presenting a challenging issue for otolaryngologists.1 CSOM is regarded as a multifactorial illness stemming from a series of complex interactions involving the environment, bacteria, hosts, and genetic risk factors. Bacterial infection is the primary cause of CSOM, with coagulase-negative Staphylococcus, Pseudomonas aeruginosa, Staphylococcus aureus (S. aureus), and Proteus mirabilis being the most prevalent pathogens in CSOM.2,3 Thus, animals or cells inoculation with bacterial were used for CSOM model conduction.4,5 It is well established that S. aureus and Bacillus cereus (B. cereus) are critical foodborne pathogens in terms of frequency and seriousness of the disease. S. aureus is the most predominant pathogens that cause CSOM6 and B. cereus is a major cause of post-traumatic endophthalmitis7 and could be observed in calves with otitis media (OM).8 Currently, first-line treatment for uncomplicated CSOM, encompassing antibiotics and anti-inflammatory drugs, is the most effective approach, with minimal side effects.9 Nevertheless, the abuse of antibiotics in recent years, particularly their non-standard utilization, may give rise to certain variations in pathogenic bacteria and their susceptibility to antibiotics.3 Therefore, the discovery of novel therapies for CSOM is crucial.
Probiotics are defined as follows “live microorganisms that, when administered in adequate amounts, serve a health benefit on the host” according to the World Health Organization (WHO). Probiotics participate in regulating the microbial balance and ameliorate diseases stemming from microbial community imbalance, especially in the intestinal mucosa.10 For over a decade, probiotics have been successfully used to treat common infectious diseases of the digestive tract.11 It is indicated that probiotics showed strong antibacterial activity against gram-positive bacterial including S. aureus and B. cereus.12,13 In studies on OM, Scott and Coleman discovered that probiotic therapy holds considerable promise for alleviating OM. They demonstrated its ability to inhibit pathogen growth in the ear canal in vitro, suggesting its potential to offer a more effective alternative to antibiotics in the future and warranting further exploration.14,15 Lactobacillus have become one of the commonly used probiotics.16 Accumulating evidence suggested that Lactobacillus displayed antibacterial activity against S. aureus and B. cereus.17,18 In addition, the protective role of Lactobacillus in CSOM has been characterized.19 However, the underlying mechanism by which Lactobacillus improve S. aureus and B. cereus-induced CSOM remains unknown.
At present, investigations into the pathogenic biology of OM not only focus on the disparity of pathogenic bacteria among distinct types of OM but also on the inflammatory mechanisms of bacteria, such as the different toxins produced by various pathogenic bacteria or the release of specific inflammatory factors. Lipid rafts are microstructural domains rich in cholesterol and sphingomyelin on the plasma membrane with a size of approximately 70 nm. They are dynamic structures localized on the outer leaflets of the plasma membrane. Lipid rafts function as platforms anchored to proteins and are closely related to messenger formation, synaptic transmission, transmembrane transport, membrane signal transduction, protein sorting, cell proliferation, and apoptosis. Lipid rafts have been reported to play a crucial role in regulating the anti-inflammatory signal of cells and are regarded as the preferred site for the assembly of multimolecular complexes that mediate endocytosis of activated toll-like receptor 4 (TLR4).20
RFTN1 encodes a lipid raft linker that may play a key role in the formation and maintenance of lipid rafts.21 In addition, under the stimulation of bacterial lipopolysaccharide, RFTN1 mediates the internalization of TLR4 into endosomes in dendritic cells and macrophages, and poly (I: C) into TLR3-positive endosomes in myeloid dendritic cells and epithelial cells, leading to the activation of TICAM1-mediated signals and subsequent IFnB1.22,23 It been implicated in T cell antigen receptor-mediated signal transduction by regulating tyrosine kinase LCK localization, T cell-dependent antibody production, and cytokine secretion. It may regulate signal transduction mediated by B cell antigen receptors.21 The concept of lipid rafts remains an emerging field of study and further research is required to elucidate its role in bacterial inflammation.
In this study, we sequenced the granulation of middle ear cavity from CSOM patients and lavage fluid of middle ear from normal volunteer and found that RFTN1 expression was enhanced compared to that in the normal group. Further mechanistic research using S. aureus and B. cereus to induce CSOM models demonstrated that RFTN1 promotes lipid raft formation and stability on the surface of middle ear epithelial cells, leading to TLR4 activation and an inflammatory response. Additionally, this mechanism was verified in a rat model of CSOM, and the probiotic Lactobacillus was shown to improve CSOM induced by S. aureus and B. cereus. In conclusion, our results suggest that Lactobacillus may inhibit inflammation in CSOM induced by S. aureus and B. cereus through the RFTN1-lipid raft-TLR4 pathway, providing novel insights into the pathogenesis of CSOM and probiotic treatments.
Materials and Methods
Ethics Statement
The clinical study was reviewed and approved by the ethics committee of The First Affiliated Hospital of Kunming Medical University (No. 2022-L-171). The study was conducted according to the standards set by the Declaration of Helsinki. All animal procedures were approved by the Institutional Animal Ethics Committee Kunming Medical University (No. KMMU2020225).
RNA Sequence
The clinical samples were part of the routine hospital laboratory procedure. RNA sequencing of granulation of middle ear cavity from CSOM patients (n=15) and lavage fluid of middle ear from normal volunteer (n=9) was conducted. Total RNA from was extracted with Trizol reagent (Invitrogen) and quantity was determined on Nanodrop (Thermo Fisher Scientific). The extracted total RNA samples were stored at a low temperature and sent to the Beijing Cnkingbio Biotechnology Corporation for the determination of total RNA concentration, integrity, purity, and total amount. After passing the test, the samples were used to construct the mRNA bank and sequenced.
Cell Culture
Human middle ear epithelial cells (HMEEC, HX-0344) was obtained from Shanghai Hexu Biotechnology Co., Ltd. Cells were cultured in a mixture of Dulbecco’s modified Eagle’s medium (DMEM, Thermo Fisher Scientific) containing 10% fetal bovine serum (Thermo Fisher Scientific), 100 u/mL penicillin (Thermo Fisher Scientific) and 100 μg/mL streptomycin (Thermo Fisher Scientific) and bronchial epithelial basal medium (BEBM)(1:1) containing a bag of BEGM Singlequots (Lonza Bioscience) per 500 mL of medium, the cells were cultured in 37 °C and 5% CO2 cell incubators. Fresh medium was changed every 2–3 days, and the culture was fused to 80% or more, digested with 0.25% trypsin (Yeasen) and 0.05% EDTA, digested with complete medium, and centrifuged for 5 min (1000r/min). The cells were resuspended in fresh medium.
Cell Transfection
Scrambled siRNA of RFTN1 (siRFTN1) or siRNA control (siCtrl), RFTN1 overexpressed plasmid (oeRFTN1), and vector plasmid (oeNC) were purchased from GeneChem (Shanghai, China). Cells were transfected with siRNA or plasmids using Lipofectamine 3000 (Thermo Fisher Scientific).
Preparation of Bacterial
B. cereus, S. aureus and were Lactobacillus were cultured at 37°C on plates of Luria-Bertani broth (LB, Becton Dickinson (BD)) with 1% agar (w/v, Sigma). Overnight cultures and subcultures were routinely cultured in LB broth with shaking at 37°C (250 rpm). The strains were dissolved and diluted to the desired concentration of sterile saline.
Inflammatory Model Induced by Bacterial-Cell Co-Culture
The flow chamber was precoated with 2 mg/mL bovine collagen, and 1×105 HMEEC cells were inoculated with RPMI1640 medium containing 20% serum at 37 °C, 5% CO2 incubator for 24 h. The bottom of the chamber was used for all experiments. S. aureus and B. cereus were inoculated at 2.5×107 CFU/mL in an outer dish and co-cultured for 2–3 day.
ELISA
The ELISA kit was employed to determine the levels of TNF-α, IL1-β, IL-6, IL-8, IL-10 in the supernatant of indicated HMEEC cells and lavage fluid of indicated rats.
Quantitative Polymerase Chain Reaction Assay (qPCR)
Total RNAs was isolated from the cell lines using Trizol reagent. The mRNA levels were measured using SYBR Premix Ex Taq (TaKaRa) on an ABI‐7500 Fast system (Applied Biosystems, CA). Relative mRNA levels were normalized to those of GAPDH using the 2−ΔΔCt method. The sequences used were as follows: RFTN1, ATGGGTTGCGGATTGAACAAG/AGCGGTATTCATAGGACACATCT; FLOT1, GCCCTGCATCCAACAGATCC/AATGCCAGTGACTGAGATGGG, FLOT2, TTGCTGACTCTAAGCGAGCC/TCCACGGCAATCTGTTTCTTG; TNF-α, CCTCTCTCTCTAATCAGCCCTCTG/GAGGACCTGGGAGTAGATGAG; IL-1β, ATGATGGCTTATTACAGTGGCAA/GTCGGAGATTCGTAGCTGGA; IL-6, ACTCACCTCTTCAGAACGAATTG/CCATCTTTGGAAGGTTCAGGTTG; IL-8, TTTTGCCAAGGAGTGCTAAAGA/AACCCTCTGCACCCAGTTTTC; IL-10, GACTTTAAGGGTTACCTGGGTTG/TCACATGCGCCTTGATGTCTG; GAPDH, GGAGCGAGATCCCTCCAAAAT/GGCTGTTGTCATACTTCTCATGG.
Membrane Protein Extraction
Cell membrane proteins were extracted using the Mem-PERTM Plus membrane protein extraction kit. The cells were scraped off the surface of the plate with a cell scraper, and 5×106 cells were resuspended in culture medium. The resulting cell suspension was centrifuged at 300 g for 5 min. The cell pellet was washed with 3 mL of cell wash solution and centrifuged at 300 × g for 5 min. The supernatant was carefully removed and discarded. Cells were resuspended in 1.5 mL of cell wash solution and transferred to a 2 mL centrifuge tube. Centrifugation (300 × g) was performed for 5 min and the supernatant was discarded. 0.75 mL permeabilization buffer was added to the cell pellet. Short vortices used to obtain a homogeneous cell suspension. It was incubated at 4 °C for 10 min with continuous mixing condition. The permeabilized cells were centrifuged at 16000 × g for 15 mins. The supernatant containing cytosolic proteins was carefully removed and transferred to a new tube. Solubilization buffer (0.5 mL) was added to the precipitate, which was then resuspended using pipette suction. It was incubated at 4 °C for 30 min with continuous mixing conditions. At 4 °C, the tube was centrifuged for 15 min at a speed of 16000 × g. Supernatants containing soluble membrane proteins and membrane-associated proteins were transferred to a new tube. Proceed with downstream applications. Cytosolic and membrane components on ice can be used immediately or stored at −80 °C after packaging for future use.
Western Blot
Proteins in the culture medium of cell lysates were separated using 12% SDS-PAGE and transferred to immobile transfer membranes (Millipore, Bedford, MA). The transferred blots were incubated with a blocking solution containing 5% dry milk in TBST (0.15% Tween 20, 200 mm NaCl, and 25 mm Tris-HCl [pH 7.6]). The blots were washed and incubated for 1 h with secondary peroxidase-conjugated antibodies (Santa Cruz Biotechnology). Immunoreactive bands were detected using ECL detection reagents (Amersham, Buckinghamshire) according to the manufacturer’s instructions.
Detection of Lipid Raft Content in HMEEC Cells by Vybrant™ Alexa Fluor™ 488 Lipid Raft Labeling Kit
The cells were labeled with a fluorescent CtxB conjugate. The cells were centrifuged, and the cell particles were gently resuspended in complete medium on ice. The cells were centrifuged again, and the cell particles were suspended in 2 mL of the fluorescent CtxB coupling working solution. The cells were incubated at 4 °C for 10 min. After incubation, cells were gently washed several times with 1× ice PBS. CtxB-labelled lipid rafts were cross-linked with anti-CtxB antibodies. The cells were centrifuged, and the cell particles were gently resuspended in 2 mL of ice anti-CtxB antibody working solution. The cells were incubated at 4 °C for 15 min. After incubation, cells were gently washed several times with 1 × ice PBS. The cells were placed in 1× ice PBS and observed under a fluorescence microscope with the corresponding filter groups.
CSOM Rat Model Establishment and Treatment
Animal studies were performed in accordance with the Guide for the Care and Use of Laboratory Animals. For the S. aureus and B. cereus infection experiment, Sprague-Dawley (SD) rats weighing 200–250 g were anesthetized by intramuscular injection of ketamine-toluene thiazide mixture at a dose of 0.25 ~ 3 mL/100 g body weight. Animals were placed in the supine position, and a transverse incision was made at a position approximately 0.5 cm below the hyoid level to separate and expose the anterior cervical muscles. The hyoid scapula and hyoid sternum muscles were separated near the hyoid bone, so that the bottom wall of the ear bleb could be seen. Sterile saline was injected into the left ear of the rats as a control, and 200 μL of the bacterial suspension (containing 3×108 CFU) was injected into the right ear. The injection hole in the ear wall was sealed with bone wax and the incision was sutured layer-by-layer.
For Lactobacillus treatment, Lactobacillus suspension (200 μL, 1×109 CFU) was applied to the right ears of the rats.
HE Staining
After the administration ended, all the rats in each group were sacrificed through cervical vertebra dislocation. The inner ear was dissected, fixed with 4% paraformaldehyde, decalcified, dehydrated, paraffin-embedded for sectioning, and subjected to HE staining. The pathological morphological changes of the middle ear mucosa were observed under a microscope.
Statistical Analysis
Statistical analyses were performed using Statistical Program for Social Science version 26.0 (SPSS 26.0), R software (version 4.0.3), and GraphPad Prism (version 9). Quantitative data with a normal distribution is presented as the mean ± SD and were analyzed using Student’s t-test or one-way analysis of variance. To estimate the significance of gene expression differences between the patient subgroups, we performed a two-sample t test for each gene. P≤0.05 indicates a statistically significant difference between measured values. The significance was set as *P < 0.05, **P < 0.01, and ***P < 0.001. Differentially expressed genes were screened using P-value < 0.05 and |logFC| >1.5 as cut-off criteria, respectively.
Results
The Role of RFTN1 and Lipid Rafts in CSOM
Variable differentially expressed genes (DEGs) between middle ear cavity from CSOM patients and lavage fluid of middle ear from normal volunteer were identified. A total of 3646 DEGs (1620 up-regulated and 2026 down-regulated) in CSOM were identified. Among them, we found that RFTN1 was highly expressed in the CSOM (P=0.045; Figure 1A and B). To explore the role of RFTN1 in CSOM, we co-cultured S. aureus and B. cereus with the HMEEC cell and constructed a CSOM cell model. The mRNA and protein expression levels of RFTN1 and lipid raft proteins FLOT1 and FLOT2 were increased in the CSOM cell model (Figure 1C–E). We observed changes in the expression of lipid rafts under inflammatory conditions using a lipid raft labeling kit. The results showed that the lipid raft content of the cell membrane increased significantly in the CSOM group (Figure 1F). These results indicate that CSOM induced by S. aureus and B. cereus can induce the expression of RFTN1 in HMEEC and the formation of lipid rafts on the cell membrane.
 |
Figure 1 The role of RFTN1 and lipid rafts in CSOM. (A) Volcano plot and (B) A hierarchical clustering heatmap were used to display the differentially expressed mRNA patterns between normal and CSOM group. The cell model of CSOM was induced by co-culture of S. aureus and B. cereus with HMEEC cells. The same number of cells were collected, and RNA, whole cell lysate (WCL) and membrane protein (Mem) were extracted. (C and D) The expression of RFTN1 and raft protein FLOT1/2 mRNA was detected by qPCR. (E) Western blot was used to detect the expression of RFTN1 and FLOT1/2 proteins in cells and cell membranes. (F) The lipid raft content of HMEEC cells was detected by Vybrant™ Alexa Fluor™ 488 lipid raft labeling kit. *P< 0.05, **P< 0.01, ***P< 0.001. |
RFTN1 Regulates Lipid Raft Formation and Inflammation in CSOM
To determine the function of RFTN1 in CSOM, we knocked down RFTN1 in HMEEC cells (Figure 2A and B). RFTN1 inhibition reduced TNF-α, IL-1β, IL-6, IL-8, and IL-10 levels (Figure 2C and D). In addition, lipid rafts on the cell membrane were reduced in RFTN1 knockdown cells (Figure 2E). These results indicated that RFTN1 is a key regulatory factor in the formation of lipid rafts and inflammatory responses.
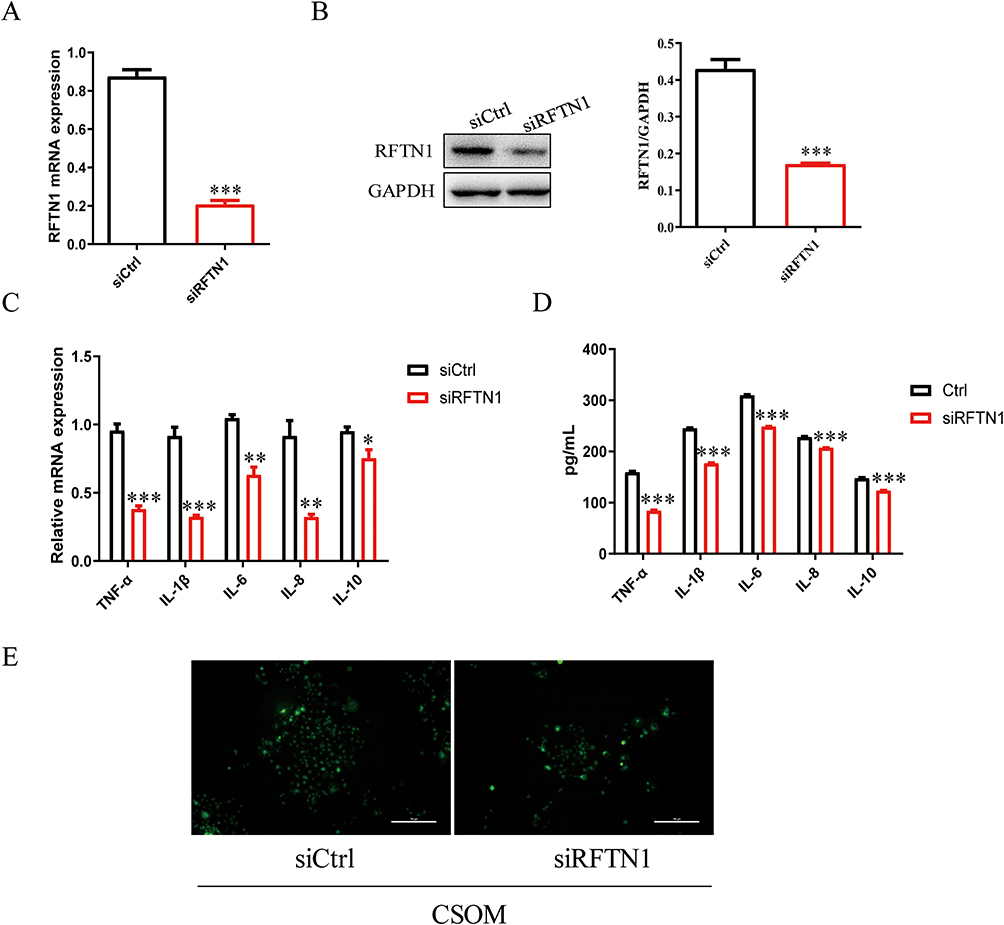 |
Figure 2 RFTN1 regulates lipid raft formation and inflammation in CSOM. siRFTN1 were transfected into HMEEC cells knock down RFTN1, and the transfected cells were cultured for 48 hours. (A and B) The mRNA and protein expression level of RFTN1 were detected using qPCR and Western blot. (C and D) qPCR and ELISA were used to detect the levels of inflammatory cytokines TNF-α, IL1-β, IL-6, IL-8, IL-10. (E) Vybrant™ Alexa Fluor™ 488 lipid raft labeling kit was used to detect the content of cell lipid rafts. *P< 0.05, **P< 0.01, ***P< 0.001. |
RFTN1 Modulates the Inflammation Response in CSOM via Lipid Raft Formation
We further determined the mechanism underlying RFTN1 expression in CSOM. We firstly conducted the RFTN1 overexpressed cells and introduced methyl-β-cyclodextrin (MβCD), an inhibitor of lipid raft24 to treat cells. The expression of RFTN1 in RFTN1 overexpressed cells was also determined (Figure 3A). Administration of MβCD counteracted the promoting effect of RFTN1 overexpression on the inflammatory response (Figure 3B and C) and the formation of lipid rafts (Figure 3D).
 |
Figure 3 RFTN1 modulates the inflammation response in CSOM via lipid raft formation. The cells transfected with oeRFTN1/control plasmid (oeNC) were treated with MβCD for 8 hours, and the cells were collected. (A) Expression of RFTN1 was determined by western blot. (B and C) qPCR and ELISA were used to detect the expression level of inflammatory cytokines. (D) The content of lipid rafts in cells was detected by Vybrant™ Alexa Fluor™ 488 lipid raft labeling kit. *P< 0.05, ***P< 0.001. |
RFTN1 Modulated the Expression of TLR4 in CSOM Cells
Previous studies have suggested that raft proteins are involved in the activation of TLR420 and that the recruitment of TLR4 to lipid rafts promotes a proinflammatory response.25 However, the function of TLR4 in RFTN1-induced inflammatory response in CSOM remains unknown. RFTN1 inhibition reduce the expression of TLR4 (Figure 4A). MβCD administration alleviated the promoting effect of RFTN1 overexpression on TLR4 expression (Figure 4B), indicating that RFTN1 modulated TLR4 expression via lipid rafts. In addition, immunofluorescence analysis confirmed that TLR4 expression was suppressed by RFTN1 inhibition (Figure 4C).
 |
Figure 4 RFTN1 modulated the expression of TLR4 in CSOM cells. (A) Expression of TLR4 in RFTN1 knocked down cells was detected by Western blot. (B) Expression of TLR4 in RFTN1 overexpressed cells with MβCD administration was detected by Western blot. (C) Immunofluorescence of TLR4 and RFTN1 in RFTN1 knocked down cells. *P< 0.05, **P< 0.01, ***P< 0.001. |
RFTN1 Positively Regulates S. aureus and B. cereus -Induced Inflammation in vivo
To further confirm the key role of RFTN1 in the pathogenesis of CSOM, we injected S. aureus and B. cereus suspensions into the right ears of rats to induce the CSOM model. After five days, MβCD was injected subcutaneously into rats in the control and CSOM groups. The middle ear was perfused with 1 mL of PBS thrice, and the levels of TNF-α, IL-1β, IL-6, IL-8, and IL-10 in the lavage fluid were measured. The rats were killed, pathological changes were observed by HE staining, and the expression of RFTN1 and TLR4 was detected by immunofluorescence. The results showed that In CSOM rats, the middle ear mucosa and mucosal layer were thickened, and tissue edema and gland dilatation were observed. Eosinophils of plasma cells were aggregated, goblet cells were increased, and nuclei were deeply stained, indicating that S. aureus and B. cereus can effectively induce CSOM development (Figure 5A). Consistent with CSOM cells, upregulated expression of RFTN1 and TLR4 induced by S. aureus and B. cereus was observed in middle ear tissue (Figure 5B). The colocalization of RFTN1 and TLR4 also increased in CSOM rats, and the expression of TNF-α, IL-1β, IL-6, IL-8, and IL-10 was higher in CSOM rats than in normal rats (Figure 5B and C). Additionally, MβCD treatment partially eliminated the effects of S. aureus- and B. cereus-induced pathological changes and inflammatory responses (Figure 5A and C). These results further prove that RFTN1 regulates the expression of TLR4, subsequently promoting the progression of CSOM in vivo.
 |
Figure 5 RFTN1 positively regulates S. aureus and B. cereus-induced inflammation in vivo. After 5 days establishing an CSOM model, MβCD is administered subcutaneously to rats in the control group and CSOM group. After 48 hours, the middle ear lavage fluid was collected, and the middle ear tissues are separated. (A) HE staining was used to observe the pathological changes of middle ear tissue; (B) GM1, RFTN1 and TLR4 were detected by immunofluorescence; (C) Determination of inflammatory factors in the middle ear lavage fluid. ***P< 0.001. |
Lactobacillus Improves CSOM Induced by S. aureus and B. cereus Through Lipid Raft-Dependent TLR4 Inflammatory Signal
Microbiological research on healthy children shows that Lactobacillus have a potential protective effect on OM,26 and Lactobacillus can be used as probiotics to treat diseases caused by other pathogenic bacteria, such as α hemolytic streptococcus.27 To study the therapeutic effect of Lactobacillus on CSOM caused by S. aureus and B. cereus, we treated a rat model with Lactobacillus. The results showed that Lactobacillus administration significantly reduced the inflammatory reaction of the middle ear epithelium and swelling of the tympanic membrane and pus (Figure 6A). The expression and colocalization of RFTN1 and TLR4 decreased, as did the expression of cytokines (Figure 6B and C). In summary, Lactobacillus therapy may effectively improve CSOM induced by S. aureus and B. cereus by inhibiting the RFTN1-TLR4 axis.
 |
Figure 6 Lactobacillus alleviates CSOM induced by S. aureus and B. cereus in rat. CSOM model was established, and Lactobacillus solution was injected at the same time. After 7 days, the middle ear lavage fluid was collected, and the middle ear tissue was separated. (A)HE staining was used to observe the pathological changes of middle ear tissue; (B) GM1, RFTN1 and TLR4 were detected by immunofluorescence; (C) Determination of inflammatory factors in the middle ear lavage fluid. **P< 0.01, ***P< 0.001. |
Discussion
In the present study, we performed RNA sequence analysis and the DEGs consisting of 1620 up-regulated genes and 2026 down-regulated genes in CSOM were identified. The expression of RFTN1 was significantly increased in CSOM patients. We the treated HMEEC cell and rats with S. aureus and B. cereus to induce CSOM and confirmed the key regulatory role of RFTN1 in the progression of CSOM. RFTN1 can affect the formation of lipid rafts on the cell membrane and the localization of TLR4 on lipid rafts, subsequently promoting the development of CSOM. We treated a rat model of CSOM with Lactobacillus and found that Lactobacillus treatment alleviated CSOM progression by inhibiting RFTN1 and TLR4 expression. Our findings suggest a crucial role for Lactobacillus in alleviating CSOM progression and uncovered the molecular mechanism involving Lactobacillus-regulated inhibition of the RFTN1-lipid raft-TLR4 signaling pathway under CSOM conditions.
OM refers to inflammation and/or infection of the middle ear, including a series of acute and chronic diseases clinically characterized by middle ear effusion.28 CSOM is characterized by a persistent and threatening pattern of severe ear discharge, tympanal perforation, and varying degrees of hearing loss over a period of more than six weeks. CSOM is a multi-etiological disease associated with irreversible pathological changes in middle ear tissue.29 It is estimated that 65–300 million patients, especially children, are affected. Currently, the most common risk factors for CSOM include persistent local gram-positive or gram-negative bacterial infections, recurrent acute OM, recurrent upper respiratory tract infections, poor sanitation, middle ear trauma, internal tympanic canals, and nutritional deficiencies.30 At present, although there are a variety of options for the standard treatment of CSOM, no significant breakthrough has been made in either surgical or conservative treatment, or in solving otorrhea or hearing loss. Therefore, probiotic therapy may be a potential target for the treatment and prevention of CSOM in the future.19 Many randomized controlled studies have reported that Lactobacillus rhamnosus GG,31–33 Bifidobacterium lactis BB-12,32,34 Streptococcus alpha hemolyticus,35 Streptococcus salivarius 24SMB,36 Streptococcus salivarius K1237 have good effects on the treatment and prevention of OM.
Lipid rafts are microstructure domains rich in cholesterol and sphingomyelin in the plasma membrane that play important roles in cell signal transduction. The structure of lipid rafts is dynamic, and lipid and protein contents are constantly changing. The types of lipids and proteins in the lipid rafts determine their size, stability, structure, and function. Most pathological conditions associated with lipid rafts are caused by an increase in their abundance or stability. Using lipid rafts as therapeutic targets has a significant effect on slowing the disease process. Studies have shown that lipid rafts play an important role in the regulation of diseases, such as inflammation, cancer, systemic lupus erythematosus, neurodegenerative diseases, and atherosclerosis. Considering the important role of inflammation in various pathological processes, targeting lipid rafts can alleviate various inflammation-induced diseases.38 Based on their different contents, lipid rafts can be divided into inflammatory, apoptosis signal molecule enrichment, and immune lipid rafts. Inflammatory rafts contain activated receptors, which mediate cellular inflammatory reactions.
TLR4 forms dimer in the ligand-activated reaction, which requires the microenvironment of lipid rafts, and the integrity of lipid rafts is very important for normal TLR4 signal transduction.39 A decrease in cellular cholesterol reduces the number of lipid rafts and destroys their structure of lipid rafts, thus inhibiting receptor dimerization and TLR4 recruitment to lipid rafts and weakening TLR4 signal transduction. However, an increase in cellular cholesterol increases the number of lipid rafts and enhance the signal transduction of TLR4.40
Saeki et al21 knocked out RFTN1 in chicken DT40B cells and observed a decrease in cell proliferation. By reintroducing the gene, raftlin was shown to be required for normal growth rate. In the absence of raftlin, the number of lipid rafts decreased and increased due to the overexpression of raftlin, indicating that raftlin is necessary for the formation or maintenance of lipid rafts. By consuming cholesterol, the lipid raft was destroyed, which led to a decrease in raftlin and a decrease in other components of lipid rafts similar to raftlin gene deletion, which confirmed previous observations. BCR-mediated tyrosine phosphorylation and calcium mobilization were impaired due to the lack of raftlin in DT40 cells, and the overexpression of raftlin was enhanced, indicating that raftlin is necessary for effective BCR signal transduction by forming or maintaining lipid rafts. Tatematsu et al22 studied the role of raftlin in TLR4 signal transduction in raftlin knockdown HEK293 cells and found that raftlin is necessary for activation of the IFN-β promoter mediated by TLR4 induced by lipopolysaccharide (LPS) in the presence or absence of membrane-bound CD14. Raftlin is essential for LPS-induced IFN-β production and the regulation of TLR4 signal transduction in human primary immune cells. Confocal microscopy showed that under the stimulation of endotoxin, the internalization of TLR4 also required raftlin. In response to LPS stimulation, raftlin moves from the cytoplasm to the plasma membrane, colocalizes with TLR4 on the cell surface, and is internalized by TLR4. Immunoprecipitation experiments confirmed that raftlin interacted with the clathrin/AP2 complex and mediated endocytosis of TLR4 by participating in the selection of goods with AP2. Our research, using humans and rats as representatives, confirmed the key role of RFTN1 in the signal transduction of cell membrane lipid rafts and TLR4 inflammation, and expounded the effects of RFTN1 on TLR4 activation either directly (binding with TLR4) or indirectly (affecting the formation of lipid rafts). The main and side effects of these two agents must be proven through further experiments.
Lactic acid bacteria are recognized as safe and non-pathogenic food-grade microorganisms that widely exist in the digestive tract of plants, dairy products, human beings, animals, and female vaginas, and mainly include 43 genera, such as Bifidobacterium, Lactobacillus, Lactococcus, Pediococcus, Leuconostoc and Streptococcus.41 Some strains of lactic acid bacteria are considered to be probiotics. According to the definition of probiotics by the World Health Organization (WHO), probiotics refer to a kind of living microorganisms that can exert beneficial effects on host health when ingested in sufficient quantities. The most common types are Lactobacillus and Bifidobacterium.16 Recently, probiotic lactic acid bacteria have attracted wide attention because of their important functions in improving intestinal flora balance, promoting host immunity, and preventing and treating diseases, such as reducing inflammatory response, anti-diabetes, and anti-tumor.42–46 However, at present, most research on probiotic therapy focuses on improving intestinal flora ecology by oral administration, and there is little research on the application of probiotics outside the intestine. Here, we propose a therapeutic strategy for implanting Lactobacillus into the skin or mucosa of the middle ear canal and other parts to inhibit the occurrence of inflammation and demonstrate the possible mechanism by which Lactobacillus can improve OM through the lipid raft -TLR4 axis. Many studies have shown that lactic acid bacteria play a regulatory role in cholesterol metabolism in vivo. Lactic acid bacteria in the intestine can inhibit cholesterol absorption in vivo by inhibiting the expression of NPC1L1, mediating the upregulation of the cholesterol transporter ABCG5/G8, and producing bile salt hydrolase during metabolism, which is the main regulator of serum cholesterol content.47–50 However, at present, research on the mechanism by which lactic acid bacteria regulate cholesterol degradation lacks systematization, and most studies only analyze and measure the expression and enzyme activity of one or a limited number of metabolic enzymes or proteins. There are few studies on the material basis, signal pathway, and the relationship between these proteins regulated by lactic acid bacteria.51,52 There are relatively few reports on whether the substances that lactic acid bacteria regulate cholesterol metabolism are metabolites of strains or cell wall components, and whether they regulate cholesterol metabolism directly or indirectly. Therefore, we speculate that one of the mechanisms by lactic acid bacteria regulate inflammation may be to affect cholesterol metabolism in epithelial cells and subsequently affect the formation of lipid rafts.
It is important to emphasize the potential limitations of this study. For example, a suspension of Lactobacillus was employed to directly treat the infected ear in this study. In other studies, means such as intraduodenal injection,53 intraperitoneal injection54 and oral administration55 were utilized. Whether there are any differences among these treatment methods, and which one is the best remain to be explored. And some studies indicated that the bactericidal effect of Lactobacillus,56 whether Lactobacillus can exert a bactericidal effect in CSOM remains to be investigated. Additionally, there have been clinical studies applying Lactobacillus to the treatment of diseases.57 How to enhance the therapeutic efficacy of Lactobacillus is also something that requires exploration. Whether Lactobacillus can exert a better effect through certain modifications or packaging remains a subject that we will continue to explore.
To sum up, although we have preliminarily proved the potential of Lactobacillus to improve CSOM by inhibiting lipid raft -TLR4 signaling pathway, there is still a lack of sufficient experiments to prove the molecular mechanism of Lactobacillus inhibiting lipid raft and affecting cholesterol metabolism in epithelial cells, which may be an issue worth exploring. In the subsequent study, we aim to utilize Lactobacillus in the clinical treatment of CSOM.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (81960187), Yunnan Provincial Department of Science and Technology, Kunming Medical University joint project (202201AY070001-051, 202101AY070001-082), and the Project of Yunnan Province to build a Science and Technology Innovation Center for South and Southeast Asia (202303AP140005).
Disclosure
The authors declare no competing interests.
References
1. Rosario DC, Mendez MD. Chronic Suppurative Otitis. StatPearls. Treasure Island (FL): StatPearls Publishing LLC; 2023.
2. Santos-Cortez RL, Hutchinson DS, Ajami NJ, et al. Middle ear microbiome differences in indigenous Filipinos with chronic otitis media due to a duplication in the A2ML1 gene. Infect Dis Poverty. 2016;5(1):97. doi:10.1186/s40249-016-0189-7
3. Khairkar M, Deshmukh P, Maity H, Deotale V. Chronic suppurative otitis media: a comprehensive review of epidemiology, pathogenesis, microbiology, and complications. Cureus. 2023;15(8):e43729. doi:10.7759/cureus.43729
4. Kelly MK, Ali K, Anping X, et al. A novel mouse model of chronic suppurative otitis media and its use in preclinical antibiotic evaluation. Sci Adv. 2020;6(33):eabc1828.
5. Khomtchouk KM, Joseph LI, Khomtchouk BB, et al. Treatment with a neutrophil elastase inhibitor and ofloxacin reduces P. aeruginosa burden in a mouse model of chronic suppurative otitis media. NPJ Biofilms Microbiomes. 2021;7(1). doi:10.1038/s41522-021-00200-z
6. Shefali D, Dharam V, Jaimanti B, Praveen R. Mapping of audiometric analysis with microbiological findings in patients with chronic suppurative otitis media (CSOM): a neglected clinical manifestation. Crit Rev Clin Lab Sci. 2023;60(3):212–232.
7. Durand ML. Endophthalmitis. Clin Microbiol Infect. 2013;19(3):227–234. doi:10.1111/1469-0691.12118
8. Samad L, Taghi Zahraei S, Mohammad Zare E, et al. Microbiological study of the auditory canal in dairy calves with otitis media. Microb Pathog. 2024;188:106547.
9. Daniel SJ. Topical treatment of chronic suppurative otitis media. Curr Infect Dis Rep. 2012;14(2):121–127. doi:10.1007/s11908-012-0246-8
10. Williams NT. Probiotics. Am J Health Syst Pharm. 2010;67(6):449–458. doi:10.2146/ajhp090168
11. Kim SK, Guevarra RB, Kim YT, et al. Role of probiotics in human gut microbiome-associated diseases. J Microbiol Biotechnol. 2019;29(9):1335–1340. doi:10.4014/jmb.1906.06064
12. Perihan Kubra A, Gozde K, Fatih T. Development and characterization of a novel sodium alginate based active film supplemented with Lactiplantibacillus plantarum postbiotic. Int J Biol Macromol. 2023;244:125240.
13. Kantapich S, Mongkol T. Bioprophylactic potential of novel human colostrum probiotics via apoptotic induction of colon cancer cells and cell immune activation. Biomed Pharmacother. 2022;149:112871.
14. Coleman A, Cervin A. Probiotics in the treatment of otitis media. The past, the present and the future. Int J Pediatr Otorhinolaryngol. 2019;116:135–140. doi:10.1016/j.ijporl.2018.10.023
15. Scott AM, Clark J, Julien B, et al. Probiotics for preventing acute otitis media in children. Cochrane Database Syst Rev. 2019;6(6):Cd012941. doi:10.1002/14651858.CD012941.pub2
16. Shanahan F, Dinan T, Ross P, Hill C. Probiotics in transition. Clin Gastroenterol Hepatol. 2012;10(11):1220–1224. doi:10.1016/j.cgh.2012.09.020
17. Maxs UES, Annytha IRD, Nelsi Kurniawati R. Detection of antibacterial activity of lactic acid bacteria, isolated from Sumba mare’s milk, against Bacillus cereus, Staphylococcus aureus, and Escherichia coli. J Adv Vet Anim Res. 2022;9(1):53.
18. Chetan S, Brij Pal S, Nishchal T, et al. Antibacterial effects of Lactobacillus isolates of curd and human milk origin against food-borne and human pathogens. 3 Biotech. 2017;7(1):1–9.
19. Carol N, Aren B, Shyamali S, Satya P, Lily HPN, Sam JD. Safety of transtympanic application of probiotics in a chinchilla animal model. J Otolaryngol Head Neck Surg. 2017;46(1):63.
20. Płóciennikowska A, Hromada-Judycka A, Borzęcka K, Kwiatkowska K. Co-operation of TLR4 and raft proteins in LPS-induced pro-inflammatory signaling. Cell Mol Life Sci. 2015;72(3):557–581. doi:10.1007/s00018-014-1762-5
21. Saeki K, Miura Y, Aki D, Kurosaki T, Yoshimura A. The B cell-specific major raft protein, Raftlin, is necessary for the integrity of lipid raft and BCR signal transduction. EMBO J. 2003;22(12):3015–3026. doi:10.1093/emboj/cdg293
22. Tatematsu M, Yoshida R, Morioka Y, et al. Raftlin controls Lipopolysaccharide-induced TLR4 internalization and TICAM-1 signaling in a cell type-specific manner. J Immunol. 2016;196(9):3865–3876. doi:10.4049/jimmunol.1501734
23. Watanabe A, Tatematsu M, Saeki K, et al. Raftlin is involved in the nucleocapture complex to induce poly(I:C)-mediated TLR3 activation. J Biol Chem. 2011;286(12):10702–10711. doi:10.1074/jbc.M110.185793
24. Sha Y, Liu Y, Yang J, et al. B3GALT4 remodels the tumor microenvironment through GD2-mediated lipid raft formation and the c-met/AKT/mTOR/IRF-1 axis in neuroblastoma. J Exp Clin Cancer Res. 2022;41(1):314. doi:10.1186/s13046-022-02523-x
25. Zou J, Feng D, Ling W, Duan R. Lycopene suppresses proinflammatory response in lipopolysaccharide-stimulated macrophages by inhibiting ROS-induced trafficking of TLR4 to lipid raft-like domains. J Nutr Biochem. 2013;24(6):1117–1122. doi:10.1016/j.jnutbio.2012.08.011
26. van den Broek MFL, De Boeck I, Kiekens F, Boudewyns A, Vanderveken OM, Lebeer S. Translating recent microbiome insights in otitis media into probiotic strategies. Clin Microbiol Rev. 2019;32(4). doi:10.1128/CMR.00010-18
27. Niittynen L, Pitkäranta A, Korpela R. Probiotics and otitis media in children. Int J Pediatr Otorhinolaryngol. 2012;76(4):465–470. doi:10.1016/j.ijporl.2012.01.011
28. Leichtle A, Hoffmann TK, Wigand MC. Otitis media: definition, pathogenesis, clinical presentation, diagnosis and therapy. Laryngorhinootologie. 2018;97(7):497–508. doi:10.1055/s-0044-101327
29. Mahmood FB, Amanda JL, Christopher GB-J. Chronic suppurative otitis media. Lancet. 2024;403(10441).
30. Elemraid MA, Mackenzie IJ, Fraser WD, et al. A case-control study of nutritional factors associated with chronic suppurative otitis media in Yemeni children. Eur J Clin Nutr. 2011;65(8):895–902. doi:10.1038/ejcn.2011.58
31. Näse L, Hatakka K, Savilahti E, et al. Effect of long-term consumption of a probiotic bacterium, Lactobacillus rhamnosus GG, in milk on dental caries and caries risk in children. Caries Res. 2001;35(6):412–420. doi:10.1159/000047484
32. Rautava S, Salminen S, Isolauri E. Specific probiotics in reducing the risk of acute infections in infancy—a randomised, double-blind, placebo-controlled study. Br J Nutr. 2009;101(11):1722–1726. doi:10.1017/S0007114508116282
33. Hatakka K, Blomgren K, Pohjavuori S, et al. Treatment of acute otitis media with probiotics in otitis-prone children-a double-blind, placebo-controlled randomised study. Clin Nutr. 2007;26(3):314–321. doi:10.1016/j.clnu.2007.01.003
34. Taipale T, Pienihäkkinen K, Isolauri E, et al. Bifidobacterium animalis subsp. lactis BB-12 in reducing the risk of infections in infancy. Br J Nutr. 2011;105(3):409–416. doi:10.1017/S0007114510003685
35. Roos K, Håkansson EG, Holm S. Effect of recolonisation with “interfering” alpha streptococci on recurrences of acute and secretory otitis media in children: randomised placebo controlled trial. BMJ. 2001;322(7280):210–212. doi:10.1136/bmj.322.7280.210
36. Marchisio P, Santagati M, Scillato M, et al. Streptococcus salivarius 24SMB administered by nasal spray for the prevention of acute otitis media in otitis-prone children. Eur J Clin Microbiol Infect Dis. 2015;34(12):2377–2383. doi:10.1007/s10096-015-2491-x
37. Di Pierro F, Colombo M, Giuliani MG, et al. Effect of administration of Streptococcus salivarius K12 on the occurrence of streptococcal pharyngo-tonsillitis, scarlet fever and acute otitis media in 3 years old children. Eur Rev Med Pharmacol Sci. 2016;20(21):4601–4606.
38. Bukrinsky MI, Mukhamedova N, Sviridov D. Lipid rafts and pathogens: the art of deception and exploitation. J Lipid Res. 2020;61(5):601–610. doi:10.1194/jlr.TR119000391
39. Chansrichavala P, Chantharaksri U, Sritara P, Ngaosuwankul N, Chaiyaroj SC. Atorvastatin affects TLR4 clustering via lipid raft modulation. Int Immunopharmacol. 2010;10(8):892–899. doi:10.1016/j.intimp.2010.04.027
40. Chowdhury SM, Zhu X, Aloor JJ, et al. Proteomic analysis of ABCA1-null macrophages reveals a role for stomatin-like protein-2 in raft composition and toll-like receptor signaling. Mol Cell Proteomics. 2015;14(7):1859–1870. doi:10.1074/mcp.M114.045179
41. Bermúdez-Humarán LG, Aubry C, Motta JP, et al. Engineering lactococci and lactobacilli for human health. Curr Opin Microbiol. 2013;16(3):278–283. doi:10.1016/j.mib.2013.06.002
42. Hutchinson AN, Tingö L, Brummer RJ. The potential effects of probiotics and ω-3 fatty acids on chronic low-grade inflammation. Nutrients. 2020;12(8):2402. doi:10.3390/nu12082402
43. Panebianco C, Latiano T, Pazienza V. Microbiota manipulation by probiotics administration as emerging tool in cancer prevention and therapy. Front Oncol. 2020;10:679. doi:10.3389/fonc.2020.00679
44. Sun Z, Sun X, Li J, et al. Using probiotics for type 2 diabetes mellitus intervention: advances, questions, and potential. Crit Rev Food Sci Nutr. 2020;60(4):670–683. doi:10.1080/10408398.2018.1547268
45. Castro-Mejía JL, O’Ferrall S, Krych Ł, et al. Restitution of gut microbiota in Ugandan children administered with probiotics (Lactobacillus rhamnosus GG and Bifidobacterium animalis subsp. lactis BB-12) during treatment for severe acute malnutrition. Gut Microbes. 2020;11(4):855–867. doi:10.1080/19490976.2020.1712982
46. Zeng Z, Yuan Q, Yu R, Zhang J, Ma H, Chen S. Ameliorative effects of probiotic Lactobacillus paracasei NL41 on insulin sensitivity, oxidative stress, and beta-cell function in a type 2 diabetes mellitus rat model. Mol Nutr Food Res. 2019;63(22):e1900457. doi:10.1002/mnfr.201900457
47. Duan LP, Wang HH, Ohashi A, Wang DQ. Role of intestinal sterol transporters Abcg5, Abcg8, and Npc1l1 in cholesterol absorption in mice: gender and age effects. Am J Physiol Gastrointest Liver Physiol. 2006;290(2):G269–276. doi:10.1152/ajpgi.00172.2005
48. Kumar M, Nagpal R, Kumar R, et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp Diabetes Res. 2012;2012:902917. doi:10.1155/2012/902917
49. Kikuchi T, Orihara K, Oikawa F, et al. Intestinal CREBH overexpression prevents high-cholesterol diet-induced hypercholesterolemia by reducing Npc1l1 expression. Mol Metab. 2016;5(11):1092–1102. doi:10.1016/j.molmet.2016.09.004
50. Tomaro-Duchesneau C, Jones ML, Shah D, Jain P, Saha S, Prakash S. Cholesterol assimilation by Lactobacillus probiotic bacteria: an in vitro investigation. Biomed Res Int. 2014;2014:380316. doi:10.1155/2014/380316
51. Chen K, Pachter L. Bioinformatics for whole-genome shotgun sequencing of microbial communities. PLoS Comput Biol. 2005;1(2):106–112. doi:10.1371/journal.pcbi.0010024
52. Lye HS, Rusul G, Liong MT. Removal of cholesterol by lactobacilli via incorporation and conversion to coprostanol. J Dairy Sci. 2010;93(4):1383–1392. doi:10.3168/jds.2009-2574
53. Tanida M, Yamano T, Maeda K, Okumura N, Fukushima Y, Nagai K. Effects of intraduodenal injection of Lactobacillus johnsonii La1 on renal sympathetic nerve activity and blood pressure in urethane-anesthetized rats. Neurosci Lett. 2005;389(2):109–114. doi:10.1016/j.neulet.2005.07.036
54. Wan F, Wang H, Wang M, Lv J, Zhao M, Zhang H. Sustained release of Lactobacillus casei cell wall extract can induce a continuous and stable IgA deposition model. J Pathol. 2022;257(3):262–273. doi:10.1002/path.5884
55. Nápoles-Medina AY, Aguilar-Uscanga BR, Solís-Pacheco JR, et al. Oral administration of lactobacillus inhibits the permeability of blood-brain and gut barriers in a parkinsonism model. Behav Neurol. 2023;2023:6686037. doi:10.1155/2023/6686037
56. Wu CC, Lin CT, Wu CY, Peng WS, Lee MJ, Tsai YC. Inhibitory effect of Lactobacillus salivarius on Streptococcus mutans biofilm formation. Mol Oral Microbiol. 2015;30(1):16–26. doi:10.1111/omi.12063
57. Huang R, Wu F, Zhou Q, et al. Lactobacillus and intestinal diseases: mechanisms of action and clinical applications. Microbiol Res. 2022;260:127019. doi:10.1016/j.micres.2022.127019
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.