Back to Journals » Journal of Inflammation Research » Volume 18
ITGB7 Remodels Inflammation and Immune Microenvironment and Enhances Checkpoint Inhibitor-Based Immunotherapy in Pancreatic Cancer
Authors Zhu Y, Lei Y, Luo A, Zhang Q, Cai L, Zhang Y, Li X, Wang B, Sun D, Li Y, Sun P, Gong S, Cheng Y
Received 15 April 2025
Accepted for publication 2 December 2025
Published 16 December 2025 Volume 2025:18 Pages 17633—17649
DOI https://doi.org/10.2147/JIR.S534605
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Yun Zhu,1 Yi Lei,2 Anni Luo,1 Qingqing Zhang,2 Linqian Cai,1 Yongxin Zhang,1 Xiangzhao Li,3 Biao Wang,4 Danning Sun,1 Yanqi Li,1 Penghui Sun,5 Sitang Gong,2 Yang Cheng2
1State Key Laboratory of Organ Failure Research, Guangdong Provincial Key Laboratory of Viral Hepatitis Research, Department of Infectious Diseases, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China; 2Digestive Department, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, Guangzhou, Guangdong, 510623, People’s Republic of China; 3Department of Pathology, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong, 510515, People’s Republic of China; 4Department of Hepatobiliary Surgery, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong, 510515, People’s Republic of China; 5Nanfang PET Center, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong, 510515, People’s Republic of China
Correspondence: Yang Cheng, Department of Digestive, Guangzhou Women and Children’s Medical Center, Guangzhou Medical University, No. 9 Jinsui Road, Guangzhou, Guangdong, 510623, People’s Republic of China, Email [email protected]
Background: The role of ITGB7 in pancreatic ductal adenocarcinoma (PDAC) remains unreported. This study aims to investigate the effects of ITGB7 on inflammation and immune microenvironment, and its correlation with the response to immune checkpoint inhibitors (ICIs).
Methods: The correlation between ITGB7 expression and the response to ICIs treatment was evaluated in our clinical cohort and in the TCGA dataset. TMT and PRM proteomic analysis identified ITGB7-associated proteins, biological processes, and signal pathways. The role of ITGB7 in macrophage polarization was explored both in vivo and in vitro. Additionally, ITGB7-associated immune and inflammatory regulatory molecules, and immune cells infiltration in PDAC were assessed using TCGA dataset. An ITGB7-associated ceRNA network was constructed to explore post-transcriptional regulation.
Results: Clinical sample analysis and TCGA analysis showed that ITGB7 was significantly overexpressed in PDAC compared to normal pancreatic tissue. Patients with high ITGB7 expression demonstrated a better response to ICIs blockade therapy and exhibited a favorable prognosis. Proteomic analysis indicated that ITGB7 regulates immune- and inflammation- related proteins. ITGB7 expressed in pancreatic tumor cells promoted M2 macrophage polarization. ITGB7 modulated immune-related signaling pathways and was positively correlated with expression of immune checkpoint molecules, inflammatory cytokines, and immune-stimulating molecules. ITGB7 correlated with increased immune cell infiltration.
Conclusion: We provide the first evidence that ITGB7 expression correlates with immune regulation, inflammatory modulation, and immune cell infiltration in PDAC. ITGB7 is associated with enhanced immunotherapy response and improved prognosis. This study provides novel insights into the regulation of PDAC immune responses.
Keywords: pancreatic ductal adenocarcinoma, ITGB7, immunotherapy response, inflammatory modulation, prognosis
Introduction
Pancreatic cancer, particularly pancreatic ductal adenocarcinoma (PDAC), is highly aggressive with a five-year survival rate of only 13%. Globally, it accounts for approximately 67,440 new cases annually and 51,980 deaths.1 PDAC is characterized by its highly aggressive nature, early recurrence, and metastasis, resulting in poor prognosis. For advanced PDAC, conventional chemotherapy remains the primary treatment therapy but exhibits limited efficacy. Immune checkpoint inhibitors (ICIs) have shown limited success in improving survival rates for patients with advanced PDAC.2,3 The mechanisms underlying this resistance remain incompletely understood.
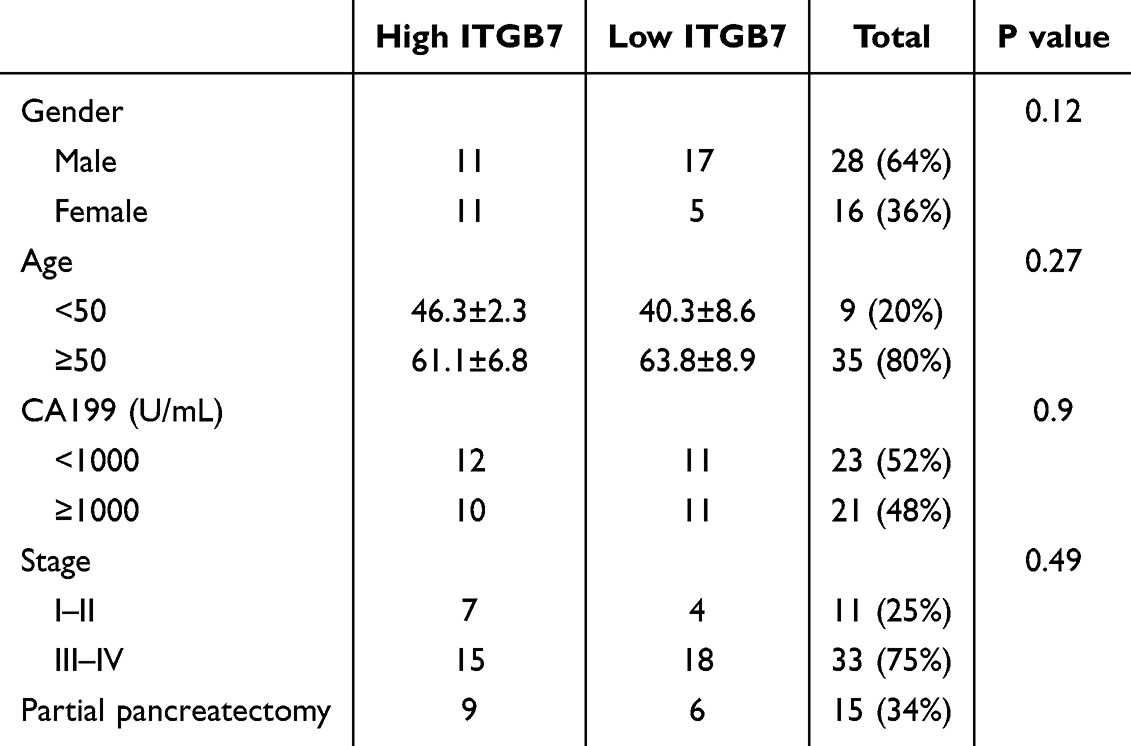 |
Table 1 The Characteristics of Patients with PACD Treated with PD-1 Antibody Therapy |
The tumor microenvironment (TME) in PDAC is a complex system critical for tumor progression and response to therapy. It is composed of a complex network of cellular and non-cellular components, including cancer cells, immune cells, fibroblasts, and inflammatory cytokines and chemokines.4 As an immunologically “cold” tumor, the pancreatic TME is composed of numerous populations of fibroblasts, dense extracellular matrix, a poorly formed vascular system, and a diverse and largely suppressive populations of immune cells.5 PDAC’s complex immunosuppressive tumor microenvironment that acts to insulate the tumor against an effective cytotoxic immune response.6 Chronic inflammation recruits tumor associated macrophages (TAMs) and other immunosuppressive cells that enhance tumor progression by facilitating immune evasion.7 Chronic inflammation promotes cancer cells survival, proliferation, and invasion.8
Integrin-β (ITGB) superfamily, belongs to the superfamily of integrins.9 Extensive alterations in integrin expression have been observed in malignant tumors where they promote cancer metastasis through transmission of signals from growth factor receptors.10 The ITGB family encompasses eight subtypes in the human body. In the intestine, ITGB7 promotes the homing of Treg to gut-associated lymphoid tissues and maintains the intrinsic function of Treg.11 ITGB7 could inhibit the tumorigenesis and progression of colorectal cancer by maintaining antitumor immunity in colorectal cancer.12 However, the role of ITGB7 in PDAC remains unclear.
In this study, we performed the first proteomic study of ITGB7 intervention in PDAC. We also evaluated the impact of ITGB7 on macrophage polarization, immune and inflammatory regulation in PDAC. Furthermore, we examined the association of ITGB7 expression with responses to ICIs and patient prognosis. Investigating the role of integrin ITGB7 in regulating immune responses and inflammatory processes in PDAC may provide valuable insights into mechanisms of immune evasion and contribute to the enhancement of immunotherapeutic efficacy.
Methods
Clinical Samples
We collected tumor tissues and clinical data of patients with PDAC who were treated with anti programmed cell death protein 1 (PD1) antibody at Nanfang Hospital, South Medical University from October 2018 to April 2024. The inclusion criteria comprised a pathological diagnosis of PDAC, availability of tumor biopsy samples or residual tissue following surgical resection, completion of at least three cycles of PD-1 monoclonal antibody treatment, and at least one documented tumor assessment. Patients with incomplete follow-up data or no tumor assessment were excluded. A total of 44 eligible patients were enrolled in the study, of whom 64% were male. The median age was 59 years (range: 31–82 years). Fifteen patients had undergone partial pancreatectomy. All patients provided written informed consent, which was approved by the Ethics Committee of Nanfang Hospital, South Medical University. The patient characteristics are presented in Table 1. The median follow-up time was 11.5 months. Our study complies with the Declaration of Helsinki.
Immunohistochemistry (IHC)
The clinical patient samples were immunostained with anti-ITGB7 antibody (Novus, China) as described in a previous study.13 A negative control was included using only secondary antibodies to ensure specificity. The staining score was independently evaluated by two researchers and comprised the integrated staining intensity score and the average proportion of positive cells score.
Survival Analysis for Clinical PDAC Patients
Patients were stratified into high and low ITGB7 expression groups based on the median score of ITGB7 expression determined by IHC. Their response to PD1 antibody treatment was assessed using RECIST 1.1 criteria every 6–12 weeks. Responses were classified as complete response (CR), partial response (PR), stable disease (SD), or progressive disease (PD). Objective response rate (ORR) was defined as the proportion of patients achieving CR and PR. Disease control rate (DCR) referred to the proportion of patients without disease progression. Univariate survival analysis was conducted using Kaplan–Meier (KM) survival method, and the Log rank test was applied to compare survival differences between groups.
Construction of Virus-Infected Cells
The human pancreatic cancer cell line PANC-1 was obtained from the Cell Bank of the Typical Culture Collection Committee at the Chinese Academy of Sciences. PANC-1 cells were cultured to logarithmic phase and infected with LV-ITGB7-sgRNA/sgRNA-control (Supplementary Table 1). Cell selection was carried out using 200 μg/mL G418 and 1.00 μg/mL Puromycin. Western blot analysis was conducted to validate the successful generation of stable expressing cells for ITGB7 and β-actin (Supplementary Figure 1).
Tandem Mass Tag (TMT) Analysis
Total protein was extracted from negative control (NC) and ITGB7 overexpressioned (OE) PANC-1 cells (n=3), digested with trypsin, and labeled with TMT reagents (Thermo Fisher Scientific). The peptides were fractionated using an Agilent 1260 infinity II HPLC system and separated using a nanoflow rate Easy nLC system. Mass spectrometry analysis was conducted using a Q Exactive HF-X mass spectrometer. Detailed detection methods can be found in the Supplementary Method.
Proteins with a fold change > 1.2 and P-value < 0.05 were considered as differentially expressed proteins (DEPs). Gene Ontology (GO) annotation and enrichment were performed using Blast2GO and Fisher’s exact test. KEGG pathway enrichment analysis was conducted for the target proteins. Subcellular localization of DEPs was predicted using WoLF PSORT. Functional domains of DEPs were annotated using Interpro, and their distribution in the total proteome was analyzed using Fisher’s exact test. Protein expression levels were normalized, and differences between the groups were verified using Parallel Reaction Monitoring (PRM). Clustering analysis was conducted using Matplotlib to generate hierarchical cluster heatmaps. Interaction networks were constructed using Cytoscape (version 3.2.1).
Mononuclear Cells Isolation and Co-Culture Experiment
CD14+ monocytes were isolated from Peripheral blood mononuclear cells (PBMCs) obtained from healthy donors who provided informed consent, using MACS cell separation (Miltenyi Biotec GmbH). The purity of CD14+ monocytes was confirmed to be over 95% by flow cytometry (Supplementary Figure 2). Monocytes were cultured in 24-well plates at a density of 5×105 cells/well. Supernatants from ITGB7-OE or control PANC-1 cells and BxPC-3 cells were used as conditioned culture media for monocytes based on the optimal Effector: Target (E: T) ratio. After 4 days of culture, the differentiated monocytes were harvested for further experiments.
Immunofluorescence Staining and Confocal Microscopy
Immunofluorescence staining of pretreated cells or tissue sections was performed as previously described.14 Cells were fixed with 4% paraformaldehyde at 4°C for 15 minutes, and tissue underwent dewaxing and antigen retrieval. Samples were blocked with 5% goat serum at 37°C for 30 minutes. Primary antibodies, anti-iNOS antibody (Proteintech, 18985-1-AP, 1:600), and anti-CD206 antibody (abcam, ab64693, 1:1000), were added and incubated overnight at 4°C, followed by washing. CD206 staining, an Alexa Fluor 488-labeled anti-rabbit secondary antibody (Cell Signaling Technology, 4412) was incubated in the dark at room temperature for one hour. Samples were then incubated with DAPI for 10 minutes. For iNOS staining, an HRP-conjugated secondary antibody (Zhongshanjinqiao, PV-6001) was incubated for 40 minutes. Subsequently, Cy5 tyramide (AAT, 11066) was applied for 10 minutes. Then samples were washed and mounted. Images were captured using Observation with Laser Scanning Confocal microscope (Zeiss, LSM880).
Quantitative Real-Time PCR
The total RNA was extracted using the TRIZOL reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol.15 cDNA was synthesized from total RNA using PrimeScript™ RT Master Mix (Takara, Shiga, Japan). Quantitative PCR (q-PCR) analysis was conducted on the Step-One Plus Real-Time PCR System utilizing Power SYBR Green PCR Master Mix (Applied Biosystems, Foster, CA), and data were analyzed using StepOne Software. Gapdh served as an internal control. The primer sequences are provided in Supplementary Table 2.
Dataset Collection and Survival Analysis
Transcriptome data and miRNA expression data of PDAC tumor tissues were obtained from the TCGA PAAD database. Gene expression data of tumor cell lines were acquired from Cancer Cell Line Encyclopedia (CCLE). ITGB7 expression differences between tumor and normal tissues was analyzed using GEPIA2, based on gene expression data from normal tissues in GTEx and tumor tissues in TCGA with visualization via ggplot and statistical analysis using the Mann–Whitney U-test. A total of 81 patients with complete clinical information and prognosis data were categorized into a high-expression group (n=41) and a low-expression group (n=40) based on median ITGB7 levels. Overall survival (OS) and disease-free survival (DFS) analyses were conducted. KM survival curve was performed using R package survminer, using the Log rank test. Correlation analysis between ITGB7 expression and immune checkpoint molecules, MHC molecules, chemokine factors, as well as chemokine receptors was performed using correlation test and visualized through heatmaps.
Identification of DEGs and Functional Enrichment Analysis
The PAAD database contains 178 samples. Based on the median expression level of ITGB7 in tumor tissues, patients were divided into low-expression group (n=89) and high-expression group (n=89). Gene Set Enrichment Analysis (GSEA) was performed using the Hallmarks of Cancer collection, KEGG, and GO as the reference gene set. The cut-off criteria were as follows: normalized enrichment scores (NES) > 1.0, false discovery rate (FDR) q > 0.25 and nominal p < 0.05. Differentially expressed genes (DEGs) between the ITGB7 high and low expression groups were identified using the EdgeR package, with p-value < 0.05 and |log2 fold change| > 1.5. Volcano plots were constructed using the layered grammar of graphics implemented in the ggplot2 package within the R programming environment.
GO and KEGG pathway analyses were conducted using the R package “clusterProfiler”, with significant differences defined as p value < 0.05 and FDR < 0.05 for enriched signaling pathways and biological processes. The enrichment scores of molecular pathways and gene expression signatures were assessed using single-sample gene set enrichment analysis, implemented in the R package “GSVA”.
Protein-Protein Interaction (PPI) Network
According to GSEA enrichment analysis, the 100 most significant correlation hub genes were imported to construct a PPI network comprising of 50 positively correlated genes and 50 negatively correlated genes. The online STRING database (https://string-db.org) was used to screen the hubs and construct the network. The PPI network was constructed by setting medium confidence at 0.400 Disconnected nodes were excluded. The correlations between ITGB7 and IRF4, or PTPRC, or PRKCH were evaluated.
Comprehensive Correlation Analysis of Tumor-Infiltrating Immune Cells
To elucidate the immune cell landscape in high and low-ITGB7 expression groups, we employed CIBERSORT (https://cibersort.stanford.edu/) with the LM22 signature. Pearson correlation analysis identified the relationship between immune cell phenotypes and ITGB7. TIP database (http://biocc.hrbmu.edu.cn/TIP/index.jsp) was used to evaluate the immune status of patients with high and low ITGB7 expression levels, with visualization performed using R script. The TIMER tool (https://cistrome.shinyapps.io/timer) was used to analyze the composition of tumor-infiltrating immune cells and their correlations with ITGB7 or gene hubs identified in the PPI network.16 TIDE and IPS signatures were assessed for predicting response to ICIs.
Chemotherapeutic Response Prediction
The comprehensive Genomics of Drug Sensitivity in Cancer (GDSC) database (https://www.cancerrxgene.org/) was used to predict the response of PDAC samples to small molecule targeted drugs and chemotherapy agents, based on existing literature.17 The prediction process was executed using the R package “pRRophetic”, wherein ridge regression estimated the half-maximal inhibitory concentration (IC50) of samples.
Cell Viability Assay (CCK-8) and IC50 Determination
ITGB7 OE and NC PANC-1 cells were treated with different concentrations of cisplatin (0, 5, 25, 100 μM), erlotinib (0, 5, 10, 20, 40 μM), docetaxel (0, 0.5, 1, 1.5, 2 μM), and all-trans retinoic acid (ATRA; 0, 20, 40, 60, 80 μM) for 48 h. After treatment, 10% CCK-8 solution (Elabscience, China) was added, and cells were incubated for 1 h at 37 °C. Absorbance at 450 nm was measured using a microplate reader (Thermo Fisher Scientific, USA). Dose–response curves were generated using a four-parameter logistic regression model in GraphPad Prism 9 (GraphPad Software, USA) to determine IC50 values.
lncRNA-miRNA-mRNA Network Construction
The mRNA, lncRNA, and miRNA data were stratified by the median expression of ITGB7. Differential analysis was conducted using thresholds of |log2FC| > 1 and FDR < 0.05. Differentially expressed lncRNAs and miRNAs were paired using miRcode. Target genes for the identified miRNAs were predicted using miRDB, miRTarBase, and TargetScan. A competing endogenous RNA (ceRNA) network was constructed from 113 intersecting genes and miRNAs, and visualized with Cytoscape (version 3.8.0).
Results
ITGB7 is Upregulated in PDAC and Correlates with Favorable Response to ICIs Treatment
IHC analysis revealed significantly higher ITGB7 expression in PDAC tissues compared to adjacent normal tissues in 44 patients in the PD-1 antibody immunotherapy cohort (Figure 1A). Patients were divided into high- and low-expression groups based on ITGB7 expression levels in tumor tissues. No significant differences in baseline characteristics were observed between the two groups (Table 1). Patients with high ITGB7 expression exhibited better ORR and DCR to PD-1 antibody treatment (Table 2). IHC also revealed higher PDL1 expression in the ITGB7 high-expression group compared to the low-expression group (Figure 1B). Patients with high ITGB7 expression exhibited longer PFS than those with low ITGB7 expression (Figure 1C). Analysis of TCGA-PAAD and GTEx database revealed significantly higher ITGB7 expression in tumor tissues compared with normal tissues (Figure 1D). Among 178 samples from TCGA, no significant differences were observed between the two groups in age, gender, TNM stage, grade (Supplementary Table 3), OS or DFS (Supplementary Figure 3). Multivariate analyses indicated that ITGB7 was not an independent prognostic indicator for PDAC (Supplementary Table 4). TCGA data indicated that patients with high ITGB7 expression had prolonged OS when treated with PD-L1 antibody (Figure 1E). These findings suggest that ITGB7 is overexpressed in PDAC and is positively correlated with a better response to ICIs.
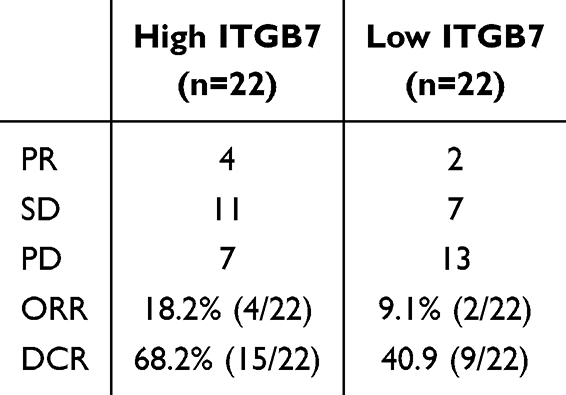 |
Table 2 Treatment Response to PD1 Antibody in PACD Patients with High Versus Low ITGB7 Expression in Tumor Tissue |
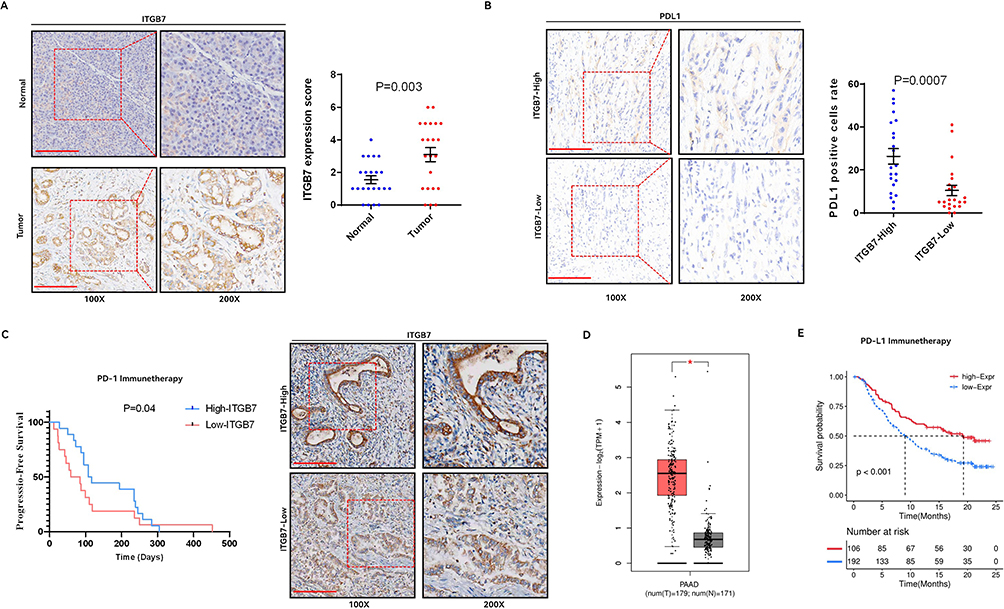 |
Figure 1 ITGB7 is upregulated in PDAC and correlates with favorable response to ICIs treatment. (A) Expression of ITGB7 in tumor and adjacent non-tumor tissues from patients in the clinical cohort using IHC. Representative pictures were shown. Bar=200um. (B) Expression of PDL1 in tumor tissues between the high and low ITGB7 expression patient groups in the clinical cohort. (C) PFS of patients in the high and low ITGB7 expression groups in the clinical cohort. (D) Differential expression of ITGB7 in pancreatic cancer versus normal tissues analyzed by the GEPIA2 database. *, p<0.05. (E) Survival analysis of patients treated with PD-L1 antibody in the PAAD dataset, comparing high vs low ITGB7 expression. |
ITGB7 Regulates Inflammation and Immune Response in PDAC
To investigate the function of ITGB7 in PDAC, we employed TMT analysis to identify DEPs in ITGB7-OE PANC-1 cells compared to NC cells, and further analyzed their potential biological functions. Clustering results were shown (Figure 2A). DEPs included 53 upregulated and 33 downregulated proteins (Figure 2B).
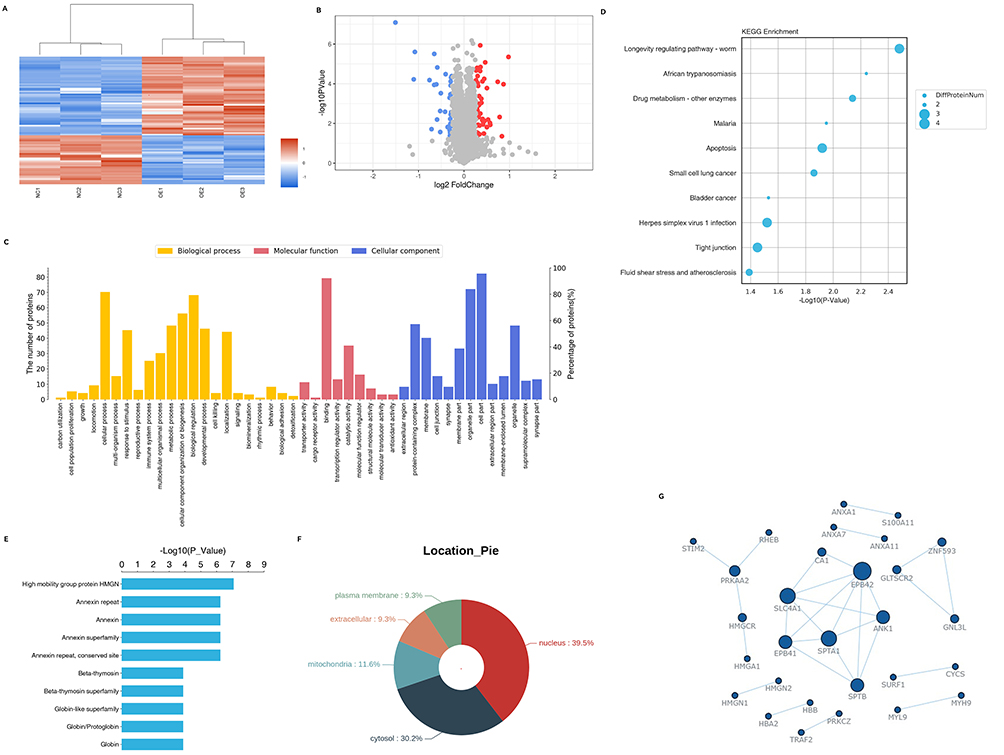 |
Figure 2 TMT-based quantitative proteomic analysis of control versus ITGB7 overexpression pancreatic cancer cells. (A) Clustering analysis of NC and ITGB7 OE PANC01 cell samples. (B) Volcano plot of DEPs in NC and OE PANC01 cell lines. Red, significantly upregulated proteins; blue, significantly downregulated proteins; gray, proteins with no significant change. (C) GO classification of DEPs into biological process, cellular component and molecular function. (D) KEGG pathway enrichment analysis of DEPs. Each bubble refers to the number of DEPs. (E) Protein domain enrichment analysis. The x-axis represents enrichment fold and y-axis means protein domain. (F) Subcellular localization analysis of DEPs in PANC01 cells. (G) The PPI network of immune-related DEPs. |
GO annotation revealed that DEPs mainly participate in cell proliferation, immune regulation, and metabolism. The main functions include binding and catalytic activity. Cellular components are mainly cell parts, organelle parts, and protein complexes (Figure 2C). KEGG analysis identified key pathways including drug metabolism, apoptosis, and tight junction signaling (Figure 2D). Domain enrichment analysis revealed that DEPs are predominantly associated with high mobility group (HMG), Annexin (ANX) superfamily, and β-thymosin (Tβ) superfamily (Figure 2E). Subcellular localization prediction indicated that DEPs are predominantly located in the nucleus (39.5%) and cytosol (30.2%) (Figure 2F). PRM analysis validated DEPs related to tumor cell immune and inflammatory responses, with expression trends consistent with TMT results (Supplementary File 1). The PPI network constructed from these DEPs (Figure 2G) aligned with domain enrichment findings, highlighting proteins involved in immune response (ANXA & HMG), inflammatory response (TRAF2), and drug metabolism (CYCS& SURF1). Our results suggest that ITGB7 is involved in regulating immune and inflammatory processes, and drug metabolism.
ITGB7 Promotes the Polarization of M2 Phenotype Macrophages
We further explored the effect of ITGB7 on the polarization of TAMs in pancreatic cancer in vivo and in vitro. IHC and immunofluorescence analyses revealed that, in the clinical PDAC cohort, patients with high ITGB7 expression exhibited significantly greater infiltration of CD206-positive cells and markedly reduced iNOS-positive cells compared to those in the low ITGB7 group (Figure 3A and B). In the co-culture model, the proportion of CD206+ macrophages increased significantly when treated with supernatant from ITGB7-overexpressing PANC-1 cells compared to the control cells (Figure 3C). Furthermore, ITGB7 overexpression upregulated M2-associated genes (IL6, IL10, ARG1, CCL22) and downregulated M1-associated genes (IL1B, CXCL9) in differentiated monocytes (Figure 3D and E). These findings suggest that ITGB7 promotes macrophages polarization toward the M2 phenotype.
 |
Figure 3 ITGB7 expression in pancreatic cancer cells promotes macrophage M2 polarization. (A) Expression of CD206 in tumor tissues from patients with high or low ITGB7 expression. Representative pictures of IHC were shown. (B) Expression of iNOS (red) in tumor tissues of patients in the high and low ITGB7 expression groups. Representative pictures of immunofluorescence were shown. (C) Immunofluorescence of CD206 (green) in human monocytes cultured with conditioned medium (CM) from ITGB7-OE or NC PANC-1 cells. (D and E) RT-PCR analysis of M1 (D) and M2 (E) gene expression in human monocytes co-cultured with PANC-1 cells. (F) Immunofluorescence of CD206 (green) in human monocytes cultured with CM from ITGB7-OE or NC BxPC3 cells. (G and H) RT-PCR was used to measure M1 (G) and M2 (H) gene expression levels in differentiated monocytes co-cultured with BxPC3 cells, p<0.05; **, p<0.01. |
ITGB7 Regulates Inflammation and Immune Signal Pathways
We further analyzed the impact of ITGB7 on inflammatory and immune-related pathways in the TCGA dataset. GSEA analysis suggested that ITGB7 is enriched in pathways related to NK cell-mediated cytotoxicity, B and T cell receptor signal, cytokine-cytokine receptor interactions, chemokine signaling, and leukocyte transendothelial migration (Figure 4A). GSVA analysis revealed that ITGB7 is closely linked to IFNγsignaling response, inflammatory response and multiple inflammatory factor signaling pathways, including TNFα-NFκB and IL6-STAT3 (Figure 4B).
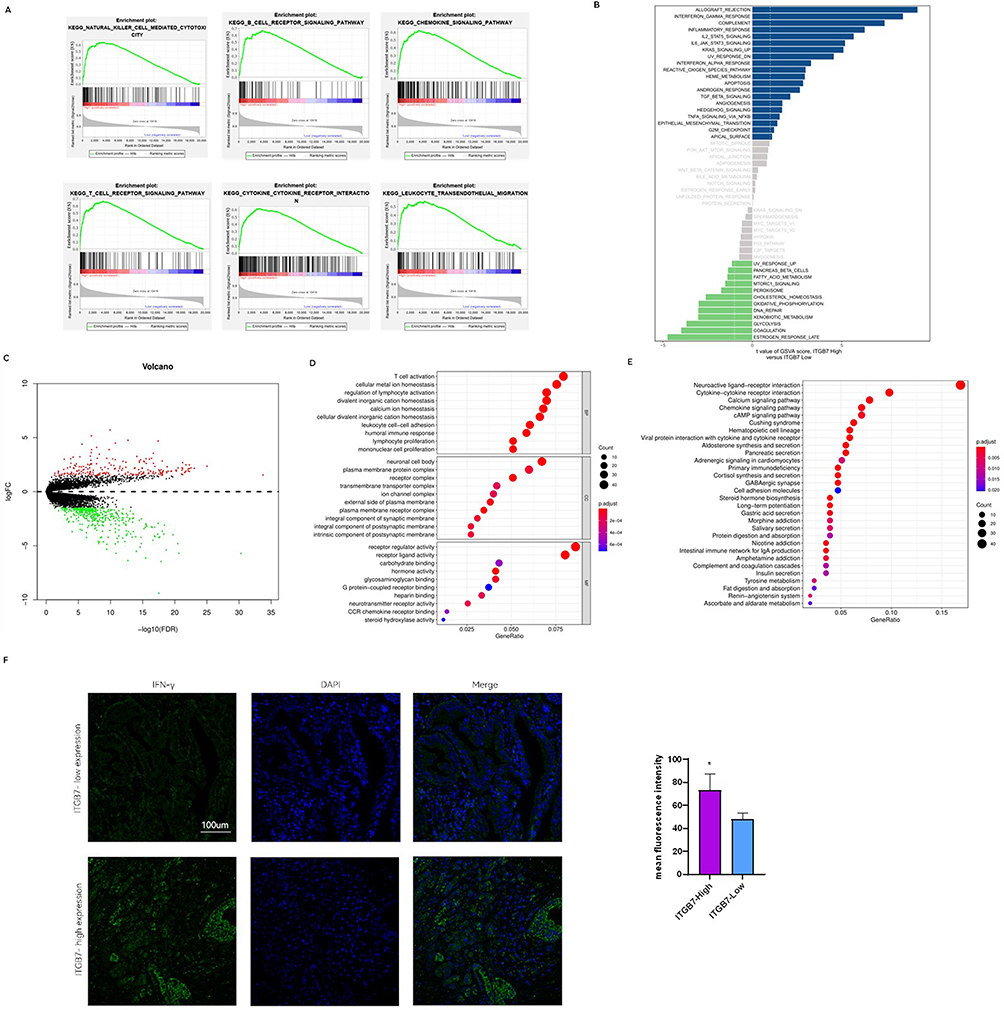 |
Figure 4 ITGB7 regulates inflammation and signal pathways. (A) Significantly different pathways in the GSEA enrichment analysis using single-gene method for ITGB7. (B) GSVA enrichment analysis results for ITGB7. Blue indicates upregulated pathways, green indicates downregulated pathways, and gray indicates non-significant pathways. (C) Volcano plot of ITGB7-associated DEGs. Red indicates upregulated genes, green indicates downregulated genes, and gray indicates non-significant genes. (D) GO enrichment analysis of DEGs related to ITGB7. The Top 10 terms for Biological Process (BP), Cellular Component (CC), and Molecular functions (MF) were shown. (E) KEGG pathway analysis of DEGs associated with ITGB7. The top 10 pathways were mapped. (F) Immunofluorescence of IFN-γ in patients with ITGB7 high and low expression in our clinical cohort. *, p<0.05. |
Differential expression analysis using TCGA data identified 635 DEGs between high and low ITGB7 expression groups (Figure 4C). The overlapping molecules between DEGs from TCGA data and proteomic DEPs from PANC-1 cells are presented in Supplementary Figure 4. GO analysis of these DEGs highlighted their involvement in T cell activation, lymphocyte proliferation, and mononuclear cell proliferation (Figure 4D). KEGG analysis showed significant enrichment of DEGs in multiple immune-related pathways (Figure 4E). We further evaluated IFN-γ expression in tumor tissues from our clinical samples. Immunofluorescence analysis revealed higher IFN-γ levels in the high ITGB7 expression group (Figure 4F). These results collectively demonstrate that ITGB7 plays a pivotal role in modulating inflammation and immune signaling pathways.
ITGB7 Correlates with the Expression of Immune and Inflammatory Regulatory Molecules
We further investigated the correlation between ITGB7 expression and immune checkpoint molecule (ICM) across pan-tumors in the TCGA database. Heatmap analysis showed significant positive correlations between ITGB7 and ICMs in most tumor types (Figure 5A). In PDAC, ITGB7 expression positively correlated with PD1, PDL1, CTLA4, TIGIT, LAG3, and TIM3 (Figure 5B and Supplementary Figure 5). To elucidate the involvement of ITGB7 in physiological immune regulation and tumorigenesis, we further examined its correlation with ICMs in normal tissues. Similar results were observed in normal pancreatic tissues using the GTEx database (Supplementary Figure 6). Analysis of 41 pancreatic cancer cell lines from CCLE database also revealed a significant positive correlation between ITGB7 and PD-L1 expression (Figure 5C). In the PAAD dataset, patients with high ITGB7 levels exhibited higher levels of immune-stimulating molecules (eg, CD28, CD80, CD86) and inflammatory molecules (eg, TNF receptor superfamily members) compared to those with low ITGB7 levels (Figure 5D). High ITGB7 expression was also associated with increased MHC molecules, inflammatory chemokines, and chemokine receptors (Figure 5E-G). These findings suggest a strong positive association between ITGB7 and inflammatory/immune-regulatory molecules in PDAC.
 |
Figure 5 Correlation between ITGB7 and inflammatory and immune regulatory molecules. (A) Heatmap of correlations between ITGB7 and immune checkpoints across various tumors. Red represents positive correlation, blue represents negative correlation, and green represents the significance of correlations. *, p<0.05; **, p<0.01; ***, p<0.001. (B) Scatter plot of correlations between ITGB7 and ICMs in pan-tumor analyses. Each point represents a tumor type. The arrow highlights pancreatic cancer. Purple indicates significant correlations, gray indicates non-significant correlations. (C) Scatter plot of the correlation between ITGB7 and PD-L1 in pancreatic cancer cell lines in CCLE database. (D) Heatmap of immune stimulatory and inflammatory molecules, MHC molecule (E), inflammatory chemokine factors (F), and chemokine receptor (G) in PDAC patients with high vs low ITGB7 expression. |
ITGB7 Correlates with Increased Immune Cell Infiltration
We compared cancer immunity cycle scores between patients with high vs low ITGB7 expression in TCGA. High ITGB7 expression showed significantly enhanced recruitment of CD4+ T cells, dendritic cells, macrophages, and NK cells (Figure 6A). Moreover, ITGB7 expression was strongly correlated with DCs, B cells, and neutrophils infiltration (Figure 6B).
 |
Figure 6 ITGB7 expression in PDAC correlates with immune cell infiltration and PDL1 treatment prognosis. (A) Cancer immune cycle score for patients with high vs low ITGB7 expression in TCGA. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.001. (B) Correlation analysis of ITGB7 expression levels with immune cells infiltration in tumor tissues. (C) Prediction of small molecule drugs sensitivity in patients with high versus low ITGB7 expression. IC50, half-maximal inhibitory concentration. (D) IC50 values of Cisplatin, ATRA, Erlotinib, and Docetaxel in control and ITGB7-OE PANC-1 cells. |
Drug sensitivity analysis revealed that the high ITGB7 group exhibited enhanced response to chemotherapy agents (cisplatin, docetaxel, all-trans retinoic acid) and TKIs (axitinib, sunitinib, erlotinib) (Figure 6C and Supplementary Figure 7). Overexpression of ITGB7 was correlated with reduced IC50 values for cisplatin, docetaxel, and all-trans retinoic acid in PANC-1 cells, indicating increased drug sensitivity (Figure 6D). These findings indicate a positive correlation between ITGB7 expression and immune cell infiltration, as well as increased sensitivity to certain agents.
Construction of ITGB7 Associated Co-Expression Network
To analyze the regulatory characteristics of ITGB7 expression, we systematically revealed the dynamic association between the differential expression profile and the ceRNA regulatory network. Differential expression analysis identified 1285 mRNAs, 449 lncRNAs, and 46 miRNAs with significant changes between high vs low ITGB7 expression groups in TCGA (Supplementary Figure 8). Volcano plots in Figure 7A-C illustrate these differences. A ceRNA network was constructed by integrating interactions between the differential expressed RNAs (Figure 7D). Correlation between miRNAs and mRNA/lncRNA in the ceRNA network was shown in Supplementary Figure 9. Key network hubs included miR-150, miR-137, and miR-216 - known PDAC tumor suppressors. The ceRNA network offers valuable insights into the molecular mechanisms underlying the progression of PDAC.
 |
Figure 7 ITGB7-specific miRNA-associated ceRNA network in PAAD. (A–C) Volcano plots showing differentially expressed mRNAs, lncRNAs (B), and miRNAs (C) based on ITGB7 expression in the PAAD dataset. Red represents significantly up-regulated genes, green represents significantly down-regulated genes, and gray represents genes with no significant differences. (D) Graphical representation of the miRNA-lncRNA-mRNA network in PDAC at varying ITGB7 expression levels. Blue, mRNA; Green, miRNA; Red, lncRNA. |
The ITGB7-Related PPI Network is Associated with Immune Cell Infiltration
To systematically analyze the regulatory mechanism of ITGB7 in the tumor immune microenvironment, we constructed a core interaction network involving ITGB7 using the TCGA dataset and further investigated its role in immune regulation in PDAC. The ITGB7-centered PPI network (62 nodes, 317 edges) revealed direct interactions with IRF4, PTPRC, and PRKCH (Figure 8A). Expression patterns of network components were visualized through a heatmap (Figure 8B). These interactions showed strong positive correlations in TIMER database validation (Figure 8C). Notably, IRF4/PTPRC/PRKCH levels were significantly correlated with infiltration of B cells, CD4+/CD8+ T cells, macrophages, and DCs in PDAC (Figure 8D–F). Our findings suggest that the ITGB7 network plays a significant role in tumor immune regulation.
 |
Figure 8 PPI network associated with ITGB7. (A) PPI network of ITGB7constructed based onTCGA dataset. Interaction types are indicated by colored edges: blue (curated database), purple (experimentally validated), dark green (gene neighborhoods), light green (text mining), red (gene fusions), and light violet (protein homology). (B) Heatmap illustrating the differential expression of key nodes in the PPI network between high and low ITGB7 expression groups. (C) Correlation scatter plot of ITGB7 with its direct interacting genes. (D-F) Correlation of IRF4 (D), PTPRC (E), and PRKCH (F) expression with immune cell infiltration in the TIMER database. |
Discussion
ITGB7 may play a significant role in tumor immune regulation, but its specific role in PDAC remains unclear. Our study used proteomics and clinical data analysis to elucidate ITGB7’s function and its impact on prognosis in PDAC. We provide the first evidence that ITGB7 expression correlates with immune regulatory molecules, inflammatory response, and immune cell infiltration in PDAC. Moreover, ITGB7 promotes M2 macrophage polarization and is associated with improved immunotherapy response.
ITGB7 plays distinct roles in various tumors. In breast cancer, ITGB7 enhances immunotherapy and serves as a prognostic marker.18,19 In colorectal cancer, ITGB7 maintains antitumor immunity, inhibits tumorigenesis and correlates with longer patient survival.12 ITGB7 is highly expressed in pancreatic cancer cells compared to other integrins.20 Our analysis of clinical cohorts and TCGA data consistently revealed significant ITGB7 overexpression in PDAC tissues compared to normal pancreatic tissues.
Our quantitative proteomic analysis utilizing TMT and PRM methods21,22 revealed that ITGB7 overexpression significantly alters the proteomic profile of pancreatic cancer cells, primarily impacting immune processes, inflammatory responses, and drug metabolism pathways. Key DEPs regulated by ITGB7 include immune-related proteins (eg, HMGN1, ANXA1, ANXA4, and Tβ superfamily members), inflammatory response proteins (eg, TRAF2), and drug metabolism proteins (eg, CYCS and SURF1).
HMGN1 promotes DC recruitment and activation, enhancing Th1-type immunity.23 Members of the Annexin superfamily, such as Annexin A1, remodel the immune microenvironment in pancreatic cancer via the cGAS/STING pathway.24 Tβ superfamily stimulates immune responses, promoting lymphocyte proliferation, differentiation, and NK cell cytotoxicity.25,26 Proteomics analysis and PPI network of ITGB7 associated DEPs highlighted the critical role of ITGB7 in tumor immune regulation and inflammatory response modulation.
The upregulation of ICM expression positively correlated with increased lymphocyte infiltration and better prognosis in patients on ICI therapy.27 In our study, ITGB7 expression positively correlated with ICM expression in most tumors, particularly showing positive correlations with PD1, PDL1, CTLA4, TIGIT, LAG3, and TIM3 in PDAC. Our study also revealed a significant positive correlation between ITGB7 and PD-L1 expression in our clinical cohort as well as multiple pancreatic cancer cell lines.
The correlation between ITGB7 expression and immunotherapy response was further explored in our clinical cohort. We showed that PDAC patients with higher ITGB7 expression had significantly better response and a more favorable prognosis when treated with PD1 antibody. Survival analysis of TCGA data demonstrated that PDAC patients with high ITGB7 expression exhibited significantly prolonged overall survival when treated with PD-L1 antibodies, corroborating the findings from the clinical cohort.
Cytokines such as interleukins, interferons and chemokines, influence the tumor microenvironment and treatment efficacy. Pro-inflammatory cytokines such as IL-6, TNF-α, promote tumor growth and metastasis by activating key signaling pathways such as NF-kB and STAT3, which drive cell survival, proliferation, and resistance to apoptosis.28 Targeting these molecules shows therapeutic potential, and may help overcome resistance to conventional therapies.29,30 Our study showed that patients with high ITGB7 levels exhibited elevated expression of immune stimulating molecules, MHC molecules, inflammatory chemokines, and chemokine receptors in pancreatic tumors. The expression of IFN γ and expanded immune gene signatures respond positively to PD1 checkpoint blockade.31 GSVA analysis in our study linked ITGB7 to IFN-γsignaling response, inflammatory response and multiple inflammatory factor signaling pathways, including TNFα-NFκB and IL6-STAT3 signaling pathways. In our clinical cohort, high ITGB7 expression was associated with significantly higher IFN-γ levels, supporting the correlation between ITGB7 and IFN-γ. Our findings support the role of ITGB7 in regulating immune and inflammatory molecules.
M2-type TAMs impair immune surveillance by secreting immunosuppressive cytokines like IL10 and TGFβ. They also enhance cancer proliferation and invasion through cytokines such as IL6. Furthermore, M2-type TAMs produce angiogenic factors to induce angiogenesis.32,33 So far, the roles of M2 macrophages in carcinogenesis and immunotherapies remain incompletely understood due to their complexity and heterogeneity across different contexts. In certain tumors, such as gliomas, M2 macrophages may represent a promising target for immunotherapy.34 IL-10 and IL-6 secreted by tumor cells have been reported to promote M2 polarization.35 The observed increase in M2 macrophages in response to ITGB7-overexpressing cell supernatant implies that ITGB7 may modulate the tumor microenvironment in a manner conducive to immune evasion and tumor progression. ITGB7 expressed in pancreatic cancer cells may contribute to M2 macrophage polarization by enhancing the expression and secretion of IL-6 and IL-10. Immune checkpoint inhibitors may exert their function by inhibiting the immunosuppressive effect mediated by M2 macrophages in pancreatic cancer tissues overexpressing ITGB7.
The infiltration of immune cells is closely linked to immunotherapy responsiveness.36 High infiltration of CD4+ and CD8+ T cells is a favorable prognostic factor for patients with PDAC.37 In the TCGA dataset analysis, we found that ITGB7-associated DEGs were involved in T cell activation and lymphocyte proliferation. ITGB7 expression significantly enhances recruitment of CD4+ T cells, CD8+ T cells, neutrophils and DCs. Our research indicates that ITGB7 may promote the tumor immune response by modulating the infiltration of various immune cells.
Drug sensitivity analysis demonstrated ITGB7 expression enhanced responsiveness to chemotherapy agents such as cisplatin and docetaxel, as well as targeted therapies such as erlotinib. These results indicate that these drugs could be promising therapeutic options for patients with high ITGB7 expression.
Our study, for the first time, constructed a pancreatic cancer-specific ceRNA interaction network associated with ITGB7 that involved a number of regulators for tumor progression. Hub miRNAs include miR150, miR137, miR216a, miR216b, miR670, and miR96. Mir-150 interacts with lncRNAs, such as LINC00261 and PART1, as well as mRNAs like MYCNOS within the regulatory network, which are involved in cell cycle regulation. Experimental evidence has demonstrated that mir-150 influences tumor proliferation in colon cancer through its regulatory effects on the cell cycle.38 Mir-216b interacts with MIR7-3HG and LINC00469, and its sequence specifically targets oncogenes, including KRAS.39 miR137, miR96, and miR150 have been reported to be significantly down-regulated in PDAC and act as tumor suppressors affecting tumor growth, invasion and chemotherapy sensitivity.40–42 The identification of well-established PDAC tumor suppressor molecules which function as network hubs suggests a novel mechanism through which ITGB7 may influence tumor progression via an epigenetic regulatory axis. The ceRNA network highlights the potential role of ITGB7 in PDAC and offers new insights for future research.
The highly connected nodes in our PPI network, such as PTPRC, IRF, PRKCH, CD86, LCP2, and CD2, are key molecules involved in immune regulation, which suggests that ITGB7 may participate in complex immune regulatory mechanisms. PTPRC, also known as CD45, serves as a critical regulatory factor in the activation of T and B cell antigen receptors, linking multiple immune receptors, such as CD28 and CD2, to downstream signaling molecules.43 PRKCH is essential for the Treg-intrinsic CTLA4/PKCη signaling pathway that suppresses tumor immunity in melanoma.44 While the roles of IRF4, PTPRC, and PRKCH in PDAC have not been reported, our study shows that these genes are positively associated with infiltration of B cells, CD4+ T cells, CD8+ T cells, macrophages, and DCs in PDAC. This indicates potential involvement in immune regulation of ITGB7 within PDAC, providing insights into the immune regulatory network.
Our research has, for the first time, reported the abnormal expression of ITGB7 in PDAC and elucidated its role in TME and immunotherapy response. ITGB7 modulates immune and inflammation-related signaling pathways, and promotes M2 macrophage polarization in PDAC. ITGB7 is associated with favorable immunotherapy responsiveness. This study provides novel insights into the regulation of PDAC immune responses. Further validation with larger sample sizes is needed to confirm its association with prognosis to immunotherapy. Additionally, the underlying mechanisms by which ITGB7 influences immune responses in PDAC require further investigation.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The studies involving human participants were reviewed and approved by the Ethics Committee of Nanfang Hospital. Consent was obtained by the study participants prior to study commencement.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the grant from National Natural Science Foundation of China [82473012 & 82073174], GuangDong Basic and Applied Basic Research Foundation [2024A1515030242 & 2022A1515012406], and Science and Technology Projects in Guangzhou[SL2022A03J01176], Clinical Research Project of Nanfang Hospital, Southern Medical University[2021CR026&2024CR022].
Disclosure
The authors declare no competing interests in this work.
References
1. Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12–49. doi:10.3322/caac.21820
2. Minato N. Toward a new stage of PD-1 blockade cancer immunotherapy. Int J Clin Oncol. 2020;25(5):787–789. [1437-7772 (Electronic)]. doi:10.1007/s10147-020-01643-8
3. Farhangnia P, Khorramdelazad H, Nickho H, Delbandi AA. Current and future immunotherapeutic approaches in pancreatic cancer treatment. J Hematol Oncol. 2024;17(1):40. 1756-8722 (Electronic)
4. Baghban R, Roshangar L, Jahanban-Esfahlan R, et al. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun Signaling. 2020;18(1). 1478-811X (Electronic)). doi:10.1186/s12964-020-0530-4
5. Halbrook CJ, Lyssiotis CA, di Magliano MP, Maitra A. Pancreatic cancer: advances and challenges. Cell. 2023;186(8):1729–54. 1097-4172 (Electronic)
6. Ullman NA-O, Burchard PR, Dunne RF, et al. Immunologic strategies in pancreatic cancer: making cold tumors hot. J Clin Oncol. 2022;40(24):2789–2805. [1527-7755 (Electronic)]. doi:10.1200/JCO.21.02616
7. Huang R, Kang T, Chen S. The role of tumor-associated macrophages in tumor immune evasion. J Cancer Res Clin Oncol. 2024;150(5):238. 1432-1335 (Electronic).
8. Zhao H, Wu L, Yan G, et al. Inflammation and tumor progression: signaling pathways and targeted intervention. Signal Transduction Targeted Ther. 2021;6(1):263. 2059-3635 (Electronic)
9. Hynes RO. Integrins: versatility, modulation, and signaling in cell adhesion. Cell. 1992;69(1):11–25. 0092-8674 (Print).
10. Bianconi D, Unseld M, Prager GW. Integrins in the spotlight of cancer. Int J Mol Sci. 2016;17(12):2037. 1422-0067 (Electronic)
11. Sun H, Kuk W, Rivera-Nieves J, et al. β7 integrin inhibition can increase intestinal inflammation by impairing homing of CD25(hi)FoxP3(+) regulatory T cells. Cell Mol Gastroenterol Hepatol. 2020;9(3):369–85. 2352-345X (Electronic)
12. Zhang Y, Xie R, Zhang H, et al. Integrin β7 inhibits colorectal cancer pathogenesis via maintaining antitumor immunity. Cancer Immunol Res. 2021;9(8):967–980. [2326-6074 (Electronic)]. doi:10.1158/2326-6066.CIR-20-0879
13. Cheng Y, Wang K, Geng L, et al. Identification of candidate diagnostic and prognostic biomarkers for pancreatic carcinoma. EBioMedicine. 2019;40:382–393. 2352-3964 (Electronic). doi:10.1016/j.ebiom.2019.01.003
14. Cheng Y, Zheng H, Wang B, et al. Sorafenib and fluvastatin synergistically alleviate hepatic fibrosis via inhibiting the TGFβ1/Smad3 pathway. Digest Liver Dis. 2018;50(4):381–388. [1878-3562 (Electronic)]. doi:10.1016/j.dld.2017.12.015
15. Cheng Y, Zhu Y, Xu W, et al. PKCα in colon cancer cells promotes M1 macrophage polarization via MKK3/6-P38 MAPK pathway. Mol Carcinogenesis. 2018;57(8):1017–1029. [1098-2744 (Electronic)]. doi:10.1002/mc.22822
16. Li T, Fu J, Zeng Z, et al. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020;48(W1):W509–W514. [1362-4962 (Electronic)]. doi:10.1093/nar/gkaa407
17. Lu X, Jiang L, Zhang L, et al. Immune signature-based subtypes of cervical squamous cell carcinoma tightly associated with human papillomavirus type 16 expression, molecular features, and clinical outcome. Neoplasia. 2019;21(6):591–601. [1476-5586 (Electronic)]. doi:10.1016/j.neo.2019.04.003
18. Yang L, Wang M, Hu X, et al. EccDNA-oriented ITGB7 expression in breast cancer. Annals Transl Med. 2022;10(24):1344. [2305-5839 (Print)]. doi:10.21037/atm-22-5716
19. Xu T, Liu J, Xia Y, et al. Integrated analysis reveals the participation of IL4I1, ITGB7, and FUT7 in reshaping the TNBC immune microenvironment by targeting glycolysis. Annals Med. 2021;53(1):916–928. [1365-2060 (Electronic)]. doi:10.1080/07853890.2021.1937694
20. Farrow B, Rychahou P, O’Connor KL, et al. Butyrate inhibits pancreatic cancer invasion. J Gastrointestinal Surg. 2003;7(7):864–870. [1091-255X (Print)]. doi:10.1007/s11605-003-0031-y
21. Guo H, Guo H, Zhang L, et al. Dynamic TMT-based quantitative proteomics analysis of critical initiation process of totipotency during cotton somatic embryogenesis transdifferentiation. Int J Mol Sci. 2019;20(7):1691. [1422-0067 (Electronic)]. doi:10.3390/ijms20071691
22. Urisman AA-O, Levin RS, Gordan JD, et al. An optimized chromatographic strategy for multiplexing in parallel reaction monitoring mass spectrometry: insights from quantitation of activated kinases. Mol Cell Proteomics. 2017;16(2):265–277. [1535-9484 (Electronic)]. doi:10.1074/mcp.M116.058172
23. Yang D, Han Z, Alam MM, et al. High-mobility group nucleosome binding domain 1 (HMGN1) functions as a Th1-polarizing alarmin. Seminars Immunol. 2018;38:49–53. 1096-3618 (Electronic). doi:10.1016/j.smim.2018.02.012
24. Hou Z, Lu F, Lin J, et al. Loss of Annexin A1 in macrophages restrains efferocytosis and remodels immune microenvironment in pancreatic cancer by activating the cGAS/STING pathway. J ImmunoTher Cancer. 2024;12(9):e009318. [2051-1426 (Electronic)]. doi:10.1136/jitc-2024-009318
25. Lee H-R, Yoon SY, Kang H-B, et al. Thymosin beta 4 enhances NK cell cytotoxicity mediated by ICAM-1. Immunol Lett. 2009;123(1):72–76. [1879-0542 (Electronic)]. doi:10.1016/j.imlet.2009.02.008
26. Huff T, Müller CS, Otto AM, Netzker R, Hannappel E. beta-Thymosins, small acidic peptides with multiple functions. Int J Biochem Cell Biol. 2001;33(3):205–20. (1357-2725 (Print))
27. Hu -F-F, Liu C-J, Liu -L-L, et al. Expression profile of immune checkpoint genes and their roles in predicting immunotherapy response. Briefings Bioinfo. 2021;22(3). 1477-4054 (Electronic). doi:10.1093/bib/bbaa176
28. Tošić I, Frank DA. STAT3 as a mediator of oncogenic cellular metabolism: pathogenic and therapeutic implications. Neoplasia. 2021;23(12):1167–1178. [1476-5586 (Electronic)]. doi:10.1016/j.neo.2021.10.003
29. Yi M, Li T, Niu M, et al. Targeting cytokine and chemokine signaling pathways for cancer therapy. Signal Transduction Targeted Ther. 2024;9(1):176. 2059-3635 (Electronic)
30. Vilgelm AE, Richmond A. Chemokines modulate immune surveillance in tumorigenesis, metastasis, and response to immunotherapy. Front Immunol. 2019;10:333. 1664-3224 (Electronic).
31. Ayers M, Lunceford J, Nebozhyn M, et al. IFN-γ-related mRNA profile predicts clinical response to PD-1 blockade. J Clin Investig. 2017;127(8):2930–2940. [1558-8238(Electronic)]. doi:10.1172/JCI91190
32. Zhang Y, Yan W, Collins MA, et al. Interleukin-6 is required for pancreatic cancer progression by promoting MAPK signaling activation and oxidative stress resistance. Cancer Res. 2013;73(20):6359–6374. [1538-7445 (Electronic)]. doi:10.1158/0008-5472.CAN-13-1558-T
33. Chen P, Huang Y, Bong R, et al. Tumor-associated macrophages promote angiogenesis and melanoma growth via adrenomedullin in a paracrine and autocrine manner. Clin Cancer Res. 2011;17(23):7230–7239. [1557-3265 (Electronic)]. doi:10.1158/1078-0432.CCR-11-1354
34. Ye F, Wang L, Li Y, et al. IL4I1 in M2-like macrophage promotes glioma progression and is a promising target for immunotherapy. Front Immunol. 2024. 14. 1664-3224 (Electronic). doi:10.3389/fimmu.2023.1338244
35. Bingle L, Brown NJ, Lewis CE. The role of tumour-associated macrophages in tumour progression: implications for new anticancer therapies. J Pathol. 2002;196(3):254–65. 0022-3417 (Print)
36. Jochems C, Schlom J. Tumor-infiltrating immune cells and prognosis: the potential link between conventional cancer therapy and immunity. Exp Biol Med. 2011;236(5):567–579. [1535-3699(Electronic)]. doi:10.1258/ebm.2011.011007
37. Fukunaga A, Miyamoto M, Cho Y, et al. CD8+ tumor-infiltrating lymphocytes together with CD4+ tumor-infiltrating lymphocytes and dendritic cells improve the prognosis of patients with pancreatic adenocarcinoma. Pancreas. 2004;28(1):e26–e31. [1536-4828 (Electronic)]. doi:10.1097/00006676-200401000-00023
38. Zhang ZC, Wang GP, Yin LM, Li M, Wu LL. Increasing miR-150 and lowering HMGA2 inhibit proliferation and cycle progression of colon cancer in SW480 cells. Eur Rev Med Pharmacolog Sci. 2018;22(20):1. 2284-0729 (Electronic)
39. Wu X, Chen W, Cai H, et al. MiR-216b inhibits pancreatic cancer cell progression and promotes apoptosis by down-regulating KRAS. Arch Med Sci. 2018;14(6):1321–1332. [1734-1922 (Print)]. doi:10.5114/aoms.2018.72564
40. Xiao J, Peng F, Yu C, et al. microRNA-137 modulates pancreatic cancer cells tumor growth, invasion and sensitivity to chemotherapy. Int J Clin Exp Pathol. 2014;7(11):7442. 1936-2625 (Electronic)
41. Yu S, Lu Z, Liu C, et al. miRNA-96 suppresses KRAS and functions as a tumor suppressor gene in pancreatic cancer. Cancer Res. 2010;70(14):6015–6025. [1538-7445 (Electronic)]. doi:10.1158/0008-5472.CAN-09-4531
42. Srivastava SK, Bhardwaj A, Singh S, et al. MicroRNA-150 directly targets MUC4 and suppresses growth and malignant behavior of pancreatic cancer cells. Carcinogenesis. 2011;32(12):1832–1839. [1460-2180 (Electronic)]. doi:10.1093/carcin/bgr223
43. Rheinländer A, Schraven B, Bommhardt U. CD45 in human physiology and clinical medicine. Immunol Lett. 2018;196:22–32. [1879-0542 (Electronic)]. doi:10.1016/j.imlet.2018.01.009
44. Pedros C, Canonigo-Balancio AJ, Kong K-F, et al. Requirement of Treg-intrinsic CTLA4/PKCη signaling pathway for suppressing tumor immunity. JCI Insight. 2017;2(23). doi:10.1172/jci.insight.95692
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Significance of Immunogenic Cell Death-Related Prognostic Gene Signature in Cervical Cancer Prognosis and Anti-Tumor Immunity
Jiang S, Cui Z, Zheng J, Wu Q, Yu H, You Y, Zheng C, Sun Y
Journal of Inflammation Research 2023, 16:2189-2207
Published Date: 22 May 2023
Interleukin Family-Based Signature Relates to Cancer-Associated Fibroblasts Spatial Distribution and Immune Therapy Response in Pancreatic Carcinoma
Cheng Y, Xiao S, Li X, Wang B, Lei Y, Sun P, Ma L, Zhu Y
Journal of Inflammation Research 2025, 18:10129-10146
Published Date: 29 July 2025