Back to Journals » Journal of Inflammation Research » Volume 18
Isoorientin Modulates Gut Microbes and Their Metabolites to Alleviate Caco-2 Cell Monolayer Inflammation by Reducing Intestinal Permeability via P-Gp/eCBs
Authors Wang Z, Jiang Y ![]() , Duan B, Zhang H, Li A, Yang J, Lv M, Yang J
, Duan B, Zhang H, Li A, Yang J, Lv M, Yang J
Received 26 March 2025
Accepted for publication 9 September 2025
Published 16 October 2025 Volume 2025:18 Pages 14365—14379
DOI https://doi.org/10.2147/JIR.S530562
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Nadia Andrea Andreani
Zhenzhen Wang,1 Yuang Jiang,2 Bensong Duan,3 Haibin Zhang,3 Aoxiang Li,1 Jin Yang,1 Mengke Lv,1 Jingya Yang1,4
1College of Food Science and Technology, Shanghai Ocean University, Shanghai, 201306, People’s Republic of China; 2College of Fisheries and Life Science, Shanghai Ocean University, Shanghai, 201306, People’s Republic of China; 3Department of Gastroenterology Shanghai East Hospital, School of Medicine, Endoscopy Center, Tongji University, Shanghai, People’s Republic of China; 4Marine Biomedical Science and Technology Innovation Platform of Lin-Gang Special Area, Shanghai, 201306, People’s Republic of China
Correspondence: Jingya Yang, College of Food Science and Technology, Shanghai Ocean University, No. 999 Hu-Cheng Huan Road, Shanghai, 201306, People’s Republic of China, Email [email protected]
Introduction: Isoorientin (ISO) is a naturally occurring flavonoid that studies have shown to have strong experimental antioxidant, anti-inflammatory, anti-diabetic and anti-obesity properties. It has been shown that ISO alleviates Dextran sodium sulfate (DSS) induced colitis in mice by modulating gut microbes and their metabolites. The aim of this study was to modulate gut microbes and their metabolism by ISO to investigate its anti-inflammatory effects and its specific molecular mechanisms in a lipopolysaccharide (LPS)-induced monolayer inflammation model in Caco-2 cells.
Methods: Feces from ISO-treated DSS colitis mice were collected and gut flora culture supernatants were prepared. Detection of the effect of intestinal flora supernatants on the monolayer barrier of Caco-2 cells by inoculation of Caco-2 cells into the Transwell transmembrane culture system to simulate the intestinal mucosal barrier.
Results: The results revealed that ISO-conditioned intestinal flora supernatant significantly increased TEER values, decreased intestinal epithelial FITC-dextran flux permeability, and restored LPS-induced occludin, ZO-1 protein expression in Caco-2 cells. Meanwhile, intestinal flora supernatant significantly ameliorated the LPS-induced inflammatory response. In addition, ISO further enhanced its protective effect on intestinal permeability by regulating the expression of P-glycoprotein (P-gp) and endogenous cannabinoid system (eCB)-related proteins, which may attenuate the inflammatory response by activating the P-gp/eCB signaling pathway.
Conclusion: The present study offers fresh perspectives into the application of ISO-conditioned intestinal flora supernatant as a potential anti-inflammatory agent and intestinal barrier protector in vitro. The unique regulation of the P-gp/eCBs pathway by ISO-conditioned intestinal flora supernatant was the novel mechanistic insights provided in this study.
Keywords: isoorientin, Caco-2 cell, intestinal mucosal barrier, ZO-1, occludin, P-glycoprotein
Introduction
Inflammatory bowel disease (IBD) is characterized by inflammatory damage caused by intestinal flora and is predisposed to occur in genetically susceptible individuals presenting with intestinal epithelial barrier (IEB) dysfunction. Intestinal epithelial cells (IECs) are the major players in maintaining the integrity of the intestinal mucosal barrier. A compromised IEB is a key factor contributing to the development of IBD and other gut-related inflammatory disorders. In the pathological state of IBD, different pathogenic factors including bacteria, pathogens and other antigens may trigger the overproduction of inflammatory factors by the IECs, which in turn leads to an uncontrolled state of intestinal inflammation.1 The absence of the intestinal mucosal barrier due to the damage of the IECs allows a large number of bacterial flora in the lumen of the intestinal cavity to come into direct contact with the intestinal mucosa, resulting in the translocation of bacteria from the lumen of the intestinal cavity to the bloodstream and organs, which in turn leads to the systemic inflammatory response.2–4
The integrity of the IEB largely depends on tight junction (TJ) proteins which are apical multiprotein structures. These TJs link adjacent epithelial cells at the apical region of the lateral membrane, connecting transmembrane proteins to the intracellular actin cytoskeleton. This interaction is essential for controlling paracellular permeability and ensuring the stability of the epithelial layer.5 The expression and distribution of TJs in IECs are influenced by a range of both intracellular and extracellular factors, including inflammatory cytokines, gut microbes and their metabolites. These factors play a dynamic role in modulating TJ function.6–8 Since damage to the IEC TJ barrier caused by different pathogens promotes the development of IBD by increasing intestinal permeability,9 prevention of intestinal diseases by maintaining the barrier function of the IECs is necessary and particularly important.
Lipopolysaccharide (LPS) is a key factor in triggering both intestinal and systemic inflammation, and it has been demonstrated that LPS disrupts the structure of the TJ and induces, directly or indirectly, mucosal hyperpermeability through the secretion of cytokines and inflammatory mediators.10,11 In addition, persistent inflammatory stimulation leads to impaired intestinal TJ barrier function, which in turn produces proinflammatory cytokines, creating a vicious cycle.12 Relevant studies have shown that intestinal microecological disorders can produce strong inflammatory substances such as LPS, which can increase the permeability of the IEB, leading to “intestinal leakage” and allowing a variety of disease-causing substances to escape to the peripheral circulatory system, causing systemic inflammation.13 Consequently, strategies aimed at minimizing the inflammatory response could serve as potential therapeutic approaches for treating IBD. Several in vitro studies have demonstrated that natural compounds, such as vitamin A, can reverse LPS-induced damage to the intestinal barrier.10 Extracts from Boswellia serrata and turmeric have shown a protective effect on the intestinal epithelium, evidenced by increased transepithelial electrical resistance (TEER) values compared with LPS-treated cells.14 Additionally, a combination of immobilized probiotics and herbal extracts was found to prevent inflammation-induced reductions in TEER, paracellular permeability, and TJ protein translocation.15 Paeoniflorin markedly elevated TEER levels, reduced FITC-dextran flux permeability and restored the expression of occludin, zinc occludens-1 (ZO-1) and claudin-5 in Caco-2 cells following LPS exposure.16
The proper functioning of the gut relies on the formation and preservation of a mucosal barrier, which consists of a single layer of IECs. The establishment of a normal gut microbiota is crucial for the development and continued maintenance of this epithelial barrier. An increasing number of studies indicate that gut microbiota plays a crucial role in the initiation and progression of intestinal inflammation in IBD.4,17 A large number of microorganisms parasitizing the organism’s intestinal tract constitute the biological barrier of the intestinal mucosa and maintain a dynamic equilibrium in the normal intestinal environment, and the production of these microorganisms and their metabolites, short-chain fatty acids (SCFAs), plays an important role in the maintenance of the intestinal barrier.18,19 There are some innovative therapeutic strategies reported to aim at efficiently restoring IEB integrity. Zhang et al reported that tannic acid and zinc ion coordination of nanase for the treatment of IBD by removing reactive oxygen and nitrogen species.20 In addition, CO-releasing polyoxometalates nanozyme with gut mucosal immunity and, microbiota homeostasis Remodeling was able to restore IEB integrity.21
Flavonoids are important components of the human diet and are widely found in a variety of edible plants. Metabolic conversion of dietary flavonoids by bacteria in the intestinal tract may provide potential benefits to human health. Isoorientin (ISO), a naturally occurring C-glycoside flavonoid, has demonstrated a variety of pharmacological effects. Our previous study have found that ISO is able to alleviate DSS-induced mice and has a modulatory effect on gut microbes and their metabolites in mice with colitis, markedly increasing the content of gut microbial metabolites SCFAs and secondary bile acids (SBAs), and regulating P-glycoprotein (P-gp) expression to alleviate colonic inflammation.22 Production of SCFAs and SBAs in the intestine is important in the treatment of colonic inflammation and may also play a significant role in preserving the integrity of the intestinal barrier.5,23
eCBs are strongly associated with the production and resolution of inflammation.24,25 P-gp, a transmembrane pump with broad substrate specificity, is able to balance the expression of relevant receptor proteins in the eCB system and thus participate in inflammatory processes.24 Modulation of the P-gp/eCB axis and increased P-gp expression reduce intestinal inflammation, thereby decreasing IEB permeability.24,26,27 In the present study, the IEB inflammation model was simulated by LPS-induced Caco-2 cells in vitro, and ISO-regulated intestinal fecal supernatant was co-cultured with the Caco-2 cell model. The effects of ISO-regulated intestinal microbial metabolites on inflammatory response and P-gp/eCB axis-related protein expression were investigated, and an experimental basis was provided for ISO to regulate P-gp expression through intestinal microbial metabolites, reduce intestinal permeability and regulate the intestinal barrier to alleviate colitis.
Materials and Methods
Materials
The purity of ISO was ≥98% and it was provided by the Chinese Academy of Sciences (Shanghai, China). A total of 21 6-7-week-old female SPF class C57BL/6 mice weighing 18–20 g, were provided by Spearfish (Beijing) Biotechnology Co. LPS from Salmonella enterica was sourced from Sigma-Aldrich (CAS:93572–42-0); Merck KGaA. The cell culture media, which included fetal bovine serum (FBS), trypsin/EDTA and antibiotics (penicillin/streptomycin), was acquired from Gibco; Thermo Fisher Scientific, Inc (Cat:21773-1-AP). Antibodies targeting occluding (Cat:21773-1-AP), ZO-1 (Cat:27260-1-AP) and cannabinoid receptor 2 (CB2, Cat:29371-1-AP) were purchased from Wuhan Sanying Biotechnology, while P-gp (Cat:13978) and GAPDH (Cat:2118) antibodies were obtained from Cell Signaling Technology, Inc. Secondary antibodies conjugated to goat anti-rabbit IgG and Alexa Fluor® 488 (Cat:SA00001-2) were supplied by Wuhan Sanying Biotechnology. All additional reagents were provided by Sigma-Aldrich; Merck KGaA.
Animal Experimentation
C57BL/6 mice were randomly divided into a control group (CON), a model group (DSS, MP Biomedicals, CAS:9011–18-1) and an ISO group (ISO; n=7). All mice were given normal water for the first 7 days. On the second set of 7 days, mice in all groups, except for the CON group, continued to drink 2.5% DSS solution ad libitum for 7 days, and the experimental cycle totaled 14 days. During the whole experimental period, the ISO group was treated with 50 mg/kg/day ISO by gavage, while the CON and DSS groups were given the same dose of 0.5% CMC-Na (Accela, CAS:9004–32-4) solution by gavage. On the last day of administration, the feces of mice from different groups were taken separately and stored at −80°C.
Fecal Supernatant Preparation
Fecal supernatants from mice were prepared following previously established protocols.25 Fecal samples, freshly collected and stored at −80°C, were first weighed and then resuspended in serum-free growth medium to a concentration of 0.25 g/mL (wt/vol). These samples were homogenized using gentle vortexing and manual grinding with a sterile pipette tip, followed by centrifugation at 10,000×g for 15 min. The resulting supernatant was filtered aseptically through a 0.22 μm polyethersulfone filter and subsequently diluted in serum-free DMEM High Glucose Medium, making the samples suitable for cellular assays.
Cell Culture
Generation 22–60 human IEC Caco-2 cells (ATCC) were grown at 37°C and 5% CO2 in DMEM containing 10% FBS and 1% penicillin-streptomycin. The medium was changed every 2 days. Monolayers grew on collagen-coated tissue culture-treated 6-well plates and were used 7–9 days after inoculation to induce an inflammatory response according to a previous study.16 After 2 h of incubation, fecal supernatant, 5 mM butyric acid (CAS:9024–15-1) or 50 μM deoxycholic acid (CAS:83–44-3), ursodeoxycholic acid (CAS:128–13-2) and lithocholic acid (CAS:434–13-9) (Sigma-Aldrich; Merck KGaA) were added, and metabolite concentrations were as previously described.28 Cells were collected after 24 h for further analysis. The cells were deprived of serum by incubating in serum-free DMEM growth medium for 12 h before being exposed to fecal supernatant and metabolites.
Cell Viability Assay
The Cell Counting Kit-8 (CCK-8, CAT:04483231) assay was performed following the manufacturer’s guidelines (Adamas, China). Caco-2 cells were seeded in 96-well plates at a density of 1×104 cells/well and incubated for 24 h. Then, the cells were treated with LPS (0, 0.1, 1, 10, 20 and 50 μg/mL) or different groups of fecal supernatants (at concentrations of 0, 1, 10, 25 and 50 mg/mL) by incubated for 24 h. The cells were discarded. After 24 h of fecal supernatants treatment (FST), the supernatant was discarded, and 110 μL CCK-8 diluted with DMEM was added to each well, and then the cells were incubated in the incubator for 2 h. The absorbance at 450 nm was measured by a full-wavelength zymography and cell viability was calculated. A total of 10 μL CCK-8 solution was added to each well and cells were incubated for an additional 2 h. Following this, the absorbance was measured at 450 nm using a microplate reader to assess cell viability. Each experiment was performed five times.
ELISA
In this experiment, Caco-2 cells were plated in 6-well plates at a density of 2.5×105 cells/mL and were cultured until confluence was achieved. Fecal supernatant was then added; 10 mg/mL fecal supernatant and 10 μg/mL LPS were added to the cells in the following three different groups, and the fecal supernatant was pretreated for 2 h if LPS was stimulated for 24 h. At the end of the intervention, the culture supernatant was aspirated, and the levels of the cell-secreted cytokines TNF-α (CAT:JM-02415M2) and IL-6 (CAT:JM-02446M2) were determined by ELISA fractionation (Jinmei Biotechnology, China); the cells were then assessed using ELISA kits, following the procedures outlined by the manufacturer. The levels of IL-6 and TNF-α were measured in cell supernatants.
Reverse Transcription-Quantitative PCR (RT-qPCR)
Total RNA was isolated from 2×105 cultured cells using 1 mL TRIzol reagent (Beyotime, China, CAT:R0016). Subsequently, cDNA was generated from 1 μg RNA using M-MLV reverse transcriptase, and 100 ng synthesized cDNA was then amplified. cDNA was quantified in real time with the BIO-RAD system (Bio-Rad Laboratories, Inc.) according to the manufacturer’s instructions, and qPCR was performed using the SuperStar Universal SYBR Master Mix kit (Kangwei Reagent, China, CAT:CW3371M). The thermocycling conditions used were the following: Initial cycle at 95°C for 30 sec, followed by 45 cycles of 95°C for 15 sec and 60°C for 30 sec. A melting curve analysis was performed to evaluate the specificity of the products. Each sample was analyzed in triplicate, and the CT values for the target transcripts were recorded. Gene expression levels were normalized to those of the housekeeping gene GAPDH using the 2-ΔΔCq method. The primers used for qPCR are listed in Table 1. qPCR was used to measure relative mRNA expression of the pro-inflammatory markers TNF-α and IL-6 to determine the anti-inflammatory status of fecal supernatants subjected to epithelial permeability testing.
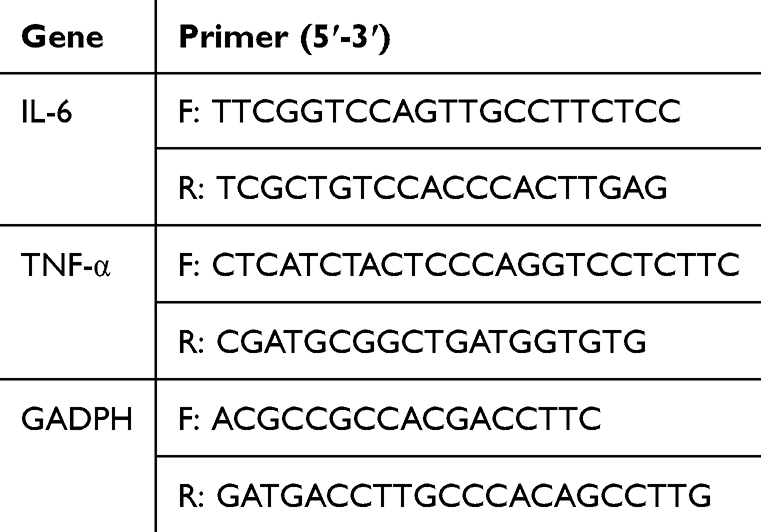 |
Table 1 PCR Primer Sequences for Each Gene |
Measurement of TEER and Permeability
Cells with good growth status were taken, trypsin digested to prepare cell suspension, inoculated in 24-well Transwell plates (polycarbonate membrane with 0.4 μm pore size) at a concentration of 1×104 cells/mL, 200 μL cell suspension was added to the AP test and 600 μL complete medium was added to the BL side; cells were then incubated at 37°C and 5% CO2 for 21 days, and then the different groups of fecal supernatants with or without LPS (10 μg/mL) were added to the apical medium after 24 h of incubation. A TEER assay was performed as previously described.29 In both compartments, the medium of the Caco-2 cell monolayer was exchanged with PBS, and 0.1 mg/mL FITC-Dextran (Beyotime Biotechnology, China, CAT:ST2935) was introduced to the apical side of the Transwell. After incubating for 1 h with the FITC-Dextran, the medium from the basolateral side of the Transwell was collected. Fluorescence was then quantified using a fluorescence microplate reader (Thermo Fisher Scientific, Inc)., with excitation set at 480 nm and emission measured at 520 nm.
Western Blotting
Cells were lysed using RIPA buffer containing a protease inhibitor cocktail, and protein concentrations were determined using the BCA protein assay with bovine serum albumin as the standard. Proteins from each sample were then resolved by SDS-PAGE with 10–13% acrylamide gels and subsequently transferred to PVDF membranes. The proteins separated on the membrane were blocked with 5% skimmed milk for 1 h, and then subjected to immunoblotting with primary antibodies targeting GADPH (1:5000; Proteintech Group, Inc), TJ protein ZO-1 (1:5000; Proteintech Group, Inc), Occludin (1:5000; Proteintech Group, Inc), P-gp (1:1000; Cell Signaling Technology, Inc)., and CB2 (1:5000; Proteintech Group, Inc) overnight at 4°C. After washing the membranes three times with buffer containing 1% Tween-20 (TBST), the membrane was incubated with the appropriate secondary antibody (1:1000; Cell Signaling Technology) for 1 h at room temperature. The intensity of the bands was detected using Al600 images and quantified using Image J (version 1.53; NIH).
Cellular Immunofluorescence Assay
Cells were cultured on coverslips within 24-well plates for 24 h. Subsequently, the cells were rinsed three times with PBS, fixed using 4% paraformaldehyde for 15 min, and blocked with 5% bovine serum albumin for 1 h at 37°C in a humidified chamber. Following another PBS wash, primary antibodies, including ZO-1 (1:300, Proteintech Group, Inc) and Occludin (1:300, Proteintech Group, Inc), were applied and incubated with the cells overnight at a 4°C humidified environment. The slides were then washed three times with PBS, and Alexa Fluor® 488-conjugated secondary antibody (1:500, Proteintech Group, Inc) was added for 2 h at room temperature. Cell nuclei were subsequently stained with DAPI. Finally, fluorescence microscopy was performed, and three random fields per slide were selected for observation.
Statistical Analysis
Data are presented as mean ± SEM. Statistical analysis was conducted using GraphPad Prism (version 9.5; Dotmatics). Group comparisons were performed using one-way ANOVA followed by Tukey’s post-hoc test. P<0.05 was considered to indicate a statistically significant difference.
Results
Effect of ISO-Treated Mouse Fecal Supernatant on Caco-2 Cell Viability
To determine the reparative effects of ISO-conditioned mouse fecal supernatants on LPS-induced Caco-2 cell cytotoxicity, the cytotoxicity of fecal supernatants or LPS on Caco-2 cell viability was first evaluated by treating them with different concentrations of fecal supernatants (0, 1, 10, 25 and 50 mg/mL) or LPS (0, 0.1, 1, 10, 20 and 50 μg/mL) toxicity effects by continuous co-incubation for 24 h. The results of CCK-8 assay suggested that fecal supernatant at a concentration of 25 mg/mL significantly reduced cell viability (P<0.05) compared with that of untreated cells after 24 h of incubation (Figure S1A–C). LPS affected cell viability at a concentration of 10 μg/mL (P<0.05; Figure S1D). It is suggested that fecal supernatant is not toxic to Caco-2 cells at concentrations ≤25 mg/mL for 24 h. Therefore, a 10-mg/mL dose of fecal supernatant was used with 10 μg/mL LPS in subsequent experiments.
{kind=link}
Next, the results after co-incubation with LPS-treated Caco-2 cells for 24 h with the action of mouse fecal supernatants from different treatment groups showed that the ISO-treated group (ISO-FST, DSS and ISO treated mouse) and CON group (CON-FST, no DSS treated) mouse fecal supernatants significantly reversed the reduction of Caco-2 cell viability caused by LPS compared with the LPS group, while the DSS-treated group (DSS-FST, DSS treated) fecal supernatant did not have this effect (Figure 1A). Meanwhile, the ISO-treated mouse fecal supernatant completely reversed the LPS-stimulated reduction in cell viability compared with the DSS-treated group (Figure 1A).
 |
Figure 1 Dose-effect fecal supernatant on Caco-2 cell viability and inhibition of LPS-induced inflammation reaction. (A) The cytotoxic effects of fecal supernatant on the Caco-2 cell viability were assessed by Cell Counting Kit-8 assay. (B and C) ELISA of Caco-2 secreted inflammatory factors IL-6 and TNF-α. (D) The mRNA expression of IL-6 and TNF-α was detected using reverse transcription-quantitative PCR. The data are presented as mean ± SEM. *P<0.05, **P<0.01, ***P<0.001, n=5. LPS, lipopolysaccharide. |
To further explore whether the regulation of gut microbial metabolites by ISO has the biological activity to inhibit colitis by studying the anti-inflammatory effect of ISO-treated mouse fecal supernatant on Caco-2 cells, 10 μg/mL LPS-induced inflammation in Caco-2 cells was treated using mouse fecal supernatant from different treatment groups. Our results showed that the expression of the proinflammatory factors IL-6 and TNF-α was increased after LPS stimulation of Caco-2 cells, and the cecal supernatant of the ISO-treated group significantly reduced the secretion of IL-6 and TNF-α induced by LPS into the culture medium from Caco-2 cells, whereas the DSS-treated group did not reduce the secretion of inflammatory cytokines by Caco-2 cells (Figure 1B and C). The changes in mRNA expression of IL-6 and TNF-α were also detected by RT-qPCR, and the results showed that LPS stimulation increased the expression of mRNA levels of IL-6 and TNF-α in cells, and the treatment of cecal supernatants in the DSS group exacerbated this effect, whereas the cecal supernatants in the ISO group reversed these upregulations (Figure 1D). It suggests that ISO-treated mouse fecal supernatant is able to attenuate LPS-stimulated inflammatory responses in Caco-2 cells.
Effect of ISO-Treated Mouse Fecal Supernatant on LPS-Induced Permeability of Caco-2 Monolayer Cells
TEER was employed to assess the integrity of the Caco-2 cell monolayer, and cell monolayers were complete and TJ formed when the TEER value was >200 Ω/cm2. The TEER value increased gradually with the increase of culture time and reached a plateau on day 9 until day 11, when the TEER value reached 1188 Ω/cm2 (Figure 2A).
 |
Figure 2 TJ protein damage in Caco-2 cells caused by LPS inhibition by ISO-regulated microbial metabolites. (A) TEER values to detect Caco-2 monolayer barrier formation. (B) Caco-2 cell large solute permeability was detected in each group by using FITC-dextran flux experiment. (C–H) Western blotting of occludin and ZO-1 protein expression in Caco-2 cells treated with different concentrations of LPS. (F–H) Western blotting of occluding and ZO-1 protein expression in fecal supernatant-treated Caco-2 cells. (I) Immunofluorescence detection of occludin and ZO-1 protein expression in fecal supernatant-treated Caco-2 cells (magnification, x40). The data are presented as mean ± SEM. *P<0.05, **P<0.01, ***P<0.001, n=3. |
To determine the effect of different treatments of mouse fecal supernatant on the permeability of monolayer Caco-2 cells, FITC-Dextran permeability was used to detect the permeability of Caco-2 cells to macromolecular solutes. The results showed that the FITC-Dextran permeability of Caco-2 cells was increased after induction by LPS stimulation, and the FITC-Dextran concentration was significantly higher compared with the CON group, and compared with the ISO fecal supernatant group. In addition, treatment in the ISO fecal supernatant group significantly reduced LPS-stimulation-induced FITC-dextran leakage compared with the CON and DSS fecal supernatant groups (Figure 2B).
Effect of ISO-Treated Mouse Fecal Supernatant on LPS-Induced ZO-1 and Occludin in Caco-2 Cells
The results of the effect of LPS on the expression of ZO-1 and occludin in Caco-2 cells determined by Western blotting showed that 10–50 μg/mL LPS significantly reduced the expression of ZO-1 and occludin in Caco-2 cells, and the difference was statistically significant (P≤0.05) compared with that in the control group (Figure 2C–E), and of which 10 μg/mL LPS had a significant effect, suggesting that LPS could break the TJ structure between Caco-2 cells to some extent.
To investigate the effect of ISO-treated mouse fecal supernatant on the LPS-induced breakage of Caco-2 TJ structure, the expression of ZO-1 and occludin was measured in Caco-2 cells. Western blotting results showed that the expression of ZO-1 and occludin, TJ proteins, in Caco-2 cells was significantly reduced in the LPS group (Figure 2F–H), suggesting that the TJ structure of Caco-2 cells was broken, while the expression of occludin and ZO-1 could be restored after treatment in the ISO fecal supernatant-treated group. Meanwhile, compared with the DSS-treated group, the ISO group also significantly increased the expression of occludin and ZO-1, suggesting that the ISO-treated mouse fecal supernatant could significantly repair the LPS-induced breakage of the TJ structure of Caco-2 cells. In addition, the results of ZO-1 and occludin immunofluorescence analyses were consistent with the Western blotting results (Figure 2I). In conclusion, these data suggest that ISO fecal supernatant maintains Caco-2 cell integrity by inhibiting LPS-induced loss of TJ proteins in Caco-2 cells.
Effect of ISO-Conditioned Fecal Supernatant on P-Gp/eCB-Related Proteins in Caco-2 Cells
We next investigated the expression of P-gp/eCB-related proteins in Caco-2 cells by Western blotting. As a result, the expression of P-gp and CB2 proteins in Caco-2 cells was significantly decreased after induction by LPS stimulation (P≤0.05), while the treatment with ISO fecal supernatant (10 mg/mL) significantly reversed the aforementioned response and increased the expression of P-gp and CB2 (Figure 3A–C). In addition, the ISO fecal supernatant-treated group significantly increased the expression of P-gp and CB2 compared with the DSS group-treated group. These results suggest that the ISO fecal supernatant may ameliorate LPS-induced inflammation in Caco-2 cell monolayers by modulating the expression of P-gp/eCB axis-related proteins.
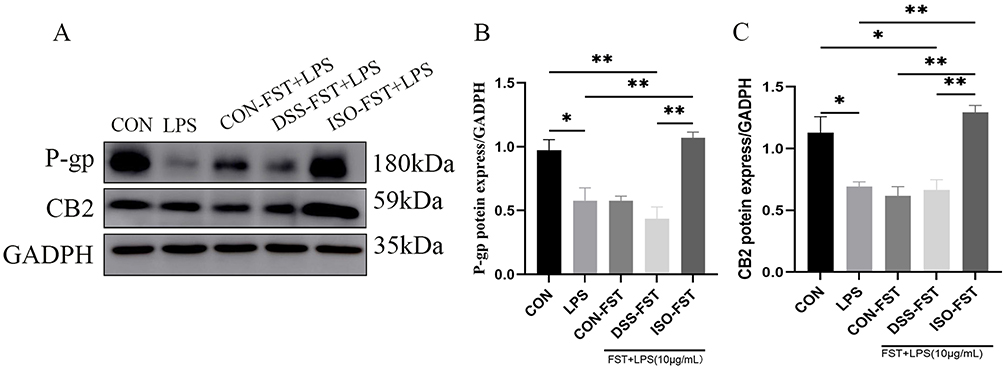 |
Figure 3 ISO-regulated microbial metabolites regulate the expression of P-gp/eCB axis-related proteins P-gp and CB2. (A–C) Western blotting detected the expression of P-gp and CB2 proteins inhibited by LPS by ISO-treated microbial metabolites. The data are presented as mean ± SEM. *P<0.05, **P<0.01, n=3. Abbreviations: ISO, isoorientin; P-gp, P-glycoprotein; eCB, endogenous cannabinoid system; CB2, cannabinoid receptor 2. |
ISO-Regulated Microbial Metabolites Alleviate LPS-Induced Structural Damage and Inflammatory Response in Caco-2 Cells by Modulating P-Gp/eCBs
In the previous results it was found that the ISO-treated mouse fecal supernatant could exert a reparative effect on LPS-induced inflammation in damaged Caco-2 cells and was highly associated with a decrease in the expression of P-gp/eCB-related proteins. Meanwhile, previous experiments showed that the metabolites of mouse intestinal flora regulated by ISO mainly included butyric and deoxycholic acids, lithocholic acid and ursodeoxycholic acid.22 Here, in order to verify that ISO-regulated metabolites of intestinal flora can enhance the expression of P-gp/eCB-related proteins thereby realizing the repairing effect on the TJ structure of Caco-2 cells as well as attenuating the inflammatory response, the P-gp protein inhibitor verapamil with different concentrations of chemobutyric acid, deoxycholic acid (DCA), lithobionic acid (LCA) and ursodeoxycholic acid (UDCA) was used alone or in combination to treat LPS-induced Caco-2 cells. The effects of ISO-regulated microbial metabolites on LPS-induced inflammation and the expression of the TJ proteins ZO-1 and occludin in Caco-2 cells were observed. The results showed that the P-gp inhibitor verapamil could not alter the LPS-stimulated inflammatory response in Caco-2 cells, whereas the four major microbial metabolites regulated by ISO significantly reduced the secretion of IL-6 and TNF-α in Caco-2 cells (Figure 4A and B). It is suggested that the expression of the ISO-regulated P-gp affects the inflammatory response of Caco-2 cells.
 |
Figure 4 Microbial metabolites inhibit LPS-induced inflammation and TJ protein damage in Caco-2 cells. (A and B) ELISA inhibited the expression of microbial metabolites against the inflammatory factors IL-6 and TNF-α induced in Caco-2 cells by LPS. (C) Western blotting detected the microbial metabolites that increased the expression of the TJ proteins ZO-1 and occludin in Caco-2 cells inhibited by LPS. (D–G). Western blotting detection of microbial metabolites to regulate the expression of P-gp/eCBs axis-related proteins P-gp and CB2. The data are presented as mean ± SEM. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001, n=3. Abbreviations: LPS, lipopolysaccharide; TJ, tight junction; ZO-1, zinc occludens-1; P-gp, P-glycoprotein; eCB, endogenous cannabinoid system; CB2, cannabinoid receptor 2. |
Western blotting showed that ISO-regulated microbial metabolites significantly ameliorated the LPS-induced decrease in ZO-1 and occludin expression, and the combination of the four major metabolites was even more pronounced in the rise of ZO-1 and occludin expression in Caco-2 cells (P≤0.05; Fig. 4C-3). In addition, the results of ISO-regulated microbial metabolites on the protein expression related to P-gp/eCBs showed that the combination of microbial metabolisms significantly increased the reduction of P-gp and CB2 protein expression in LPS-stimulated Caco-2 cells, and also after the use of verapamil, a P-gp inhibitor (Figure 4C–G). Notably, the combined intervention of the four major metabolites exhibited a significant synergistic effect on the upregulation of ZO-1 and occludin expression compared to a single metabolite in the Caco-2 cell model (P≤0.05). This result was corroborated with the results of immunofluorescence assay: as shown in Figure 5, the metabolite treatment group could effectively improve the structural integrity damage of LPS on tight junction proteins (ZO-1/occludin). The aforementioned results suggest that ISO-treated microbial metabolites can alleviate LPS-induced structural damage and inflammatory responses in Caco-2 cells by regulating P-gp and CB2 protein expression and may be associated with the P-gp/eCB signaling pathway.
 |
Figure 5 The combined of the four major metabolites exhibited a significant synergistic effect on the upregulation of ZO-1 and occludin expression Immunofluorescence detection of occludin and ZO-1 protein expression in fecal supernatant-treated Caco-2 cells (magnification, x40). |
Discussion
The intestinal barrier prevents microorganisms, antigens and toxic macromolecules in the intestinal lumen from invading the body, and is the body’s first line of defense against the external environment.30 The intestinal epithelial TJ is an important component of the intestinal mucosal barrier. Under physiological conditions, a large number of microorganisms in the intestinal tract interact with the IECs to maintain a dynamic balance, and once the TJ is reduced or missing, the permeability of the cellular space will increase, and bacteria and various macromolecular substances can enter the circulation through the TJ, causing diseases, including Necrotizing enterocolitis (NEC), IBD, celiac disease, diabetes mellitus, HIV-associated diarrhea, food allergies and others.31,32 The IEC monolayer barrier formed by Caco-2 cell culture mimics the intestinal barrier in vitro and is a well-established in vitro intestinal model for immune-related studies or analyses, such as intestinal inflammation, mucosal defense and immune tolerance to intrinsic flora.2
LPS, also known as endotoxin, is a major bacterial pathogen, and its direct contact with IECs disrupts intercellular TJ proteins and thus breaks the TJ structure leading to increased intestinal permeability. When the TJs are compromised or become permeable, LPS and other luminal antigens can breach the barrier through paracellular pathways, leading to the onset of intestinal inflammation. Persistent inflammatory stimuli can exacerbate the damage to the intestinal barrier, resulting in further dysfunction and an increased production of proinflammatory cytokines.12,33 Studies have shown that circulating levels of LPS are elevated in human subjects with IBD; similarly, high levels of LPS have been detected in the intestinal tissue and serum in animal models of IBD.34 Therefore, elevated levels of LPS can adversely affect IECs, leading to compromised integrity of the intestinal barrier. In the present study, a Caco-2 cell damage model was created through LPS exposure. The findings revealed that as the concentration of LPS increased, there was a marked decline in the viability of Caco-2 cells, accompanied by more pronounced apoptosis, and a progressive reduction in the protein levels of ZO-1 and occludin.
Earlier research has demonstrated that ISO notably reduces colonic inflammation induced by DSS and mitigates ulcerative colitis in mice. This effect is achieved through the modulation of gut microbiota and their metabolic products.22 However, the effect of ISO-regulated microbial metabolites on the inflammatory response of IECs under damaging somatic conditions is unknown. CCK-8 assay results showed that ISO-regulated microbial metabolites significantly increased the reduction in cell viability caused by LPS at a concentration of 10 mg/mL. ELISA and RT-qPCR were used to detect the effects of ISO-regulated microbial metabolites on LPS-stimulated inflammatory responses in Caco-2 cells showed that ISO-regulated microbial metabolites significantly reduced the expression of the pro-inflammatory factors IL-6 and TNF-α in LPS-induced Caco-2 cells. TJ proteins are essential for preserving the integrity of the IEB during oxidative stress or inflammatory states as they physically close the gaps between IECs.35 Notably, experimental data from colonic biopsies of patients with IBD highlight the damage to the intestinal barrier, accompanied by a decrease in TJ protein expression.36,37 These findings indicate that regulating TJ proteins enhances the integrity of the intestinal barrier, which could serve as a promising therapeutic approach for IBD. In the current study, both TEER measurements and cell monolayer permeability tests were conducted to evaluate the functionality of the intestinal barrier.15,16 The findings revealed that microbial metabolites regulated by ISO notably enhanced the levels of TJ proteins, specifically ZO-1 and occludin, in Caco-2 cells that were affected by LPS. Furthermore, there was a significant reduction in FITC-Dextran permeability across the monolayer cells, effectively counteracting the increase in monolayer cell permeability induced by LPS. It is suggested that ISO-regulated microbial metabolites inhibited the LPS-induced decrease in Caco-2 cell viability and attenuated the inflammatory response, possibly regulating barrier function by modulating the expression of TJ proteins.
P-gp, as a transporter protein, plays an important role in preventing the absorption of exogenous substances and harmful metabolites into the body, and can also inhibit the transepithelial migration of neutrophils by mediating the metabolic pathway associated with eCBs, thereby modulating the immune-inflammatory response.24 CB1 and 2 are the 2 most common receptors in eCBs: CB1 can oxidize macrophages in association with nitric oxide synthase in the body, thereby exacerbating the inflammatory response,38 and CB2 has an anti-inflammatory effect; when there is systemic inflammatory response, the activated CB2 can not only reduce inflammatory mediators, but also reduce the content of proinflammatory factors and can regulate the stability of the intestinal microbiota.39,40 In this experiment, the treatment of microbial metabolites regulated by ISO increased the cell viability reduction caused by LPS, decreased IEB permeability and proinflammatory factor secretion, and thus alleviated the IEB inflammatory response. In addition, it increased the expression of P-gp and CB2 proteins in Caco-2 cells, suggesting that it may play a therapeutic role by regulating CB2 through the “P-gp pump”. To verify this result, the effect of the P-gp inhibitor verapamil on LPS-induced inflammation and TJ structure was next investigated in Caco-2 cells. The results revealed that the P-gp inhibitor verapamil could not alleviate LPS-induced inflammation in Caco-2 cells, while Western blotting results showed that the expression levels of the TJ proteins ZO-1 and occludin, as well as a reduction in the expression of the CB2 proteins in Caco-2 cells in the verapamil group. When P-gp was inhibited, the LPS-induced inflammation was not successfully alleviated, and there was a concurrent reduction in CB2 expression. This observation implies that P-gp plays a critical role not only in preserving the integrity of the intestinal barrier but also in potentially modulating the expression and function of CB2 receptors through a specific underlying mechanism.
SBAs and SCFAs are two of the major bacterial metabolites in the colon, and there is growing evidence that SCFAs and SBAs have biological activities that inhibit inflammation and promote the intestinal barrier.18,28,41 In previous studies, the major gut microbial metabolites regulated by ISO were found to include butyrate, UDCA, LCA, and DCA; therefore, to investigate the role of these ISO-regulated metabolites on LPS-induced inflammation in Caco-2 cells, we designed further based on the results of the existing studies and in conjunction with the results of the iso-regulated cell viability experiments in mouse fecal supernatant Experiments.28 By treating LPS-stimulated Caco-2 cells with butyric acid, DCA, LCA, and UDCA alone or in combination, the experimental results showed that the combination of these four key metabolites was effective in attenuating the LPS-stimulated inflammatory response in Caco-2 cells and inhibiting the damage of cellular tight junction (TJ) proteins.28,42 Consistently with the results of the ISO-regulated microbial metabolites, the aforementioned results suggest that ISO-regulated microbial metabolites are protective against LPS-induced Caco-2 cell inflammatory injury. The unique regulation of the P-gp/eCBs pathway by ISO-conditioned intestinal flora supernatant was the novel mechanistic insights provided in this study. However, the detailed mechanisms underlying the regulation of the P-gp/eCBs pathway by ISO-conditioned intestinal flora supernatant remain unclear and more investigations are needed.
In conclusion, the intestinal microbial metabolites regulated by ISO had a significant protective effect on permeability in the IEB inflammation model, which may reduce Caco-2 monolayer cell permeability by regulating the expression of the P-gp/eCB axis-related proteins P-gp and CB2, decreasing the disruption of the TJ proteins ZO-1 and occludin, enhancing the intestinal barrier function, decreasing the secretion of IL-6 and TNF-α and attenuating Caco-2 cell monolayer inflammation, thereby exerting a therapeutic effect (Figure 6). Overall, these findings could be instrumental in formulating treatment approaches involving ISO for managing inflammation-associated conditions in patients with IBD.
 |
Figure 6 A model illustration of the potential molecular mechanism involved in the fecal supernatant-treated Caco-2 cells. |
Ethics Approval and Consent to Participate
The animal experiments were approved by the Animal Ethics Committee of Shanghai Ocean University under the review and approval number SHOU-DW-2022-047. US National Institutes of Health Guide for the Care and Use of Laboratory Animals was followed in this study.
Acknowledgments
The authors thank all the technicians at the Laboratory Animal Facility of Shanghai Ocean University for providing daily care for the mice.
Funding
This work received funding from the Shanghai Pudong New Area Health System Young Medical Talents Training Program (PWRq2023-26) and NSFC (No. 82200689).
Disclosure
The authors report no competing interests in this work.
References
1. Gibson PR, Anderson RP, Mariadason JM, Wilson AJ. Protective role of the epithelium of the small intestine and colon. Inflamm Bowel Dis. 1996;2(4):279–302. doi:10.1097/00054725-199612000-00011
2. Van De Walle J, Hendrickx A, Romier B, Larondelle Y, Schneider YJ. Inflammatory parameters in Caco-2 cells: effect of stimuli nature, concentration, combination and cell differentiation. Toxicol In Vitro. 2010;24(5):1441–1449. doi:10.1016/j.tiv.2010.04.002
3. Darfeuille-Michaud A, Boudeau J, Bulois P, et al. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology. 2004;127(2):412–421. doi:10.1053/j.gastro.2004.04.061
4. Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflammatory bowel disease. Nature. 2007;448(7152):427–434. doi:10.1038/nature06005
5. Yan H, Ajuwon KM. Butyrate modifies intestinal barrier function in IPEC-J2 cells through a selective upregulation of tight junction proteins and activation of the Akt signaling pathway. PLoS One. 2017;12(6):e0179586.
6. Rawat M, Nighot M, Al-Sadi R, et al. IL1B increases intestinal tight junction permeability by up-regulation of MIR200C-3p, which degrades occludin mRNA. Gastroenterology. 2020;159(4):1375–1389. doi:10.1053/j.gastro.2020.06.038
7. Smyth D, McKay CM, Gulbransen BD, Phan VC, Wang A, McKay DM. Interferon-gamma signals via an ERK1/2-ARF6 pathway to promote bacterial internalization by gut epithelia. Cell Microbiol. 2012;14(8):1257–1270. doi:10.1111/j.1462-5822.2012.01796.x
8. Garcia-Hernandez V, Quiros M, Nusrat A. Intestinal epithelial claudins: expression and regulation in homeostasis and inflammation. Ann N Y Acad Sci. 2017;1397(1):66–79. doi:10.1111/nyas.13360
9. Guan Q, Zhang J. Recent advances: the imbalance of cytokines in the pathogenesis of inflammatory bowel disease. Mediators Inflamm. 2017;2017:4810258. doi:10.1155/2017/4810258
10. He C, Deng J, Hu X, et al. Vitamin A inhibits the action of LPS on the intestinal epithelial barrier function and tight junction proteins. Food Funct. 2019;10(2):1235–1242. doi:10.1039/C8FO01123K
11. Omonijo FA, Liu S, Hui Q, et al. Thymol improves barrier function and attenuates inflammatory responses in porcine intestinal epithelial cells during Lipopolysaccharide (LPS)-induced inflammation. J Agric Food Chem. 2019;67(2):615–624. doi:10.1021/acs.jafc.8b05480
12. Guo S, Al-Sadi R, Said HM, Ma TY. Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14. Am J Pathol. 2013;182(2):375–387. doi:10.1016/j.ajpath.2012.10.014
13. Dirisina R, Katzman RB, Goretsky T, et al. p53 and PUMA independently regulate apoptosis of intestinal epithelial cells in patients and mice with colitis. Gastroenterology. 2011;141(3):1036–1045. doi:10.1053/j.gastro.2011.05.032
14. Governa P, Marchi M, Cocetta V, et al. Effects of Boswellia serrata Roxb. and Curcuma longa L. in an In vitro intestinal inflammation model using immune cells and Caco-2. Pharmaceuticals. 2018;11(4):126. doi:10.3390/ph11040126
15. Cocetta V, Catanzaro D, Borgonetti V, et al. A fixed combination of probiotics and herbal extracts attenuates intestinal barrier dysfunction from inflammatory stress in an in vitro model using Caco-2 cells. Recent Pat Food Nutr Agric. 2019;10(1):62–69. doi:10.2174/2212798410666180808121328
16. Wu XX, Huang XL, Chen RR, et al. Paeoniflorin prevents intestinal barrier disruption and inhibits Lipopolysaccharide (LPS)-induced inflammation in Caco-2 cell monolayers. Inflammation. 2019;42(6):2215–2225. doi:10.1007/s10753-019-01085-z
17. Clemente JC, Manasson J, Scher JU. The role of the gut microbiome in systemic inflammatory disease. BMJ. 2018;360:j5145. doi:10.1136/bmj.j5145
18. Peng L, He Z, Chen W, Holzman IR, Lin J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatr Res. 2007;61(1):37–41. doi:10.1203/01.pdr.0000250014.92242.f3
19. Johnson CD, Kudsk KA. Nutrition and intestinal mucosal immunity. Clin Nutr. 1999;18(6):337–344. doi:10.1016/S0261-5614(99)80012-0
20. Zhang C, Li Q, Xing J, et al. Tannic acid and zinc ion coordination of nanase for the treatment of inflammatory bowel disease by promoting mucosal repair and removing reactive oxygen and nitrogen species. Acta Biomater. 2024;177:347–360. doi:10.1016/j.actbio.2024.02.015
21. Lu H, Zhou Q, Li J, et al. CO-releasing polyoxometalates nanozyme with gut mucosal immunity and microbiota homeostasis remodeling effects for restoring intestinal barrier integrity. Adv. Sci. 2025;12(17):e2500116. doi:10.1002/advs.202500116
22. Wang Z, Yang L, Feng Y, et al. Isoorientin alleviates DSS-treated acute colitis in mice by regulating intestinal epithelial P-glycoprotein (P-gp) expression. DNA Cell Biol. 2024;43(10):520–536. doi:10.1089/dna.2024.0101
23. Guo C, Wang Y, Zhang S, et al. Crataegus pinnatifida polysaccharide alleviates colitis via modulation of gut microbiota and SCFAs metabolism. Int J Biol Macromol. 2021;181:357–368. doi:10.1016/j.ijbiomac.2021.03.137
24. Szabady RL, Louissaint C, Lubben A, et al. Intestinal P-glycoprotein exports endocannabinoids to prevent inflammation and maintain homeostasis. J Clin Invest. 2018;128(9):4044–4056. doi:10.1172/JCI96817
25. Neish AS. Acute inflammation: endogenous cannabinoids mellow the harsh proinflammatory environment. J Clin Invest. 2018;128(9):3750–3751. doi:10.1172/JCI122885
26. Foley SE, Tuohy C, Dunford M, et al. Gut microbiota regulation of P-glycoprotein in the intestinal epithelium in maintenance of homeostasis. Microbiome. 2021;9(1):183. doi:10.1186/s40168-021-01137-3
27. Haran JP, Bhattarai SK, Foley SE, et al. Alzheimer’s disease microbiome is associated with dysregulation of the anti-inflammatory P-glycoprotein pathway. mBio. 2019;10(3). doi:10.1128/mBio.00632-19
28. Foley SE, Dente MJ, Lei X, et al. Microbial metabolites orchestrate a distinct multi-tiered regulatory network in the intestinal epithelium that directs P-glycoprotein expression. mBio. 2022;13(4):e0199322. doi:10.1128/mbio.01993-22
29. Li F, Du P, Yang W, Huang D, Nie S, Xie M. Polysaccharide from the seeds of Plantago asiatica L. alleviates nonylphenol induced intestinal barrier injury by regulating tight junctions in human Caco-2 cell line. Int J Biol Macromol. 2020;164:2134–2140. doi:10.1016/j.ijbiomac.2020.07.259
30. Förster C. Tight junctions and the modulation of barrier function in disease. Histochem Cell Biol. 2008;130(1):55–70. doi:10.1007/s00418-008-0424-9
31. Matamoros S, Gras-Leguen C, Le Vacon F, Potel G, de La Cochetiere MF. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013;21(4):167–173. doi:10.1016/j.tim.2012.12.001
32. Mankertz J, Schulzke JD. Altered permeability in inflammatory bowel disease: pathophysiology and clinical implications. Curr Opin Gastroenterol. 2007;23(4):379–383. doi:10.1097/MOG.0b013e32816aa392
33. Han X, Fink MP, Yang R, Delude RL. Increased iNOS activity is essential for intestinal epithelial tight junction dysfunction in endotoxemic mice. Shock. 2004;21(3):261–270. doi:10.1097/01.shk.0000112346.38599.10
34. Guo S, Nighot M, Al-Sadi R, Alhmoud T, Nighot P, Ma TY. Lipopolysaccharide regulation of intestinal tight junction permeability is mediated by TLR4 signal transduction pathway activation of FAK and MyD88. J Immunol. 2015;195(10):4999–5010. doi:10.4049/jimmunol.1402598
35. Zhang B, Guo Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br J Nutr. 2009;102(5):687–693. doi:10.1017/S0007114509289033
36. Kuo WT, Zuo L, Odenwald MA, et al. The tight junction protein ZO-1 is dispensable for barrier function but critical for effective mucosal repair. Gastroenterology. 2021;161(6):1924–1939. doi:10.1053/j.gastro.2021.08.047
37. Ibrahim S, Zhu X, Luo X, Feng Y, Wang J. PIK3R3 regulates ZO-1 expression through the NF-kB pathway in inflammatory bowel disease. Int Immunopharmacol. 2020;85:106610. doi:10.1016/j.intimp.2020.106610
38. Khan N, Laudermilk L, Ware J, et al. Peripherally selective CB1 receptor antagonist improves symptoms of metabolic syndrome in mice. ACS Pharmacol Transl Sci. 2021;4(2):757–764. doi:10.1021/acsptsci.0c00213
39. Chayasirisobhon S. Cannabis and neuropsychiatric disorders: an updated review. Acta Neurol Taiwan. 2019;28(2):27–39.
40. Ellermann M, Pacheco AR, Jimenez AG, et al. Endocannabinoids inhibit the induction of virulence in enteric pathogens. Cell. 2020;183(3):650–665.e615. doi:10.1016/j.cell.2020.09.022
41. Zeng H, Umar S, Rust B, Lazarova D, Bordonaro M. Secondary bile acids and short chain fatty acids in the colon: a focus on colonic microbiome, cell proliferation, inflammation, and cancer. Int J Mol Sci. 2019;20(5):1214.
42. Peng L, Li ZR, Green RS, Holzman IR, Lin J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J Nutr. 2009;139(9):1619–1625. doi:10.3945/jn.109.104638
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.