Back to Journals » Journal of Inflammation Research » Volume 18
Interleukin-31 in Respiratory Type 2 Inflammatory Diseases: Mechanistic Insights and Therapeutic Potential
Authors Yao S, Zhao Y, Li Z, Lou H, Zhang L ![]()
Received 6 July 2025
Accepted for publication 30 September 2025
Published 27 October 2025 Volume 2025:18 Pages 14881—14890
DOI https://doi.org/10.2147/JIR.S551942
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Shuang Yao,1,2,* Yan Zhao,1,2,* Zhimeng Li,3,4 Hongfei Lou,5,* Luo Zhang1,2,4,6,*
1Department of Otolaryngology Head and Neck Surgery, Beijing Tongren Hospital, Capital Medical University, Beijing, People’s Republic of China; 2Beijing Laboratory of Allergic Diseases, Beijing Municipal Education Commission and Beijing Key Laboratory of New Medicine and Diagnostic Technology Research for Nasal Disease, Beijing Institute of Otolaryngology, Beijing, People’s Republic of China; 3Department of Immunology, CAMS Key Laboratory T Cell and Cancer Immunotherapy, Institute of Basic Medical Sciences, Chinese Academy of Medical Sciences and School of Basic Medicine, Peking Union Medical College, State Key Laboratory of Common Mechanism Research for Major Diseases, Beijing, People’s Republic of China; 4Research Unit of Diagnosis and Treatment of Chronic Nasal Diseases, Chinese Academy of Medical Sciences, Beijing, People’s Republic of China; 5Department of Otolaryngology, Eye & ENT Hospital, Fudan University, Shanghai, People’s Republic of China; 6Department of Allergy, Beijing Tongren Hospital, Capital Medical University, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hongfei Lou, Department of Otolaryngology, Eye & ENT Hospital, Fudan University, 3 Fenyang Road, Xuhui District, Shanghai, 200031, People’s Republic of China, Email [email protected] Luo Zhang, Department of Otolaryngology Head and Neck Surgery, Beijing Tongren Hospital, Capital Medical University, No. 1, Dongjiaominxiang St, Dongcheng District, Beijing, 100730, People’s Republic of China, Email [email protected]
Abstract: Respiratory type 2 inflammatory diseases, including asthma and allergic rhinitis (AR), and chronic rhinosinusitis with nasal polyps (CRSwNP), are characterized by epithelial dysfunction, immune dysregulation, and chronic airway inflammation. Interleukin-31 (IL-31), primarily produced by Th2 cells and other immune and structural cells, has emerged as a pivotal mediator in these conditions. By activating the JAK/STAT and MAPK signaling, IL-31 drives chemokine release, eosinophil recruitment, mucus hypersecretion, and neuroimmune responses. Clinical studies demonstrate that IL-31 levels are elevated and correlate with disease severity, positioning IL-31 as a promising biomarker for diagnosis, prognosis, and treatment response monitoring. Importantly, IL-31 uniquely links type 2 inflammation with sensory nerve dysfunction, addressing unmet clinical needs not fully resolved by conventional IL-4/IL-5–targeted therapies. In this review, we synthesize current mechanistic and clinical evidence on IL-31 in asthma, AR, and CRSwNP, highlight its distinct value compared with other cytokines, and outline the major challenges and research questions that remain unanswered. We further discuss potential translational strategies, including biomarker development and IL-31–targeted interventions, which may provide new opportunities for precision medicine in respiratory type 2 inflammatory diseases.
Keywords: IL-31, respiratory inflammatory diseases, T type 2 cells, mechanism of action, therapeutic targets
Introduction
Type 2 inflammation is predominantly mediated by T type 2 cells, type II innate lymphoid cells (ILC2s) and their secreted cytokines, including IL-4, IL-5, and IL-13. Its key pathophysiological characteristics include epithelial dysfunction, excessive mucous gland secretion, immune cell infiltration, airway remodeling, and neuroimmune interactions.1–3 The term “ Respiratory Type 2 Inflammatory Diseases” does not represent a single clinical diagnosis but rather serves as a conceptual umbrella for a group of clinically heterogeneous chronic airway conditions that are unified by these core mechanisms of Type 2 inflammation. This grouping encompasses a spectrum of disorders affecting both the upper and lower airways, with prototypical examples including asthma, AR, and CRSwNP. Although clinically distinct, their shared immunopathological pathway often leads to significant comorbidity and mutual exacerbation. Given their chronic and recurrent nature, these diseases profoundly impair patient quality of life and impose a substantial socioeconomic burden. Therefore, a deeper understanding of their shared pathogenetic mechanisms to facilitate the development of precision-targeted therapies is a critical and urgent medical need.
IL-31 is a member of the interleukin 6 (IL-6) cytokine family and is mainly produced by activated T cells, especially T type 2 cells.4 Additionally, IL-31 is also secreted by dendritic cells, eosinophils, M2 macrophages, keratinocytes, and fibroblasts.5–7 Unlike other IL-6 family members, IL-31 does not signal through gp130 receptor subunit. Instead, it forms a heterodimers receptor complex composed of IL-31-specific receptor α (IL-31RA) and oncostatin M receptor β (OSM Rβ).4 The IL-31 receptor complex is highly expressed in epithelial cells and keratinocytes,4 as well as in gastrointestinal tract, smooth muscle cells, dorsal root ganglia, and immune cells including eosinophils, basophils, mast cells, and macrophages (Figure 1).8–10 Ip et al and Chattopadhyay, et al investigated the IL-31-activated signaling pathways using bronchial epithelial cell line (BEAS-2B) and alveolar epithelial cell line (A549), respectively. Their studies revealed that in the pulmonary epithelium, IL-31 activates JAK-STAT and MAPK pathways (including ERK, JNK, and p38 MAPK), exhibiting greater activity compared to other cytokines of the IL-6 family.11,12 However, research on IL-31-activated signaling pathways in other respiratory diseases remains limited. Moreover, the interactions between these pathways and their specific regulatory mechanisms across various respiratory conditions are not well understood and require further investigation.
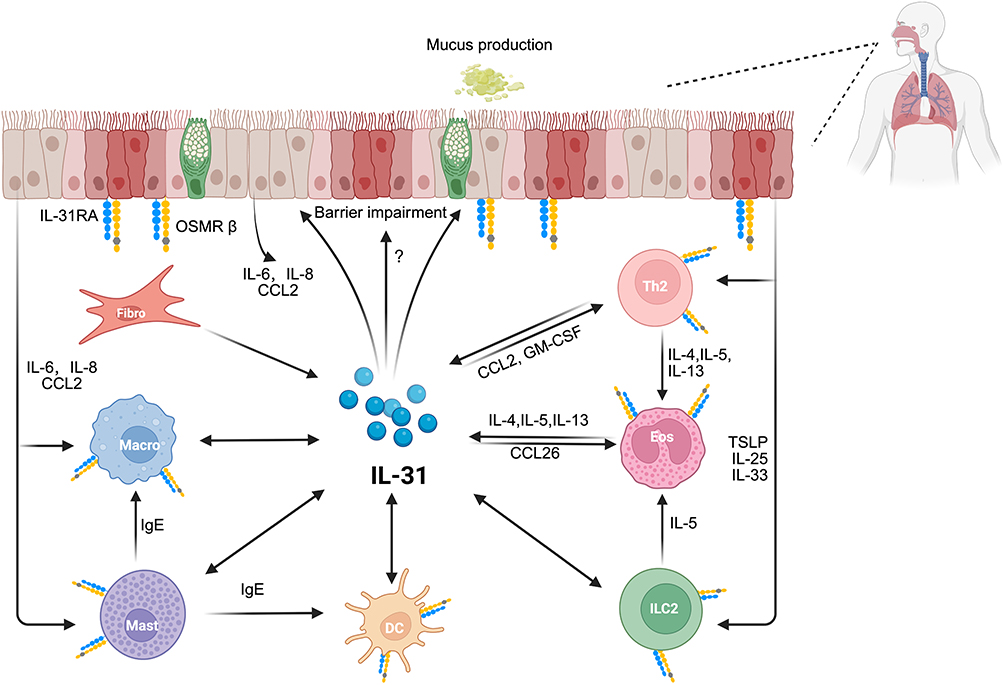 |
Figure 1 The multifaceted mechanistic role of IL-31 in respiratory type 2 inflammatory diseases. This figure illustrates the central role of Interleukin-31 (IL-31) in the pathogenesis of respiratory type 2 inflammation. IL-31 is produced by a variety of immune and structural cells, including T helper 2 cells, type 2 innate lymphoid cells, eosinophils, mast cells, and fibroblasts. Subsequently, by acting on multiple target cells, it orchestrates a complex inflammatory cascade, activating various immune cells to release a spectrum of type 2 cytokines (eg, IL-4, IL-5, IL-13), chemokines (eg, CCL2), and IgE that amplify the allergic inflammatory response. Although the expression of its receptor on airway epithelial cells has been confirmed, the comprehensive impact of IL-31 on epithelial function remains unclear. Unidirectional arrows indicate secretion of a molecule or its action upon a target cell. Bidirectional arrows suggest a feedback loop, where a cell type can both produce IL-31 and be activated by it. Abbreviations: DC, dendritic cell; Eos, eosinophil; Fibro, fibroblast; GM-CSF, granulocyte-macrophage colony-stimulating factor; ILC2, type 2 innate lymphoid cell; Macro, macrophage; Mast, mast cell; OSMRβ, oncostatin M receptor β; Th2, T helper 2 cell. |
While IL-31 has been extensively studied in the context of atopic dermatitis, yielding promising clinical results as a therapeutic target,13–16 its role in respiratory type 2 inflammatory diseases remains a comparatively underexplored and emerging field. Nonetheless, nascent evidence suggests its potential involvement in airway pathogenesis, possibly through contributions to mucus overproduction, airway remodeling, and sustained inflammatory responses. Recent studies have primarily focused on its interactions with immune and epithelial cells, and its ability to promote the production of pro-inflammatory mediators.17–19 Although research on IL-31 in respiratory diseases remains in its early stages, current findings underscore its pivotal role in regulating inflammation, modulating immune responses, and contributing to tissue remodeling. Further elucidation of its mechanistic involvement in respiratory type 2 inflammatory diseases will provide a solid scientific foundation for the development of novel therapeutic strategies.
This review summarizes the role of IL-31 in respiratory type 2 inflammatory diseases and outlines recent research advance. It provides a foundation for future basic and clinical investigations, supports the development of IL-31-targeted therapies and offers new perspectives for improving patient outcomes.
The Mechanistic Role of IL-31 in Respiratory Type 2 Inflammatory Diseases
The Role of the IL-31 in the Pathogenesis of Asthma
While initially linked to the pro-inflammatory cascade in allergic diseases, recent evidence reveals that the IL-31/IL-31RA signaling axis plays a complex and multifaceted role in asthma, exhibiting both pro- and anti-inflammatory functions. Furthermore, its receptor, IL-31RA, demonstrates critical functions independent of its ligand, IL-31 (Table 1).20–23
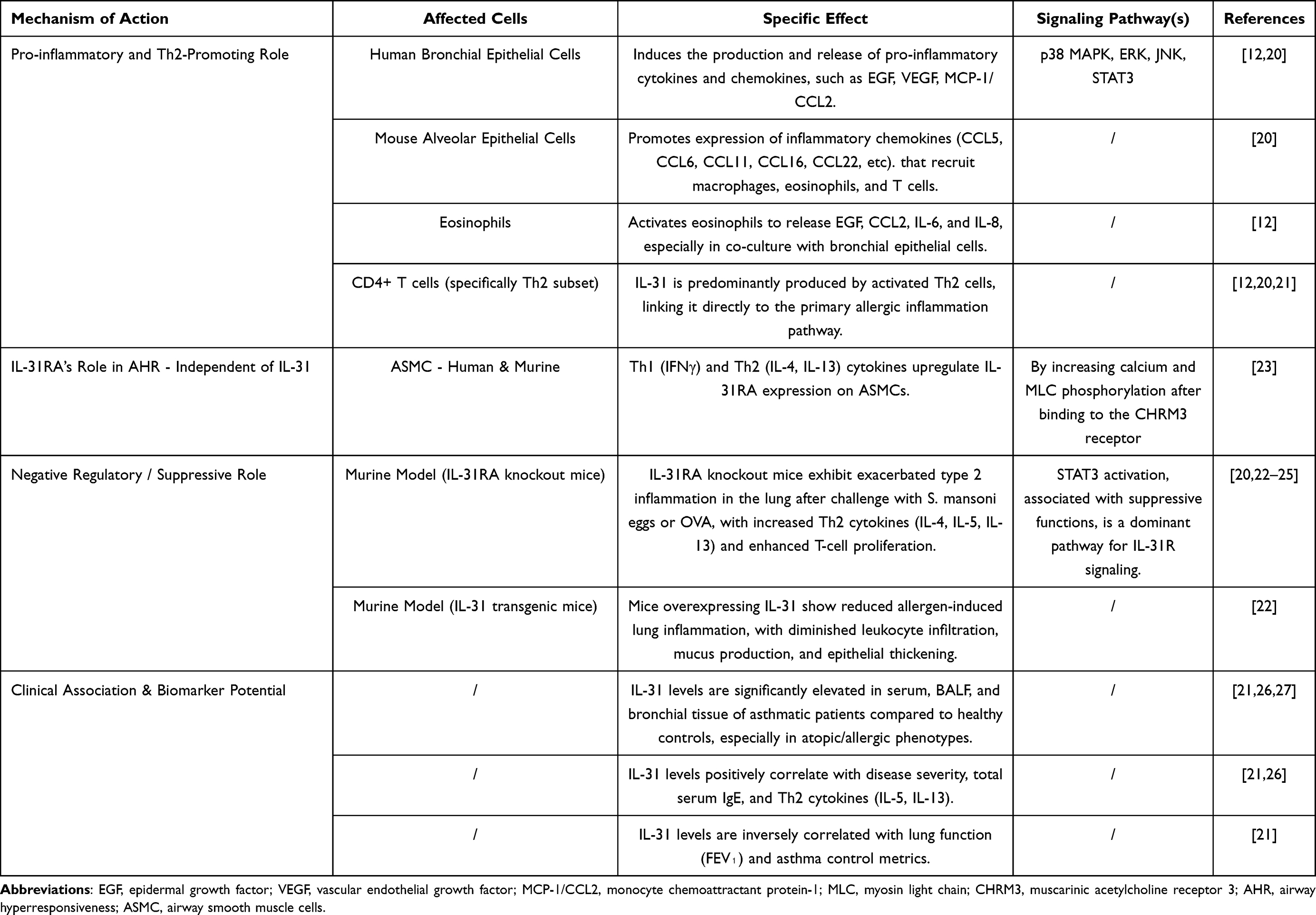 |
Table 1 The Role of IL-31 in Asthma |
The Pro-Inflammatory Role of IL-31: An Early Driver and Amplifier of Inflammation
IL-31 acts as a pro-inflammatory mediator, playing a dual role as both a driver and an amplifier of the inflammatory cascade. Its pro-inflammatory action is closely linked to the classic type 2 inflammatory pathway, as its expression levels positively correlate with key Th2 cytokines, including IL-5, IL-13, and thymic stromal lymphopoietin (TSLP), as well as total serum IgE.19,21,26,27 Moreover, IL-4 can induce IL-31 production from T cells, suggesting a positive feedback loop within the Th2 network.28 Mechanistically, following an allergen challenge, both IL-31 and its receptor IL-31RA are upregulated in the lungs of asthmatic mouse models.20 IL-31 then directly stimulates alveolar and bronchial epithelial cells to produce a wide array of chemokines, such as CCL5, CCL11 (Eotaxin-1), and CCL22. These chemokines are crucial for recruiting inflammatory cells—including eosinophils, T cells, and macrophages—to the airways, thereby initiating and amplifying local inflammation.20 Furthermore, a novel mechanism involving an IL-31/TRPV1 (Transient Receptor Potential Vanilloid 1) pathway has been identified in asthma exacerbated by environmental chemical exposure. Dermal exposure to diisononyl phthalate (DINP), for instance, elevates IL-31, which activates the TRPV1 pathway, leading to increased IL-4 secretion and a subsequent Th2 immune imbalance that aggravates asthma symptoms.29
The Anti-Inflammatory Role of IL-31: A Late-Phase “Brake” on Inflammation
Paradoxically, the IL-31/IL-31RA axis also functions as a negative regulator that limits the magnitude of type 2 inflammation.20,24 This is strongly supported by studies on IL-31 transgenic (IL-31tg) mice, which, despite constitutively high levels of IL-31, exhibit significantly reduced allergen-induced lung inflammation, including diminished leukocyte and eosinophil infiltration and less mucus production.22 Conversely, this regulatory function is further substantiated by studies on IL-31RA knockout (IL-31RA−/−) mice. These mice consistently develop exacerbated type 2 inflammation in response to allergens or parasitic infections, characterized by more severe inflammation and higher Th2 cytokine production.22 One proposed mechanism is that high concentrations of IL-31 compete with Oncostatin M (OSM) for their shared receptor subunit, OSMRβ.25 This competition could suppress OSM-mediated pro-inflammatory effects, which are driven by factors like IL-6 and VEGF, thereby exerting an overall anti-inflammatory effect.22
IL-31RA Regulation of Airway Hyperresponsiveness
Recent studies have identified a novel, ligand-independent role for IL-31RA in directly promoting airway hyperresponsiveness (AHR), a function distinct from its involvement in airway inflammation.23 In allergic asthma models, the expression of IL-31RA—but not its ligand IL-31—is significantly upregulated in the lungs. This upregulation is driven by key Th1 (IFN-γ) and Th2 (IL-4, IL-13) cytokines in airway smooth muscle cells (ASMCs).23 Crucially, the genetic loss of IL-31RA specifically attenuates AHR without affecting airway inflammation or mucus production, functionally decoupling the mechanism of AHR from the inflammatory response in these models. The underlying mechanism is a ligand-independent function where IL-31RA acts as a positive regulator of the muscarinic acetylcholine receptor 3 (CHRM3) on ASMCs. IL-31RA physically interacts with CHRM3, enhancing its protein levels and thereby augmenting agonist-induced calcium signaling and myosin light chain phosphorylation, which directly promotes smooth muscle contraction and AHR.23
The Role of IL-31 in Allergic Rhinitis (AR)
In recent years, growing evidence shows that Interleukin-31 (IL-31) plays a key role in the pathogenesis of allergic rhinitis (AR). IL-31 is an important clinical biomarker and is also involved in driving key symptoms.17,19,30
IL-31 Is a Strong Biomarker for AR Activity and Severity
Many clinical studies confirm that both adult and pediatric patients with AR have higher systemic and local levels of IL-31. Compared to healthy controls, IL-31 levels are significantly increased in the serum, nasal lavage fluid, and nasal mucosal tissue of AR patients.19,30–32 (Table 2). Notably, the level of IL-31 is closely related to disease severity. In pediatric patients, IL-31 concentration is positively correlated with total IgE levels and eosinophil counts, especially in children who also have asthma.30 After allergen challenge, IL-31 is released mainly during the late-phase reaction, and its concentration correlates with the severity of nasal symptoms and the level of the key Th2 cytokine IL-13.17 This link to severity is further solidified by findings that the amount of pollen-induced IL-31 production by PBMCs is selectively associated with a poorer quality of life (QOL) in AR patients, a correlation not observed with IL-5 or IL-13, suggesting a unique role for IL-31 in the disease’s impact.33 These findings establish IL-31 as a reliable biomarker for disease activity and Th2 inflammation in AR.
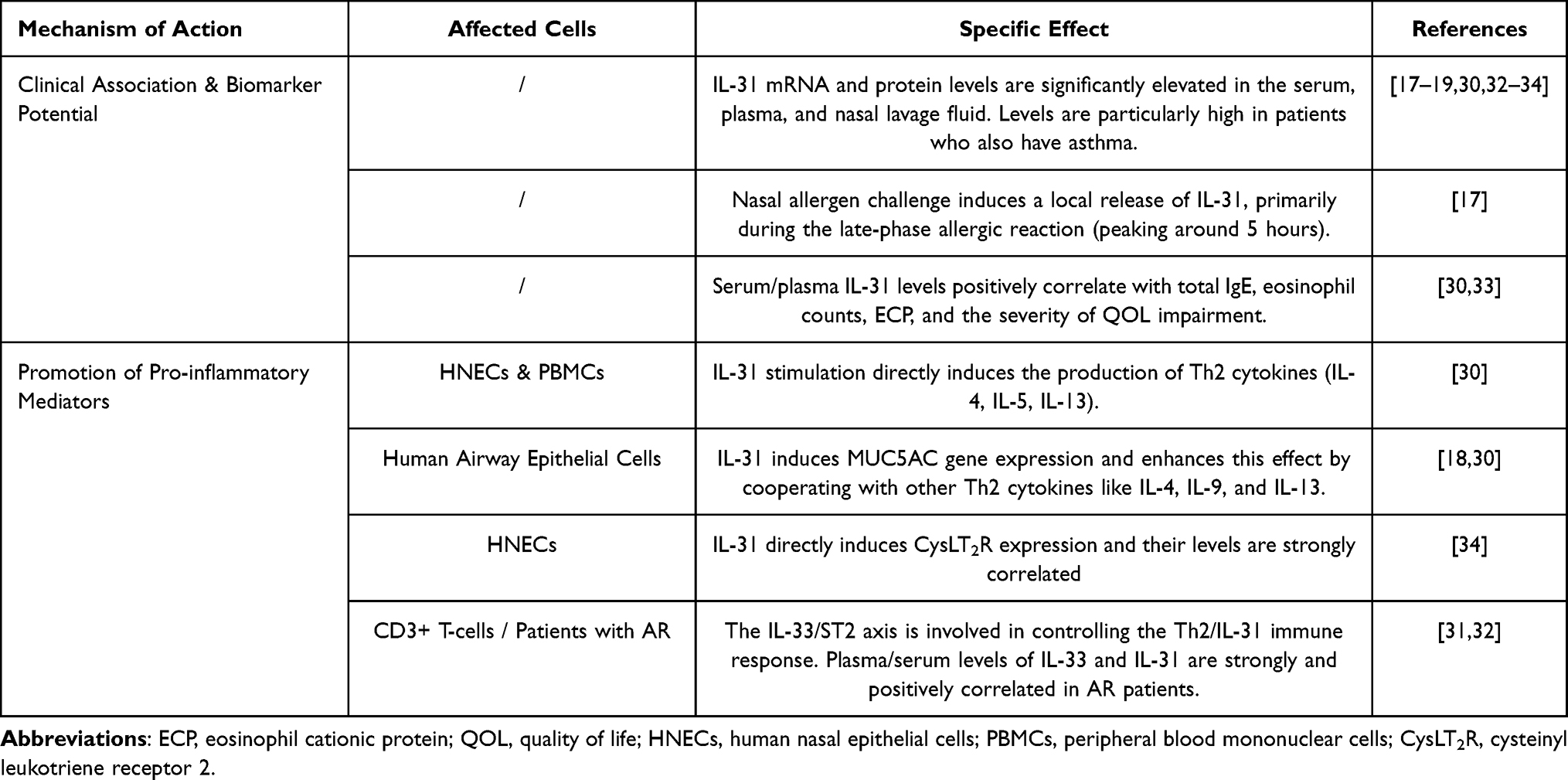 |
Table 2 The Role of IL-31 in AR |
Driving Nasal Itching and Mucus Secretion
The role of IL-31 in allergic rhinitis (AR) is especially clear in how it directly causes two main symptoms: nasal itching and mucus secretion. For nasal itching, recent research has identified the IL-31/CysLT2R axis as a key pathway. In the nasal epithelial cells of AR patients, the expression of both IL-31 and cysteinyl leukotriene receptor 2 (CysLT2R) is significantly increased. Most importantly, the levels of IL-31 and CysLT2R are specifically associated with the severity of nasal itching, but not with other symptoms like congestion or a runny nose.34 In parallel, IL-31 is an important driver of excess mucus production. In the nasal mucosa of AR patients, the expression of IL-31 and its receptor (IL-31RA) is increased, mainly in the submucosal glands. Functional studies show that IL-31 directly causes human airway epithelial cell lines to express the MUC5AC gene, a major respiratory mucin. Furthermore, IL-31 works together with other Th2 cytokines like IL-4 and IL-13 to increase MUC5AC gene expression, confirming its role as a direct cause of mucus hypersecretion in AR.18
The Role of IL-31 in Chronic Rhinosinusitis with Nasal Polyps (CRSwNP)
IL-31 is also implicated in the pathogenesis of chronic rhinosinusitis with nasal polyps (CRSwNP). In nasal polyp tissues, IL-31 expression is significantly elevated and correlates with clinical disease severity.35 Mechanistically, IL-31 appears to drive the local Th2 inflammatory environment. Its levels are positively correlated with key Th2 cytokines, and it may amplify the Th2 response and eosinophilic inflammation by directly stimulating epithelial cells.35 However, as research on its role in CRSwNP is still emerging, the comprehensive mechanisms of IL-31 in this disease warrant further investigation.
An Unbiased Summary: The Dual Role and Future Perspectives
While IL-31 is primarily recognized for its pro-inflammatory functions, an unbiased perspective must also acknowledge its potential regulatory roles. For instance, studies in chronic asthma mouse models have shown that IL-31 can reduce CD4⁺ T-cell proliferation and type 2 cytokine production, leading to attenuated eosinophilia and epithelial thickening.20 This paradoxical effect suggests that IL-31 may act as a negative feedback regulator in chronic inflammation.33
In summary, IL-31 is a critical mediator in type 2 respiratory diseases. It drives inflammatory cascades by activating Th2 cells, eosinophils, and macrophages, which in turn release type 2 cytokines and chemokines that amplify allergic inflammation (Figure 1). It contributes to distinct features of asthma, such as airway hyperresponsiveness and inflammation, and to allergic rhinitis manifestations such as pruritus. Yet, its complex and sometimes contradictory functions underscore the need for deeper investigation. These characteristics make IL-31 not only an attractive therapeutic target but also a molecule whose full biological significance remains to be elucidated.
Clinical Significance of IL-31
With advancements in immunology and molecular biology, biomarkers are increasingly playing a critical role in disease diagnosis, prognosis assessment, and treatment monitoring. Recent studies have demonstrated a strong correlation between IL-31 expression levels and the onset, progression, activity, and severity of clinical symptoms in respiratory type 2 inflammatory diseases. These findings provide both theoretical foundation and practical prospects for the use of IL-31 as a biomarker in clinical practice.
IL-31 as a Biomarker for Disease Diagnosis
In airway allergic diseases such as asthma and AR, IL-31 levels are significantly elevated in the serum, nasal secretions, and bronchoalveolar lavage fluid (BALF) of affected individuals, with its concentration positively correlated with disease severity.17,19,21 Therefore, IL-31 detection may serve as a valuable tool for screening and auxiliary diagnosis of airway allergic diseases. Furthermore, combining IL-31 detection with other established biomarkers, such as IL-5, IL-13, could enhance the sensitivity and accuracy of diagnosis.
IL-31 as a Biomarker for Monitoring Disease Activity
Given the strong correlation between IL-31 levels and the disease activity in respiratory type 2 inflammatory diseases, IL-31 holds promise as a potential biomarker for monitoring disease progression. A study by Lai et al demonstrated that serum IL-31 levels in asthma patients are positively correlated with allergen-specific immunoglobulin E (IgE) and negatively correlated with forced expiratory volume in one second (FEV₁).21 Since acute asthma exacerbations are associated with high mortality rates and are largely mediated by IgE-driven mechanisms, early detection of IL-31 levels may facilitate the identification of disease worsening, enabling timely intervention and treatment adjustments. Therefore, for chronic and recurrent diseases, routine monitoring of IL-31 concentrations could support dynamic disease assessment and inform personalized treatment strategies.
IL-31 as a Biomarker for Treatment Response Assessment
With the increasing adoption of biologics and targeted therapies, evaluation of treatment efficacy has become a critical aspect of clinical management. Given that IL-31 expression levels are closely associated with the severity and clinical manifestations in airway diseases, fluctuations in IL-31 concentrations may provide a reliable indicator of therapeutic response. For instance, studies have shown an inverse correlation between IL-31 levels and asthma control,19,21 suggesting that serum IL-31 measurements could serve as a dynamic tool for assessing disease management. This is particularly valuable in cases where patients exhibit suboptimal disease control despite presenting with mild or ambiguous clinical symptoms, thereby supporting precision medicine approaches. In addition, our study demonstrates that nasal corticosteroid treatment significantly reduces the expression of IL-31 and CysLT2R in the nasal epithelium of patients with AR, accompanied by a marked alleviation of itching. These findings suggest that IL-31 expression levels are closely correlated with the therapeutic response to corticosteroid treatment and may serve as a potential biomarker for predicting treatment efficacy.34
Potential and Challenges of IL-31 as a Therapeutic Target
With increasing insights into the role of IL-31 in immune-mediated diseases, targeting IL-31 and its receptors has emerged as a novel therapeutic strategy. IL-31 not only modulates inflammatory responses but is also closely implicated in pathological processes such as itching in allergic reactions, dyspnea, and airway remodeling. Consequently, IL-31 is considered a promising therapeutic target in respiratory type 2 inflammatory diseases. Currently, IL-31-targeted therapies can be broadly categorized into two types: (1) Direct inhibition of IL-31using monoclonal antibodies (eg, BMS-981164); (2) Inhibition of the IL-31 receptor (IL-31RA or OSMR), thereby indirectly suppressing IL-31’s biological activity, as seen with Nemolizumab (CIM331).36 Nemolizumab exerts its effects of IL-31 by binding to IL-31RA, blocking the binding of IL-31 to its receptors, thereby mitigating its pro-inflammatory effects.15 It has already been approved for the treatment of atopic dermatitis and has demonstrated significant therapeutic efficacy.37 However, clinical trials investigating IL-31-targeted therapies for respiratory type 2 inflammatory diseases have not yet been conducted, likely due to the incomplete understanding of IL-31’s specific role in these conditions. IL-31 engages in complex interactions with immune cells, epithelial cells, and other cytokines, warranting further investigation to clarify its role in chronic respiratory inflammation. This complexity poses challenges for the development of IL-31–targeted monotherapy. Future studies should define the precise contribution of IL-31 to type 2 airway inflammation and evaluate its safety and therapeutic potential.
Sensory nerve hyperresponsiveness has been recognized as a critical mechanism underlying persistent cough in patients with asthma and CRSwNP,38,39 which cannot be fully relieved by inhaled corticosteroids or anti–IL-5/IL-4Rα therapies.40,41 A recent study in a mouse model of allergic asthma offers a potential new approach. In this study, pharmacologic blockade of the IL-31/TRPV1 pathway effectively attenuated airway hyperresponsiveness, inflammation, and remodeling. These findings provide a strong rationale for investigating IL-31 antagonists as a novel therapy for allergic asthma.29 Although cytokines such as IL-4 and IL-5 play more established roles in the airways, IL-31 provides an additional pathway that may be linked to dysfunction of the neuroepithelial unit, and targeting this axis could yield more pronounced improvements in clinical symptoms.
However, to translate these promising neuroimmune functions of IL-31 into effective therapies, several mechanistic and translational questions must be addressed. Key mechanistic issues include: (1) Dissecting downstream pathways – clarifying how JAK/STAT and MAPK signaling mediate distinct functions in epithelial cells versus sensory neurons to enable cell type–specific interventions; (2) Defining the dual role of IL-31 – identifying molecular switches that shift its function from a pro-inflammatory mediator in acute phases to a potential negative regulator in chronic conditions; and (3) Exploring neuroimmune interactions – elucidating the role of the IL-31/CysLT2R axis in nasal pruritus and evaluating its potential as a target for topical therapies in allergic rhinitis.
From a translational and clinical perspective, future studies should focus on: (1) Clinical trial efficacy-assessing the safety and effectiveness of IL-31 receptor antagonists (eg, Nemolizumab) in patients with severe asthma or allergic rhinitis, particularly those with Cough Hypersensitivity Syndrome; and (2) Biomarker development – determining whether IL-31 or its receptor levels in serum or tissue can predict which patients are most likely to benefit from IL-31-targeted interventions. Collectively, these efforts will clarify the precise contribution of IL-31 to type 2 airway inflammation and its therapeutic potential.
Conclusion
In conclusion, Interleukin-31 is emerging as a pivotal and multifaceted mediator in the complex network of respiratory type 2 inflammatory diseases, extending far beyond its well-established role in atopic dermatitis. Its unique functions in orchestrating immune activation, driving airway remodeling, and modulating neuroimmune pathways—particularly its link to pruritus—highlight its significance as both a promising biomarker and a compelling therapeutic target for symptoms often refractory to conventional treatments. However, critical questions remain. A key challenge is to elucidate the molecular switch that governs IL-31’s paradoxical dual function, shifting from a pro-inflammatory role in acute phases to a regulatory role in chronic conditions. Addressing these mechanistic questions and advancing targeted therapies into clinical trials will be essential to translate our understanding of IL-31 into clinical benefit.
Acknowledgments
This work was supported by the National Key R&D Program of China (2022YFC2504100), the National Natural Science Foundation of China (Nos. 82271139, 82371115, and 82471139), the Program for Changjiang Scholars and Innovative Research Team (IRT13082), the High Level Public Health Technical Talent Training Plan (Lingjunrencai-02-09), the Beijing Municipal Public Welfare Development and Reform Pilot Project for Medical Research Institutes, Beijing Municipal Health Commission (JYY2023-1) and the CAMS Innovation Fund for Medical Sciences (2019-I2M-5-022).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no competing interests related to this work, apart from the funding sources disclosed.
References
1. Kolkhir P, Akdis CA, Akdis M, et al. Type 2 chronic inflammatory diseases: targets, therapies and unmet needs. Nat Rev Drug Discov. 2023;22(9):743–767. doi:10.1038/s41573-023-00750-1
2. McCormick JP, Lee JT. Insights into the implications of coexisting type 2 inflammatory diseases. J Inflamm Res. 2021;14:4259–4266. doi:10.2147/jir.S311640
3. Molofsky AB, Locksley RM. The ins and outs of innate and adaptive type 2 immunity. Immunity. 2023;56(4):704–722. doi:10.1016/j.immuni.2023.03.014
4. Dillon SR, Sprecher C, Hammond A, et al. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat Immunol. 2004;5(7):752–760. doi:10.1038/ni1084
5. Kasraie S, Niebuhr M, Baumert K, Werfel T. Functional effects of interleukin 31 in human primary keratinocytes. Allergy. 2011;66(7):845–852. doi:10.1111/j.1398-9995.2011.02545.x
6. Datsi A, Steinhoff M, Ahmad F, Alam M, Buddenkotte J. Interleukin-31: the “itchy” cytokine in inflammation and therapy. Allergy. 2021;76(10):2982–2997. doi:10.1111/all.14791
7. Rüdrich U, Gehring M, Papakonstantinou E, et al. Eosinophils are a Major Source of Interleukin-31 in Bullous Pemphigoid. Acta Derm Venereol. 2018;98(8):766–771. doi:10.2340/00015555-2951
8. Bağci IS, Ruzicka T. IL-31: a new key player in dermatology and beyond. J Allergy Clin Immunol. 2018;141(3):858–866. doi:10.1016/j.jaci.2017.10.045
9. Gibbs BF, Patsinakidis N, Raap U. Role of the pruritic cytokine IL-31 in autoimmune skin diseases. Front Immunol. 2019;10:1383. doi:10.3389/fimmu.2019.01383
10. Sonkoly E, Muller A, Lauerma AI, et al. IL-31: a new link between T cells and pruritus in atopic skin inflammation. J Allergy Clin Immunol. 2006;117(2):411–417. doi:10.1016/j.jaci.2005.10.033
11. Nakashima C, Otsuka A, Kabashima K. Interleukin-31 and interleukin-31 receptor: new therapeutic targets for atopic dermatitis. Exp Dermatol. 2018;27(4):327–331. doi:10.1111/exd.13533
12. Ip WK, Wong CK, Li ML, Li PW, Cheung PF, Lam CW. Interleukin-31 induces cytokine and chemokine production from human bronchial epithelial cells through activation of mitogen-activated protein kinase signalling pathways: implications for the allergic response. Immunology. 2007;122(4):532–541. doi:10.1111/j.1365-2567.2007.02668.x
13. Nilforoushzadeh MA, Heidari N, Ghane Y, et al. A systematic review of interleukin-31 inhibitors in the treatment of prurigo nodularis. Inflammopharmacology. 2024;32(2):991–1003. doi:10.1007/s10787-024-01436-9
14. Ständer S, Yosipovitch G, Lacour JP, et al. Nemolizumab efficacy in prurigo nodularis: onset of action on itch and sleep disturbances. J Eur Acad Dermatol Venereol. 2022;36(10):1820–1825. doi:10.1111/jdv.18377
15. Kwatra SG, Yosipovitch G, Legat FJ, et al. Phase 3 trial of nemolizumab in patients with prurigo nodularis. N Engl J Med. 2023;389(17):1579–1589. doi:10.1056/NEJMoa2301333
16. Kabashima K, Matsumura T, Komazaki H, Kawashima M. Trial of nemolizumab and topical agents for atopic dermatitis with pruritus. N Engl J Med. 2020;383(2):141–150. doi:10.1056/NEJMoa1917006
17. Baumann R, Rabaszowski M, Stenin I, et al. The release of IL-31 and IL-13 after nasal allergen challenge and their relation to nasal symptoms. Clin Transl Allergy. 2012;2(1):13. doi:10.1186/2045-7022-2-13
18. Shah SA, Ishinaga H, Hou B, Okano M, Takeuchi K. Effects of interleukin-31 on MUC5AC gene expression in nasal allergic inflammation. Pharmacology. 2013;91(3–4):158–164. doi:10.1159/000346609
19. Chai R, Liu B, Qi F. IL-31, IL-33, and TSLP expression and relation to severity of asthma and rhinitis in Chinese allergic patients. Int J Clin Experiment Pathol. 2017;10:1774–1782.
20. Huang J, Yue H, Jiang T, et al. IL-31 plays dual roles in lung inflammation in an OVA-induced murine asthma model. Biol Open. 2019;8(1):bio036244. doi:10.1242/bio.036244
21. Lai T, Wu D, Li W, et al. Interleukin-31 expression and relation to disease severity in human asthma. Sci Rep. 2016;6:22835. doi:10.1038/srep22835
22. Neuper T, Neureiter D, Sarajlic M, et al. IL-31 transgenic mice show reduced allergen-induced lung inflammation. Eur J Immunol. 2021;51(1):191–196. doi:10.1002/eji.202048547
23. Akkenepally SV, Yombo DJK, Yerubandi S, et al. Interleukin 31 receptor α promotes smooth muscle cell contraction and airway hyperresponsiveness in asthma. Nat Commun. 2023;14(1):8207. doi:10.1038/s41467-023-44040-1
24. Perrigoue JG, Li J, Zaph C, et al. IL-31-IL-31R interactions negatively regulate type 2 inflammation in the lung. J Exp Med. 2007;204(3):481–487. doi:10.1084/jem.20061791
25. Bilsborough J, Mudri S, Chadwick E, Harder B, Dillon SR. IL-31 receptor (IL-31RA) knockout mice exhibit elevated responsiveness to oncostatin M. J Immunol. 2010;185(10):6023–6030. doi:10.4049/jimmunol.0902769
26. Moaaz M, Abo El-Nazar S, Abd El-Rahman M, Soliman E. Stem cell factor and interleukin-31 expression: association with IgE among egyptian patients with atopic and nonatopic bronchial asthma. Immunol Invest. 2016;45(2):87–106. doi:10.3109/08820139.2015.1089890
27. Ulambayar B, Lee SH, Yang EM, Ye YM, Park HS. Association between epithelial cytokines and clinical phenotypes of elderly asthma. Allergy Asthma Immunol Res. 2019;11(1):79–89. doi:10.4168/aair.2019.11.1.79
28. Edukulla R, Singh B, Jegga AG, Sontake V, Dillon SR, Madala SK. Th2 Cytokines Augment IL-31/IL-31RA Interactions via STAT6-dependent IL-31RA Expression. J Biol Chem. 2015;290(21):13510–13520. doi:10.1074/jbc.M114.622126
29. Peng Q, Wu Y, Li Y, et al. The IL-31/TRPV1 pathway mediates allergic asthma exacerbated by DINP dermal exposure in OVA-sensitized Balb/c mice. Sci Total Environ. 2024;912:169613. doi:10.1016/j.scitotenv.2023.169613
30. Liu W, Luo R, Chen Y, et al. Interleukin-31 promotes helper T cell type-2 inflammation in children with allergic rhinitis. Pediatr Res. 2015;77(1–1):20–28. doi:10.1038/pr.2014.151
31. Vocca L, Di Sano C, Uasuf CG, et al. IL-33/ST2 axis controls Th2/IL-31 and Th17 immune response in allergic airway diseases. Immunobiology. 2015;220(8):954–963. doi:10.1016/j.imbio.2015.02.005
32. Qiao Y, Chen J. Serum levels of IL-31, IL-33 and ST2 in allergic rhinitis of children in China. Cell Mol Biol. 2018;64(12):52–55. doi:10.14715/cmb/2018.64.12.11
33. Okano M, Fujiwara T, Higaki T, et al. Characterization of pollen antigen-induced IL-31 production by PBMCs in patients with allergic rhinitis. J Allergy Clin Immunol. 2011;127(1):277–279.e1–11. doi:10.1016/j.jaci.2010.09.029
34. Du K, Zhang M, Yao S, Luo T, Yu H, Lou H. The IL-31/CysLT2R axis is associated with itching in patients with allergic rhinitis. Int Forum Allergy Rhinol. 2024;14(6):1070–1078. doi:10.1002/alr.23311
35. Ouyang H, Cheng J, Zheng Y, Du J. Role of IL-31 in regulation of Th2 cytokine levels in patients with nasal polyps. Eur Arch Otorhinolaryngol. 2014;271(10):2703–2709. doi:10.1007/s00405-014-2913-x
36. Tan HT, Sugita K, Akdis CA. Novel biologicals for the treatment of allergic diseases and asthma. Curr Allergy Asthma Rep. 2016;16(10):70. doi:10.1007/s11882-016-0650-5
37. Ständer S, Yosipovitch G, Legat FJ, et al. Efficacy and safety of nemolizumab in patients with moderate to severe prurigo nodularis: the OLYMPIA 1 randomized clinical phase 3 trial. JAMA Dermatol. 2024. doi:10.1001/jamadermatol.2024.4796
38. Song WJ, Chang YS. Cough hypersensitivity as a neuro-immune interaction. Clin Transl Allergy. 2015;5:24. doi:10.1186/s13601-015-0069-4
39. Mazzone SB, McGarvey L. Mechanisms and rationale for targeted therapies in refractory and unexplained chronic cough. Clin Pharmacol Ther. 2021;109(3):619–636. doi:10.1002/cpt.2003
40. Ito K, Kanemitsu Y, Fukumitsu K, et al. Targeting the interleukin-5 pathway improves cough hypersensitivity in patients with severe uncontrolled asthma. Ann Allergy Asthma Immunol. 2023;131(2):203–208.e1. doi:10.1016/j.anai.2023.04.022
41. Lombardi C, Berti A, Menzella F, Cottini M. Dupilumab reduces patient-reported cough and improves quality of life in patients with severe eosinophilic asthma with or without chronic rhinosinusitis with nasal polyps: a real-life prospective study. Drugs Context. 2025;14:2025. doi:10.7573/dic.2025-6-3
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.