Back to Journals » Neuropsychiatric Disease and Treatment » Volume 22
Intestinal Flora Reconfiguration via Electroacupuncture: A Strategy to Counteract Depressive-Like Symptoms in Rats
Authors Tan Y, Wu Y, Chen Z, Zhang Z ![]() , Huang S, Li Z, Huang Y
, Huang S, Li Z, Huang Y
Received 10 October 2025
Accepted for publication 13 February 2026
Published 4 March 2026 Volume 2026:22 565614
DOI https://doi.org/10.2147/NDT.S565614
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Yuhui Tan,1,* Yuebei Wu,1,* Zhuying Chen,1 Zhinan Zhang,1 Shengtao Huang,1 Zheng Li,1 Yong Huang1,2
1School of Traditional Chinese Medicine, Southern Medical University, Guangzhou, 510515, People’s Republic of China; 2Department of Acupuncture, Nanfang Hospital, Southern Medical University, Guangzhou, 510515, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yong Huang, Email [email protected] Zhinan Zhang, Email [email protected]
Background: Depression is a global health concern, and acupuncture has emerged as an effective treatment. The role of intestinal microbiota in depression remains unclear. This study, utilizing 16S rRNA high-throughput sequencing, aimed to explore the relationship between electroacupuncture (EA) and depressive behavior by examining changes in the intestinal microbiota.
Materials and Methods: Forty-eight male Sprague-Dawley rats were utilized, with 13 assigned to the normal control (NC) group. The remaining rats underwent a 28-day depression modeling process, and those exhibiting depressive symptoms were randomly divided into chronic unpredictable mild stress (CUMS) and EA groups. The EA group received 14 days of treatment. Behavioral analyses were conducted on rats from the NC, CUMS, and EA groups to assess EA’s effectiveness. Additionally, 16S rRNA sequencing was performed on randomly selected rats from each group.
Results: By examining the behavior of 39 rats and the intestinal microbiota of 18 rats, we found that EA may alter the composition of the intestinal microbiota community structure in CUMS rats, particularly modulating the abundance of Akkermansia in a manner potentially linked to gut–brain axis regulation, including stress- and inflammation-related pathways that may influence microbial composition.
Conclusion: The potential antidepressant impact of electroacupuncture (EA) might be linked to the modulation of Akkermansia abundance within the brain-gut axis.
Keywords: depression, Akkermansia, brain-gut axis, gastrointestinal microbiome, electroacupuncture
Introduction
Depression, characterized by symptoms such as sadness, loss of interest or pleasure, guilt or low self-worth, sleep and appetite disturbances, fatigue, and poor concentration, is the most prevalent mental health problem in the general population.1,2 In its most severe form, depression can even lead to an increased risk of suicide and death.3,4 It is often a chronic condition that significantly impacts an individual’s career potential and overall quality of life.5,6 The standard drug treatments for depressive disorder, mainly selective serotonin reuptake inhibitors (SSRIs), have limitations such as undesirable side effects and delayed onset of action.7,8 Given the high incidence of depressive disorder, the high rate of suicide, and the prevalence of treatment-resistant cases, it is crucial to develop new anti-depression strategies.
Acupuncture shows promise as a non-pharmacological approach to alleviate depressive symptoms, serving as a viable alternative or complementary therapy to enhance treatment outcomes.9 In contrast to pharmacological interventions, acupuncture offers the advantages of cost-effectiveness and minimal side effects.10 Siguan acupoints, namely bilateral LI4 and LR3, are frequently employed acupoints for managing depression. Research has indicated that electroacupuncture targeting the Siguan point holds potential in ameliorating the complexities associated with chronic inflammatory pain and depression,11 affects peripheral neurochemical pathways such as colonic 5-hydroxytryptamine and short-chain fatty acid production,12 and exerts regulatory effects on gut-related inflammatory and metabolic processes.13 Moreover, this intervention has been found to upregulate the expression of BDNF and TrkB in the hippocampus of rats exhibiting post-stroke depression.14 Collectively, these multifaceted effects form a preliminary basis for selecting the Siguan acupoints in the present work. In spite of this, the role of electroacupuncture at Siguan point is still not fully clarified, and its potential mechanism is worthy of further exploration.
The precise antidepressant mechanism of acupuncture likely entails a multi-organ and multi-target involvement, wherein gut microbiota may constitute one of the factors.15 Research has demonstrated the significant involvement of intestinal microbiota in both the etiology and therapeutic approaches for depression. For instance, the fecal microbiota composition is perturbed in individuals diagnosed with major depression.16 Anxiety-depressive-like behaviors improved in male mice after long-term treatment with a specific probiotic, Lactobacillus plantarum.17 Growing evidence from both animal experiments and clinical investigations indicates that dysregulation of gut microbiota composition, as well as the presence of microbe-derived short-chain fatty acids, D-amino acids, and metabolites, significantly contribute to the pathophysiology of depression through the brain-gut-microbiota axis, affecting both neural and immune systems.18–20 Our prior investigations have demonstrated the potential antidepressant efficacy of electroacupuncture through modulation of the intestinal microbiota. Specifically, this effect involves an increase in the abundance of lactic acid bacteria concomitant with a reduction in Staphylococcus aureus levels.21 Despite these findings, research examining the impact of acupuncture on gut microbiota in depression models remains limited. Notably, there exists a scarcity of pertinent observations concerning the specific composition of electroacupuncture-modulated bacteria across distinct acupoints and various species.
This focus on the gut-brain axis represents a new direction of inquiry for our research group. Our previous work on this topic explored a central nervous system mechanism, demonstrating that EA at Governor Vessel (GV20 and GV29) acupoints ameliorates depression by suppressing hippocampal autophagy.22 The present study, therefore, was designed to complement our prior findings by investigating a distinct, peripheral pathway associated with a different set of acupoints (Siguan points), with attention to intestinal microbiota as a key component of gut–brain axis regulation.
Based on accumulating evidence that disturbances in gut microbiota contribute to depressive phenotypes through immune, metabolic, and neural pathways, we hypothesized that EA may ameliorate depressive‐like behaviors in CUMS rats by modulating specific microbial taxa associated with stress- and inflammation-related processes. Thus, the objective of this study was to investigate alterations in the abundance of gut microbiota in mice following electro-acupuncture intervention and CUMS modeling using behavioral assessments and 16s RNA technology. Our aim was to identify specific gut microbiota associated with the antidepressant potentiation of acupuncture, thereby contributing new insights into the mechanisms underlying the antidepressant effects of acupuncture.
Materials and Methods
Animals
Forty-eight adult, male, SD rats, weighing 180–220 g, and aged seven weeks. This experiment has passed the Institutional Animal Care and Use Committee of Southern Medical University, China (No. L2017178). The rats were raised separately in SPF (Specific Pathogen-Free) facilities (temperature 22–26 °C, humidity 50–60% on 12/12 light-dark cycles), under identical housing conditions. Thirty-five rats began to be exposed to chronic unpredictable mild stress (the CUMS group). Rats in the NC group were fed normally without any intervention.
Chronic Unpredictable Mild Stress (CUMS)
The response to CUMS is the most commonly used rodent model of depression, and its reliability and effectiveness have been confirmed.23 The establishment of this model is based on the study of evaluating the biological effects of antidepressants.24,25 Except for the normal control (NC) group, the other 35 rats were exposed to CUMS for 28 days. During this period, rats were exposed to one of the stressors every day, including constant light (24 h), water deprivation (24 h), immobilization (2 h), swimming (5 min), level shaking (5 min), heat stress (5 min), and tail clamping (3 min; 3 cm from the tail end). The same stressor was applied for no more than two days to avoid the occurrence of adaptation. The CUMS protocol was consistent with that used in our prior work to ensure model stability and comparability.22 Successful induction of depressive symptoms was tested with a sucrose preference test (SPT). The SPT was shown to reflect the intrinsic anhedonia of mice. The percentage of sugar/water preference decreased significantly. This was taken as an indication of the success of the replication of the depression model from the perspective of behavior. The rats in CUMS group with less than a 10% sucrose preference decrease were considered insusceptible to CUMS stressors and were excluded.26,27
Depressive symptoms were successfully induced in 26 rats. These 26 were then divided, equally and randomly, between the CUMS group and the EA group. The EA group received electroacupuncture intervention, whereas rats in the CUMS group and the NC group received handling or light immobilization (the same procedures as those of the EA group rats) but without other intervention and were otherwise reared under the same routine conditions.
Intervention
Electroacupuncture of the Siguan acupoints in the 13 rats of the EA group lasted 14 days. The electroacupuncture-intervention points are the Siguan acupoints: a combination of bilateral LI4 and LR3, meaning “four gates” in Chinese.
Acupuncture needles (0.18 mm × 25 mm, Hwato Appliance Factory) were inserted horizontally into bilateral acupoints at LI4 (between the first and second metacarpals of both forelimbs) and LR3 (the indentation between the first and second metatarsals in the dorsum of both hindlimbs) to a depth of 2–3 mm. After insertion, the needles were connected to the electrical stimulator. Electrical simulations with sparse waves (current of 1 mA), frequency of 10–20 Hz, voltage of 5V were applied. The stimulation intensity is considered preferable when the limbs and trunk of the rats vibrate slightly. According to Zhang’s literature, EA was applied for 30 min, once/day for 14 days.28
SPT and Weighting
At the end of the intervention, three behavioral parameters were measured, using procedures previously described by our group.22 Sucrose preference test (SPT), weighing, and open field test (OFT), were conducted to assess depression-like behavior. The rats were euthanized the next day.
The SPT program was performed as mentioned previously.29 Rats were first taught to adjust to 1% (w/v) sucrose solution in their cages, and two bottles of this solution were placed in each cage. After 24 h, tap water was used to replace sugar water. The rats fasted and were deprived of water for 24 h. Rats were kept in separate cages for 1 h of SPT and were exposed to two bottles containing 200 mL water and sucrose solution (1% w/v). In order to prevent position preference, the bottle positions were switched during the test. The sucrose-preference score was calculated as the 1% sucrose solution intake as a percentage of total liquid intake. After the experiment, all the rats were put back into their original cages. Weight was assessed at the conclusion of the intervention.
OFT
OFT was used to measure the activity of the rats, as described in a previous study.30 The open field was a black wooden box, 100 cm × 100 cm × 38 cm, kept in a room with temperature maintained at 21–25 °C, with the light above the box kept uniform and constant. The floor was divided into squares of 25 cm × 10 cm × 10 cm. In order to record the movement of rats in the box, a video recording system was installed above the chamber. Each rat was placed, without being agitated, in the middle square of the field, and its exploratory movements were recorded for five minutes. Each rat was deodorized with 75% ethanol before the experiment, and the whole box was cleaned with 75% ethanol between rats. When the OFT was complete, SMART3.0 software was used to analyze the video and evaluate the locomotor behavior and position in the open field.
Sample Collection and Preparation
Follow the instructions in the fecal microbial genome DNA Extraction method (TIANGEN, Beijing, China). Followed the instructions in the fecal microbial genome DNA Extraction method (TIANGEN, Beijing, China). The V3-V4 region of the 16S rRNA gene was amplified by PCR using universal primers. The original sequence used Mothur version 1.31.2 (http://www.mothur.org/) for quality screening to obtain unique read fragments. Sequences smaller than 200 bp and larger than 1000 bp were excluded, as were the sequences containing primer mismatch, band mismatch, fuzzy base sequence, and homopolymer more than 6 bases. The RDP reference data base, which contains 20,712 bacterial and 601 archaeal 16S rRNA gene sequences, was used to assign all remaining sequences to the operational taxon with a 97% threshold match. The key operational classification units were identified using a random forest algorithm. Based on the results of species annotation and operational taxonomic unit (OTU), the analysis of species diversity and species complexity among sample groups was carried out; it includes bioinformatics analysis, species composition analysis, alpha diversity analysis, beta diversity analysis, species diversity analysis and Marker Species, association network analysis, and functional potential prediction.
Euthanasia
Rats were anesthetized through peritoneal injection of pentobarbital sodium at doses ranging from 30 to 90 mg/kg, based on their individual weights. Following anesthesia, the rats were positioned with their tails lifted using the experimenter’s right hand, placing it on the lid of the rat cage or another textured surface. The head and neck were gently secured using the experimenter’s left thumb and index finger, while the root of the tail was pulled with the right hand, inducing cervical dislocation. Subsequently, the spinal cord was effectively disconnected from the brainstem, resulting in immediate euthanasia of the animal.
Statistical Analysis
The data were analyzed using various statistical methods based on the data distribution. The independent samples t-test was employed for normally distributed data, while the nonparametric Kruskal–Wallis H-Test was utilized for non-normally distributed data. Pearson correlation analysis or Spearman correlation analysis was applied as appropriate. Levene’s test was used to assess the variances between the two groups. In cases where the variances were found to be dissimilar, adjusted p-values were used. Furthermore, we generated summaries of the taxonomic distribu-tions of OTUs to calculate the relative abundances of gut microbiota at different levels. Alpha diversity was assessed using different parameters, namely Shannon, Simpson, phylogenetic di-versity, and Chao, etc. Beta diversity, represented by distance matrices between samples, was evaluated through principal coordinate analysis (PCoA). The Random Forest algorithm was em-ployed to identify key discriminatory OTUs. Additionally, the linear discriminant-analysis effect size (LEfSe) method was applied to identify dominant bacterial taxa in both control and de-pressed mice. A significance level of P< 0.05 was considered statistically significant. The method of Benjamini and Hochberg False Discovery was used for multiple testing corrections.
Results
Behavior Analysis
Three groups (the NC, CUMS, and EA groups), with n = 13/group, were tested. The performance of the CUMS model was stable during treatment, and the behavioral results in both the CUMS and EA groups were significantly different from those in the normal control group (p < 0.05). In addition, the EA group also showed positive results relative to the CUMS group. EA treatment significantly improved depression-like behavior in rats (p < 0.05). The analysis results are shown in Figure 1.
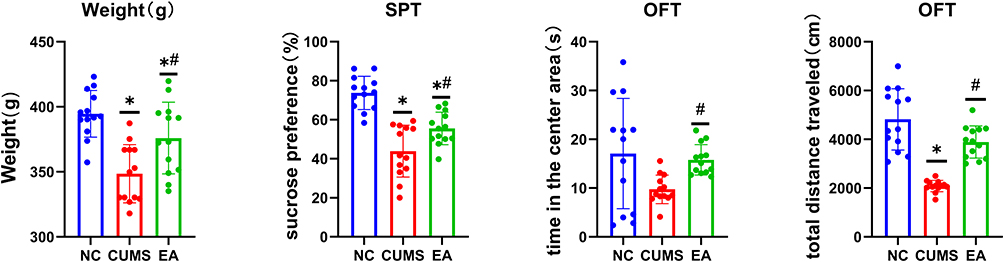 |
Figure 1 Behavior Analysis. *p<0.05 vs NC group; #p<0.05 vs CUMS group. |
Alpha Diversity Analysis
Alpha diversity, also known as within-habitat diversity, is an indicator of species richness, diversity, and evenness in locally uniform habitats. Alpha diversity is calculated by constructing curves based on sequence numbers and the OTU numbers represented by these sequences. Faith’s PD index (p = 0.02) indicated that the sample has diversity based on evolution, as shown in Figure 2A. In each panel, the abscissa is the grouping label, and the ordinate is the value of the corresponding alpha diversity index. In the box diagram, the meanings of each symbol are as follows: the upper and lower end lines of the box, the upper and lower quartile (IQR). Middle bit line, median; Upper and lower edges, maximum and minimum inner circumference (1.5 × IQR).
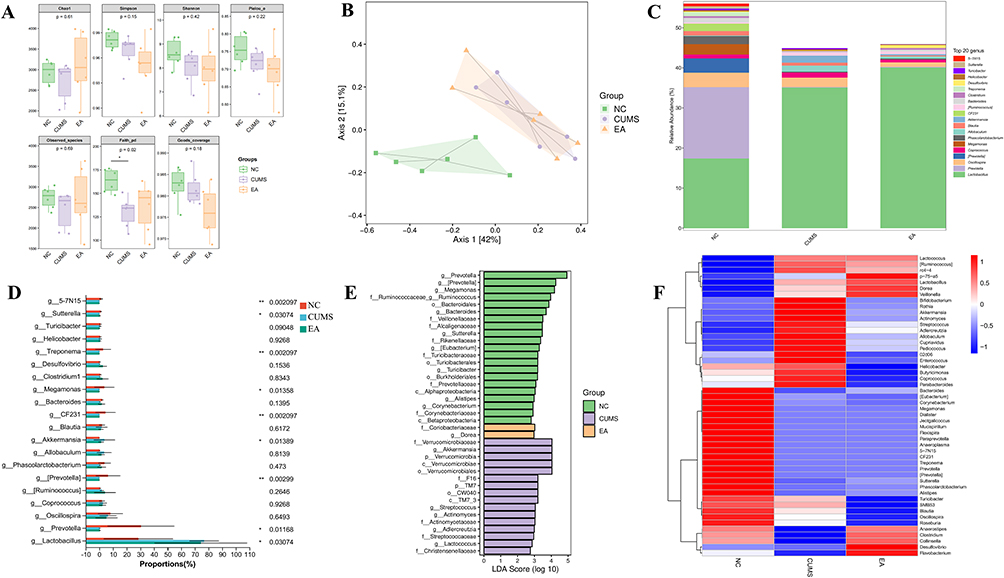 |
Figure 2 Effects of electroacupuncture on intestinal microbiota composition in CUMS rats. (A) α-diversity analysis. (B) β-diversity analysis. (C) Analysis of genus taxonomic species composition. (D) Differential analysis of genus taxonomic species composition; *p < 0.05, **p < 0.01. (E) LEfSe analysis. (F) Heat map. Abbreviations: NC, normal control; CUMS, chronic unpredictable mild stress; EA, electroacupuncture; LEfSe, linear discriminant analysis effect size. |
Beta Diversity Analysis
The Beta diversity index focuses on the comparison of diversity between different habitats, that is, on the differences between samples. The so-called beta diversity distance, which reduces the species abundance, characteristics and evolutionary relationships of representative sequences in a sample to one-dimensional data, or sample difference distance, by using different methods, describes the population differences between two samples from different perspectives.
PCoA analysis was performed using R language and the results are shown in Figure 2B. Each sample is represented by a dot in the figure, and groups are indicated by dots of different colors. For each axis, the brackets represent the proportions of the distance matrix. The physical unit length ratio of the two axes was set to a default of 1, which was different from the ratio of the explanatory degree. In this case, projection analysis was recommended instead of looking at the distance directly. That is, the closer the projection of the two points, the more similar the community composition of the sample. In the figure, the dotted circle is a 95% confidence ellipse. The NC group shows obvious separation from the other two groups, and the EA group tended to be closer to the NC group.
Species Composition Analysis
At each classification level, the specific composition tables of microbial communities can be derived by counting the flattened ASV (amplicon sequence variants)/OTU tables. Using this tables, we first calculated the number of taxa in different samples at each classification level. The number of microbial taxonomic units in the NC group was significantly higher than that in the CUMS and EA groups, indicating that the NC group exhibited greater microbial diversity. The genus-level taxonomic composition was subsequently visualized based on the average relative abundance table generated from the 16S rRNA sequencing data using the Mothur pipeline and RDP reference database, as shown in Figure 2C.
The taxonomic distribution of species within each group was visually depicted using the average genus-level species abundance table and subsequently analyzed employing the Kruskal–Wallis H-test. In Figure 2D, discernible differences in the relative abundance of Lactobacillus, Prevotella, [Prevotella], Akkermansia, CF231, Megamonas, Treponema, Sutterella, and 5–7N15 were observed among the three groups (P = 0.03074, 0.01168, 0.00299, 0.01389, 0.002097, 0.01358, 0.002097, 0.03074, 0.002097). The relative abundance of Lactobacillus and Akkermansia increased following CUMS but decreased after EA. Conversely, the relative abundance of Prevotella, [Prevotella], CF231, Megamonas, Treponema, Sutterella, and 5–7N15 decreased after CUMS and showed minimal change after EA.
Species Difference and Marker Species Analysis
Using the LEfSe analysis method, comparisons between two or more groups can be realized, and the comparative analysis between subgroups can also be carried out, so that species with significant differences in abundance between groups can be found. There are 39 taxa with LDA scores >2 that are significant in four groups, among which are 21 taxa in the NC group (Prevotella group, Megamonas, Bacteroidales, etc)., of which 2 were identified in the EA group (Coriobacteriaceae, Dorea). In the CUMS group, 16 were identified (Verrucomicrobiaceae, Akkermansia, Verrucomicrobia, Verrucomicrobiae, Verrucomicrobias, etc) in Figure 2E the ordinate is the taxon with significant differences between groups, and the abscissa shows the logarithmic score of the LDA analysis of each taxon intuitively by bar graph. Classification units are sorted according to the score value, so as to describe their specificity in sample grouping. The longer the length is, the more significant the difference of the taxon is, and the color of the bar graph indicates.
Heat maps, as shown in Figure 2F can be used for species composition analysis to display trends in species abundance distribution between samples and to compare species composition differences between samples. Red indicates a higher abundance of this genus in this sample, whereas blue indicates the opposite. The CUMS group shows a greater abundance of Bacteroidetes and Proteobacteria and a decreased abundance of Firmicutes than did the NC group. Blautia, Roseburia, and Anaerostipes have anti-inflammatory effects and belong to probiotics. Compared with the NC group, the abundance of them in the CUMS group is lower, while slightly higher in the EA group.31–33
Discussion
The behavioral outcomes indicated reduced body weight, sucrose preference, and travel distance in rats from the CUMS group compared to the NC group, signifying heightened depressive symptoms in the former. These results collectively verify the successful establishment of the CUMS model.
Depressive symptoms in rats in the EA group, after EA treatment, were less severe than those in the CUMS group. Sucrose preference and activity distance of rats in the EA group were greater than those in the CUMS group. These findings indicate that EA at Siguan acupoints is capable of mitigating depressive-like symptoms in CUMS rats. Interestingly, these results complement our previous findings, where we demonstrated that EA at GV20 and GV29 modulates central mechanisms by inhibiting hippocampal autophagy.22 The present study, by employing Siguan acupoints, suggests a distinct therapeutic mechanism involving the reconfiguration of gut microbiota. Taken together, these findings support the notion that EA’s antidepressant effects may involve multiple pathways and that the underlying mechanism may be highly dependent on the choice of acupoints. Prior studies suggest that Governor Vessel points may influence central nervous processes, whereas peripheral acupoints such as Siguan have been hypothesized to affect systemic pathways including the gut–brain axis. In the present experiment, the bilateral LI4 and LR3 (Siguan) acupoints are used, which are supposed to have the effect of soothing the liver, relieving depression, calming the nerves, and smoothing qi.14 A previous experiment using rats also observed the antidepressant effects of LI4 and LR3 stimulation, which may be related to the regulation of the expression of BDNF and TrkB in the brains of depressed rats.14 In addition, another animal experiment observe that the change of intestinal microbiota was related to the improvement of depressive symptoms.34 Our findings are consistent with the above studies, stimulation of LI4 and LR3 is accompanied by improvements in depressive-like behavior and by concurrent alterations in gut microbiota composition.
The α and β diversity analyses revealed higher species richness in the NC group compared to the CUMS and EA groups. From a species composition perspective, the EA group exhibited closer resemblance to the NC group. These findings underscore the significant involvement of intestinal microflora richness in depression’s pathogenesis, wherein reduced richness correlates with heightened susceptibility to depression. Greater bacterial diversity contributes to the equilibrium of diverse flora and metabolites, thereby constraining the proliferation of detrimental bacterial species.35
Through the analysis of the composition and abundance of intestinal flora, it was found that Lactobacillus and Akkermansia were the only bacteria that had statistical differences among the three groups and changed after the intervention of EA. Further supported by LEfSe analysis, it was revealed that only Akkermansia exhibited an LDA score greater than 2. Although Lactobacillus also showed significant differences, its abundance changes appeared to be context-dependent and varied across experimental conditions and analytical approaches.21,36 Accordingly, Lactobacillus was not prioritized as the primary taxon for downstream mechanistic interpretation in the present study. This points to the potential involvement of Akkermansia in the mechanism underlying the amelioration of depression-like behavior in rats.
Akkermansia muciniphila (A. muciniphila), an anaerobic gram-negative bacterium, represents the exclusive member of Verrucomicrobia within the human intestinal ecosystem. Widely distributed in the mucus layer, it exhibits oval non-motile cells and lacks endospore-forming capability.37 This microbe exclusively harnesses mucins for nitrogen, carbon, and energy, displaying white colonies on soft agar mucin medium.38 Constituting around 3–5% of the gut microbiota in healthy individuals, A. muciniphila’s prominence is further demonstrated by its presence among the top 20 abundant species in the human gut.38,39 Notably, its colonization and abundance are subject to multifaceted influences including age, location, genotype, dietary patterns, and the host’s physiological status.40 While detected across various segments of human mucosa and feces, A. muciniphila particularly thrives in the mucin-rich cecum.41
Akkermansia, considered a promising next-generation probiotic, is proposed to modulate the 5-HTR1A-CREB-BDNF signaling cascade through interactions with TLR5 via Amuc_1100Δ80, a mucin outer membrane protein, thus reshaping gut microbial composition to mitigate depressive-like behaviors in CUMS mice.42,43 Ding et al demonstrated enhanced depression alleviation through oral Akkermansia supplementation, involving modulation of cholinergic synapses, fat metabolism, aromatic compound degradation, vitamin absorption, fatty acid breakdown, butyrate utilization, carbon metabolism, as well as biosynthesis of pantothenic acid and coenzyme A.44 This study delineated the potential of Akkermansia to influence metabolic pathways, digestion, and absorption for depression improvement.
However, it has also been suggested that heightened Akkermansia abundance might trigger excessive intestinal mucin consumption, resulting in mucosal disruption, escalated intestinal inflammation, and noxious substance absorption.45,46 Wang et al observed that the introduction of Myxophilus supplementation to a context of profound microbial dysbiosis compromised the integrity of the intestinal barrier. This disruption facilitated the establishment of an environment conducive to disease progression, ultimately culminating in the development of colitis-associated colorectal cancer in a murine model.45 Under inflammatory conditions, Ganesh et al documented Akkermansia’s transition into a pathogenic state, exacerbating intestinal inflammation in murine models.46 Furthermore, increased abundance of Akkermansia has been observed in certain psychiatric conditions, including Alzheimer’s disease.46,47 Our findings parallel these studies: reduced intestinal microbial diversity in CUMS-affected mice correlated with escalated depression-like behaviors and heightened Akkermansia abundance, which was counteracted by EA intervention. Based on these findings, we propose that CUMS exposure is associated with maladaptive alterations in the gut microbial milieu in mice. Within this context, changes in Akkermansia abundance may reflect altered host–microbe interactions at the intestinal mucosal interface, potentially involving shifts in mucin utilization. Such mucosal-level alterations could influence gut–brain axis signaling, thereby contributing to the modulation of depressive-like behaviors. Accordingly, further studies integrating targeted mechanistic experiments are required to delineate the precise pathways linking electroacupuncture, gut microbiota modulation, and behavioral outcomes.
However, there is limited direct evidence regarding the effects of A. muciniphila on neuropsychiatric disorders. The prevailing body of clinical research has predominantly centered around establishing correlations between the bacterium and these disorders, lacking substantial causal inference. Nonetheless, multiple theoretical frameworks elucidating the mechanisms by which A. muciniphila potentially influences brain function have emerged.48 These frameworks encompass the generation of neuroactive metabolites and derivatives, fortification of the gastrointestinal mucosal barrier, orchestration of host immune responses, and modulation of metabolic processes. It is imperative to progressively amass pertinent data and empirical substantiation within these theoretical constructs.
Our study has inherent limitations. Firstly, we did not further investigate the precise mechanistic link between Akkermansia and depression, which limits causal interpretation of the observed association. Secondly, fecal microbiota transplantation experiments were not conducted, and the absence of pertinent markers like BDNF, inflammatory biomarkers, and intestinal histopathological staining limits functional and mechanistic validation of the gut–brain axis involved in the observed effects. Subsequently, further comprehensive investigations are warranted to determine whether Akkermansia plays a causal role in depression.
Conclusion
Through analysis of intestinal microbiota abundance and composition, we identified an association between the antidepressant effects of electroacupuncture (EA) and alterations in the gut microbiota. EA intervention was accompanied by changes in microbial community structure in CUMS rats, among which a reduction in Akkermansia was observed following treatment. Although these findings suggest that gut microbiota alterations may contribute to the antidepressant effects of EA, the role of Akkermansia in depression remains controversial, and further studies are required to clarify the underlying mechanisms.
Data Sharing Statement
The figure data used to support the findings of this study are available from the corresponding author via Email at [email protected].
Ethics Approval and Consent to Participate
The experiments were conducted in accordance with the NIH Guidelines for the Care and Use of Animals and were approved by the Animal Experimentation Ethics Committee of Southern Medical University (No. L2017178).
Funding
This work was supported by the National Natural Science Foundation of China (82074519), Natural Science Foundation of Guangdong Province, China (2022A1515011658, 2023A1515012235), Science and Technology Projects in Guangzhou, China (2024A04J3988), Administration of Traditional Chinese Medicine of Guangdong Province, China (20241197).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Lim GY, Tam WW, Lu Y, et al. Prevalence of Depression in the Community from 30 Countries between 1994 and2014. Sci Rep. 2018;8(1):2861. doi:10.1038/s41598-018-21243-x
2. Sinyor M, Rezmovitz J, Zaretsky A. Screen all for depression. BMJ. 2016;352:i1617. doi:10.1136/bmj.i1617
3. Choo C, Diederich J, Song I, Ho R. Cluster analysis reveals risk factors for repeated suicide attempts in amulti-ethnic Asian population. Asian J Psychiatr. 2014;8:38–10. doi:10.1016/j.ajp.2013.10.001
4. Large M. Study on suicide risk assessment in mental illness underestimates inpatientsuicide risk. BMJ. 2016;532:i267. doi:10.1136/bmj.i267
5. Daly EJ, Trivedi MH, Wisniewski SR, et al. Health-related quality of life in depression: a STAR*D report. Ann Clin Psychiatry. 2010;22(1):43–55. doi:10.1177/104012371002200107
6. Mayor S. Persistent depression doubles stroke risk despite treatment, study finds. BMJ. 2015;350:h2611. doi:10.1136/bmj.h2611
7. Rush AJ, Trivedi MH, Wisniewski SR, et al. Acute and longer-term outcomes in depressed outpatients requiring one or severaltreatment steps: a STAR*D report. Am J Psychiatry. 2006;163(11):1905–1917. doi:10.1176/ajp.2006.163.11.1905
8. Wang Z, Wang X, Liu J, et al. Acupuncture treatment modulates the corticostriatal reward circuitry in majordepressive disorder. J Psychiatr Res. 2017;84:18–26. doi:10.1016/j.jpsychires.2016.09.014
9. Smith CA, Armour M, Lee MS, Wang LQ, Hay PJ. Acupuncture for depression. Cochrane Database Syst Rev. 2018;3(3):CD004046. doi:10.1002/14651858.CD004046.pub4
10. Chan YY, Lo WY, Yang SN, Chen YH, Lin JG. The benefit of combined acupuncture and antidepressant medication for depression: A systematic review and meta-analysis. J Affect Disord. 2015;176:106–117. doi:10.1016/j.jad.2015.01.048
11. Shin KM, Park JE, Lee S, et al. Effect of siguan acupuncture on gastrointestinal motility: a randomized, sham-controlled, crossover trial. Evid Based Complement Alternat Med. 2013;2013:918392. doi:10.1155/2013/918392
12. Xu H, Li L, Kang Z, et al. Effects of electroacupuncture at “Siguan” points on the expression of colonic 5-hydroxytryptamine and fecal short-chain fatty acids in rats with post-stroke depression. Chin AcupuncT Moxibustion. 2023;43(05):545–551. doi:10.13703/j.0255-2930.20221125-k0002
13. Li Z, Li L, Xu H, et al. Modulating gut microbiota: the mechanism of electroacupuncture at the “Siguan” acupoints in alleviating post-stroke depression. Neuropsychiatr Dis Treat. 2025;21:281–294. doi:10.2147/NDT.S495460
14. Kang Z, Ye H, Chen T, Zhang P. Effect of electroacupuncture at siguan acupoints on expression of BDNF and TrkBProteins in the hippocampus of post-stroke depression rats. J Mol Neurosci. 2021;71(10):2165–2171. doi:10.1007/s12031-021-01844-4
15. Yang NN, Lin LL, Li YJ, et al. Potential mechanisms and clinical effectiveness of acupuncture in depression. Curr Neuropharmacol. 2022;20(4):738–750. doi:10.2174/1570159X19666210609162809
16. Jiang H, Ling Z, Zhang Y, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav Immun. 2015;48:186–194. doi:10.1016/j.bbi.2015.03.016
17. Barros-Santos T, Silva K, Libarino-Santos M, et al. Effects of chronic treatment with new strains of Lactobacillus plantarum oncognitive, anxiety- and depressive-like behaviors in male mice. PLoS One. 2020;15(6):e0234037. doi:10.1371/journal.pone.0234037
18. Foster JA, McVey NK. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013;36(5):305–312. doi:10.1016/j.tins.2013.01.005
19. Yang Z, Li J, Gui X, et al. Updated review of research on the gut microbiota and their relation to depressionin animals and human beings. Mol Psychiatry. 2020;25(11):2759–2772. doi:10.1038/s41380-020-0729-1
20. Chang L, Wei Y, Hashimoto K. Brain-gut-microbiota axis in depression: a historical overview and futuredirections. Brain Res Bull. 2022;182:44–56. doi:10.1016/j.brainresbull.2022.02.004
21. Qiu X, Li Z, Huang S, et al. Electroacupuncture improves depression-like behavior by regulating the abundance of lactobacillus and staphylococci in mice. J Integr Neurosci. 2023;22(2):28. doi:10.31083/j.jin2202028
22. Zhang Z, Cai X, Yao Z, et al. EA ameliorated depressive behaviors in CUMS rats and was related to its suppressing autophagy in the hippocampus. Neural Plast. 2020;2020:8860968. doi:10.1155/2020/8860968
23. Antoniuk S, Bijata M, Ponimaskin E, Wlodarczyk J. Chronic unpredictable mild stress for modeling depression in rodents: Meta-analysis of model reliability. Neurosci Biobehav Rev. 2019;99:101–116. doi:10.1016/j.neubiorev.2018.12.002
24. Lu Y, Ho CS, McIntyre RS, Wang W, Ho RC. Effects of vortioxetine and fluoxetine on the level of Brain Derived NeurotrophicFactors (BDNF) in the hippocampus of chronic unpredictable mild stress-induceddepressive rats. Brain Res Bull. 2018;142:1–7. doi:10.1016/j.brainresbull.2018.06.007
25. Chen B, Li J, Xie Y, et al. Cang-ai volatile oil improves depressive-like behaviors and regulates DA and 5-HTmetabolism in the brains of CUMS-induced rats. J Ethnopharmacol. 2019;244:112088. doi:10.1016/j.jep.2019.112088
26. Xie H, Huang H, Tang M, et al. iTRAQ-based quantitative proteomics suggests synaptic mitochondrial dysfunctionin the hippocampus of rats susceptible to chronic mild stress. Neurochem Res. 2018;43(12):2372–2383. doi:10.1007/s11064-018-2664-y
27. Zhang J, Zhang J, Zhang Z, et al. Dopaminergic signaling in prefrontal cortex contributes to the antidepressant effect of electroacupuncture: aniTRAQ‐basedproteomics analysis in a rat model ofCUMS. Anat Record. 2021;304(11):2454–2469. doi:10.1002/ar.24732
28. Yao Z, Zhang Z, Zhang J, et al. Electroacupuncture alleviated the depression-like behavior by regulating FGF2 andastrocytes in the hippocampus of rats with chronic unpredictable mild stress. Brain Res Bull. 2021;169:43–50. doi:10.1016/j.brainresbull.2021.01.005
29. Liu MY, Yin CY, Zhu LJ, et al. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat Protoc. 2018;13(7):1686–1698. doi:10.1038/s41596-018-0011-z
30. Bhat AH, Prabhu P, Balakrishnan K. A critical analysis of state-of-the-art metagenomics OTU clustering algorithms. J Biosci. 2019;44(6). doi:10.1007/s12038-019-9964-5
31. Allen-Vercoe E, Daigneault M, White A, et al. Anaerostipes hadrus comb. nov. a dominant species within the human colonicmicrobiota; reclassification of Eubacterium hadrum Moore et al. 1976. Anaerobe. 2012;18(5):523–529. doi:10.1016/j.anaerobe.2012.09.002
32. Tamanai-Shacoori Z, Smida I, Bousarghin L, et al. Roseburia spp.: a marker of health? Future Microbiol. 2017;12:157–170. doi:10.2217/fmb-2016-0130
33. Liu X, Mao B, Gu J, et al. Blautia-a new functional genus with potential probiotic properties? Gut Microbes. 2021;13(1):1–21. doi:10.1080/19490976.2021.1875796
34. Zhu HZ, Liang YD, Ma QY, et al. Xiaoyaosan improves depressive-like behavior in rats with chronic immobilizationstress through modulation of the gut microbiota. Biomed Pharmacother. 2019;112:108621. doi:10.1016/j.biopha.2019.108621
35. Liu L, Wang H, Chen X, et al. Gut microbiota and its metabolites in depression: from pathogenesis to treatment. EBioMedicine. 2023;90:104527. doi:10.1016/j.ebiom.2023.104527
36. Wang S, Ishima T, Zhang J, et al. Ingestion of Lactobacillus intestinalis and Lactobacillus reuteri causes depression- and anhedonia-like phenotypes in antibiotic-treated mice via the vagus nerve. J Neuroinflammation. 2020;17:241. doi:10.1186/s12974-020-01916-z
37. Derrien M, Vaughan EE, Plugge CM, de Vos WM. Akkermansia muciniphila gen. nov. sp. nov. a human intestinal mucin-degradingbacterium. Int J Syst Evol Microbiol. 2004;54(Pt 5):1469–1476. doi:10.1099/ijs.0.02873-0
38. Belzer C, de Vos WM. Microbes inside--from diversity to function: the case of Akkermansia. ISME J. 2012;6(8):1449–1458. doi:10.1038/ismej.2012.6
39. Qin J, Li R, Raes J, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464(7285):59–65. doi:10.1038/nature08821
40. Zhai Q, Feng S, Arjan N, Chen W. A next generation probiotic, Akkermansia muciniphila. Crit Rev Food Sci Nutr. 2019;59(19):3227–3236. doi:10.1080/10408398.2018.1517725
41. Derrien M, Van Baarlen P, Hooiveld G, et al. Modulation of mucosal immune response, tolerance, and proliferation in mice colonized by the mucin-degrader Akkermansia muciniphila. Front Microbiol. 2011;2:166. doi:10.3389/fmicb.2011.00166
42. Cheng R, Zhu H, Sun Y, Hang T, Zhang M. The modified outer membrane protein Amuc_1100 of Akkermansia muciniphila improveschronic stress-induced anxiety and depression-like behavior in mice. Food Funct. 2022;13(20):10748–10758. doi:10.1039/D2FO01198K
43. Sun Y, Zhu H, Cheng R, Tang Z, Zhang M. Outer membrane protein Amuc_1100 of Akkermansia muciniphila alleviatesantibiotic-induced anxiety and depression-like behavior in mice. Physiol Behav. 2023;258:114023. doi:10.1016/j.physbeh.2022.114023
44. Ding Y, Bu F, Chen T, et al. A next-generation probiotic: Akkermansia muciniphila ameliorates chronicstress-induced depressive-like behavior in mice by regulating gut microbiota andmetabolites. Appl Microbiol Biotechnol. 2021;105(21–22):8411–8426. doi:10.1007/s00253-021-11622-2
45. Wang K, Wu W, Wang Q, et al. The negative effect of Akkermansia muciniphila-mediated post-antibioticreconstitution of the gut microbiota on the development of colitis-associatedcolorectal cancer in mice. Front Microbiol. 2022;13:932047. doi:10.3389/fmicb.2022.932047
46. Ganesh BP, Klopfleisch R, Loh G, Blaut M. Commensal Akkermansia muciniphila exacerbates gut inflammation in SalmonellaTyphimurium-infected gnotobiotic mice. PLoS One. 2013;8(9):e74963. doi:10.1371/journal.pone.0074963
47. Khedr EM, Omeran N, Karam-Allah RH, Ahmed GK, Abdelwarith AM. Alteration of gut microbiota in Alzheimer’s disease and their relation to the cognitive impairment. J Alzheimers Dis. 2022;88(3):1103–1114. doi:10.3233/JAD-220176
48. Lei W, Cheng Y, Gao J, et al. Akkermansia muciniphila in neuropsychiatric disorders: friend or foe? Front Cell Infect Microbiol. 2023;13:1224155. PMCID: PMC10051028. doi:10.3389/fcimb.2023.1224155
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.