Back to Journals » International Journal of General Medicine » Volume 19
Integrating Bioinformatics and Experimental Validation Identify CCNA2 as a Novel Prognostic Biomarker and Tumor Promoter via the PI3K/AKT Pathway in Lung Adenocarcinoma
Authors Li JP, Zhang MY, Li R, Huo C, Qu JJ ![]() , Qu YQ
, Qu YQ ![]()
Received 1 October 2025
Accepted for publication 21 January 2026
Published 29 January 2026 Volume 2026:19 571529
DOI https://doi.org/10.2147/IJGM.S571529
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Ching-Hsien Chen
Jian-Ping Li, Meng-Yu Zhang, Rui Li, Chen Huo, Jia-Jia Qu, Yi-Qing Qu
Department of Pulmonary and Critical Care Medicine, Qilu Hospital of Shandong University, Jinan, 250012, People’s Republic of China
Correspondence: Yi-Qing Qu, Department of Pulmonary and Critical Care Medicine, Qilu Hospital of Shandong University, Wenhuaxi Road 107#, Jinan, 250012, People’s Republic of China, Tel +86 531 8216 9335, Fax +86 531 8296 7544, Email [email protected]
Purpose: To investigate the prognostic value of cell cyclin A2 (CCNA2) in lung adenocarcinoma (LUAD) and to explore its mechanisms in promoting cancer progression.
Patients and Methods: In this study, we employed an integrated strategy combining bioinformatics, clinical analysis and molecular biology to elucidate the role of CCNA2 in LUAD. First, comprehensive bioinformatics analyses were performed using public datasets. This included detecting the differential expression of CCNA2 in LUAD versus normal tissues, analyzing its correlation with patient survival and clinical characteristics, and employing Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes pathway (KEGG) analysis to predict the functions of CCNA2-associated genes. The relationship between CCNA2 expression and immune infiltration was further examined via the tumor immune estimation resource (TIMER) platform. The expression level of CCNA2 was also confirmed through reverse transcription-quantitative PCR and Western blotting. Additionally, the biological function of CCNA2 was evaluated by constructing an in vitro transfection model.
Results: The results of the present study indicated that CCNA2 was upregulated in LUAD tissues. Cox regression analysis revealed that CCNA2 upregulation is a independent prognostic biomarker for LUAD. Additionally, CCNA2 was markedly associated with immune cell infiltration and immune checkpoint molecules. The results of in vitro experiments confirmed that knockdown of CCNA2 significantly inhibited the proliferation, invasion and migration of H1975 and H1299 cells. Furthermore, CCNA2 was found to promote the invasion and migration of lung cancer cells through the PI3K/AKT signaling pathway.
Conclusion: The present research identified the prognostic signature and biological function of CCNA2 in LUAD, which suggested that CCNA2 may be a potential prognostic biomarker and a pivotal oncogenic driver for this disease.
Keywords: LUAD, CCNA2, predictive significance, biological function, PI3K/AKT
Introduction
Lung cancer (LC) has a high morbidity and cancer-related mortality rate globally.1 The incidence of LUAD has increased in recent years and accounts for 50% of deaths attributed to LC.2 In long-term clinical practice, LUAD is liable to metastasize at the early stage of disease, and some patients with LUAD may have already reached the advanced stage when diagnosed.3 Despite the advancements in immunotherapy and targeted therapy, the 5-year survival rate of patients with LUAD is only 22%.4,5 Hence, there is an urgent need to further explore the molecular mechanism of LUAD to enhance the survival and prognosis of patients.
The occurrence of LUAD is the outcome of the combined action of multiple factors. The study of the potential abnormal expression of genes can provide an important basis for the diagnosis, clinical prognosis and treatment choice of LUAD.6–8 CCNA2 is an essential cell cycle regulator that is involved in the regulation of mitosis.9 CCNA2 is also the core regulator of the hematopoietic process and the development of embryonic cells.10 Previous studies have shown that CCNA2 participates in tumor metastasis. For instance, Gopinathan et al11 revealed that the function of CDK2 and CCNA2 in maintaining cell cycle accuracy and regulating tumorigenesis. CCNA2 exhibits abnormal expression patterns in various tumor tissues, including breast, gastric and ovarian cancer tissues.12–14 CCNA2, regulated by microRNA-508-3p, is also involved in the proliferation and invasion of ovarian tumor cells.15 Furthermore, from the perspective of etiology, the high expression of CCNA2 in LUAD is specifically associated with smoking, which is the main environmental risk factor for this disease.16 A study found that CCNE1 can drive the occurrence of hepatocellular carcinoma by activating the PI3K/AKT pathway.17 This further supports the view that cyclins may cross-regulate classical signaling pathways. Another study revealed that CCNA2 is one of the core targets of curcumin’s anti-tumor effect, and the PI3K-AKT signaling pathway is a key pathway that is significantly enriched in this context.18 This indicates that there is a functional connection between CCNA2 and the PI3K/AKT pathway. The prognostic value of CCNA2 in LUAD as well as its potential molecular mechanisms in this context remain unknown. Our hypothesis that CCNA2 promotes LUAD progression via the PI3K/AKT pathway is further supported by the external analytical evidence, providing a broader rationale for investigating this specific mechanistic link in our study.
This study investigates the prognostic significance and molecular mechanisms of CCNA2 in LUAD, aiming to establish it as a reliable biomarker for predicting patient clinical outcomes.
Materials and Methods
Expression of CCNA2 in LUAD
The differential expression of CCNA2 between tumor and adjacent normal tissues was verified by analyzing RNA sequencing data from The Cancer Genome Atlas (TCGA, https://portal.gdc.cancer.gov/). To independently validate the mRNA expression findings, two publicly available microarray datasets from the Gene Expression Omnibus (GEO, https://www.ncbi.nlm.nih.gov/geo/) database were utilized: GSE10072 and GSE19804.
Prognostic Analysis
Prognostic analysis was performed to evaluate the association between CCNA2 expression and patient survival. Overall survival (OS) data, along with corresponding clinical annotations (including sex, pathological stage, tumor size, and nodal metastasis status), were extracted from the TCGA-LUAD cohort. The cohort was then divided into CCNA2-high and CCNA2-low expression groups based on the median expression value of CCNA2. Kaplan-Meier (KM) survival curves were generated and compared using the “survival” and “survminer” packages in R software version 4.3.2. To investigate the association between CCNA2 expression and prognosis with key clinical characteristics, a stratified analysis was performed. Patients were stratified by sex, pathological stage (including T, N and M stages), and CCNA2 expression levels were compared across each subgroup. To validate the prognostic significance of CCNA2 identified in the TCGA cohort, an independent validation dataset (GSE68465) was obtained from the GEO database. The same analytical method was applied to this external cohort to verify the robustness of our findings.
Gene Function and Pathway Enrichment Analysis
Using the GSE68465 dataset, we screened for CCNA2-associated genes and performed functional enrichment analysis. Firstly, to identify genes whose expression patterns were closely related to CCNA2, samples were divided into a CCNA2-high expression group and a CCNA2-low expression group according to the median expression value of CCNA2. Subsequently, differential expression analysis between the two groups was conducted using the limma package in R software (version 4.3.2). This yielded a set of differentially expressed genes associated with CCNA2 co-expression. To further explore the biological functions and pathways associated with this gene set, we performed a systematic functional enrichment analysis. GO enrichment analysis annotated gene function from three aspects: Biological Process, Cellular Component, and Molecular Function. KEGG pathway enrichment analysis was used to identify significantly enriched signaling pathways. Enrichment analysis plots were generated through the online data analysis platform available at http://www.bioinformatics.com.cn/.
The Correlation Between CCNA2 and Immune Characteristics
The relationship between CCNA2 expression and tumor immune microenvironment was investigated using TIMER platform (https://cistrome.shinyapps.io/timer/), a comprehensive web platform for systematic analysis of immune infiltrates across diverse cancer types. Within the TCGA-LUAD cohort, we applied the deconvolution algorithm embedded in TIMER to estimate the abundance of six immune cell populations: B cells, CD4+ T cells, CD8+ T cells, neutrophils and macrophages. Subsequently, Spearman’s rank correlation analysis was performed to evaluate the association between CCNA2 mRNA expression and the estimated infiltration levels of each immune cell type. Correlation strength and statistical significance (p-value) were reported, with a threshold of p<0.05 considered significant.
Cell Culture and Gene Transfection
The cell lines (H1975, H1299, A549 and 16HBE) used in the present study were obtained from Procell Life Science & Technology Co., Ltd. All cell lines were cultured in RPMI-1640 medium (Gibco) supplemented with 10% fetal bovine serum (FBS; SinSage) and 1% penicillin-streptomycin (Solarbio), under standard culture conditions (37°C, 5% CO2 in a humidified incubator). Cells were seeded in antibiotic-free growth medium at six well plate and allowed to adhere for 24h prior to transfection. When the cells had covered approximately 60–80% of the culture plate area, they were ready for transfection. Cells were harvested 48 h post-transfection for downstream analyses. CCNA2 small interfering RNA was purchased from Shanghai GenePharma Co., Ltd. Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.) was utilized to transfect the cells. The sequence of siRNA was as follows: CCAN2 (Sense: 5′-GUAGCAGAGUUUGUGUACATT-3′; Antisense: 5′- UGUACACAAACUCUGCUACTT-3′); Control (Sense: 5′-UUCUCCGAACGUGUCACGUTT-3′; Antisense: 5′-ACGUGACACGUUCGGAGAATT-3′). CCNA2 overexpression plasmid [Homo-CCNA2 (NM_001237.5) in PCDNA3.1] were obtained from BioSune Co., Ltd. For transfection with CCNA2 plasmid, the cells were treated with Lipofectamine 3000.
RNA Extraction and Reverse Transcription-Quantitative PCR (RT-qPCR)
Total RNA was extracted from the 16HBE, H1975, H1299 and A549 cell lines using TRIzol reagent. qPCR was conducted with SYBR Green Premix Pro Taq HS qPCR Kit [Accurate Biotechnology (Hunan) Co., Ltd.]. Thermal cycling was conducted on the Bio-Rad CFX Connect Real-Time System. GAPDH was used as the endogenous control and calculated by the 2−ΔΔCt method.19 The primers were purchased from BioSune Co., Ltd., the sequences of which were as follows: CCNA2 forward, 5′-TAAACAGCCTGCGTTCACCA-3′ and reverse, 5′-CAGGGCATCTTCACGCTCTA-3′; GAPDH forward, 5′-AAATCAAGTGGGGCGATGCT-3′ and reverse, 5′-CAAATGAGCCCCAGCCTTCT-3′.
Western Blotting
The protein content from cells was extracted with RIPA lysis buffer (Beyotime Institute of Biotechnology) and normalized after measuring the concentration using a BCA assay kit according to the provided instructions. SDS-PAGE (10%) was utilized to separate the total proteins, which were subsequently transferred onto nitrocellulose membranes. These membranes were then blocked with 5% skimmed milk for 60 min at room temperature before incubation with the CCNA2 (cat. no. ab181591; 1:2000 dilution; Abcam) and GAPDH (cat. no. 5174; 1:1000 dilution; Cell Signaling Technology, Inc.) primary antibodies overnight at 4°C. The membrane was then incubated with a rabbit secondary antibody (ZB-2301; 1:2000 dilution; Beijing Zhongshan Golden Bridge Biotechnology Co. Ltd.) for 1h at room temperature. Images of the results were collected using the ChemiDoc MP Imaging System (Bio-Rad Laboratories, Inc).
Cell Proliferation Assay
The EDU (5-ethynyl-2′-deoxyuridine) incorporation assay was performed to assess cellular proliferation. The cells were seeded into 96-well plates and incubated at 37°C with 5% CO2 for 24 h. Cultured cells were first incubated with EDU (50μM, RiboBio) diluted in complete growth medium for 2 hours at 37°C under 5% CO2. Following EDU exposure, cells were fixed with 4% paraformaldehyde for 15 minutes at room temperature, permeabilized with 0.5% Triton X-100 in PBS for 20 minutes, and incubated with a 1× Apollo reaction solution for 30 min in the dark. Cells were subsequently counterstained with 1× Hoechst 33342 (RiboBio) for DNA staining. Fluorescent images were acquired using a confocal microscope (ZEISS, axio observer7).
Cell Wound Healing Assay
H1975 and H1299 cell lines were seeded into 6-well dishes and cultured for a specific timeframe to achieve ~90% confluency. The confluent cells were then scratched using 200 µL pipette tips and washed with PBS three times. These cells were subsequently cultured in RPMI‑1640 medium (Gibco) without FBS for 48 h. Microscopic image (Olympus CKX53) of the dishes were captured at 0 h and the closure of the gap was assessed at 48 h.
Transwell Migration and Invasion Assays
For invasion assays, 8-μm pore transwell inserts (Corning) were coated with 50 μL growth factor-reduced Matrigel (Solarbio, 1:8 dilution in serum-free medium) and polymerized at 37°C for 1 h. Migration assays used uncoated inserts. A total of 3×10⁴ cells in 200 μL serum-free medium were seeded into upper chambers. Lower chambers contained 600 μL complete medium with 20% FBS as chemoattractant. Assembled plates were incubated at 37°C with 5% CO2 for 24 h. Cells were fixed in 4% paraformaldehyde (30 min) and then stained with 0.1% crystal violet (30min) at room temperature. Non-migratory/invasive cells were removed from upper chambers using cotton swabs. The chambers were placed on a slide and the migration of cells was observed under an inverted microscope.
Statistical Analysis
R software (version 4.3.2) was utilized for data processing and analysis. p<0.05 was considered to indicate a statistically significant difference. Student’s t tests were applied to continuous variables with normal distribution and equal variance, whereas Mann–Whitney U-test addressed non-normal data. When comparing the expression levels of CCNA2 between tumor and normal tissues, we employed independent samples t-tests for statistical analysis. For cases where both tumor and adjacent normal tissues were obtained from the same patient, paired-sample t-tests were used. Data involving comparisons across the three or more groups were analyzed using one-way ANOVA or the Kruskal–Wallis test. One-way ANOVA followed by Tukey’s post-hoc test was applied to normally distributed data with equal variances. For data that violated the assumption of normality or homogeneity of variances, the non-parametric Kruskal–Wallis test was performed, followed by Dunn’s post-hoc test for pairwise comparisons. Survival analysis was conducted using the KM method, and the survival curve was evaluated using the Log rank test. In cases where survival curves intersect, the Renyi test was applied for statistical comparison.20
Results
CCNA2 Expression in LUAD
To determine the differences in CCNA2 expression in LUAD, the CCNA2 mRNA levels were analyzed using data from TCGA database. The data from 52 matched pairs of LUAD and adjacent normal tissues were chosen for this analysis. The results indicated that CCNA2 was highly expressed in LUAD tissues (Figure 1a). Furthermore, similar results were observed in GSE10072 and GSE19804 datasets (Figure 1b and c). The GSE10072 dataset includes the expression profiling of 58 LUAD tissues and 49 normal lung tissues. The GSE19804 dataset originates from a transcriptional profiling of female lung cancer patients without smoking, encompassing 60 lung cancer tissue samples and 60 normal adjacent tissue samples (Table 1). The diagnostic ability of CCNA2 to distinguish tumor and normal samples was assessed using receiver operating characteristic curves. Notably, the area under the curve values were 0.970 (TCGA), 0.905 (GSE10072) and 0.833 (GSE19804) (Figure 1d–f).
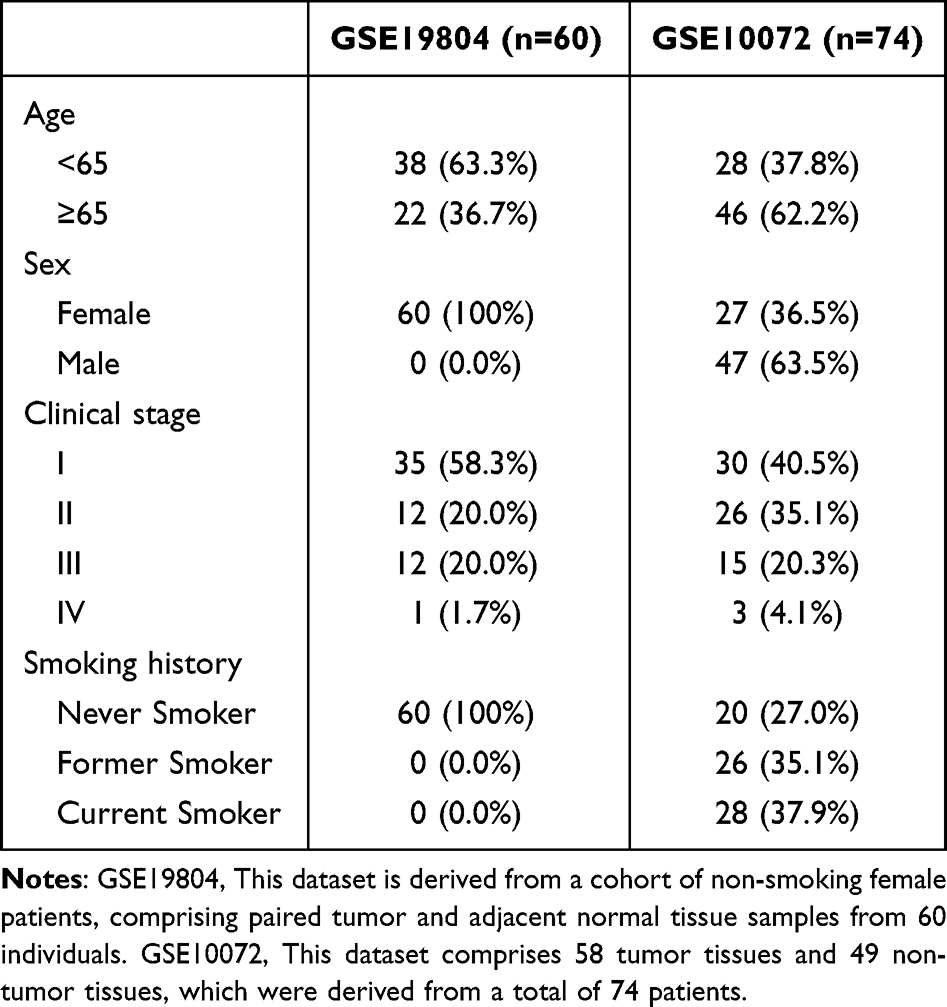 |
Table 1 Patient Demographic and Clinical Characteristics of GSE19804 and GSE10072 |
 |
Figure 1 The expression levels of CCNA2 in LUAD. (a) The CCNA2 mRNA expression between paired LUAD tumor tissues and adjacent normal tissues from the TCGA-LUAD dataset. Box plot comparing CCNA2 expression in normal and tumor tissues based on (b) GSE10072 dataset and (c) GSE19804 dataset. ROC curve of CCNA2 in LUAD based on TCGA-LUAD dataset (d), GSE10072 dataset (e), GSE19804 dataset (f). X-axis indicates false-positive rates, and Y-axis indicates true-positive rates. ***p< 0.001. Statistical significance was determined by paired Student’s t-test (Figure 2a) and unpaired Student’s t-test (Figure 2b and 2c). Abbreviations: LUAD, lung adenocarcinoma; LC, lung cancer; GSE10072, GSE19804, dataset from GEO. |
Association of CCNA2 Expression and Clinical Parameters
The demographic and clinical characteristics of patients from the TCGA-LUAD and GSE68465 cohorts are presented in Table 2. Analysis of data from TCGA database showed a significant association between the CCNA2 expression level and sex, clinical stage, tumor (T) classification and node (N) classification. However, there was no association between CCNA2 expression and the metastasis stage of patients with LUAD (Figure 2a). Additionally, the analysis of data from the GSE68465 cohort indicated a link between higher CCNA2 expression and male sex, T classification and N classification. Higher CCNA2 levels were also observed in those patients with LUAD who were smokers (Figure 2b). We analyzed the relationship between the mutations of common driver genes in LUAD and the expression of CCNA2 based on TCGA data. We observed that patients with ALK mutations exhibited higher expression levels of CCNA2. However, no significant differences were observed between the wild-type and mutant forms of EGFR and KRAS. Furthermore, for the key cell cycle gene CDKN2A, there was no difference in the expression of CCNA2 between the mutant and wild-type forms (Figure 2c).
 |
Table 2 Patient Demographic and Clinical Characteristics of TCGA-LUAD and GSE68465 Cohort |
 |
Figure 2 Association of CCNA2 expression with clinicopathologic parameters. (a) The relationship between CCNA2 expression and clinicopathologic parameters in TCGA-LUAD dataset. (b) The relationship between CCNA2 expression and clinicopathologic parameters in GSE68465 dataset. (c) CCNA2 expression according to the mutation of EGFR, KRAS, ALK and CDKN2A. *p < 0.05. **p< 0.01. ***p < 0.001. Statistical significance was determined by student’s t test and one-way ANOVA. Abbreviations: ns, no significance; T, Tumor; N, Node; M, Metastasis. |
Prognostic Performance of CCNA2 in LUAD
The patient survival status and gene expression of CCNA2 in LUAD is shown in Figure 3a and b. The results indicated that patients with higher CCNA2 expression had poorer outcomes. Survival analysis was further performed using LUAD data from TCGA database. As is shown in Figure 3c, The KM survival curve showed that patients with low CCNA2 expression had a significantly better overall survival. Furthermore, LUAD data acquired from the GSE68465 dataset in the GEO database were applied to verify the survival analysis results (Figure 3d). Univariable and multivariable Cox proportional hazards regression analyses indicted that CCNA2 was an independent risk factor for patients with LUAD (Figure 3e and f).
 |
Figure 3 Survival analysis of CCNA2. Distribution of the survival status and CCNA2 expression in (a) TCGA-LUAD dataset and (b) GSE68465 dataset. Correlation between CCNA2 expression and OS of LUAD in (c) TCGA-LUAD dataset and (d) GSE68465 dataset. Univariate cox regression analysis (e) and multivariate cox regression analysis (f) in the GSE68465 dataset. Statistical significance was determined by Log rank test (Figure 4c) and Renyi test (Figure 4d). Abbreviation: OS, overall survival. |
Functional Enrichment Analysis of CCNA2
As shown in the volcano plot, 454 downregulated and 468 upregulated genes were screened (|log Fold Change|>0.5, adjust p<0.05; Figure 4a). The heatmap in Figure 4b shows the top 50 genes. This result illustrated that multiple differentially expressed genes were associated with the CCNA2. By applying the Betweenness algorithms, the principal genes were identified through evaluating the connection degree of each node within the protein-protein interaction network (Figure 4c). Analysis of the GO functions revealed that CCNA2 may play a significant role in cellular processes, specifically in cell division activities such as “DNA replication”, “Mitotic nuclear division” and “DNA-dependent ATPase activity” (Figure 4d). KEGG pathway analysis demonstrated that the genes co-expressed with CCNA2 are mainly linked to the “Cell cycle”, “DNA replication processes” and “the IL-17 signaling pathway” (Figure 4e).
 |
Figure 4 Functional analysis of CCNA2. (a) The volcano plot of CCNA2-related genes. (b) The heatmap shows the top 50 genes related to CCNA2. (c) Protein–protein interaction network of CCNA2-related genes. (d) GO analysis of CCNA2-related genes. (e) KEGG analysis of CCNA2-related genes. Abbreviations: BP, Biological Process; CC, Cellular Component; MF, Molecular Function. |
Immune Signatures of CCNA2 in LUAD
As depicted in Figure 5a, CCNA2 exhibited an inverse relationship with the levels of CD4+ T cells and B cells but exhibited a positive relationship with CD8+ T cells, neutrophils and macrophages. Concurrently, CCNA2 expression was positively correlated with key immune checkpoint molecules such as CTLA-4, PD-L1 (PDCD1), TIM-3 (Havcr2), LAG3 and TIGIT (Figure 5b).
 |
Figure 5 The correlation between CCNA2 expression and tumor immune microenvironment using the TIMER database. (a) Correlation between CCNA2 expression and immune cells. (b) Correlation between CCNA2 expression and major immune checkpoints. cor, spearman’s rank correlation coefficient. partial.cor, partial spearman’s rank correlation coefficient. p, p value. |
Relationship Between CCNA2 and LUAD Cell Function
RT-qPCR and Western blotting verified that CCNA2 was upregulated in LUAD cells (Figure 6a and b). To investigate the effects of CCNA2 expression on LUAD cell function, CCNA2 was knocked down in H1299 and H1975 cells (Figure 6c and d). The results of the EdU proliferation assays indicated that reducing CCNA2 expression led to a decrease in the proliferation rate of H1975 cells (Figure 6e). The results from the cell wound healing and transwell assays confirmed that the migration ability of H1975 and H1299 cells was affected by CCNA2. Specifically, the migratory ability of cells with CCNA2 knockdown was suppressed (Figure 6f–i).
 |
Figure 6 CCNA2 regulates LUAD cell proliferation, migration and invasion. (a) The mRNA level of CCNA2 in LUAD cell and normal cell based on RT-qPCR. (b) The protein level of CCNA2 in LUAD cell and normal cell based on Western blotting. (c) The mRNA level of knockdown efficiency of CCNA2 in H1975 and H1299 cell lines. (d)The protein level of knockdown efficiency of CCNA2 in H1975 and H1299 cell lines. (e) EDU assay detects the cell proliferation ability. (f and g) Scratch wound-healing assays were utilized to compare the distance of cell migration between si-CCNA2 group and control group at 0h and 48h after scratching. (h and i) Transwell assay demonstrating changes in the number of cells invaded after knockdown of CCNA2. *p<0.05. ***p<0.001. Statistical significance was determined by Mann–Whitney U-test. |
CCNA2 Promote LUAD Cell Migration and Invasion by Activating the PI3K/AKT Signaling Pathway
To investigate the potential molecular mechanisms underlying CCNA2-mediated LUAD development, we examined the effect of CCNA2 on the PI3K/AKT signaling pathway using Western blotting. The results showed that overexpression of CCNA2 significantly activated the PI3K/AKT signaling pathway in H1299 cells. In contrast, downregulation of CCNA2 expression reduced the levels of phosphorylated PI3K (p-PI3K) and phosphorylated AKT (p-AKT) (Figure 7a–c). Furthermore, treatment with the PI3K/AKT signaling pathway inhibitor LY294002 markedly abolished CCNA2-mediated migration and invasion of H1299 cells (Figure 7d and e). This suggest that CCNA2 may facilitate the invasion and migration of LUAD cells by modulating the PI3K/AKT signaling pathway.
 |
Figure 7 CCNA2 potentially regulates migration and invasion of H1299 cell by activating the PI3K/AKT signaling pathway. (a) WB detected the protein expression in H1299 cell with CCNA2 overexpression. (b) WB showing the expression of PI3K/AKT pathway related proteins in H1299 cell with CCNA2 overexpression. (c) WB showing the expression of PI3K/AKT pathway related proteins in H1299 cell with CCNA2 knockdown. (d) The expression of PI3K/AKT pathway related proteins in H1299 cell with CCNA2 overexpression and LY294002 treatment. (e) Transwell assay showing the migration and invasion potential of CCNA2 over expressed H1299 cells after treated with LY294002. *p < 0.05. **p< 0.01. ***p < 0.001. Statistical significance was determined by Mann–Whitney U-test. |
Discussion
In this study, we found that the expression of CCNA2 in LUAD was significantly upregulated compared to normal tissues. Studies found that the expression of CCNA2 increased in both colorectal cancer and liver cancer tissues, which is consistent with our research results.21,22 This indicates that CCNA2 may play a role in promoting cancer during the tumor formation process. The increase in its expression might be a common molecular event in the development of various malignant tumors. We analyzed the relationship between CCNA2 expression and the clinical and pathological characteristics of LUAD patients. The results showed that the expression of CCNA2 was elevated in male patients and those who were smokers. Smoking induces lung cancer by influencing epigenetics and the tumor microenvironment.23 He et al discovered that smoking induces the expression of CCNA2, thereby promoting the differentiation of AT2 cells. This may be the mechanism by which smoking causes lung cancer.16 Furthermore, The expression level of CCNA2 was significantly higher in advanced lung adenocarcinoma. Further research revealed a significant correlation between CCNA2 and lymph node metastasis (p < 0.001). This suggests that CCNA2 may directly participate in the invasion and metastasis process of LUAD. This study also found that in patients with LUAD who have ALK gene mutations, the expression of CCNA2 is elevated. ALK gene mutations can lead to abnormal activation of the downstream signaling pathways of ALK, thereby promoting cell cycle regulation, proliferation and apoptosis.24 CCNA2 regulates the transition of cells from G1 phase to S phase of the cell cycle and promotes DNA replication, thereby participating in the process of tumor proliferation.25,26 This suggests that the high expression of CCNA2 observed in this study may be one of the important molecular events driving the cell cycle disorder in LUAD cells driven by ALK mutations. This study further suggests that CCNA2 may serve as an auxiliary marker for evaluating the prognosis of ALK-mutated LUAD patients, and also provides new ideas for reversing drug resistance.
Exploration of the tumor immune microenvironment has become a research hotspot in tumor diagnosis and treatment.27,28 Studying the LUAD tumor immune microenvironment is important for exploring pathogenesis, formulating a diagnosis and treatment plan as well as for evaluating the prognosis of patients. There are numerous types of cells involved in immune cell infiltration in NSCLC, including natural killer cells, T lymphocytes, macrophages, dendritic cells and B lymphocytes.29 A number of scholars acknowledge that the prognostic model of LUAD is correlated with immune cell infiltration.30–32 The results of the present study showed that CCNA2 was negatively correlated with the infiltration degree of B lymphocytes and CD4+ T lymphocytes as well as positively correlated with the infiltration degree of CD8+ T lymphocytes, neutrophils and macrophages. In addition, the expression of CCNA2 in LUAD was associated with the expression of immune checkpoints such as CTLA-4, PD1 (PDCD1), TIM3 (HAVCR2), LAG3 and TIGIT. These results suggest that CCNA2 may be involved in the recruitment and activation of immune cells.
The basic characteristic of tumor cells is deregulated proliferation and metastasis.33,34 Abnormal cell cycle progression is an important mechanism in tumorigenesis.35,36 Yao et al37 reported that Cyclin K promotes tumorigenesis and radiation resistance through the β-catenin/Cyclin D1 axis. Another study has illustrated that CCNA1 could act as a diagnostic marker for papillary thyroid cancer.38 Cyclin Y regulates the proliferation, invasion and metastasis of HCC cells.39 Additionally, knockdown of CCNA2 significantly inhibits proliferation and migration, while promoting the apoptosis of HCC cells.22 Regarding the treatment of LC, Wang et al40 discovered that metformin reduces the expression of CCNA2 in LUAD cells and that the downregulation of CCNA2 promotes the antitumor activity of metformin. Another study revealed that RSV suppresses LUAD progression and enhances sensitivity to chemotherapy by downregulating CCNA2.41 In the present study, it was found that CCNA2 knockdown could inhibit the proliferation, invasion and migration of LUAD cells, which may lay the foundation for further study on the malignant progression of LUAD. Previous study identified that LINC00665-miR-let-7b-CCNA2 as the key ceRNA network associated with the progression of LUAD.42 However, they did not fully elucidate the precise molecular mechanisms. In this research, we found that CCNA2 contributes to the tumorigenesis and progression of LUAD by activating the PI3K/AKT signaling pathway, suggesting its potential as a promising therapeutic target for LUAD, which enhanced the novelty of this study. However, this study has several limitations. TCGA and GEO datasets may introduce inherent selection biases, and multicenter cohorts would strengthen clinical generalizability. Secondly, further research is necessary to reveal the relationship between CCNA2 expression and drug treatment.
Conclusion
CCNA2 is upregulated in LUAD and may have an important role in the prognosis of this disease. The results of in vitro experiments confirmed that CCNA2 promoted the progression of LUAD cells. Notably, these findings highlight the potential therapeutic significance of CCNA2. As a key molecule driving LUAD progression, CCNA2 may serve as a promising therapeutic target, particularly for ALK-mutant LUAD patients who are prone to drug resistance. This not only provides new perspectives for the development of individualized treatment in the future but also lays a foundation for exploring targeted combination therapy strategies to improve therapeutic efficacy.
Abbreviations
LUAD, Lung adenocarcinoma; TCGA, The cancer genome atlas program; GEO, Gene expression omnibus; TIME, Tumor immune to assess resource; GO, Gene ontology; KEGG, Kyoto encyclopedia of genes and genomes.
Ethics Approval and Informed Consent
This study is exempt from ethical review as it utilizes publicly available, de-identified data from the TCGA and GEO databases, in accordance with item 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects (National Health Commission of China, effective February 18, 2023).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (grant no. 82173173).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Peng W, Li B, Li J, et al. Clinical and genomic features of Chinese lung cancer patients with germline mutations. Nat Commun. 2022;13(1):1268. doi:10.1038/s41467-022-28840-5
2. Yu B, Li T, Chen J, et al. Identification of activated pathways in lung adenocarcinoma based on network strategy. J Cancer Res Ther. 2020;16(4):793–16. doi:10.4103/0973-1482.199458
3. Fang H, Sun Q, Zhou J, et al. m(6)A methylation reader IGF2BP2 activates endothelial cells to promote angiogenesis and metastasis of lung adenocarcinoma. Mol Cancer. 2023;22(1):99. doi:10.1186/s12943-023-01791-1
4. Wang XY, Zhao YF, Yang L, Liu Y, Yang YK, Wu N. Correlation analysis between metabolic tumor burden measured by positron emission tomography/computed tomography and the 2015 World Health Organization classification of lung adenocarcinoma, with a risk prediction model of tumor spread through air spaces. Transl Cancer Res. 2020;9(10):6412–6422. doi:10.21037/tcr-20-1934
5. Chen Y, Li E, Chang Z, et al. Identifying potential therapeutic targets in lung adenocarcinoma: a multi-omics approach integrating bulk and single-cell RNA sequencing with mendelian randomization. Front Pharmacol. 2024;15:1433147. doi:10.3389/fphar.2024.1433147
6. He L, Chen J, Xu F, Li J, Li J. Prognostic implication of a metabolism-associated gene signature in lung adenocarcinoma. Mol Ther Oncolytics. 2020;19:265–277. doi:10.1016/j.omto.2020.09.011
7. Wang W, Ren S, Wang Z, Zhang C, Huang J. Increased expression of TTC21A in lung adenocarcinoma infers favorable prognosis and high immune infiltrating level. Int Immunopharmacol. 2020;78:106077. doi:10.1016/j.intimp.2019.106077
8. Yu L, Cao S, Li J, Han B, Zhong H, Zhong R. Prognostic value and immune infiltration of a novel stromal/immune score-related P2RY12 in lung adenocarcinoma microenvironment. Int Immunopharmacol. 2021;98:107734. doi:10.1016/j.intimp.2021.107734
9. Bendris N, Loukil A, Cheung C, et al. Cyclin A2: a genuine cell cycle regulator? Biomol Concepts. 2012;3(6):535–543. doi:10.1515/bmc-2012-0027
10. Arsic N, Bendris N, Peter M, et al. A novel function for Cyclin A2: control of cell invasion via RhoA signaling. J Cell Biol. 2012;196(1):147–162. doi:10.1083/jcb.201102085
11. Gopinathan L, Tan SL, Padmakumar VC, Coppola V, Tessarollo L, Kaldis P. Loss of Cdk2 and cyclin A2 impairs cell proliferation and tumorigenesis. Cancer Res. 2014;74(14):3870–3879. doi:10.1158/0008-5472.CAN-13-3440
12. Gao T, Han Y, Yu L, Ao S, Li ZY, Ji JF. CCNA2 is a prognostic biomarker for ER plus breast cancer and tamoxifen resistance. PLoS One. 2014;9(3):e91771.
13. Yamamoto S, Tsuda H, Miyai K, Takano M, Tamai S, Matsubara O. Cumulative alterations of p27-related cell-cycle regulators in the development of endometriosis-associated ovarian clear cell adenocarcinoma. Histopathology. 2010;56(6):740–749. doi:10.1111/j.1365-2559.2010.03551.x
14. Lee YJ, Lee CE, Oh S, et al. Pharmacogenomic analysis reveals CCNA2 as a predictive biomarker of sensitivity to polo-like kinase I Inhibitor in gastric cancer. Cancers. 2020;12(6):1418.
15. Guo F, Zhang K, Li MY, et al. miR-508-3p suppresses the development of ovarian carcinoma by targeting CCNA2 and MMP7. Int J Oncol. 2020;57(1):264–276.
16. He Q, Qu M, Xu C, et al. Smoking-induced CCNA2 expression promotes lung adenocarcinoma tumorigenesis by boosting AT2/AT2-like cell differentiation. Cancer Lett. 2024;592:216922. doi:10.1016/j.canlet.2024.216922
17. Zhang K, Hu X, Yao L, Guo W. CCNE1 promotes the progression of hepatic precancerous lesion and the malignant phenotype of hepatocellular carcinoma. J Clin Transl Hepatol. 2025;13(7):555–567. doi:10.14218/JCTH.2024.00428
18. He Q, Liu C, Wang X, et al. Exploring the mechanism of curcumin in the treatment of colon cancer based on network pharmacology and molecular docking. Front Pharmacol. 2023;14:1102581. doi:10.3389/fphar.2023.1102581
19. Li P, Mi Q, Yan S, et al. Characterization of circSCL38A1 as a novel oncogene in bladder cancer via targeting ILF3/TGF-beta2 signaling axis. Cell Death Dis. 2023;14(1):59. doi:10.1038/s41419-023-05598-2
20. Li H, Han D, Hou Y, Chen H, Chen Z. Statistical inference methods for two crossing survival curves: a comparison of methods. PLoS One. 2015;10(1):e0116774. doi:10.1371/journal.pone.0116774
21. Lei X, Qiu L, Chen Q, et al. Exploring the regulatory mechanism of CCNA2 in colorectal cancer: insights from multiomics and experimental analysis. J Biol Chem. 2025;301(8):110216. doi:10.1016/j.jbc.2025.110216
22. Wang T, Li W, Wu Y, et al. Construction of a prognostic model based on disulfidptosis-related genes and identification of CCNA2 as a novel biomarker for hepatocellular carcinoma. Biol Direct. 2024;19(1):128. doi:10.1186/s13062-024-00569-9
23. Desrichard A, Kuo F, Chowell D, et al. Tobacco smoking-associated alterations in the immune microenvironment of squamous cell carcinomas. J Natl Cancer Inst. 2018;110(12):1386–1392. doi:10.1093/jnci/djy060
24. He P, Li H, Yang Z, et al. Discovery and preclinical evaluations of drug candidate DA-0157 capable of overcoming EGFR drug-resistant mutation C797S and EGFR/ALK co-mutations. Eur J Med Chem. 2025;287:117323. doi:10.1016/j.ejmech.2025.117323
25. Blanchard JM. Cyclin A2 transcriptional regulation: modulation of cell cycle control at the G1/S transition by peripheral cues. Biochem Pharmacol. 2000;60(8):1179–1184. doi:10.1016/S0006-2952(00)00384-1
26. Hegarat N, Crncec A, Suarez Peredo Rodriguez MF, et al. Cyclin A triggers Mitosis either via the Greatwall kinase pathway or Cyclin B. EMBO J. 2020;39(11):e104419. doi:10.15252/embj.2020104419
27. Altorki NK, Markowitz GJ, Gao D, et al. The lung microenvironment: an important regulator of tumour growth and metastasis. Nat Rev Cancer. 2019;19(1):9–31. doi:10.1038/s41568-018-0081-9
28. Fu T, Dai LJ, Wu SY, et al. Spatial architecture of the immune microenvironment orchestrates tumor immunity and therapeutic response. J Hematol Oncol. 2021;14(1):98. doi:10.1186/s13045-021-01103-4
29. Zheng X, Hu Y, Yao C. The paradoxical role of tumor-infiltrating immune cells in lung cancer. Intractable Rare Dis Res. 2017;6(4):234–241. doi:10.5582/irdr.2017.01059
30. Liu X-S, Zhou L-M, Yuan -L-L, et al. NPM1 is a prognostic biomarker involved in immune infiltration of lung adenocarcinoma and associated with m6A modification and glycolysis. Front Immunol. 2021;12:724741.
31. Yu Y, Wang Z, Zheng Q, Li J. GREB1L overexpression correlates with prognosis and immune cell infiltration in lung adenocarcinoma. Sci Rep. 2021;11(1):13281. doi:10.1038/s41598-021-92695-x
32. Zheng Y, Tian H, Zhou Z, et al. A novel immune-related prognostic model for response to immunotherapy and survival in patients with lung adenocarcinoma. Front Cell Dev Biol. 2021;9:651406. doi:10.3389/fcell.2021.651406
33. Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis in cancer. Nature. 2001;411(6835):342–348. doi:10.1038/35077213
34. Sun M, Angelillo J, Hugues S. Lymphatic transport in anti-tumor immunity and metastasis. J Exp Med. 2025;222(3):e20231954.
35. Wang L, Xu W, Mei Y, et al. CHRDL2 promotes cell proliferation by activating the YAP/TAZ signaling pathway in gastric cancer. Free Radic Biol Med. 2022;193:158–170. doi:10.1016/j.freeradbiomed.2022.09.006
36. Collins I, Garrett MD. Targeting the cell division cycle in cancer: CDK and cell cycle checkpoint kinase inhibitors. Curr Opin Pharmacol. 2005;5(4):366–373. doi:10.1016/j.coph.2005.04.009
37. Yao G, Tang J, Yang X, et al. Cyclin K interacts with beta-catenin to induce Cyclin D1 expression and facilitates tumorigenesis and radioresistance in lung cancer. Theranostics. 2020;10(24):11144–11158. doi:10.7150/thno.42578
38. da Silva RM, Santos JN, Uno M, et al. CCNA1 gene as a potential diagnostic marker in papillary thyroid cancer. Acta Histochem. 2020;122(8):151635. doi:10.1016/j.acthis.2020.151635
39. Shi K, Ru Q, Zhang C, Huang J. Cyclin Y modulates the proliferation, invasion, and metastasis of hepatocellular carcinoma cells. Med Sci Monit. 2018;24:1642–1653. doi:10.12659/MSM.906075
40. Wang L, Huang Y, Tian M, et al. Metformin inhibits the development of lung adenocarcinoma by regulating the expression of CCNA2 via E2F1. Comb Chem High Throughput Screen. 2025. doi:10.2174/0113862073348968241101112455
41. Tan XD, Luo CF, Liang SY. Antihyperlipidemic drug rosuvastatin suppressed tumor progression and potentiated chemosensitivity by downregulating CCNA2 in lung adenocarcinoma. J Chemother. 2024;36(8):662–674. doi:10.1080/1120009X.2024.2308975
42. Huang Y, Zhong L, Nie K, et al. Identification of LINC00665-miR-let-7b-CCNA2 competing endogenous RNA network associated with prognosis of lung adenocarcinoma. Sci Rep. 2021;11(1):4434. doi:10.1038/s41598-020-80662-x
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.