Back to Journals » International Journal of Nanomedicine » Volume 17
Ionizing Radiation-Induced Ferroptosis Based on Nanomaterials
Authors Zhang S ![]() , Zhang J, Fan X, Liu H, Zhu M, Yang M, Zhang X, Zhang H, Yu F
, Zhang J, Fan X, Liu H, Zhu M, Yang M, Zhang X, Zhang H, Yu F
Received 2 May 2022
Accepted for publication 28 July 2022
Published 6 August 2022 Volume 2022:17 Pages 3497—3507
DOI https://doi.org/10.2147/IJN.S372947
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yan Shen
Shenghong Zhang,1– 3 Jiajia Zhang,1,3 Xin Fan,1,3 Hanhui Liu,1– 3 Mengqin Zhu,1– 3 Mengdie Yang,1,3 Xiaoyi Zhang,1,3 Han Zhang,1,3 Fei Yu1,3
1Department of Nuclear Medicine, Shanghai Tenth People’s Hospital, Tongji University School of Medicine, Shanghai, People’s Republic of China; 2Shanghai Clinical College, Anhui Medical University, Shanghai, People’s Republic of China; 3Institute of Nuclear Medicine, Tongji University School of Medicine, Shanghai, People’s Republic of China
Correspondence: Fei Yu, Department of Nuclear Medicine, Shanghai Tenth People’s Hospital, Tongji University School of Medicine, Shanghai, People’s Republic of China, Tel +86-021-66302721, Fax +86-021-66300588, Email [email protected]
Abstract: Ferroptosis is an iron-dependent form of regulated cell death (RCD), that is associated with peroxidative damage to cellular membranes. A promising therapeutic method is to target ferroptosis. Nanomaterial-induced ferroptosis attracts enormous attention. Nevertheless, there are still certain shortcomings in ferroptosis, such as inadequate triggered immunogenic cell death to suit clinical demands. Various investigations have indicated that ionizing radiation (IR) can further induce ferroptosis. Consequently, it is a potential strategy for cancer therapy that combines nanomaterials and IR to induce ferroptosis. Initially, we discuss various ferroptosis inducers based on nanomaterials in this review. Furthermore, mechanisms of IR-induced ferroptosis are briefly introduced. Ultimately, we assess the feasibility of combining nanomaterials with IR to induce ferroptosis, paving the way for future research.
Keywords: radiation therapy, lipid peroxidation, ROS, nanoparticles, combined therapy
Introduction
Ferroptosis, one of the rare non-apoptotic cell deaths, is an iron-dependent form of RCD induced by peroxidative damages of polyunsaturated-fatty-acid-containing phospholipids (PUFA–PLs) in cellular membranes.1 Studies have demonstrated that ferroptosis is distinct from other forms of RCD. Ferroptosis cells typically have constricted mitochondria, higher mitochondrial membrane density, and fewer mitochondrial cristae.1 Mechanistically, cells undergoing ferroptosis is regulated by multiple cellular metabolic pathways such as redox homeostasis,2 iron handling,3 and metabolism of amino acids,4 lipids5 and energy.6 Ferroptosis as a critical cell death response is involved in multiple cancer therapies.7 As a result, ferroptosis has sparked a wave of interest in recent years as a viable therapeutic approach. Ferroptosis, on the other hand, is dependent on reactive oxygen species (ROS) and lipid peroxidation. Targeting ferroptosis treatment has obstacles because of limited efficiency and selectivity.8 For example, ferroptosis-induced immunogenic cell death is insufficient to fulfill clinical requirements.
Various nanomaterials have garnered a great deal of attention as nanotechnology has advanced. The benefits of nanomaterials in terms of biocompatibility, cytotoxicity, targeted delivery characteristics, dispersion throughout the organism, and metabolism are increasingly being revealed.9 Consequently, Nanomaterials-induced ferroptosis has gained significant attention. By boosting ROS expression and activating long-chain-fatty-acid CoA ligase 4 (ACSL4) via IR, ferroptosis can be strengthened.10 In addition, recent studies have discovered that cancer cells can be hypersensitive to radiation by enhancing ferroptosis,11 which can increase the efficacy and indications of IR.12 In this review, firstly, we discuss various ferroptosis inducers based on nanomaterials. Secondly, mechanisms of IR-induced ferroptosis are briefly introduced. Finally, we evaluate the viability of using nanomaterials in conjunction with IR to induce ferroptosis, opening the door for further investigation.
Ferroptosis Inducer Based on Nanomaterials
Numerous therapeutically authorized drugs and ferroptosis inducers are being studied. These drugs primarily affect ferroptosis via influencing the ferroptosis executive system and defense system, redox and iron balance, and cellular metabolism. Ferroptosis inducers are mainly grouped into three categories: I FINs, which inhibit the activity of SLC7A11 or deplete GSH (Erastin, Sulfasalazine (SAS)), class II FINs that inhibit the activity of GPX4 (RSL3), and Class III FIN (Idebenone), which indirectly depletes CoQ and GPX4 by activating tricosahexaene synthase(SQS).1 There are several drawbacks of small-molecule ferroptosis inducers, but nanomaterials can improve the accumulation and release of drugs on the tumor microenvironment (TME), hence exerting due to their strong biocompatibility, considerable benefits owing to nano-size effects, and enhanced permeability. Recently, several reviews have discussed the mechanism and application of ferroptosis in nanomedicine.13,14 In comparison to small-molecule ferroptosis inducers, nanomaterial-based ferroptosis inducers not only promote water solubility, cell targeting ability, tumor-specific drug accumulation and prolong circulation times in vivo,15 but also induce ferroptosis by loading and releasing Fe2+/Fe3+ ions in cells, accelerating Fenton reactions and lipid peroxidation accumulation.16,17 Therefore, nanomaterial-based ferroptosis inducers have received a huge amount of attention. The ferroptosis execution system and the ferroptosis defense system are crucial to the rational design of nanomaterial-based ferroptosis inducers (Table 1).
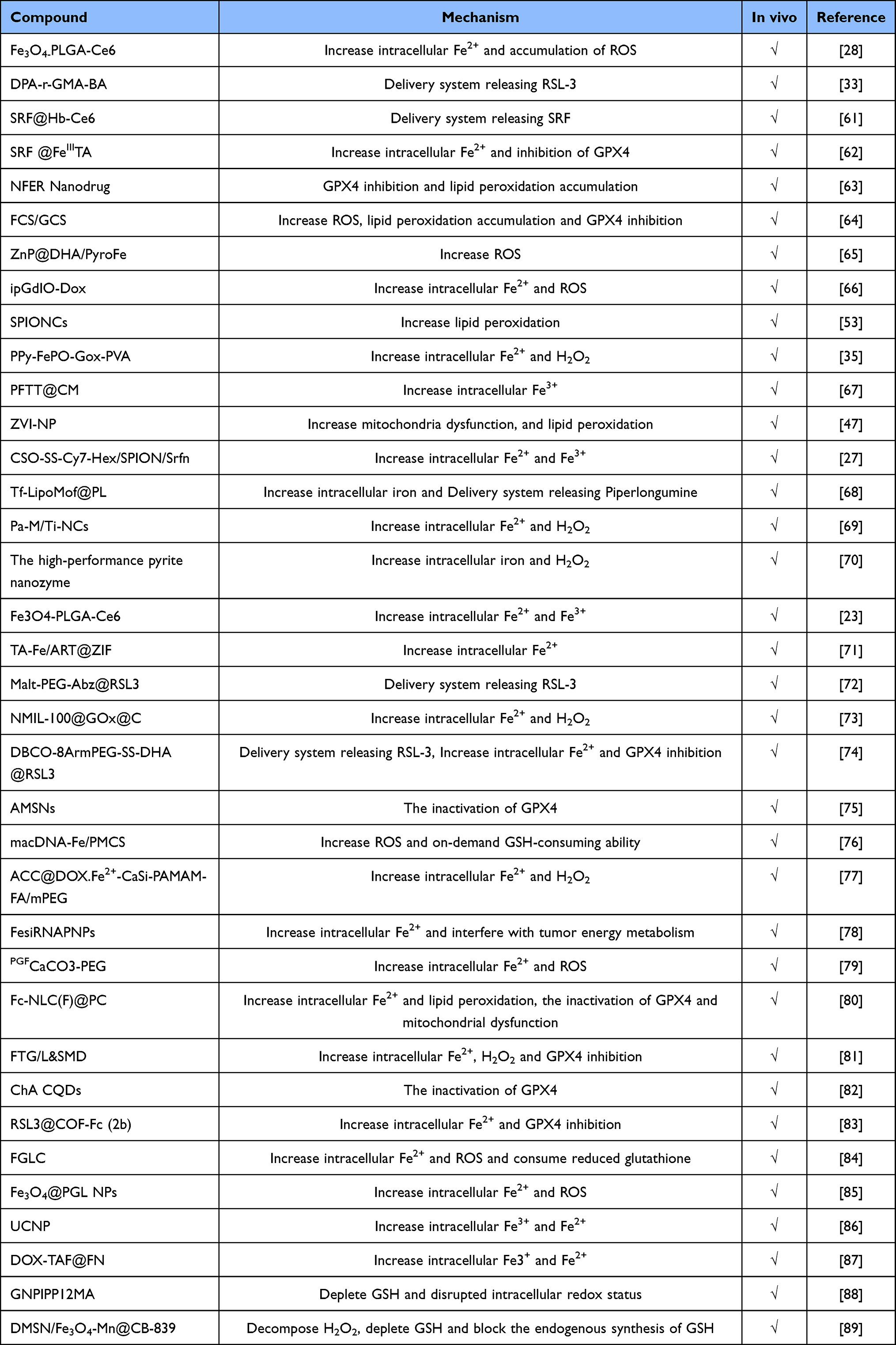 |
Table 1 The Identified Nanomaterials for Inducing Ferroptosis |
PUFA-PL Synthesis and Peroxidation
Lipid peroxidation is a common feature of ferroptosis. Nanomaterials cause ferroptosis predominantly by accelerating PUFA oxidation and esterification18 and rewriting lipid metabolism.19 Additionally, ferroptosis was studied by stimulating the synthesis of polyunsaturated ether phospholipids (PUFA-ePLs) as a substrate for lipid peroxidation.20
Iron Metabolism
Due to the formation of iron-mediated ROS via the Fenton reaction, iron metabolism plays a crucial role in ferroptosis.21 Iron-based nanoparticles (NPs) and non-iron-based nanoparticles are the two primary categories of nanomaterials used to induce ferroptosis (NNPs). NPs were developed to induce ferroptosis by regulating the intracellular iron, utilizing iron oxide nanoparticles,20 amorphous nanometallic glasses22 and metal organic frameworks (MOFs). In order to directly release ferrous/ferric ions and trigger ferroptosis, Chen et al created the Fe3O4-PLGA-Ce6 nanosystem. This system undergoes the Fenton reaction with intracellular excess hydrogen peroxide.23 Additionally, some NPs-induced ferroptosis is intimately connected to lysosomal dysfunction. They primarily employ redox processes, denaturation of lysosomal biomolecules, and physical interactions to treat lysosomal impairment, which eventually culminates in ferroptosis.13 Additionally, Some NPs-induced ferroptosis is intimately connected to lysosomal dysfunction. They primarily employ redox processes, denaturation of lysosomal biomolecules, and physical interactions to treat lysosomal impairment, which eventually culminates in ferroptosis.24 By indirectly boosting intracellular iron and controlling genes or proteins involved in iron metabolism, NNPs can alter cellular iron metabolism. By completely exploiting endogenous iron stored in endo-lysosomes and an artificial intracellular favorable feedback loop, Xiong et al established a nano-activator to induce ferroptosis.25
Mitochondrial Metabolism
Mitochondria are the primary site of intracellular iron utilization and a major source of cellular ROS.26 Nanomaterials can stimulate mitochondrial metabolism in a multitude of ways to induce ferroptosis. One of the key mechanisms of nanomaterials-induced ferroptosis is Mitochondria iron overload. Superparamagnetic iron oxide nanoparticles (SPION) were degraded to free ferrous iron in lysosomes and subsequently entered into mitochondria, leading to mitochondrial iron overload, and finally mitochondrial lipid peroxidation accumulation, causing ferroptosis.16 Mitochondrial membrane anchored oxidation/reduction response and Fenton-Reaction-Accelerable magnetic nanophotosensitizer complex self-assemblies loading sorafenib (CSO-SS-Cy7-Hex/SPION/Srfn) were constructed, which can anchor to the mitochondrial membrane to induces ferroptosis.27 Other investigations have discovered that ferroptosis can be induced by affecting mitochondrial metabolism by upregulating mitochondrial voltage-dependent anion channel (VDAC) proteins.28 Simultaneously, the research reveals that mitochondrial morphological deformations promote mitochondrial collapse, which can eventually lead to ferroptosis.29 Therefore, nanomaterials that directly target mitochondria can boost ferroptosis efficiency even more.
GPX4 Dependent Systems
The SLC7A11-GSH-GPX4 axis constitutes GPX4 dependent systems against ferroptosis. SLC7A11 is the critical transporter subunit in the glutamate-cystine reverse transporter (system Xc-), which is the primary route for intracellular cysteine to be obtained.30 Intracellular cysteine is largely responsible for GSH synthesis, and GSH peroxidases 4 (GPX4) plays an essential role in converting GSH into oxidized glutathione disulfide (GSSG) to inhibit ferroptosis.31 Inhibition of system Xc-32 and GPX433 and depletion of intracellular GSH34 are the chief mechanisms by which nanomaterials for inducing ferroptosis regulate GPX4 dependent systems to induce ferroptosis.
Other Mechanisms
FSP1, which is recruited to the plasma membrane as an oxidoreductase, lowers coenzyme Q10 (CoQ10), preventing the growth of lipid peroxides.35 To amplify ferroptosis, a ferroptosis-driven nanomaterial is intended to disrupt a GPX4- and GSH-independent ferroptosis regulation the FSP1-CoQ10-NADPH pathway.36
Mechanisms of Ferroptosis Induced by Radiotherapy
The crux of ferroptosis execution is regulating the balance between the ferroptosis execution system and the ferroptosis defense system. Cells conduct ferroptosis when the ferroptosis execution system exceeds the ferroptosis defense system. PUFA–PLs synthesis and peroxidation,37 iron metabolism,38 and mitochondrial metabolism7 constitute the ferroptosis execution system. The ferroptosis defense system mostly consists of GPX4 dependent and GPX4 non-dependent systems. The ferroptosis defense system is heavily reliant on the solute carrier family 7 member 11-glutathione (SLC7A11)-GSH-GPX4 axis.39 The ferroptosis inhibitory protein 1 (FSP1)-CoQ10 pathway,35 dihydroorotate dehydrogenase (DHODH)-dihydroubiquione (CoQH2) pathway,40 and GTP cyclohydrolase-1 (GCH1)-tetrahydrobiopterin (BH4) pathway41 constitute the ferroptosis defense system. Specifically, IR predominantly induces ferroptosis via the parallel mechanisms listed below (Figure 1).
- IR not only causes excess ROS formation by initiating radiolysis of cellular water and stimulating oxidative enzymes, altering mitochondrial structure or function, but also induces ACSL4-catalyzed PUFA production of PUFA-CoA, which is subsequently stimulated by LPCAT3 to generate PUFA-PLs. ROS and PUFA-PLs collaborate together to peroxide PUFA-PLs and induce ferroptosis.1
- The insertion of multi-PUFA into phospholipids of membrane components can be promoted via IR by upregulating the expression of ACSL4 to construct PUFA-PLs, which is subsequently mediated by ALOX to undergo lipid peroxidation and induce ferroptosis.42
- IR induces DNA damage, which activates ATM, inhibiting the production of SLC7A11, a crucial component of the cystine/glutamate transporter. It can continuously deplete GSH and inhibit GPX4, weakening the SLC7A11-GSH-GPX4 signaling axis-mediated ferroptosis defense system and further promoting ferroptosis.12
- IR-induced DNA breakage and subsequent activation of the DNA sensor cyclic GMP-AMP synthase (cGAS) signal activates the cGAS-STING1 pathway, leading to autophagy-dependent ferroptosis via lipid peroxidation.38
- IR can contribute to the production of tumor cell release particles (RT-MPs), which reverses the tumor microenvironment, increases antitumor effects, and mediates bystander effects (RIBE) in tumor cells essentially by triggering ferroptosis.43
 |
Figure 1 Mechanisms of ferroptosis induced by IR. It mainly affects ferroptosis through 5 pathways.1: IR not only causes excess ROS to induce ferroptosis by triggering radiolysis of cellular water and stimulating oxidative, altering mitochondrial structure or function. 2: IR can mediate lipid peroxidation and induce ferroptosis by upregulating the expression of ACSL4. 3: IR induced DNA damage, inhibited the expression of SLC7A11, weakened the ferroptosis defense system mediated by the SLC7A11-GSH-GPX4 signaling axis, and further promoted ferroptosis. 4: IR-induced DNA breakage activates the cGAS-STING1 pathway, leading to autophagy-dependent ferroptosis via lipid peroxidation. 5: IR, primarily by causing ferroptosis, can promote the production of tumor cell-released particles (RT-MPs), reverse the tumor microenvironment, increase antitumor effects, and mediate bystander effects (RIBE). |
Combined Ferroptosis-Driven Nanomaterials and Ionizing Radiation
Some studies have revealed that inducing ferroptosis is closely related to TME. Interferon-γ (IFN-γ) secreted by CD8+ cytotoxic T cells augment lipid peroxidation and ferroptosis through downregulating SLC7A11 expression.44 Nanomaterials may be used to transport therapeutic radioisotopes into malignancies, according to several studies.45 In recent investigations, nanoparticles operate as radio-sensitizers, depositing radiation energy and improving the therapeutic impact.46 A novel nanomaterial leads to ferroptosis by generating mitochondria dysfunction, intracellular oxidative stress, and lipid peroxidation, and a technique to synergistically reprogram TME is developed.47 Nonetheless, this therapy still has certain shortcomings in that induced immunogenic cell death is insufficient to fulfill clinical demands. Radiation therapy (RT) induces tumor-cell ferroptosis, whereas ferroptosis inducers boost radiation effectiveness.48 When IR is paired with ferroptosis inducers, antitumor immune responses are strengthened even more.48
External beam radiotherapy (EBRT) and internal radioisotope therapy (RIT) are composed of RT.46 RT utilizes IR to induce cell death directly through DNA double-strand breaks.49 IR induces cellular effects in both direct and indirect ways (Figure 2A). On the one hand, IR induces various type of DNA damages namely base damage, single-strand breaks (SSBs) and double-strand breaks (DSBs).50 DNA damage response (DDR) induced by ATM and ATR detection activates downstream checkpoint kinase 1/2 (CHEK1/2) to cause cell cycle arrest and encourage repair of the damaged DNA. If the damage cannot be fully repaired, the cell will initiate the apoptotic program.1,42 IR generate ROS by eliciting radiolysis of cellular water, activating oxidative enzymes and altering mitochondrial structure or function.1 Then ROS reaches a specific dose and can attack nucleic acids, lipids and proteins to induce ferroptosis. Specialized cells react differently to IR. TME, which is made up of several cells comprising tumor cells, immune cells, and cancer-associated fibroblasts (CAFs) determines how tumor cells respond to IR (Figure 2B).51 Monocytes and granulocytes, for instance, are extremely susceptible to radiation, whereas B cells, T cells, NK cells, macrophages, and dendritic cells (DCs), which have effective DNA repair and DDR capabilities, respond differently to radiation.50 Ferroptosis is one of the multiple deaths brought on by IR, which also releases TAAs and DAMPs, enhances antigen presentation, prompts DC activation, and subsequently enhances T cell activation. IR induces immunogenic cell death (ICD) including ferroptosis, then releases damage-associated molecular patterns (DAMPs) to facilitate the proliferation of CD8+ T cells through antigen cross-presentation from DCs.49
 |
Figure 2 (A) IR possesses direct and indirect impacts on cellular function. Base damage, SSBs, and DSBs are only a couple of minor types of DNA damage that IR may induce. In addition, IR causes radiolysis of cellular water, activates oxidative enzymes, and modifies mitochondrial structure or function to yield ROS. (B) TME is composed of different cell populations, such as tumor cells, immune cells, and CAFs, each of which reacts differently to radiation. Granulocytes and monocytes are particularly vulnerable to radiation, while B cells, T cells, NK cells, macrophages, and dendritic cells react differently. |
Jiang et al developed photosynthetic microcapsules (PMCs) powered by external near infrared photons. X-rays have the potential to efficiently generate oxidative radicals and construct micro oxygen factory in the body. Nanomaterial PMCs triggered by NIR-II laser irradiation may create a hyperoxia microenvironment in malignancy via controlled photosynthesis. The combination of X-rays and hyperoxia was discovered to cause Radiation-hyperoxia-induced lipid peroxidation (RHILP), which interferes with Fe2+ metabolism in cells, inhibits GPX4 production, and consumes GSH, ultimately leading to ferroptosis. Combining IR with nanomaterials can create an oxygen-rich microenvironment in the tumor and boost the therapeutic use of radiation by triggering ferroptosis, according to this study.52 Li et al reported superparamagnetic iron oxide nanoclusters (SPIONCs) that can release more iron ions on TME, and stimulate more hydrogen peroxide when exposed to X-rays. Ultimately, ferroptosis ensued because IR and NPs exacerbated generation of hydroxyl radicals and tumor lipid peroxidation.53 Additionally, this analysis demonstrated that SPIONCs have no effect on the level of blood biochemical and normal kidney and liver function and has the potential for clinical translation.53 In addition, certain studies have employed nanomaterials as radionuclide carriers to supply sufficient tumor-targeted delivery and improve effect of radionuclide therapy. Chen et al utilized the therapeutic radionuclide iodine-131(131I) to label HSA-CAT nanoreactors (NRs), and dramatically attenuate tumor hypoxia, hence boosting the therapeutic efficacy of radionuclide 131I.54 Shi et al designed a nano-radiopharmaceutical (177Lu-SPN-GIP) that displayed greater tumor-killing impact than conventional RT and considerably improved therapeutic outcomes.55 Consequently, combining nanomaterials-induced ferroptosis and ionizing radiation will be a viable approach. Because it can improve stability, cell uptake rate and tumor site retention time. Rational design of ionizing radiation-induced ferroptosis based on nanomaterials can not only improve the outcomes of conventional RT, but also enhance ferroptosis therapy, providing better antitumor effects than systemic approaches.
Conclusions and Perspectives
Ferroptosis inducer based on nanomaterials may not only directly induce cell ferroptosis, but also operate as radiosensitizers to attenuate the radio-resistance of cancer cells, and at the same time serve as targeted transport vehicles. It has the capacity to effectively limit tumor growth by enhancing immune responses further and observing a long-term immunological memory impact.56 Nanomaterial-based ferroptosis inducers have also been suggested as contrast agents for T1-weighted magnetic resonance imaging.57
Moreover, there are several issues with nanomaterials that have not yet been resolved. Nanomaterial-based ferroptosis inducers must also have minimal adverse effects, strong anticancer properties, biocompatibility, superior pharmacokinetics, and features for targeted distribution.58 We must first take into account the biosafety of nanomaterials. The linked organs may be harmed or toxicated by iron produced from nanomaterials.59 Second, it is apparent that multiple cancer forms differ in their susceptibility to radiation. Exist any unique variations in nanomaterials for clinical usages? Finally, can nanomaterial stability and biodegradability satisfy pharmacokinetics within the therapeutic window?59 Phagocytes may readily target nanomaterials, which hinders their ability to execute their intended objectives. Although the therapeutic efficacy of radionuclides for treatment is raised by incorporating them into nanomaterials, a significant barrier is their delayed elimination.60 The elevated levels of ROS in tumor cells have a variety of functional implications that, like a “double-edged sword”, cause ferroptosis. The susceptibility to ferroptosis and IR varies among people, cell types, and developmental stages, which is noteworthy.14 Therefore, more research is needed to determine if ionizing radiation and nanomaterial-induced ferroptosis are safe when used together in preclinical trials. IR combined with nanomaterial-induced ferroptosis can enhance immunogenicity and further support antitumor immune responses. Immune checkpoint inhibition facilitates the activation of systemic antitumor immune responses. An innovative concept for the therapy of cancer in the future is provided by coupled nanomaterial-induced ferroptosis and IR.
Abbreviations
RCD, regulated cell death; IR, ionizing radiation; PUFA–PLs, polyunsaturated-fatty-acid-containing phospholipids; ROS, reactive oxygen species; ACSL4, long-chain-fatty-acid CoA ligase 4; TME, the tumor microenvironment; PUFA-ePLs, polyunsaturated ether phospholipids; NPs, iron-based nanoparticles; NNPs, non-iron-based nanoparticles; MOFs, metal organic frameworks; GPX4, GSH peroxidases 4; CoQ10, coenzyme Q10; SLC7A11, The solute carrier family 7 member 11-glutathione; FSP1, The ferroptosis inhibitory protein 1; DHODH, dihydroorotate dehydrogenase (DHODH); CoQH2, dihydroubiquione; GCH1, GTP cyclohydrolase-1; BH4, tetrahydrobiopterin; cGAS, the DNA sensor cyclic GMP-AMP synthase; IFN-γ, Interferon-γ; RT, Radiation therapy; EBRT, External beam radiotherapy; RIT, internal radioisotope therapy; SSBs, single-strand breaks; DSBs, double-strand breaks; DDR, DNA damage response; CHEK1/2, checkpoint kinase 1/2; CAFs, cancer-associated fibroblasts; DCs, dendritic cells; ICD, immunogenic cell death; DAMPs, damage-associated molecular patterns; SPION, Superparamagnetic iron oxide nanoparticles; VDAC, voltage-dependent anion channel; PMCs, photosynthetic microcapsules; RHILP, radiation-hyperoxia-induced lipid peroxidation; SPIONCs, superparamagnetic iron oxide nanoclusters; NRs, nanoreactors.
Acknowledgments
This study was supported by grants from National Natural Science Foundation of China and Department of Nuclear Medicine, Shanghai Tenth People’s Hospital, Tongji University School of Medicine.
Funding
We acknowledge funding support by National Natural Science Foundation of China (No.82071956) and Clinical Research Plan of SHDC (No.2020CR4065).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lei G, Mao C, Yan Y, Zhuang L, Gan B. Ferroptosis, radiotherapy, and combination therapeutic strategies. Protein Cell. 2021;12:836–857. doi:10.1007/s13238-021-00841-y
2. Sharma A, Flora SJS. Positive and negative regulation of ferroptosis and its role in maintaining metabolic and redox homeostasis. Oxid Med Cell Longev. 2021;2021:1–13. doi:10.1155/2021/9074206
3. Chen X, Yu C, Kang R, Tang D. Iron metabolism in ferroptosis. Front Cell Dev Biol. 2020;8:590226. doi:10.3389/fcell.2020.590226
4. Yu X, Long YC, Parlanti P. Crosstalk between cystine and glutathione is critical for the regulation of amino acid signaling pathways and ferroptosis. Sci Rep. 2016;6(1):1–11. doi:10.1038/srep30033
5. Li D, Li Y. The interaction between ferroptosis and lipid metabolism in cancer. Sig Transduct Target Ther. 2020;5(1):1–10. doi:10.1038/s41392-020-00216-5
6. Ma Y, Han F, Min J, Lin W. Energy metabolism as a regulator of ferroptosis. Cell Cycle. 2020;19(22):2960–2962. doi:10.1080/15384101.2020.1838781
7. Chen Z, Jiang J, Fu N, Chen L. Targetting ferroptosis for blood cell-related diseases. J Drug Target. 2021;1–15. doi:10.1080/1061186X.2021.1971237
8. Yue R, Zhang C, Xu L, et al. Dual key co-activated nanoplatform for switchable MRI monitoring accurate ferroptosis-based synergistic therapy. Chem. 2022:S2451929422001462. doi:10.1016/j.chempr.2022.03.009
9. Shan X, Li S, Sun B, et al. Ferroptosis-driven nanotherapeutics for cancer treatment. J Control Release. 2020;319:322–332. doi:10.1016/j.jconrel.2020.01.008
10. Lei G, Zhang Y, Koppula P, et al. The role of ferroptosis in ionizing radiation-induced cell death and tumor suppression. Cell Res. 2020;30(2):146–162. doi:10.1038/s41422-019-0263-3
11. Ye LF, Chaudhary KR, Zandkarimi F, et al. Radiation-induced lipid peroxidation triggers ferroptosis and synergizes with ferroptosis inducers. ACS Chem Biol. 2020;15(2):469–484. doi:10.1021/acschembio.9b00939
12. Wang H, Jiang H, Van De Gucht M, De Ridder M. Hypoxic radioresistance: can ROS be the key to overcome it? Cancers. 2019;11(1):112. doi:10.3390/cancers11010112
13. Zheng H, Jiang J, Xu S, et al. Nanoparticle-induced ferroptosis: detection methods, mechanisms and applications. Nanoscale. 2021;13(4):2266–2285. doi:10.1039/D0NR08478F
14. Wang Y, Sun T, Jiang C. Nanodrug delivery systems for ferroptosis-based cancer therapy. J Control Release. 2022;344:289–301. doi:10.1016/j.jconrel.2022.01.034
15. Amreddy N, Babu A, Muralidharan R, et al. Recent advances in nanoparticle-based cancer drug and gene delivery. Adv Cancer Res. 2018;137:115–170. doi:10.1016/bs.acr.2017.11.003
16. Zheng H, You J, Yao X, Lu Q, Guo W, Shen Y. Superparamagnetic iron oxide nanoparticles promote ferroptosis of ischemic cardiomyocytes. J Cell Mol Med. 2020;24(18):11030–11033. doi:10.1111/jcmm.15722
17. Chen L, Lin Z, Liu L, et al. Fe 2+/Fe 3+ ions chelated with ultrasmall polydopamine nanoparticles induce ferroptosis for cancer therapy. ACS Biomater Sci Eng. 2019;5:9. doi:10.1021/acsbiomaterials.9b00461
18. He YJ, Liu XY, Xing L, Wan X, Chang X, Jiang HL. Fenton reaction-independent ferroptosis therapy via glutathione and iron redox couple sequentially triggered lipid peroxide generator. Biomaterials. 2020;241:119911. doi:10.1016/j.biomaterials.2020.119911
19. Xie S, Sun W, Zhang C, et al. Metabolic control by heat stress determining cell fate to ferroptosis for effective cancer therapy. ACS Nano. 2021;15(4):7179–7194. doi:10.1021/acsnano.1c00380
20. Zou Y, Henry WS, Ricq EL, et al. Plasticity of ether lipids promotes ferroptosis susceptibility and evasion. Nature. 2020;585(7826):603–608. doi:10.1038/s41586-020-2732-8
21. Wang S, Liao H, Li F, Ling D. A mini-review and perspective on ferroptosis-inducing strategies in cancer therapy. Chin Chem Lett. 2019;30(4):847–852. doi:10.1016/j.cclet.2019.03.025
22. Zhang C, Bu W, Ni D, et al. Synthesis of iron nanometallic glasses and their application in cancer therapy by a localized Fenton reaction. Angew Chem Int Ed. 2016;55(6):2101–2106. doi:10.1002/anie.201510031
23. Chen Q, Ma X, Xie L, et al. Iron-based nanoparticles for MR imaging-guided ferroptosis in combination with photodynamic therapy to enhance cancer treatment. Nanoscale. 2021;13(9):4855–4870. doi:10.1039/D0NR08757B
24. Xu S, Zheng H, Ma R, et al. Vacancies on 2D transition metal dichalcogenides elicit ferroptotic cell death. Nat Commun. 2020;11(1):3484. doi:10.1038/s41467-020-17300-7
25. Xiong H, Wang C, Wang Z, Lu H, Yao J. Self-assembled nano-activator constructed ferroptosis-immunotherapy through hijacking endogenous iron to intracellular positive feedback loop. J Control Release. 2021;332:539–552. doi:10.1016/j.jconrel.2021.03.007
26. Battaglia AM, Chirillo R, Aversa I, Sacco A, Costanzo F, Biamonte F. Ferroptosis and cancer: mitochondria meet the “iron maiden” cell death. Cells. 2020;9(6):E1505. doi:10.3390/cells9061505
27. Sang M, Luo R, Bai Y, et al. Mitochondrial membrane anchored photosensitive nano-device for lipid hydroperoxides burst and inducing ferroptosis to surmount therapy-resistant cancer. Theranostics. 2019;9(21):6209–6223. doi:10.7150/thno.36283
28. Zhang C, Liu Z, Zhang Y, Ma L, Song E, Song Y. “Iron free” zinc oxide nanoparticles with ion-leaking properties disrupt intracellular ROS and iron homeostasis to induce ferroptosis. Cell Death Dis. 2020;11(3):183. doi:10.1038/s41419-020-2384-5
29. Gao F, Wang F, Nie X, et al. Mitochondria-targeted delivery and light controlled release of iron prodrug and CO to enhance cancer therapy by ferroptosis. New J Chem. 2020;44(8):3478–3486. doi:10.1039/C9NJ05860E
30. Liu X, Olszewski K, Zhang Y, et al. Cystine transporter regulation of pentose phosphate pathway dependency and disulfide stress exposes a targetable metabolic vulnerability in cancer. Nat Cell Biol. 2020;22(4):476–486. doi:10.1038/s41556-020-0496-x
31. Zhang Y, Swanda RV, Nie L, et al. mTORC1 couples cyst(e)ine availability with GPX4 protein synthesis and ferroptosis regulation. Nat Commun. 2021;12(1):1589. doi:10.1038/s41467-021-21841-w
32. Xu Y, Han X, Li Y, et al. Sulforaphane mediates glutathione depletion via polymeric nanoparticles to restore cisplatin chemosensitivity. ACS Nano. 2019;13(11):13445–13455. doi:10.1021/acsnano.9b07032
33. Song R, Li T, Ye J, et al. Acidity‐activatable dynamic nanoparticles boosting ferroptotic cell death for immunotherapy of cancer. Adv Mater. 2021;33(31):2101155. doi:10.1002/adma.202101155
34. Meng X, Deng J, Liu F, et al. Triggered all-active metal organic framework: ferroptosis machinery contributes to the apoptotic photodynamic antitumor therapy. Nano Lett. 2019;19(11):7866–7876. doi:10.1021/acs.nanolett.9b02904
35. Bersuker K, Hendricks JM, Li Z, et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature. 2019;575(7784):688–692. doi:10.1038/s41586-019-1705-2
36. Li K, Lin C, Li M, et al. Multienzyme-like reactivity cooperatively impairs glutathione peroxidase 4 and ferroptosis suppressor protein 1 pathways in triple-negative breast cancer for sensitized ferroptosis therapy. ACS Nano. 2022;16(2):2381–2398. doi:10.1021/acsnano.1c08664
37. Chen X, Kang R, Kroemer G, Tang D. Broadening horizons: the role of ferroptosis in cancer. Nat Rev Clin Oncol. 2021;18(5):280–296. doi:10.1038/s41571-020-00462-0
38. Mumbauer S, Pascual J, Kolotuev I, Hamaratoglu F. Ferritin heavy chain protects the developing wing from reactive oxygen species and ferroptosis. PLoS Genet. 2019;15(9):e1008396. doi:10.1371/journal.pgen.1008396
39. Liu M, Kong XY, Yao Y, et al. The critical role and molecular mechanisms of ferroptosis in antioxidant systems: a narrative review. Ann Transl Med. 2022;10(6):368. doi:10.21037/atm-21-6942
40. Mao C, Liu X, Zhang Y, et al. DHODH-mediated ferroptosis defence is a targetable vulnerability in cancer. Nature. 2021;596(7873):E13–E13. doi:10.1038/s41586-021-03820-9
41. Kraft VAN, Bezjian CT, Pfeiffer S, et al. GTP cyclohydrolase 1/tetrahydrobiopterin counteract ferroptosis through lipid remodeling. ACS Cent Sci. 2020;6(1):41–53. doi:10.1021/acscentsci.9b01063
42. Huang RX, Zhou PK. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Sig Transduct Target Ther. 2020;5(1):60. doi:10.1038/s41392-020-0150-x
43. Wan C, Sun Y, Tian Y, et al. Irradiated tumor cell–derived microparticles mediate tumor eradication via cell killing and immune reprogramming. Sci Adv. 2020;6(13):eaay9789. doi:10.1126/sciadv.aay9789
44. Wang W, Green M, Choi JE, et al. CD8+ T cells regulate tumour ferroptosis during cancer immunotherapy. Nature. 2019;569(7755):270–274. doi:10.1038/s41586-019-1170-y
45. Shi X, Li Q, Zhang L, et al. 211 at-labeled polymer nanoparticles for targeted radionuclide therapy of Glucose-Dependent Insulinotropic Polypeptide Receptor (GIPR)-overexpressed cancer. Bioconjugate Chem. 2021;32(8):1763–1772. doi:10.1021/acs.bioconjchem.1c00263
46. Song G, Cheng L, Chao Y, Yang K, Liu Z. Emerging nanotechnology and advanced materials for cancer radiation therapy. Adv Mater. 2017;29(32):1700996. doi:10.1002/adma.201700996
47. Hsieh CH, Hsieh HC, Shih FS, et al. An innovative NRF2 nano-modulator induces lung cancer ferroptosis and elicits an immunostimulatory tumor microenvironment. Theranostics. 2021;11(14):7072–7091. doi:10.7150/thno.57803
48. Lang X, Green MD, Wang W, et al. Radiotherapy and immunotherapy promote tumoral lipid oxidation and ferroptosis via synergistic repression of SLC7A11. Cancer Discov. 2019;9(12):1673–1685. doi:10.1158/2159-8290.CD-19-0338
49. Zhu M, Yang M, Zhang J, et al. Immunogenic cell death induction by ionizing radiation. Front Immunol. 2021;12:705361. doi:10.3389/fimmu.2021.705361
50. Ragunathan K, Upfold NLE, Oksenych V. Interaction between fibroblasts and immune cells following DNA damage induced by ionizing radiation. IJMS. 2020;21(22):8635. doi:10.3390/ijms21228635
51. Rodriguez-Ruiz ME, Vitale I, Harrington KJ, Melero I, Galluzzi L. Immunological impact of cell death signaling driven by radiation on the tumor microenvironment. Nat Immunol. 2020;21(2):120–134. doi:10.1038/s41590-019-0561-4
52. Jiang J, Wang W, Zheng H, et al. Nano-enabled photosynthesis in tumours to activate lipid peroxidation for overcoming cancer resistances. Biomaterials. 2022;285:121561. doi:10.1016/j.biomaterials.2022.121561
53. Li Y, Yang J, Gu G, et al. Pulmonary delivery of theranostic nanoclusters for lung cancer ferroptosis with enhanced chemodynamic/radiation synergistic therapy. Nano Lett. 2022;22(3):963–972. doi:10.1021/acs.nanolett.1c03786
54. Chen J, Liang C, Song X, et al. Hybrid protein nano‐reactors enable simultaneous increments of tumor oxygenation and iodine‐131 delivery for enhanced radionuclide therapy. Small. 2019;15(46):1903628. doi:10.1002/smll.201903628
55. Shi X, Li Q, Zhang C, et al. Semiconducting polymer nano-radiopharmaceutical for combined radio-photothermal therapy of pancreatic tumor. J Nanobiotechnol. 2021;19(1):337. doi:10.1186/s12951-021-01083-0
56. Yang Y, Liu B, Liu Y, et al. DNA-based MXFs to enhance radiotherapy and stimulate robust antitumor immune responses. Nano Lett. 2022;22(7):2826–2834. doi:10.1021/acs.nanolett.1c04888
57. He H, Du L, Guo H, et al. Redox responsive metal organic framework nanoparticles induces ferroptosis for cancer therapy. Small. 2020;16(33):2001251. doi:10.1002/smll.202001251
58. Chen S, Xing C, Huang D, et al. Eradication of tumor growth by delivering novel photothermal selenium-coated tellurium nanoheterojunctions. Sci Adv. 2020;6(15):eaay6825. doi:10.1126/sciadv.aay6825
59. Yang G, Phua SZF, Bindra AK, Zhao Y. Degradability and clearance of inorganic nanoparticles for biomedical applications. Adv Mater. 2019;31:10. doi:10.1002/adma.201805730
60. Pellico J, Gawne PJ, de Rosales R. Radiolabelling of nanomaterials for medical imaging and therapy. Chem Soc Rev. 2021;50(5):3355–3423. doi:10.1039/D0CS00384K
61. Xu T, Ma Y, Yuan Q, et al. Enhanced ferroptosis by oxygen-boosted phototherapy based on a 2-in-1 nanoplatform of ferrous hemoglobin for tumor synergistic therapy. ACS Nano. 2020;14(3):3414–3425. doi:10.1021/acsnano.9b09426
62. Liu T, Liu W, Zhang M, et al. Ferrous-supply-regeneration nanoengineering for cancer-cell-specific ferroptosis in combination with imaging-guided photodynamic therapy. ACS Nano. 2018;12(12):12181–12192. doi:10.1021/acsnano.8b05860
63. Li Y, Wang X, Yan J, et al. Nanoparticle ferritin-bound erastin and rapamycin: a nanodrug combining autophagy and ferroptosis for anticancer therapy. Biomater Sci. 2019;7(9):3779–3787. doi:10.1039/c9bm00653b
64. Luo S, Ma D, Wei R, et al. A tumor microenvironment responsive nanoplatform with oxidative stress amplification for effective MRI-based visual tumor ferroptosis. Acta Biomaterialia. 2022;138:518–527. doi:10.1016/j.actbio.2021.11.007
65. Han W, Duan X, Ni K, Li Y, Chan C, Lin W. Co-delivery of dihydroartemisinin and pyropheophorbide-iron elicits ferroptosis to potentiate cancer immunotherapy. Biomaterials. 2022;280:121315. doi:10.1016/j.biomaterials.2021.121315
66. Zhu L, Wang J, Tang X, et al. Efficient magnetic nanocatalyst-induced chemo- and ferroptosis synergistic cancer therapy in combination with T 1 –T 2 dual-mode magnetic resonance imaging through doxorubicin delivery. ACS Appl Mater Interfaces. 2022;14(3):3621–3632. doi:10.1021/acsami.1c17507
67. Pan WL, Tan Y, Meng W, et al. Microenvironment-driven sequential ferroptosis, photodynamic therapy, and chemotherapy for targeted breast cancer therapy by a cancer-cell-membrane-coated nanoscale metal-organic framework. Biomaterials. 2022;283:121449. doi:10.1016/j.biomaterials.2022.121449
68. Xu R, Yang J, Qian Y, et al. Ferroptosis/pyroptosis dual-inductive combinational anti-cancer therapy achieved by transferrin decorated nanoMOF. Nanoscale Horiz. 2021;6(4):348–356. doi:10.1039/d0nh00674b
69. Zhang F, Li F, Lu GH, et al. Engineering magnetosomes for ferroptosis/immunomodulation synergism in cancer. ACS Nano. 2019;13(5):5662–5673. doi:10.1021/acsnano.9b00892
70. Meng X, Li D, Chen L, et al. High-performance self-cascade pyrite nanozymes for apoptosis-ferroptosis synergistic tumor therapy. ACS Nano. 2021;15(3):5735–5751. doi:10.1021/acsnano.1c01248
71. Li Z, Wu X, Wang W, et al. Fe(II) and tannic acid-cloaked MOF as carrier of artemisinin for supply of ferrous ions to enhance treatment of triple-negative breast cancer. Nanoscale Res Lett. 2021;16(1):37. doi:10.1186/s11671-021-03497-z
72. Li W, Liu X, Cheng X, et al. Effect of Malt-PEG-Abz@RSL3 micelles on HepG2 cells based on NADPH depletion and GPX4 inhibition in ferroptosis. J Drug Target. 2022;30(2):208–218. doi:10.1080/1061186X.2021.1953511
73. Wan X, Song L, Pan W, Zhong H, Li N, Tang B. Tumor-targeted cascade nanoreactor based on metal-organic frameworks for synergistic ferroptosis-starvation anticancer therapy. ACS Nano. 2020;14(9):11017–11028. doi:10.1021/acsnano.9b07789
74. Li Y, Li M, Liu L, et al. Cell-specific metabolic reprogramming of tumors for bioactivatable ferroptosis therapy. ACS Nano. 2022;16(3):3965–3984. doi:10.1021/acsnano.1c09480
75. Wang S, Li F, Qiao R, et al. Arginine-rich manganese silicate nanobubbles as a ferroptosis-inducing agent for tumor-targeted theranostics. ACS Nano. 2018;12(12):12380–12392. doi:10.1021/acsnano.8b06399
76. Cao F, Sang Y, Liu C, et al. Self-adaptive single-atom catalyst boosting selective ferroptosis in tumor cells. ACS Nano. 2022;16:855–868. doi:10.1021/acsnano.1c08464
77. Xue CC, Li MH, Zhao Y, et al. Tumor microenvironment-activatable Fe-doxorubicin preloaded amorphous CaCO3 nanoformulation triggers ferroptosis in target tumor cells. Sci Adv. 2020;6(18):eaax1346. doi:10.1126/sciadv.aax1346
78. Wang Y, Chen J, Lu J, et al. Metal ions/nucleotide coordinated nanoparticles comprehensively suppress tumor by synergizing ferroptosis with energy metabolism interference. J Nanobiotechnology. 2022;20:199. doi:10.1186/s12951-022-01405-w
79. Han Y, Dong Z, Wang C, et al. Ferrous ions doped calcium carbonate nanoparticles potentiate chemotherapy by inducing ferroptosis. J Control Release. 2022;348:346–356. doi:10.1016/j.jconrel.2022.06.002
80. Wang W, Fu F, Huang Z, et al. Inhalable biomimetic protein corona-mediated nanoreactor for self-amplified lung adenocarcinoma ferroptosis therapy. ACS Nano. 2022;16(5):8370–8387. doi:10.1021/acsnano.2c02634
81. Zhu G, Chi H, Liu M, et al. Multifunctional “ball-rod” Janus nanoparticles boosting Fenton reaction for ferroptosis therapy of non-small cell lung cancer. J Colloid Interface Sci. 2022;621:12–23. doi:10.1016/j.jcis.2022.04.021
82. Yao L, Zhao MM, Luo QW, et al. Carbon quantum dots-based nanozyme from coffee induces cancer cell ferroptosis to activate antitumor immunity. ACS Nano. 2022;16:9228–9239. doi:10.1021/acsnano.2c01619
83. Zhou LL, Guan Q, Li WY, Zhang Z, Li YA, Dong YB. A ferrocene-functionalized covalent organic framework for enhancing chemodynamic therapy via redox dyshomeostasis. Small. 2021;17(32):e2101368. doi:10.1002/smll.202101368
84. Xu XL, Zhang NN, Shu GF, et al. A luminol-based self-illuminating nanocage as a reactive oxygen species amplifier to enhance deep tumor penetration and synergistic therapy. ACS Nano. 2021;15(12):19394–19408. doi:10.1021/acsnano.1c05891
85. Liang X, Chen M, Bhattarai P, Hameed S, Tang Y, Dai Z. Complementing cancer photodynamic therapy with ferroptosis through iron oxide loaded porphyrin-grafted lipid nanoparticles. ACS Nano. 2021;15(12):20164–20180. doi:10.1021/acsnano.1c08108
86. Bao W, Liu X, Lv Y, et al. Nanolongan with multiple on-demand conversions for ferroptosis-apoptosis combined anticancer therapy. ACS Nano. 2019;13(1):260–273. doi:10.1021/acsnano.8b05602
87. Xu Y, Guo Y, Zhang C, et al. Fibronectin-coated metal-phenolic networks for cooperative tumor chemo-/chemodynamic/immune therapy via enhanced ferroptosis-mediated immunogenic cell death. ACS Nano. 2022;16:984–996. doi:10.1021/acsnano.1c08585
88. Cao K, Du Y, Bao X, et al. Glutathione-bioimprinted nanoparticles targeting of N6-methyladenosine FTO demethylase as a strategy against leukemic stem cells. Small. 2022;18(13):e2106558. doi:10.1002/smll.202106558
89. Wu F, Du Y, Yang J, et al. Peroxidase-like active nanomedicine with dual glutathione depletion property to restore oxaliplatin chemosensitivity and promote programmed cell death. ACS Nano. 2022:
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
An Magnetic-Targeting Nano-Diagnosis and Treatment Platform for TNBC
Zhang M, Bao S, Qiu G, Liang J, Wang Q, Zhu X, Qin G, Liu J, Zhao C
Breast Cancer: Targets and Therapy 2023, 15:101-119
Published Date: 3 February 2023
Is Emerging Nanomedicine a Friend or Foe to Germ Cells?
Yu H, Hu C, Wang X, Zhang Y, Zhang N, Yu P, Lian K, Huang J, Duan P
International Journal of Nanomedicine 2025, 20:13621-13639
Published Date: 11 November 2025