Back to Journals » Journal of Inflammation Research » Volume 18
Interleukin-27 and Interleukin-2 Cooperatively Regulate CD4⁺ T Cell Subsets and Immune Imbalance in Coronary Artery Disease
Authors Cai Y ![]() , Tang H
, Tang H ![]() , Tang W, Tang W, Xu W, Wang Y, Ding Y, Yu J, Pan C, Li Z, Peng Y, Zhu R, Yu K, Zeng Q, Zhong Y
, Tang W, Tang W, Xu W, Wang Y, Ding Y, Yu J, Pan C, Li Z, Peng Y, Zhu R, Yu K, Zeng Q, Zhong Y ![]()
Received 8 June 2025
Accepted for publication 9 November 2025
Published 10 December 2025 Volume 2025:18 Pages 17253—17269
DOI https://doi.org/10.2147/JIR.S545568
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Qing Lin
Yifan Cai,1– 3,* Hongxia Tang,4,* Wenwen Tang,5,* Wenjuan Tang,6 Wenbin Xu,1– 3 Yue Wang,1– 3 Yan Ding,1– 3 Jian Yu,1– 3 Chengliang Pan,1– 3 Zhiyang Li,1– 3 Yudong Peng,1– 3 Ruirui Zhu,1– 3 Kunwu Yu,1– 3 Qiutang Zeng,1– 3 Yucheng Zhong7
1Department of Cardiology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China; 2Hubei Key Laboratory of Biological Targeted Therapy, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China; 3Hubei Provincial Engineering Research Center of Immunological Diagnosis and Therapy for Cardiovascular Diseases, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China; 4Department of Rheumatology and Immunology, Wuhan Children’s Hospital, Tongji Medical College, Huazhong University of Science & Technology, Wuhan, Hubei, People’s Republic of China; 5Cardiovascular Department, Wuhan No.9 Hospital, Wuhan, Hubei, People’s Republic of China; 6Radiology Department, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China; 7Department of Cardiovascular Surgery, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yucheng Zhong, Department of Cardiovascular Surgery, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, 1277 Jiefang Avenue, Jianghan District, Wuhan, Hubei, 430022, People’s Republic of China, Tel/Fax +86 158 7237 4991, Email [email protected]
Background: Coronary artery disease (CAD) is an immune-mediated disorder driven by dysregulated T cell responses. Interleukin-27 (IL-27) has immunoregulatory properties, but its role in CAD remains unclear. This study is the first to investigate the effects of IL-27 on CD4⁺LAP⁺ T cells in CAD and to explore its interaction with interleukin-2 (IL-2) in modulating immune imbalance.
Methods: CAD severity was quantified by the Gensini score. Plasma IL-27 and oxidized low-density lipoprotein (ox-LDL) were measured by ELISA. Flow cytometry assessed CD4⁺ T cell subsets, while qRT-PCR and Western blot evaluated lineage-specific transcription factors.
Results: IL-27 levels were elevated in acute coronary syndrome and correlated with ox-LDL and Gensini scores. Patients with severe CAD showed a Th1/Th17-dominant profile and reductions in Th2, CD4⁺LAP⁺, and Tregs. In vitro, IL-27 promoted Th1 differentiation via T-bet/IFN-γ upregulation and suppressed Th2, Th17, and regulatory subsets, counteracting IL-2–induced expansion of Tregs and CD4⁺LAP⁺ cells. These effects were dose dependent and favored pro-inflammatory responses.
Conclusion: IL-27 drives immune imbalance in CAD by reinforcing Th1 polarization and antagonizing IL-2–mediated regulation. Beyond mechanistic insights, these findings identify IL-27 as a potential biomarker for disease severity and a candidate therapeutic target in CAD.
Keywords: IL-27, coronary artery disease, Th1/Th2, Th17/CD4+LAP+ T cells
Introduction
Coronary artery disease (CAD) is the most important cause of death and disability worldwide, causing 8.9 million deaths and 164 million years of healthy life lost (disability-adjusted life years, DALYs), in 2015, with lower- and middle-income countries have a higher burden.1 Clinically, CAD is categorized as chronic coronary syndromes (CCS, which encompass conditions previously referred to as stable angina pectoris, SAP) and acute coronary syndromes (ACS), which include unstable angina (UA) and acute myocardial infarction (AMI). Numerous reports increasing evidence indicate atherosclerosis is a chronic inflammatory disease, and inflammatory cells and the cytokine milieu are involved in plaque instability.2,3 CD4 T lymphocytes (T lymphocytes) are the largest subset of T lymphocyte populations and differentiate into Th1, Th2, Th17, and regulatory T cells (Tregs) with distinct roles in the disease progression.4–7 Recently, CD4+LAP+ T cells have been described as a discrete regulatory population, but their role, in CAD is poorly characterized.
The interleukin (IL)-12 family of cytokines, (including IL-12, 23, 27, 35) is becoming appreciated as critical modulators of cardiovascular disease.8 IL-27, identified universally in 2002, is secreted by antigen presenting cells and and signals through a heterodimeric receptor composed of IL-27Rα and gp1309.9 Laboratory studies showed a protective effect of IL-27 in murine atherosclerosis, as IL-27 receptor-deficient mice develop more severe lesions, while IL-27 administration alleviates disease.10,11 Although human studies may be reflective of opposite trend, plasma IL-27 is increased in UA and AMI, correlates with worsening left ventricular function, and is higher in patients with CAD versus controls.12
In terms of its mechanism, IL-27 has been shown to regulate CD4⁺ T cell differentiation via the activation of the STAT1/STAT3 signaling axis, promoting Th1 polarization through T-bet and IFN-γ while suppressing the Th2- and Th17-associated transcription factors GATA-3 and RORγt.13 In addition, it inhibits the generation of induced Tregs in mice, but not natural Tregs.14 However, despite these observations, our understanding of whether IL-27 regulates CD4⁺CD25⁺Foxp3⁺ Tregs and CD4⁺LAP⁺ T cells in individuals with CAD is lacking. As a consequence, our study intended to assess plasma IL-27 levels and their association with disease severity, as well as the impact of IL-27 and interleukin-2 (IL-2) on CD4⁺ T cell subset differentiation in CAD patients.
Methods
Research Subjects
A total of 95 patients diagnosed with coronary artery disease (CAD) and hospitalized at Wuhan Union Hospital from August 2021 to August 2022 were included in this study. Sample size was estimated based on preliminary data and prior literature, aiming to achieve sufficient statistical power (>80%) to detect differences in inflammatory biomarker levels between CAD subgroups with a two-sided α of 0.05. Based on coronary angiography (CAG) results, laboratory biomarkers, and electrocardiograms, the CAD patients were divided into three groups: SAP (n = 32), UA (n = 33), and AMI (n = 30). AMI was defined according to the 2018 ESC diagnostic criteria without further subdivision into ST-segment elevation and non-ST-segment elevation types.15 Additionally, 30 patients without evidence of CAD, who underwent CAG for evaluation of chest pain or suspected ischemia but were confirmed to have normal coronary arteries, were included as the healthy controls. CAG was performed for clinical indications rather than purely for research purposes. The average age of the healthy controls was 60.34 ± 5.42 years, comprising 19 males and 11 females.
Inclusion criteria: (1) Patients who met the 2019 diagnostic criteria for CAD.16 Diagnostic criteria included the presence of typical symptoms (eg, angina pectoris), objective evidence of myocardial ischemia on electrocardiogram or biomarkers, and/or angiographic evidence of ≥50% luminal stenosis in at least one major coronary artery. All patients underwent coronary angiography for confirmation; (2) All subjects underwent CAG for the first time. (3) For the AMI group, diagnosis established by clinical presentation, electrocardiogram (ECG) changes, and elevated cardiac biomarkers [ST-Elevation Myocardial Infarction (STEMI) or Non-ST-Elevation Myocardial Infarction (NSTEMI)].
Exclusion criteria: (1) Patients with arrhythmia (clinically documented atrial fibrillation, atrial flutter, or sustained ventricular tachyarrhythmias), heart failure (meeting 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure, New York Heart Association [NYHA] class II–IV17), valvular heart disease (moderate to severe stenosis or regurgitation confirmed by echocardiography), congenital heart disease (structural abnormalities diagnosed by imaging), cardiomyopathy (dilated, hypertrophic, or restrictive cardiomyopathy per established diagnostic criteria); (2) Patients with cardiogenic shock, cardiac arrest, or those undergoing staged rather than primary PCI were excluded; (3) Patients with a history of active infection or chronic inflammatory diseases affecting the skeletal system; (4) Patients with chronic liver disease, active hepatitis, acute/chronic biliary diseases, or chronic kidney disease (eGFR <60 mL/min/1.73 m2); (5) Patients with malignant tumors, autoimmune diseases, or parathyroid disorders. Blood samples were collected within 24–48 hours after symptom onset and always prior to coronary angiography to avoid procedure-related changes in inflammatory markers.
The study was approved by the hospital’s ethics committee, and all patients provided signed informed consent.
Gensini Score
All CAD patients were evaluated using the Gensini score based on their CAG results.18 The Gensini score is determined by summing the scores of stenotic segments in the affected coronary vessels. The score for each affected vessel is calculated by multiplying the degree of stenosis by the specific coefficient assigned to the location of the stenosis. The degree of luminal stenosis is assigned the following points: 1%-25%, 26%-50%, 51%-75%, 76%-90%, 91%-99%, and total occlusion receive 1, 2, 4, 8, 16, and 32 points, respectively. For vessels with multiple stenoses, each lesion was evaluated and scored individually based on both the degree of stenosis and its topographic importance. The total Gensini score was then calculated by multiplying the stenosis severity score by the weighting factor of each segment, followed by summation of all individual lesion scores.
In addition to the Gensini score, detailed coronary angiographic characteristics were recorded. These included the presence of multivessel disease, left main involvement, intracoronary thrombosis, plaque morphology (stable or vulnerable), and the severity of luminal narrowing. Severe stenosis was defined as luminal narrowing >70%. The mean number of diseased vessels was also calculated for each patient. Coronary angiographic features were compared among patients with stable angina pectoris (SAP), unstable angina (UA), and acute myocardial infarction (AMI).
Measurement of Ox-LDL and IL-27 Levels
Plasma levels of IL-27 were measured using enzyme-linked immunosorbent assay (ELISA) according to the manufacturer’s protocol (R&D Systems, USA, DY2526). The minimum detectable concentration of IL-27 was 9.5 pg/mL, with intra-assay and inter-assay coefficients of variation <10%. Each sample was analyzed in duplicate. Total cholesterol(TC), low-density lipoprotein cholesterol (LDL-C), and triglyceride(TG) levels were assessed using enzymatic assays and determined with an autoanalyzer (Hitachi 917).
Plasma and Peripheral Blood Mononuclear Cell (PBMC) Preparation
Peripheral blood samples were collected from all study participants. Plasma was isolated and stored at −80°C until further analysis. PBMCs were separated from blood samples using LSM-Lymphocyte Separation Medium (1.0770–1.0800 g/mL, MP Biomedicals, Santa Ana, CA) through density gradient centrifugation.
Cell Cultures
PBMCs were resuspended in 1 mL complete RPMI 1640 medium (containing 10% fetal bovine serum(FBS) and 2 mmol/L glutamine/penicillin/streptomycin). Cells were then deposited into 2-mL wells in 24-well plates and stimulated with or without 50 ng/mL recombinant human IL-27 (rhIL-27, 2526-IL) and 100 U/mL rhIL-2 (R&D Systems, Minneapolis, MN, BT-002). The cultures were incubated at 37°C in a 5% CO2 atmosphere for 48 hours. After incubation, cell culture supernatants were stored at −80°C, and cells were harvested for subsequent analyses.
CD4+ T Cell Stimulation with Recombinant IL-27
PBMCs were incubated with anti-CD4-FITC (BD Pharmingen, USA) for 30 min at 4°C. Following surface staining, CD4+ T cells were sorted using a FACSAria™ cell sorting system (BD Biosciences, MAGH102), achieving a purity >95%. CD4+ T cells obtained from the healthy controls were cultured with different concentrations of recombinant human IL-27 (rhIL-27) at concentrations of 0, 50, 100, and 200 ng/mL. The cells were maintained in RPMI-1640 medium (Gibco, Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum, 25 mM HEPES, 2 mM glutamine, 0.1 mM non-essential amino acids, 100 U/mL penicillin, and 100 U/mL streptomycin. T cells were stimulated with plate-bound anti-CD3 (2 μg/mL; Abclonal, A25761) and soluble anti-CD28 (2 μg/mL; Abclonal, A27700) at 37°C in a 5% CO2 atmosphere for 72 hours.
Flow Cytometry Analysis of Th1, Th2, Th17, and Treg Cells
After 48 hours of incubation with or without IL-27/IL-2, PBMCs were stimulated with phorbol 12-myristate 13-acetate (20 ng/mL, MCE, HY-18739), ionomycin (1 μg/mL, MCE, HY-13434), and monensin (2 μg/mL, MCE, HY-N0150) at 37°C in a 5% CO2 atmosphere for 4 hours. Cells were subsequently harvested and incubated with FITC-conjugated anti-CD4 (eBioscience, CA, USA, 11–0049-42). After fixation and permeabilization, cells were stained with PE-conjugated anti-IFN-γ (eBioscience, CA, USA, 12–7319-42), PE-conjugated anti-IL-4 (eBioscience, CA, USA, 12–7049-42), and PE-conjugated anti-IL-17 (eBioscience, CA, USA, 12–7178-42).
In another set of experiments, cells from each group were incubated with FITC-conjugated anti-CD4 (eBioscience, CA, USA, 11–0049-42), APC-conjugated anti-CD25 (eBioscience, CA, USA, 17–0289-42), and PE cy7-conjugated anti-LAP (eBioscience, CA, USA, 25–9829-42). Following fixation and permeabilization, PE-conjugated anti-Foxp3 (eBioscience, CA, USA, 12–4776-42) staining was performed in accordance with the manufacturer’s instructions. Flow cytometry was performed using a FACS Calibur system (BD Biosciences).
Real-Time PCR Analysis
Total RNA from PBMCs was isolated using Trizol reagent (TaKaRa Biotechnology, Dalian, China). Reverse transcription was performed with One Step SYBR Green Mix (TaKaRa), and real-time PCR was conducted on an ABI Prism 7900 system (Applied Biosystems, Foster City, CA). Primer sequences for the target genes are listed in Table 1. PCR reactions included 40 cycles of denaturation at 94°C for 10 minutes, followed by 30 seconds at 94°C, 30 seconds at 57°C, and 30 seconds at 72°C. After PCR, melting curves were generated to confirm amplification specificity.
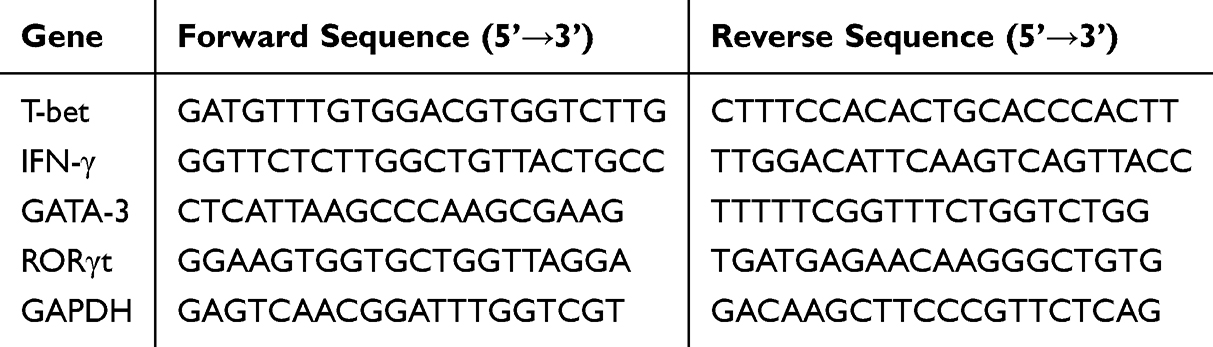 |
Table 1 Primer Sequences for Real-Time Polymerase Chain Reaction |
Western Blotting
Proteins were extracted from splenic lymphocytes using a lysing buffer and incubated on ice for 30 minutes. Lysates were centrifuged at 12,000 g for 15 minutes at 4°C. Protein concentrations in the supernatants were determined using a BCA Protein Assay Kit (Beyotime Institute of Biotechnology, P0009). Equal amounts (20 μg) of protein were separated by SDS-PAGE and transferred onto polyvinylidene fluoride membranes. Membranes were blocked at room temperature for 1 hour and incubated overnight at 4°C with primary antibodies against T-bet (ab275959, 1:1000), IFN-γ (ab267369, 1:1000), RORγt (ab113434, 0.2 μg/mL), GATA3 (ab199428, 1:1000), and GAPDH (ab8245, 1:1000). Membranes were subsequently incubated with HRP-conjugated secondary antibodies (proteintech, SA00001-1, SA00001-2) for 30 minutes at room temperature. Blots were visualized using ECL detection reagents (Bio-Rad Laboratories, Hercules, CA, 1705061).
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 8.0 software (GraphPad Software Inc., San Diego, CA). Data are expressed as mean ± standard error. The t-test was used for comparisons between two groups with normally distributed data, while Welch’s t-test was applied for non-normally distributed data. Enumeration data were analyzed using the chi-square test. For comparisons among multiple groups, one-way ANOVA followed by appropriate post hoc multiple comparison tests (Tukey’s or Dunnett’s test, as appropriate) was performed to control for type I error. Pearson’s correlation test was applied for correlation analyses. A p-value < 0.05 was considered statistically significant.
Results
Plasma IL-27 Levels Were Significantly Increased in ACS Patients
There were no significant differences in age, gender, serum TC, TG, LDL-C, blood glucose, history of hypertension, or smoking history among the three groups. However, the ox-LDL, C-reactive protein(CRP), and Gensini scores were significantly elevated in AMI patients compared to UA, SAP, and healthy controls. Additionally, UA patients exhibited higher ox-LDL, CRP, and Gensini scores than SAP and healthy controls, and SAP patients showed significantly elevated levels of these markers compared to the healthy controls (Table 2).
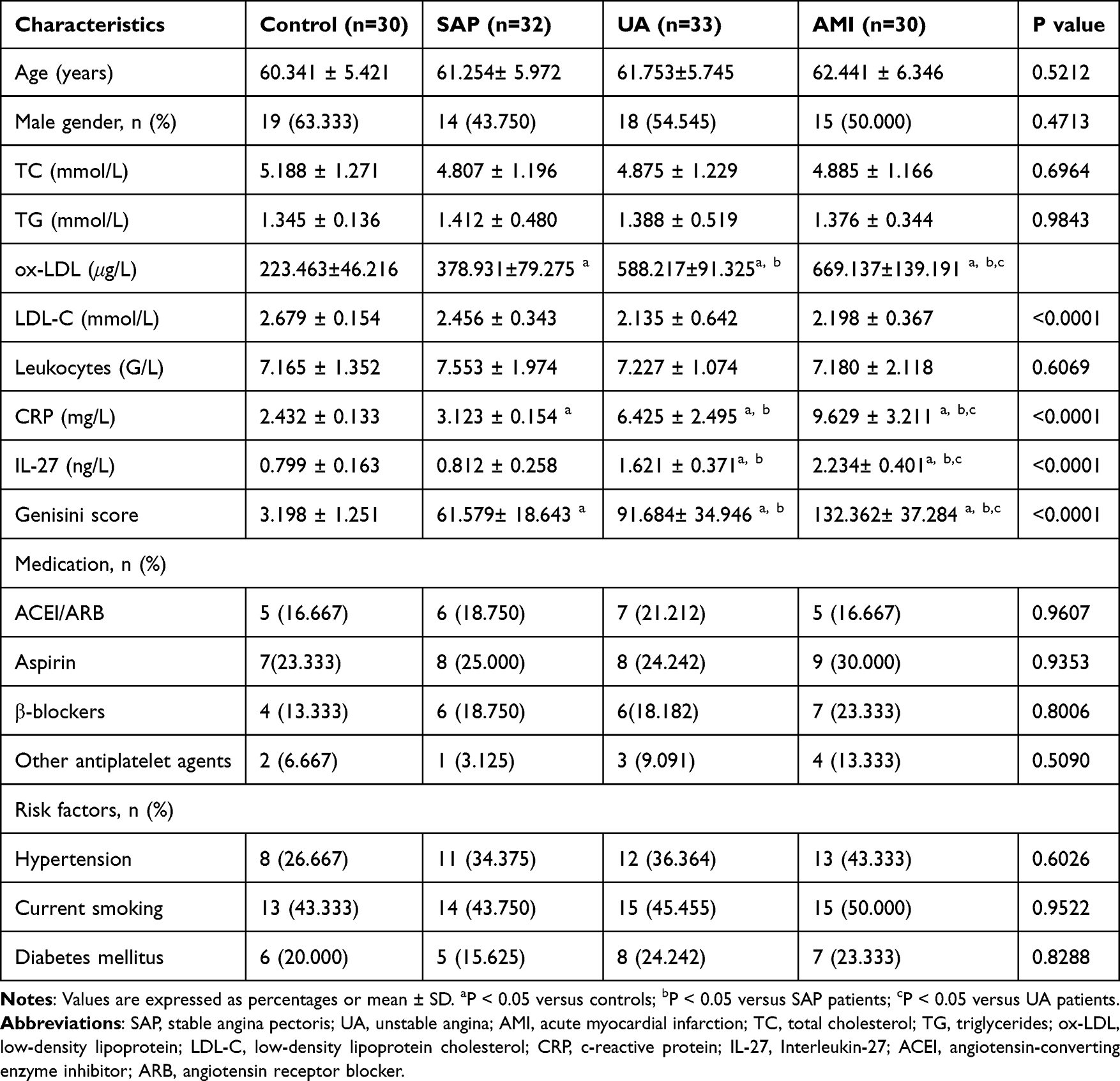 |
Table 2 Clinical Characteristics of Patients |
As shown in Figure 1a, plasma IL-27 levels in AMI patients (2.234 ± 0.401 ng/L) were significantly higher than those in SAP patients (0.812 ± 0.258 ng/L) and healthy controls (0.799 ± 0.163 ng/L) (P < 0.05). Moreover, IL-27 levels in UA patients (1.621 ± 0.371 ng/L) were significantly higher than those in SAP patients and healthy controls, although there was no significant difference between SAP patients and healthy controls.
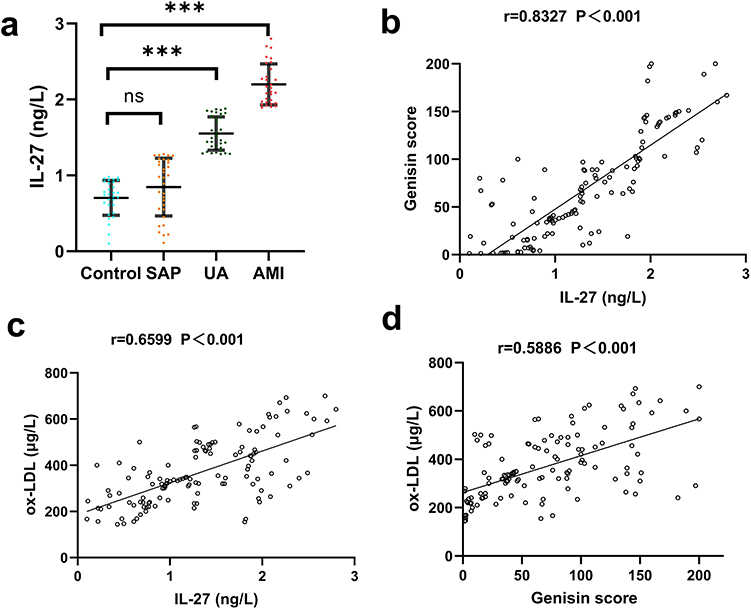 |
Figure 1 Comparison of molecule levels in control, SAP, UA, and AMI patients. (a) Plasma IL-27 levels. (b) Correlation between IL-27 and Gensini score. (c) Correlation between IL-27 and ox-LDL. (d) Correlation between Gensini score and ox-LDL. Colors indicate groups: blue = control, Orange = SAP, green = UA, red = AMI. ***P < 0.001, ns: not significant. |
Coronary Angiographic Findings
Compared with SAP, patients with UA and AMI showed more severe and complex coronary lesions. The proportions of multivessel disease, left main involvement, intracoronary thrombosis, vulnerable plaques, and severe stenosis were significantly higher in the acute coronary syndrome groups. In parallel, the mean number of diseased vessels increased progressively from SAP to UA and AMI (all P < 0.05). The detailed angiographic findings are summarized in Table 3.
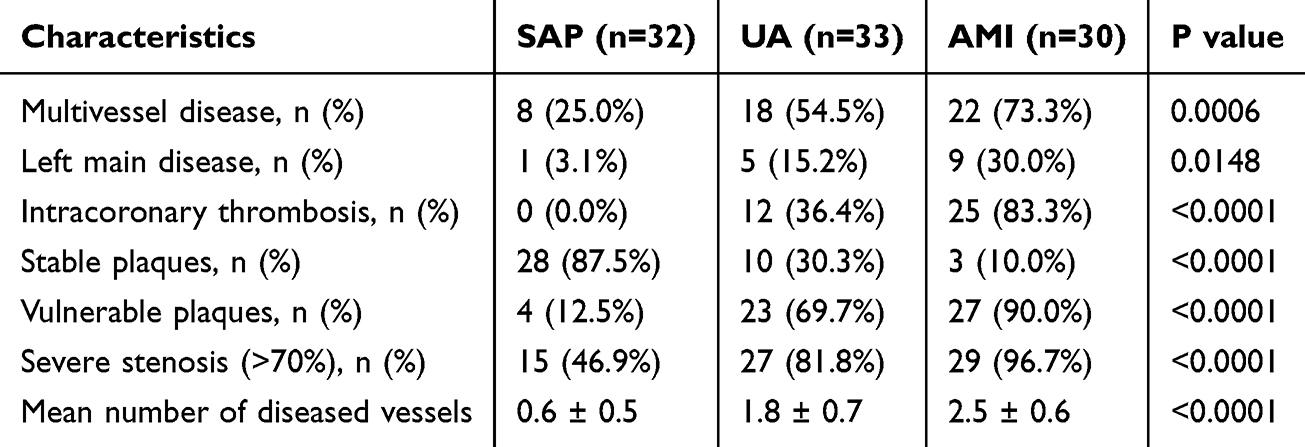 |
Table 3 Coronary Angiographic Characteristics of Study Participants |
Relationship Between IL-27 Levels and Key Clinical Indicators in CAD Patients
To investigate the association between IL-27 levels and key clinical indicators in CAD patients, we analyzed correlations between IL-27 levels, ox-LDL, and Gensini scores. As shown in Figure 1b and c, circulating IL-27 levels were positively correlated with both ox-LDL and Gensini scores (P < 0.01). Similarly, a significant positive correlation was observed between ox-LDL levels and Gensini scores (P < 0.01) (Figure 1d). These findings suggest that elevated IL-27 levels are associated with the severity of coronary artery stenosis.
Comparison of Th1, Th2, Th17, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T Cell Frequencies Among CAD Patients
The frequencies of Th1, Th2, Th17, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T cells in peripheral blood were measured by flow cytometry in healthy controls, SAP, UA, and AMI patients. As shown in Figure 2, no significant differences were found in these cell types between healthy controls and SAP patients (P > 0.05). However, AMI patients exhibited significantly higher frequencies of Th1 and Th17, along with lower frequencies of Th2, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T cells, compared to SAP patients and healthy controls (P < 0.05). Similarly, UA patients had elevated Th1 and Th17 frequencies and decreased Th2, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T cells compared to SAP patients and healthy controls (P < 0.05).
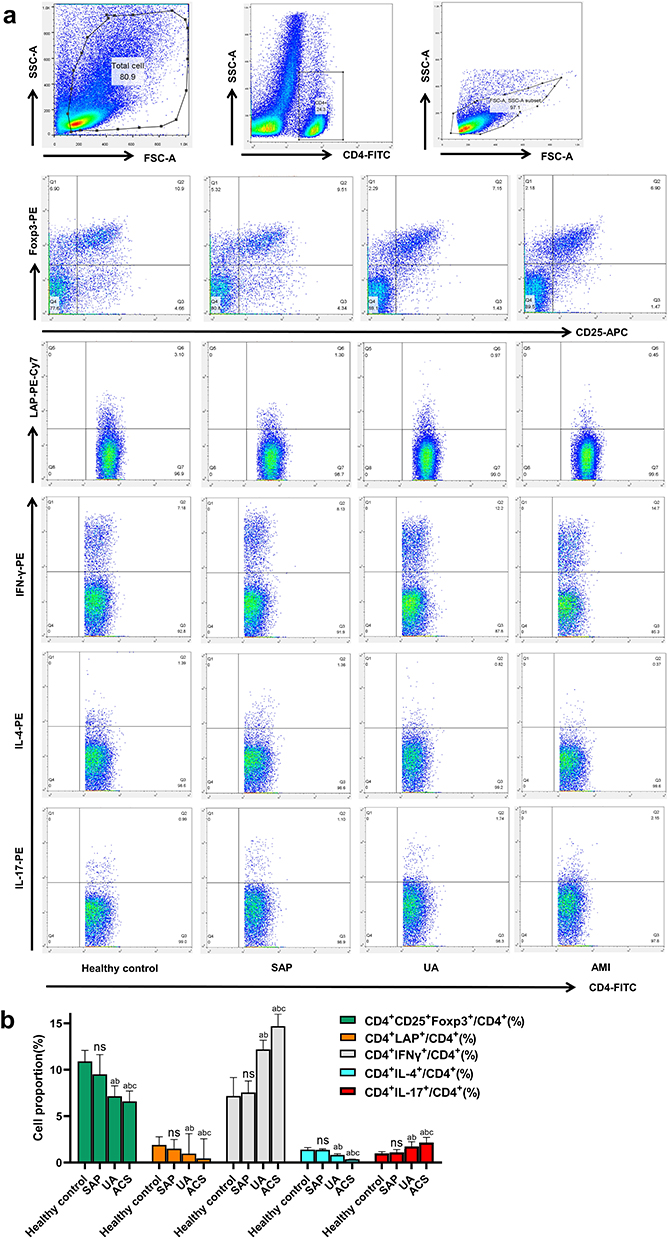 |
Figure 2 Th1, Th2, Th17, CD4 + LAP + T cells and CD4 + CD25 + Foxp3 + Tregs in peripheral blood of healthy controls (n=30), SAP (n=32), UA (n=33) and AMI (n=30) were detected. (a) Representative FACS images; (b) Comparison of Th1, Th2, Th17, CD4 + LAP + T cells and CD4 + CD25 + Foxp3 + Tregs. Bar colors indicate cell subsets: green = CD4⁺CD25⁺Foxp3⁺ Tregs, Orange = CD4⁺LAP⁺, gray = CD4⁺IFN-γ⁺ (Th1), cyan = CD4⁺IL-4⁺ (Th2), red = CD4⁺IL-17⁺ (Th17). a: P < 0.05 versus healthy controls; b: P < 0.05 versus SAP patients; c: P < 0.05 versus UA patients. Abbreviation: ns, No significant difference. |
Effects of IL-2 and IL-27 on the Frequencies of Th1, Th2, Th17, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T Cells
PBMCs from healthy controls were divided into four groups: blank control, IL-2, IL-27, and IL-2 + IL-27. Flow cytometry analysis revealed that IL-2 significantly increased the frequencies of CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells compared to the blank healthy controls (P < 0.05). In contrast, IL-27 significantly decreased the frequencies of CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells (P < 0.05). When IL-2 and IL-27 were added together, the frequencies of CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells were significantly reduced compared to IL-2 alone (P < 0.05) (Figure 3).
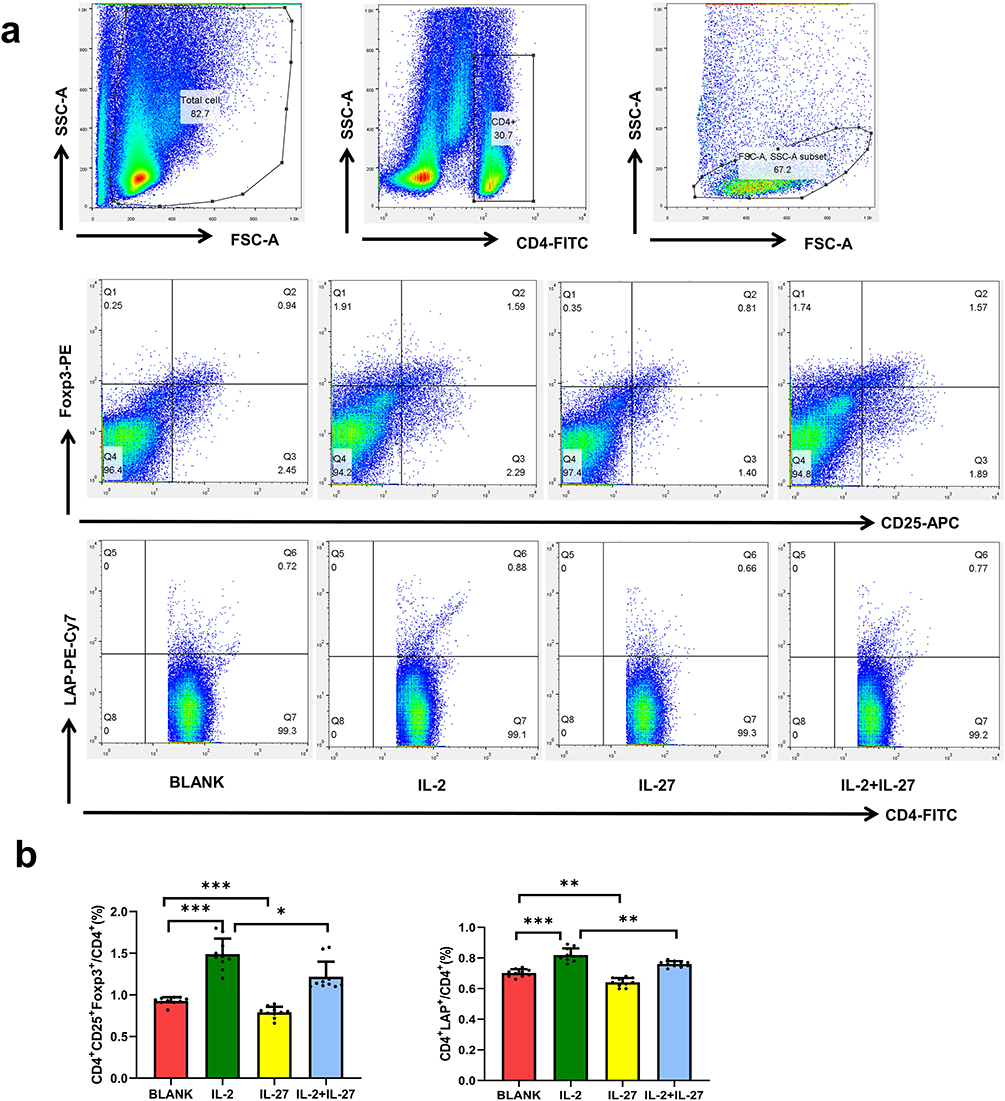 |
Figure 3 Effects of IL-2 and IL-27 on CD4⁺LAP⁺ T cells and CD4⁺CD25⁺Foxp3⁺ Tregs in vitro. (a) Representative FACS images; (b) Comparison of CD4 + LAP + T cells and CD4 + CD25 + Foxp3 + Tregs. Bar colors: red = BLANK, green = IL-2, yellow = IL-27, blue = IL-2+IL-27. ***P < 0.001, **P < 0.01, *P < 0.05 (n=10). |
For Th1, Th2, and Th17 cells, IL-2 significantly increased their frequencies compared to the healthy controls (P < 0.05). IL-27 alone increased Th1 frequency and decreased Th2 and Th17 frequencies (P < 0.05). When IL-2 and IL-27 were co-administered, Th1 frequency increased, Th17 frequency showed no significant change (P > 0.05), and Th2 frequency decreased compared to IL-2 alone (P < 0.05) (Figure 4).
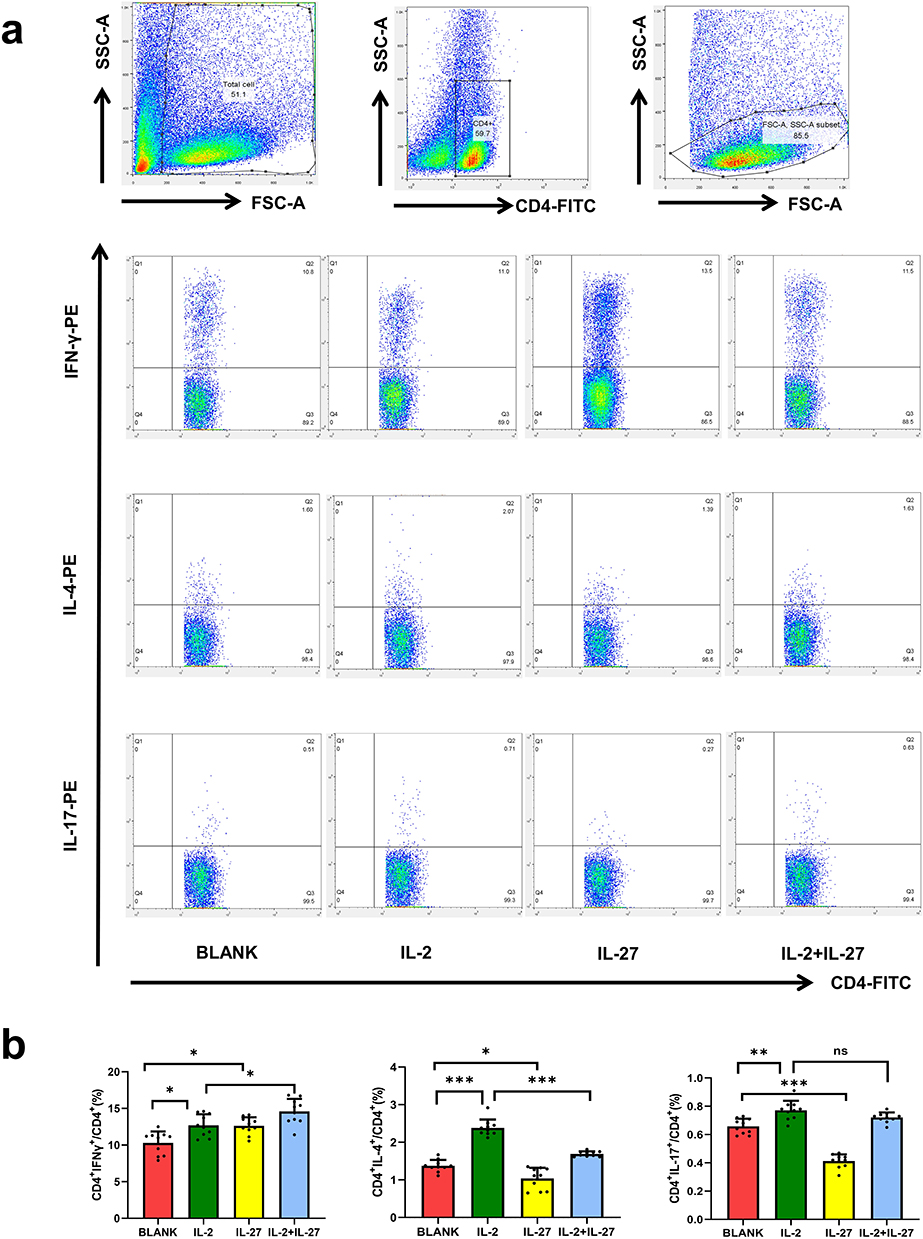 |
Figure 4 Effects of IL-2 and IL-27 on Th1, Th2, and Th17 cells in vitro. (a) Representative FACS images; (b) Comparison of Th1, Th2 and Th17. Bar colors: red = BLANK, green = IL-2, yellow = IL-27, blue = IL-2+IL-27. ***P < 0.001, **P < 0.01, *P < 0.05, ns: No significant difference (n=10). |
Effects of Different Concentrations of IL-27 on the Frequencies of Th1, Th2, Th17, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T Cells
To further examine the concentration-dependent effects of IL-27 on these cell frequencies, PBMCs were treated with IL-27 at concentrations of 0, 50, 100, and 200 ng/mL. The results showed that increasing IL-27 concentrations led to a gradual increase in Th1 frequency and a corresponding decrease in Th17, Th2, CD4+CD25+Foxp3+ Tregs, and CD4+LAP+ T cells (Figure 5).
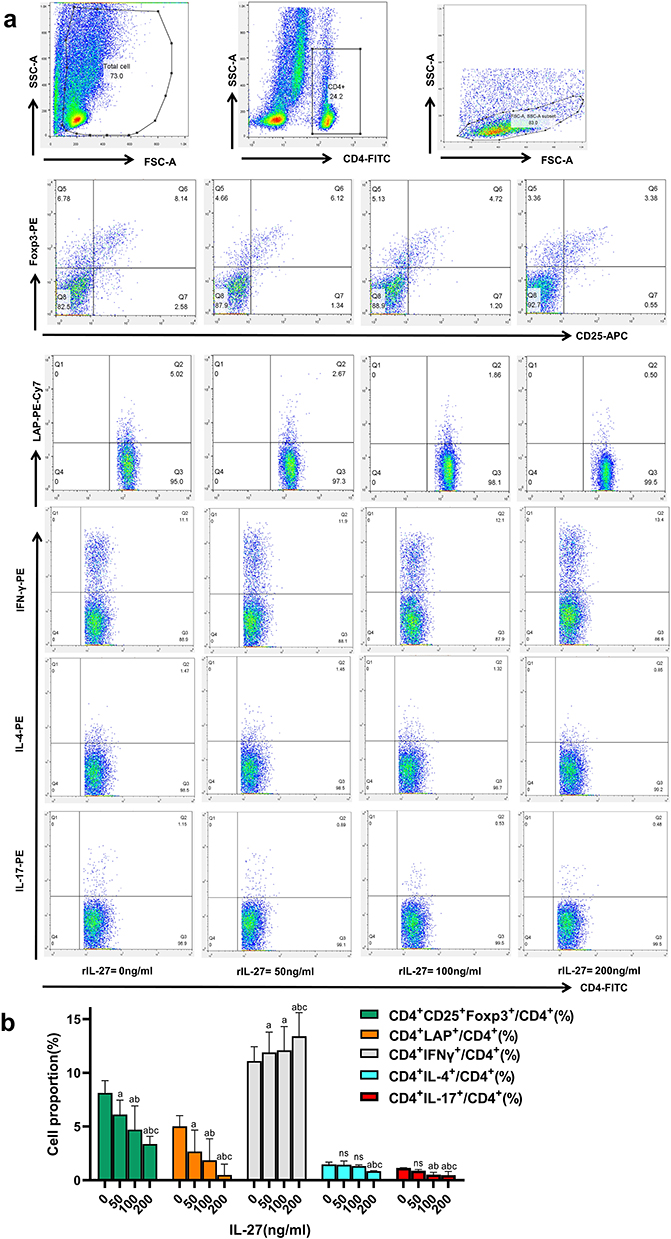 |
Figure 5 Dose-dependent effects of IL-27 on Th1, Th2, Th17, CD4⁺LAP⁺ T cells, and CD4⁺CD25⁺Foxp3⁺ Tregs. (a) Representative FACS images; (b) Comparison of Th1, Th2, Th17, CD4 + LAP + T cells and CD4 + CD25 + Foxp3 + Tregs. Bar colors denote cell subsets: green = CD4⁺CD25⁺Foxp3⁺ Tregs; Orange = CD4⁺LAP⁺; gray = CD4⁺IFN-γ⁺ (Th1); cyan = CD4⁺IL-4⁺ (Th2); red = CD4⁺IL-17⁺ (Th17). Data are expressed as the percentage of each subset within total CD4⁺ T cells (/CD4⁺%). a: P < 0.05 when compared to IL-27 concentration=0ng/mL; 0.05; b: P < 0.05 when compared to IL-27 concentration=50ng/mL; c: P < 0.05 when compared to IL-27 concentration=100ng/mL. Abbreviation: ns, No significant difference. |
IL-27 Increases T-Bet and IFN-γ Expression While Inhibiting RORγt and GATA-3 Expression
To further confirm the effects of IL-27 on Th1, Th2, and Th17 cells, PBMCs from healthy controls were stimulated with either 200 ng/mL rhIL-27 alone or 200 ng/mL rhIL-27 combined with anti-IL-27p28. The relative mRNA and protein expression levels of Th1-related T-bet and IFN-γ, Th2-related GATA-3, and Th17-related RORγt were measured. As shown in Figure 6, IL-27 significantly increased the expression of T-bet and IFN-γ, while inhibiting the expression of RORγt and GATA-3 (Figure 6).
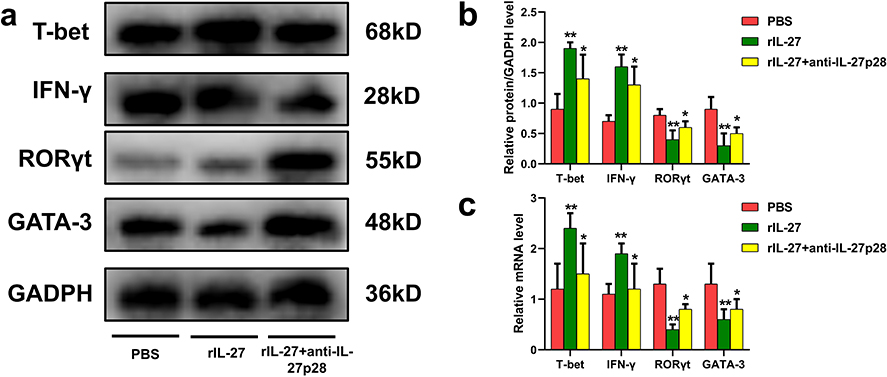 |
Figure 6 Effects of IL-27 and anti-IL-27p28 on Th1-, Th2-, and Th17-related transcription factors. (a) Representative WB images of T-bet, IFN-γ, RORγt, GATA-3, and GAPDH. (b) Relative protein expression normalized to GAPDH. (c) Relative mRNA expression of T-bet, IFN-γ, RORγt, and GATA-3. Bar colors: red = PBS, green = rhIL-27, yellow = rhIL-27+anti-IL-27p28. **P < 0.01 when versus the PBS group, *P < 0.05 when versus the PBS group. |
Discussion
CAD includes SAP, UA and AMI, which can be considered as atherosclerosis of different severity.19 The pathogenesis of atherosclerosis has not been completely understood. At present, there are theories of lipid infiltration, smooth muscle cloning, thrombosis, inflammatory reaction, etc. Among them, the inflammatory reaction theory has a more comprehensive explanation of the pathogenesis of atherosclerosis, which has been widely recognized in the academic community.20 Inflammatory response can lead to atherosclerotic plaque instability. The greater the inflammatory response, the worse the plaque stability and the more severe the CAD.21 Unstable plaque rupture is the most common cause of ACS, which is typically characterized by thin fibrous cap, lipid core, and infiltration of numerous inflammatory cells and inflammatory factors (including macrophages, T lymphocytes, etc).22 Studies have shown that serum inflammatory marker C-reactive protein (CRP) can predict the instability of coronary atherosclerotic plaque and is proportional to the incidence and severity of cardiovascular disease.23 In addition, oxidized low-density lipoprotein (ox-LDL) degrades the fibrous cap of coronary atherosclerosis and increases plaque instability, and studies have shown that elevated levels of cholesterol and ox-LDL are the main causes of atherosclerosis.24 To further verify the above views, we also focused on measuring CRP, ox-LDL, cholesterol and other indicators of CAD patients. Our results showed that the levels of ox-LDL and CRP in healthy controls, SAP, UA and AMI patients increased in turn with statistical significance. But there was no significant difference in their cholesterol level. In addition, the Gensini score has been widely accepted as an angiographic scoring system for quantifying the severity of CAD. It can be assumed that the higher the score, the more severe the CAD. We demonstrated a positive correlation between ox-LDL level and Gensini score. These conclusions confirm that ox-LDL and CRP are closely related to the development of CAD.
Among the various cytokines involved in CAD, IL-27 is an interesting presence. Firstly, IL-27 is considered to be a cytokine with biphasic effects on CD4+ T cells, regulating CD4+ T cell function in a pro-inflammatory and anti-inflammatory manner. Secondly, IL-27 has been shown to protect against atherosclerosis in animal studies, but it remains controversial in clinical studies. Our study showed that the level of IL-27 in patients with AMI and UA was significantly higher than that in SAP and healthy people, and the level of IL-27 in patients with AMI was significantly higher than that in patients with UA. And IL-27 was significantly positively correlated with ox-LDL and Gensini score. The above results suggested that IL-27 may be used as a biomarker of CAD, and the higher the level of IL-27, the higher the severity of CAD. Our results are consistent with J Wen13 However, Miura et al25 found that the level of IL-27 in patients with CAD was higher than that in patients without CAD, but there was no difference in the level of IL-27 between patients with ACS and SAP, indicating that the level of IL-27 may only be an independent factor of CAD rather than ACS. It could not provide a good evaluation of the severity of the CAD patients. Unexpectedly, Wang et al26 demonstrated that the level of IL-27 in patients with AMI and UA was significantly lower than that in patients with SAP and healthy controls, and the reduction of IL-27 may be an important factor in the development of ACS. The differences in these results could be due to a variety of factors, including selection bias. In conclusion, our results tend to suggest that IL-27 can be a good biomarker of CAD, representing the severity of CAD.
In the area of atherosclerotic plaque, a large number of macrophages and CD4+ T cells infiltrate, but the changes of CD4+ T cell subsets in CAD are controversial. Studies have shown that the expression of Th1 and Th17 in peripheral blood of patients with ACS is increased, accompanied by decreased expression of Treg and Th2, resulting in decreased Th2/Th1 and Treg/Th17 ratios.27 Other studies have shown that circulating Th1 and Th17 cells are increased in ACS patients compared with SAP patients and healthy controls, but there is no significant difference between SAP patients and healthy controls.28 Surprisingly, Kim et al29 found that the frequency of Th1 and Th17 cells in SAP patients was significantly increased compared with healthy controls, while there was no significant difference in Th1 and Th17 cells between AMI patients and healthy controls, possibly because of the different time of blood collection. Kim JD’s study collected blood 6 hours after the onset of symptoms, while other studies, including ours, peripheral blood was collected within 24–48 hours after symptom onset. Studies have shown that Th1 and Th17 cells gradually increase with time after the onset of AMI. In our study, we collected peripheral blood between 24 and 48 hours after the onset of AMI, and found that there was no significant difference in the frequencies of five types of cells in healthy controls and SAP patients, while the frequencies of Th1 and Th17 in SAP, UA and AMI patients gradually increased. The frequencies of Th2, CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells decreased gradually. The most innovative aspect of this result is the measurement of the frequency of CD4+LAP+ T cells. CD4+LAP+ T cells are a new type of regulatory T cells. Animal experiments have shown that this cell population is more immunosuppressive than CD4+CD25+ Tregs. CD4+LAP+ T cells also exist in human peripheral blood.30 Some studies have shown that CD4+CD25+ Treg cells may be related to plaque instability.31 However, few studies have reported the changes of CD4+LAP+ T cells in CAD patients. Our study proved that the changes of CD4+LAP+ T cells were consistent with the changes of CD4+CD25+Foxp3+ Tregs. The changes in these five cells further support the inflammatory theory of atherosclerosis.
At present, many studies on the association between IL-27 and CAD are limited to the changes of IL-27 levels in CAD patients, and few studies have studied the effects of IL-27 on the number of various subtypes of CD4+ T cells.25 Our study proved that IL-2 could increase the frequency of Th1, Th2, Th17, CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells, while IL-27 could increase the frequency of Th1 cells and significantly reduce Th2, Th17, CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells. Firstly, it is well known that IL-2 is one of the essential cytokines for T cell proliferation and activation. In addition to stimulating T cell activation, IL-2 can also activate intracellular STAT5 phosphorylation by binding to receptors, thereby promoting the expression of specific transcription factor Foxp3 to promote Treg cell differentiation.32 In view of this effect of IL-2, four groups were established in this study: blank group, IL-2 group, IL-27 group, and IL-2+IL-27 group, so as to make the effect of IL-27 on the five types of cells more convincing. IL-2 can increase the frequency of these five types of cells probably because it has the effect of activating and proliferating T cells, while IL-27 can increase the frequency of Th1 cells and significantly reduce the frequency of Th2, Th17, CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells, and the effect is enhanced with the increase of concentration. In order to investigate the mechanism, we treated monocytes with the most effective concentration of 200 ng/mL, and found that IL-27 could increase the expression of T-bet and IFN-γ, and inhibit the expression of RORγt and GATA-3. T-bet, GATA-3 and RORyt are specific transcription factors that control the differentiation of Th1, Th2 and Th17 cells, respectively. Based on the results, we speculated that the mechanism of IL-27 on CAD was similar to other diseases: IL-27 increased the expression of T-bet and IFN-γ, and inhibited the expression of RORγT and GATA-3, thereby promoting the differentiation of CD4+ T cells into Th1 cells and inhibiting the differentiation into Th2, Th17 and Tregs. Studies have proved that Th1 cells secrete pro-inflammatory factors, which can promote the secretion of matrix metalloproteinases and the apoptosis of smooth muscle cells, making the fiber cap thin and easy to rupture. Th2 releases anti-inflammatory factors. Increased Th1 and decreased Th2 in CAD patients promote plaque instability and the development of ACS.33 Treg cells are typical anti-inflammatory cells that stabilize atherosclerotic plaques. Therefore, IL-27 can be considered to promote the differentiation of CD4+ T cells into Th1 cells and inhibit the differentiation into Th2 and Treg cells, thereby promoting the progression of atherosclerosis and CAD disease.34 Gregersen et al10 studied the effect of IL-27 on human Th1 cells, primary monocytes and PBMC in vitro, and found that IL-27 increased NLRP3 through LPS signal in a dose-dependent manner, thereby increasing the release of IL-1β and promoting the development of atherosclerosis. This further demonstrated the role of IL-27 in promoting atherosclerosis. However, as for Th17, it has both promoting and inhibiting effects on atherosclerosis. Cirillo et al35 found that the level of IL-17 was also higher when the number of Th17 cells was increased, and Th17 and its secreted IL-17 had pro-atherogenic effects. As a subclass of Th17, Th17/Th1 cells had stronger atherogenic effects than Th17 cells.36 However, it has also been demonstrated that IL-17 maintains plaque stability by inducing the proliferation of smooth muscle cells(SMCs)and collagen in atherosclerotic plaques,37 and the increased content of SMC and collagen in fibrous caps is essential for plaque stability.38 Our study found that IL-27 can reduce Th17 by inhibiting RORγt, so it is unknown whether IL-27 has a promoting or inhibitory effect on CAD in this respect. We suggest that unlike animal studies, where IL-27 protects against CAD, in humans, IL-27 cannot be simply generalized to inhibit or protect against CAD, but rather regulates CAD by increasing T-bet, IFN-γ expression, and inhibiting RORγt and GATA-3 expression, thereby elevating Th1 cells and decreasing the frequency of Th2, Th17, CD4+CD25+Foxp3+ Treg, and CD4+LAP+ T cells.
In fact, the number of CD4+ T cell subtypes in the normal body is dynamically balanced, so that the ratio of Th1/Th2 cells and Treg/Th17 cells remains stable Many studies have demonstrated that the number of these cells in CAD can be unbalanced, leading to changes in the ratio. In studies of osteosarcoma39 and lung cancer,40 IL-27 can induce the expression of T-BET by activating STAT1, so that initial CD4+ T cells can differentiate into Th1 cells and produce IFN-γ under the action of IL-12. On the other hand, IL-27 can inhibit Th2 cell differentiation by down-regulating the transcription factor GATA-3 expressed by initial CD4+ T cells, thus playing an anti-inflammatory and anti-tumor role. In the study of colitis,41 IL-27 can play an anti-inflammatory role by inhibiting Th2 and Th17, thus inhibiting the development of acute colitis. While the association between IL-27 and Treg is controversial, Treg is an important immunosuppressive cell of the body that helps tumors to evade the body’s immunity. Most studies have demonstrated that IL-27 can inhibit Treg production and thus achieve anti-tumor effects,42 but some studies have also found that IL-27 can promote the process of peripheral Treg recruitment to tumor tissue.43 IL-27 was also found to promote Treg differentiation in in vitro cultures, but was shown to limit Treg amplification in an in vivo model.44 We believe that the possible reason is that the effect of IL-27 on Tregs is environment-dependent, and different diseases have different specific microenvironments, leading to different targets of IL-27 and thus producing different effects. We were therefore curious to see how IL-27 acts on various subtypes of CD4+ T cells in CAD. The results showed that, similar to colitis, IL-27 increased the frequency of Th1 cells and significantly decreased the frequency of Th2 and Th17 cells. Similar to most tumor studies, IL-27 significantly decreased the frequency of CD4+CD25+Foxp3+ Tregs and CD4+LAP+ T cells. However, in colitis, IL-27 reduces the frequency of Th2 and Th17, which plays an anti-inflammatory role and inhibits the development of colitis. We can only prove that the regulatory mechanism of IL-27 on CAD is to increase the expression of T-BET and IFN-γ, and inhibit the expression of RORγt and GATA-3, thereby increasing Th1 cells and decreasing Th2 and Th17. The frequency of CD4+CD25+Foxp3+ Tregs, CD4+LAP+ T cells, and IL-27 can be used as good biomarkers of coronary heart disease, which is proportional to the stenosis degree of coronary heart disease. In addition, it has been previously demonstrated that elevated baseline IL-27 levels are independent risk factors for predicting major adverse cardiac events (MACE) in patients with ACS.45 However, it is unknown whether IL-27 has a direct pathogenic effect on CAD patients. Exogenous administration of IL-27 can inhibit the progression of CAD in animals,46 but whether it promotes or inhibits human CAD is also unknown. There are few studies on IL-27 in clinical practice, and the studies on the anti-CAD effect of IL-27 mainly focus on animal experiments. Therefore, there are still many problems to be solved before the immunotherapy of IL-27 can be applied in clinical practice.
Limitations
This study has several limitations. First, the moderate sample size (n=125) may not provide sufficient power to detect small effect sizes, although it remains acceptable for exploratory immunological analyses. Second, echocardiographic data, especially left ventricular function parameters, were not analyzed, and only peripheral blood measurements were performed, which may not fully reflect local coronary immune responses. In addition, measurements were restricted to peripheral blood and taken at a single time point, which may not capture local or temporal changes in IL-27 and T cell subsets. Third, the single-center design may limit the generalizability of the findings. To reduce type I error, adjustments for multiple comparisons were considered where appropriate.
Conclusion
In summary, this study shows that in patients with acute coronary syndrome, plasma IL-27 levels are increased and correlate positively with ox-LDL levels and Gensini scores, which indicate the severity of coronary artery disease. Furthermore, in vitro work suggested that IL-27 induces Th1 differentiation, inhibits Th2, Th17, and regulatory subsets, and inhibits IL-2–induced regulatory cell proliferation. These results indicate that IL-27 promotes immune imbalance in coronary artery disease and has the potential to be both a marker for disease severity and a therapeutic target. However, it should be noted that the mechanistic data was obtained in vitro and needs further in vivo experimentation.
Data Sharing Statement
Individual deidentified participant data underlying the results of this article, including demographic, baseline, and outcome information, as well as the study protocol and statistical analysis plan, will be available upon reasonable request. These data will not be deposited on a public platform but may be obtained by contacting the corresponding author (Dr. Yucheng Zhong, Email: [email protected]). Data will be accessible beginning 6 months after publication and for up to 5 years thereafter.
Ethics Approval
This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the Ethics Committee of the Union Hospital, Tongji Medical College (2021.07.16/ UHCT22214).
This study was registered in the Chinese Clinical Trial Registry (https://www.chictr.org.cn/) under the registration number ChiCTR1900023770.
Consent to Participate
Informed consent was obtained from all individual participants included in the study.
Acknowledgments
This paper has been uploaded to Research Square as a preprint: https://www.researchsquare.com/article/rs-1964617/v1.47
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (grant numbers 81300213, 82070310, 81770273, 81900400, 81900270, 82100339).
Disclosure
The authors have no relevant financial or non-financial interests to disclose.
References
1. Ralapanawa U, Sivakanesan R. Epidemiology and the magnitude of coronary artery disease and acute coronary syndrome: a narrative review. J Epidemiol Glob Health. 2021;11(2):169–177. doi:10.2991/jegh.k.201217.001
2. Schofer N, Ludwig S, Rübsamen N, et al. Prognostic impact of Interleukin-1 receptor antagonist in patients with documented coronary artery disease. Int J Cardiol. 2018;257:24–29. doi:10.1016/j.ijcard.2018.01.055
3. Bullenkamp J, Mengoni V, Kaur S, et al. Interleukin-7 and interleukin-15 drive CD4+CD28null T lymphocyte expansion and function in patients with acute coronary syndrome. Cardiovasc Res. 2021;117(8):1935–1948. doi:10.1093/cvr/cvaa202
4. Gang H, Peng D, Hu Y, et al. InterleukIn-9-secretIng CD4+ T cells regulate CD8+ T cells cytotoxIcIty In patIents wIth acute coronary syndromes. Apmis. 2021;129(2):91–102. doi:10.1111/apm.13094
5. Saigusa R, Winkels H, Ley K. T cell subsets and functions in atherosclerosis. Nat Rev Cardiol. 2020;17(7):387–401. doi:10.1038/s41569-020-0352-5
6. Khantakova JN, Bulygin AS, Sennikov SV. The regulatory-T-cell memory phenotype: what we know. Cells. 2022;11(10):1687. doi:10.3390/cells11101687
7. Zong Y, Deng K, Chong WP. Regulation of Treg cells by cytokine signaling and co-stimulatory molecules. Front Immunol. 2024;15:1387975. doi:10.3389/fimmu.2024.1387975
8. Ye J, Wang Y, Wang Z, et al. Roles and mechanisms of interleukin-12 family members in cardiovascular diseases: opportunities and challenges. Front Pharmacol. 2020;11:129. doi:10.3389/fphar.2020.00129
9. Kawahara E, Azuma M, Nagashima H, et al. TNF receptor-associated factor 5 limits IL-27 receptor signaling in CD4+ T lymphocytes. J Immunol. 2022;208(3):642–650. doi:10.4049/jimmunol.2001358
10. Gregersen I, Sandanger Ø, Askevold ET, et al. Interleukin 27 is increased in carotid atherosclerosis and promotes NLRP3 inflammasome activation. PLoS One. 2017;12(11):e0188387. doi:10.1371/journal.pone.0188387
11. Peshkova IO, Fatkhullina AR, Mikulski Z, et al. IL-27R signaling healthy controls myeloid cells accumulation and antigen-presentation in atherosclerosis. Sci Rep. 2017;7(1):2255. doi:10.1038/s41598-017-01828-8
12. Shi Y, Zhang S, Xue Y, et al. IL-35 polymorphisms and cognitive decline did not show any association in patients with coronary heart disease over a 2-year period: a retrospective observational study (STROBE compliant). Medicine. 2020;99(31):e21390. doi:10.1097/MD.0000000000021390
13. Jin W, Zhao Y, Yan W, et al. Elevated circulating interleukin-27 in patients with coronary artery disease is associated with dendritic cells, oxidized low-density lipoprotein, and severity of coronary artery stenosis. Mediators Inflamm. 2012;2012:506283. doi:10.1155/2012/506283
14. Zhu J, Yu J, Hu A, et al. IL-27 gene therapy induces Stat3-mediated expansion of CD11b+Gr1+ myeloid cells and promotes accumulation of M1 macrophages in the tumor microenvironment. J Immunol. 2023;211(5):895–902. doi:10.4049/jimmunol.2300176
15. Thygesen K, Alpert JS, Jaffe AS, et al. Fourth universal definition of myocardial infarction (2018). J Am Coll Cardiol. 2018;72(18):2231–2264. doi:10.1016/j.jacc.2018.08.1038
16. Knuuti J, Wijns W, Saraste A, et al. 2019 ESC Guidelines for the diagnosis and management of chronic coronary syndromes. Eur Heart J. 2020;41(3):407–477. doi:10.1093/eurheartj/ehz425
17. Ponikowski P, Voors AA, Anker SD, et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: the task force for the diagnosis and treatment of acute and chronic heart failure of the European society of cardiology (ESC) Developed with the special contribution of the heart failure association (HFA) of the ESC. Eur Heart J. 2016;37(27):2129–2200. doi:10.1093/eurheartj/ehw128
18. Wang KY, Zheng YY, Wu TT, et al. Predictive value of gensini score in the long-term outcomes of patients with coronary artery disease who underwent PCI. Front Cardiovasc Med. 2022;8:778615. doi:10.3389/fcvm.2021.778615
19. Liu L, Shi Z, Ji X, et al. Adipokines, adiposity, and atherosclerosis. Cell Mol Life Sci. 2022;79(5):272. doi:10.1007/s00018-022-04286-2
20. Yang Y, Wang D, Zhang C, et al. Piezo1 mediates endothelial atherogenic inflammatory responses via regulation of YAP/TAZ activation. Hum Cell. 2022;35(1):51–62. doi:10.1007/s13577-021-00600-5
21. Liu C, Yu Z, Chen H, et al. Relationship between immunoinflammation and coronary physiology evaluated by quantitative flow ratio in patients with coronary artery disease. Front Cardiovasc Med. 2021;8:714276. doi:10.3389/fcvm.2021.714276
22. Sagris M, Theofilis P, Antonopoulos AS, et al. Inflammation in coronary microvascular dysfunction. Int J Mol Sci. 2021;22(24):13471. doi:10.3390/ijms222413471
23. Blaum C, Brunner FJ, Kröger F, et al. Modifiable lifestyle risk factors and C-reactive protein in patients with coronary artery disease: implications for an anti-inflammatory treatment target population. Eur J Prev Cardiol. 2021;28(2):152–158. doi:10.1177/2047487319885458
24. Song B, Dang H, Dong R. Differential expression of LOXL1-AS1 in coronary heart disease and its regulatory mechanism in ox-LDL-induced human coronary artery endothelial cell pyroptosis. Cardiovasc Drugs Ther. 2023;37(1):75–87. doi:10.1007/s10557-021-07274-z
25. Miura K, Saita E, Suzuki-Sugihara N, et al. Plasma interleukin-27 levels in patients with coronary artery disease. Medicine. 2017;96(43):e8260. doi:10.1097/MD.0000000000008260
26. Wang Y, Zhou C, Yu T, et al. Correlation between changes in serum RBP4, hs-CRP, and IL-27 levels and rosuvastatin in the treatment of coronary heart disease. J Healthc Eng. 2021;2021:8476592. doi:10.1155/2021/8476592
27. Inui H, Nishida M, Ichii M, et al. XCR1+ conventional dendritic cell-induced CD4+ T helper 1 cell activation exacerbates cardiac remodeling after ischemic myocardial injury. J Mol Cell Cardiol. 2023;176:68–83. doi:10.1016/j.yjmcc.2023.01.011
28. Ma X, Liu S, Li T, et al. Intensive statin treatment ameliorate the Th17/Treg functional imbalance in patients with non-ST elevation acute coronary syndrome underwent percutaneous coronary intervention. Clin Cardiol. 2020;43(4):379–385. doi:10.1002/clc.23326
29. Kim JD, Lee SH, Seo EH, et al. Role of Th1 and Th17 cells in the development and complexity of coronary artery disease: comparison analysis by the methods of flow cytometry and SYNTAX score. Coron Artery Dis. 2015;26(7):604–611. doi:10.1097/MCA.0000000000000289
30. Martínez-Shio EB, Marín-Jáuregui LS, Rodríguez-Ortega AC, et al. Regulatory T-cell frequency and function in AMI patients and its correlation with ventricular dysfunction. Clin Exp Immunol. 2024;216(3):262–271. doi:10.1093/cei/uxae014
31. Mor A, Luboshits G, Planer D, et al. Altered status of CD4(+)CD25(+) regulatory T cells in patients with acute coronary syndromes. Eur Heart J. 2006;27(21):2530–2537. doi:10.1093/eurheartj/ehl222
32. Jones DM, Read KA, Oestreich KJ. Dynamic roles for IL-2-STAT5 signaling in effector and regulatory CD4+ T cell populations. J Immunol. 2020;205(7):1721–1730. doi:10.4049/jimmunol.2000612
33. Zhao Z, Wu Y, Cheng M, et al. Activation of Th17/Th1 and Th1, but not Th17, is associated with the acute cardiac event in patients with acute coronary syndrome. Atherosclerosis. 2011;217(2):518–524. doi:10.1016/j.atherosclerosis.2011.03.043
34. McCurdy S, Yap J, Irei J, et al. IL-37-a putative therapeutic agent in cardiovascular diseases. QJM. 2022;115(11):719–725. doi:10.1093/qjmed/hcab011
35. Cirillo P, Golino P, Piscione F, et al. Transcoronary Th-17 lymphocytes and acute coronary syndromes: new evidence from the crime scene? Int J Cardiol. 2011;153(2):215–216. doi:10.1016/j.ijcard.2011.09.063
36. Zheng Z, Lu H, Wang X, et al. Integrative analysis of genes provides insights into the molecular and immune characteristics of mitochondria-related genes in atherosclerosis. Genomics. 2025;117(2):111013. doi:10.1016/j.ygeno.2025.111013
37. Zhou L, Wang Y, Wan Q, et al. A non-clinical comparative study of IL-23 antibodies in psoriasis. MAbs. 2021;13(1):1964420. doi:10.1080/19420862.2021.1964420
38. Huang R, Sun Y, Liu R, et al. ZeXieYin formula alleviates atherosclerosis by inhibiting the MAPK/NF-κB signaling pathway in APOE-/- mice to attenuate vascular inflammation and increase plaque stability. J Ethnopharmacol. 2024;327:117969. doi:10.1016/j.jep.2024.117969
39. Larousserie F, Bsiri L, Dumaine V, et al. Frontline Science: human bone cells as a source of IL-27 under inflammatory conditions: role of TLRs and cytokines. J Leukoc Biol. 2017;101(6):1289–1300. doi:10.1189/jlb.3HI0616-280R
40. Liu JQ, Zhang C, Zhang X, et al. Intratumoral delivery of IL-12 and IL-27 mRNA using lipid nanoparticles for cancer immunotherapy. J Control Release. 2022;345:306–313. doi:10.1016/j.jconrel.2022.03.021
41. McLean MH, Andrews C, Hanson ML, et al. Interleukin-27 is a potential rescue therapy for acute severe colitis through interleukin-10-dependent, T-cell-independent attenuation of colonic mucosal innate immune responses. Inflamm Bowel Dis. 2017;23(11):1983–1995. doi:10.1097/MIB.0000000000001274
42. Lin CH, Wu CJ, Cho S, et al. Selective IL-27 production by intestinal regulatory T cells permits gut-specific regulation of TH17 cell immunity. Nat Immunol. 2023;24(12):2108–2120. doi:10.1038/s41590-023-01667-y
43. Xia S, Wei J, Wang J, et al. A requirement of dendritic cell-derived interleukin-27 for the tumor infiltration of regulatory T cells. J Leukoc Biol. 2014;95(5):733–742. doi:10.1189/jlb.0713371
44. Gong H, Ma S, Chen J, et al. Dendritic cell-derived IL-27 p28 regulates T cell program in pathogenicity and alleviates acute graft-versus-host disease. Signal Transduct Target Ther. 2022;7(1):319. doi:10.1038/s41392-022-01147-z
45. Krishnamurthy P, Rajasingh J, Lambers E, et al. IL-10 inhibits inflammation and attenuates left ventricular remodeling after myocardial infarction via activation of STAT3 and suppression of HuR. Circ Res. 2009;104(2):e9–18. doi:10.1161/CIRCRESAHA.108.188243
46. Huang Y, Hu H, Liu L, et al. Interleukin-12p35 deficiency reverses the Th1/Th2 imbalance, aggravates the Th17/Treg imbalance, and ameliorates atherosclerosis in ApoE-/- Mice. Mediators Inflamm. 2019;2019:3152040. doi:10.1155/2019/3152040
47. Cai Y, Tang H, Tang W, et al. Changes in IL-27 and its effect on CD4 + T cells in patients with coronary artery disease. 2022. doi:10.21203/rs.3.rs-1964617/v1
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.