Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 13
Insulin-Like Proteins in Plant Sources: A Systematic Review
Authors Costa IS, Medeiros AF, Piuvezam G, Medeiros GCBS ![]() , Maciel BLL, Morais AHA
, Maciel BLL, Morais AHA ![]()
Received 4 April 2020
Accepted for publication 14 July 2020
Published 30 September 2020 Volume 2020:13 Pages 3421—3431
DOI https://doi.org/10.2147/DMSO.S256883
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Izael S Costa,1,* Amanda F Medeiros,1,* Grasiela Piuvezam,2,3 Gidyenne CBS Medeiros,3,4 Bruna LL Maciel,4,5 Ana Heloneida A Morais1,4,5
1Biochemistry Postgraduate Program, Biosciences Center, Federal University of Rio Grande do Norte, Natal, RN, Brazil; 2Department of Collective Health (DSC), Federal University of Rio Grande do Norte, Natal, RN, Brazil; 3Collective Health Postgraduate Program (PPGSCoL), Center for Health Sciences, Federal University of Rio Grande do Norte, Natal, RN, Brazil; 4Department of Nutrition, Center for Health Sciences, Federal University of Rio Grande do Norte, Natal, RN, Brazil; 5Nutrition Postgraduate Program, Center for Health Sciences, Federal University of Rio Grande do Norte, Natal, RN, Brazil
*These authors contributed equally to this work
Correspondence: Ana Heloneida A Morais
Department of Nutrition, Center for Health Sciences, Federal University of Rio Grande do Norte, Natal, RN, Brazil
Tel +55 84 991061887
Email [email protected]
Abstract: Type 2 diabetes mellitus (T2DM) is characterized by hyperglycemia. Proteins in plant sources that enable the maintenance of the glycemic profile may be of interest in the context of T2DM. However, their mechanisms of action are unclear, unlike other bioactive compounds. This systematic review identified and described the mechanisms of action of isolated and purified proteins and peptides extracted from vegetables on the reduction of blood glucose in T2DM in experimental studies. The research was done in PubMed, ScienceDirect, Scopus, Web of Science, Embase and Virtual Health Library (VHL) databases in March 2019. The initial search retrieved 916 articles, and, after reading the title, abstract and keywords, 24 articles were eligible for full reading. Then, five articles were eligible to build this systematic review. The evaluation of the evidence and the strength of the recommendations of the studies was evaluated with the SYstematic Review Center for Laboratory animal Experimentation - SYRCLE. Studies with proteins or peptides extracted from soybean (Glycine max), corn (Zea mays), peas (Pisum sativum), costus (Costus igneus) and ginseng (Panax ginseng) were found, and all of them decreased glycemia but not by the same mechanisms. The mechanism of action of proteins extracted from Glycine max, Pisum sativum, Costus igneus were similar, acting in the insulin-mediated pathways. The peptide derived from Zea mays increased GLP-1 expression, and the peptide from Panax ginseng reduced NF-kB signaling, both resulting in stimulating the release of insulin. Therefore, bioactive proteins and peptides of plant sources act through biochemical pathways, in the modulation of insulin resistance and the hyperglycemic state. These compounds are promising in scientific research on T2DM, because there is a probable similarity of these proteins with insulin, which enables them to act as insulin-like molecules.
Keywords: hyperglycemia, hyperinsulinemia, bioactive proteins, plant proteins, plant peptides, hypoglycemic agent
Introduction
Diabetes mellitus (DM) is a metabolic disorder characterized by a state of hyperglycemia. Among the types of DM, the most prevalent is type 2 diabetes mellitus (T2DM), occurring in about 95% of diabetes cases worldwide, with obesity being the most relevant risk factor for the clinical onset of the disease, with an excess of adipose tissue triggering the insulin resistance process.1
After meals, insulin concentrations increase in circulation. This hormone, produced by β-pancreatic cells, is largely responsible for regulating the entry of glucose into muscle and liver cells.2 This increase occurs because, after the insulin binds to its membrane receptor (insulin receptor or IR), there is a conformational change in the structure of the IR leading to its self phosphorylation in tyrosine residues, this process allows the signal transduction inside the cell. After this process, insulin receptor substrates 1 and 2 (IRS-1 and IRS-2) are also phosphorylated in tyrosine.2 Then, phosphatidylinositol-3 kinase (PI3-K) is associated with IRSs leading to phosphorylation of phosphatidylinositol bisphosphate (PIP2) to generate phosphatidylinositol triphosphate (PIP3). These phosphorylations allow the phosphatidylinositol dependent kinase (PDK) to be phosphorylated. Consequently, the distal molecule of the insulin pathway, the protein kinase B/Akt (Akt) is phosphorylated in the serine/threonine residues and this promotes an important signal for the translocation of the type 4 glucose transporter (GLUT-4) from sarcoplasm to sarcolemma, taking up glucose from the extracellular medium to the intracellular medium.3
Insulin, a hormone composed of two polypeptide chains containing 51 amino acids, acts primarily on peripheral tissues, mainly in muscles and liver.4 The mediator of insulin action is its receptors, Insulin Receptor Substrate (IRS), which are a protein family, and IRS-1 was the first member to be identified. These proteins trigger a signaling complex and one of the most studied involves the amplification of phosphoinositide 3-kinase (PI3K), which activates protein kinase B - AKT and culminates, among several aspects, in glycolysis and gluconeogenesis. This activation directly implicates in glucose homeostasis through glucose uptake through the cell membrane by glucose transporter (GLUT).5
Despite the detailed knowledge of the mechanism of action of insulin,2,3 and how most hypoglycemic agents act,4 the search for molecules with insulin-like activity is incessant. This effort is justified considering the effectiveness of this peptide, insulin, in the process of regulating blood glucose. It is known that the use of synthetic insulin has been a game-changer in the treatment of both T1DM and T2DM.6 However, looking for molecules or compounds with insulin-like characteristics, natural, mainly of plant origin, undoubtedly also assume a prominent role in this search for new therapies. Notably, in an attempt to minimize the state of insulin resistance and consequently control T2DM and also T1DM.
The metabolic state of insulin resistance causes the initial state of hyperglycemia, which can culminate in T2DM. Insulin resistance causes a reduction in glucose tolerance, and if this condition persists, it creates a decrease in pancreatic β cell mass, and culminates in the establishment of T2DM.7
T2DM is associated with several comorbidities such as cardiovascular diseases,7 severe inflammatory state,8 neoplasms9,10, retinopathy and nephropathy.11 Thus, individuals affected with T2DM need constant glycemic control and, for this reason, researches have sought therapeutic agents that enable the maintenance of the glycemic profile, especially those of natural origin. Several studies12–15 show the effect of vegetables on some metabolic disorders. However, little has been published on the mechanisms of action that involve control in the insulin resistance process and most of the publications were done in experimental models. Besides, most of these studies evaluated plant extracts, which present several components. This variety of compounds present in these extracts comprises the specification of which molecules perform the biological activity.8,11
Among bioactive compounds, proteins and peptides, of plant sources, have received attention from the scientific community concerning research on their hypoglycemic potential. Their mechanisms of action on glycemic control have not been appropriately explained, and few studies defined protein structures, with their respective defined amino acid sequences and conformation.12,13
This systematic review identifies and describes the action of isolated or purified proteins and peptides extracted from vegetables on the control of glycemia in T2DM in experimental studies. Also, this work analyzes the potential of these compounds as insulin-like proteins.
Methodology
The construction of this systematic review followed the methodological criteria established by the Preferred Reporting Items for Systematic Reviews and Meta-Analyzes Protocols (PRISMA-P),14 and the guidelines of the Systematic Review Centre for Laboratory Animal Experimentation – SYRCLE.15
The protocol for the construction of the systematic review was registered in the International Prospective Register of Ongoing Systematic Reviews - PROSPERO, under the number: CRD42019110956, and was based on the protocol described by de Medeiros et al.16
Search Strategies
The articles were accessed through electronic searches in March 2019, in the databases: PubMed; ScienceDirect; Scopus; Web of Science; Embase and Virtual Health Library (VHL). Manual searches were performed, and the search strategies are described in Table 1.
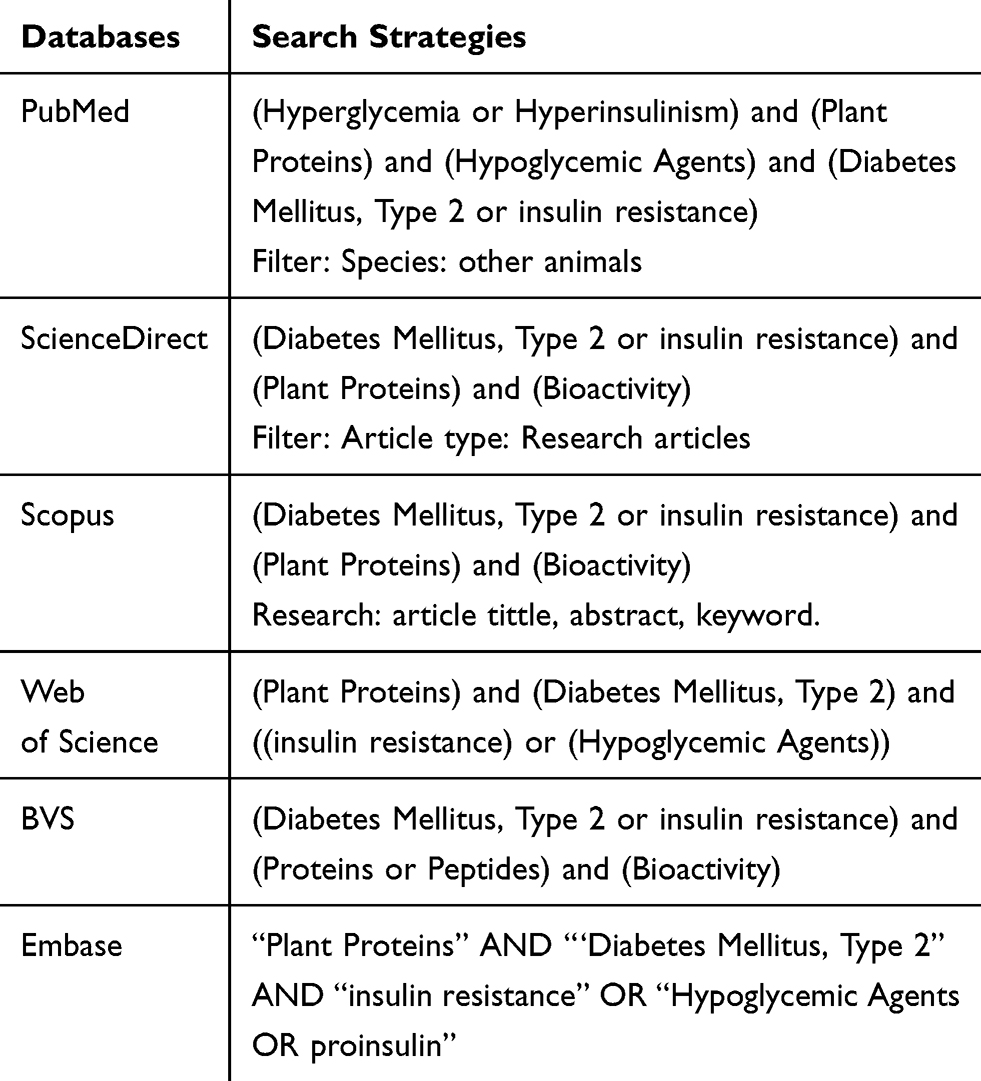 |
Table 1 Search Strategies for Databases |
The searches in the databases were performed on a computer with the IP of the Federal University of Rio Grande do Norte (UFRN), which allows access to all articles in each indexed database, through the “Portal de Periódicos Capes” at Brazil. The search in the Embase database was performed out by the University of São Paulo (USP).
Inclusion and Exclusion Criteria
Inclusion Criteria
Studies with male mice or rats with experimental T2DM (all species) with or without obesity were included. The treatment of T2DM was using purified or isolated proteins or peptides of plant sources. All times, frequencies and dosages used during the treatment of the selected studies were eligible for inclusion. Studies should present in their experimental design: well-defined groups of animals for evaluation and comparison of results, the substance used for the control group, whether it was a carrier solution (for example distilled water or saline) and/or known drugs for T2DM (for example insulin, metformin). Studies that presented measurements of serum glucose results and/or fasting serum insulin levels were included. Articles in all languages, original articles and indexed ones were included.
Exclusion Criteria
Studies with other animals, animals with experimental diabetes with comorbidities (except obesity), studies that used only an ex vivo approach, in vitro or in silico models were excluded. Studies related to the treatment of diabetes with plant extracts, non-protein vegetable origin, animal protein and non-response studies, case studies, cross-studies, studies without a separate control group and those without relevant results reported were excluded. The materials from the gray literature and the review articles were also excluded.
Data Extraction Process
The selection of articles started with the reading of the title, abstract and keywords without limitations of publication dates. Then the full text was read, in which the documents were analyzed to verify if they met the defined inclusion criteria.
The next step was the complete reading of the articles included in the systematic review. Qualitative assessment of the articles was performed with the SYRCLE Protocol. The scores were established according to compliance with the ten items of the protocol.
The full text was then read for data extraction: bibliographic data (authors, year of publication, publication period), characteristics of the animal model (species, lineage, age, sex, diabetic), of the exposure (protein or peptide, administration time, dose, frequency and route of exposure), of the study design (number of animals in the experimental and control groups, duration of follow-up and data collection time). The following outcomes were assessed: serum glucose and/or insulin (mg/dL), HOMA-IR index, glycated hemoglobin (HbA1C), oral glucose tolerance test (OGTT), oral insulin tolerance test (OITT).
Risk of Bias and Quality Evaluation
Two researchers were responsible for independent readings, inserted in Mendeley® software. Doubts were clarified with the help of a third researcher. Discrepancies were solved with the help of a third researcher. The reviewers were previously trained and calibrated to ensure uniformity in the evaluation of the criteria, and Cohen’s kappa concordance coefficient ranged from 0.6 to 1.0, with an average of 0.8. The evaluation of the evidence and the strength of the recommendations of the studies was evaluated with the SYstematic Review Center for Laboratory animal Experimentation - SYRCLE.15 The specifications for the assessment of each SYRCLE item for this review are described in the protocol.16
SYRCLE, a tool for analyzing the risk of bias, is designed specifically for animal studies. The SYRCLE consists of ten questions used to select and detect the performance, attrition and bias of the scientific articles included in the studies. The tool items were scored with “yes”, indicating low risk of bias or “No”, indicating high risk of bias.15 The specifications for the assessment of each SYRCLE item for this review are described in the study protocol.16
Results
Selection and Characteristics of Studies
Figure 1 shows the flowchart of the processes for selecting the studies. Using the specific search strategies for each database, a total of 916 articles were found, of which 46 were duplicated, leaving 870 publications for analysis. After screening the title, abstract and keywords, 24 articles were selected for full-text screening, and 05 publications were eligible to be included in the systematic review.
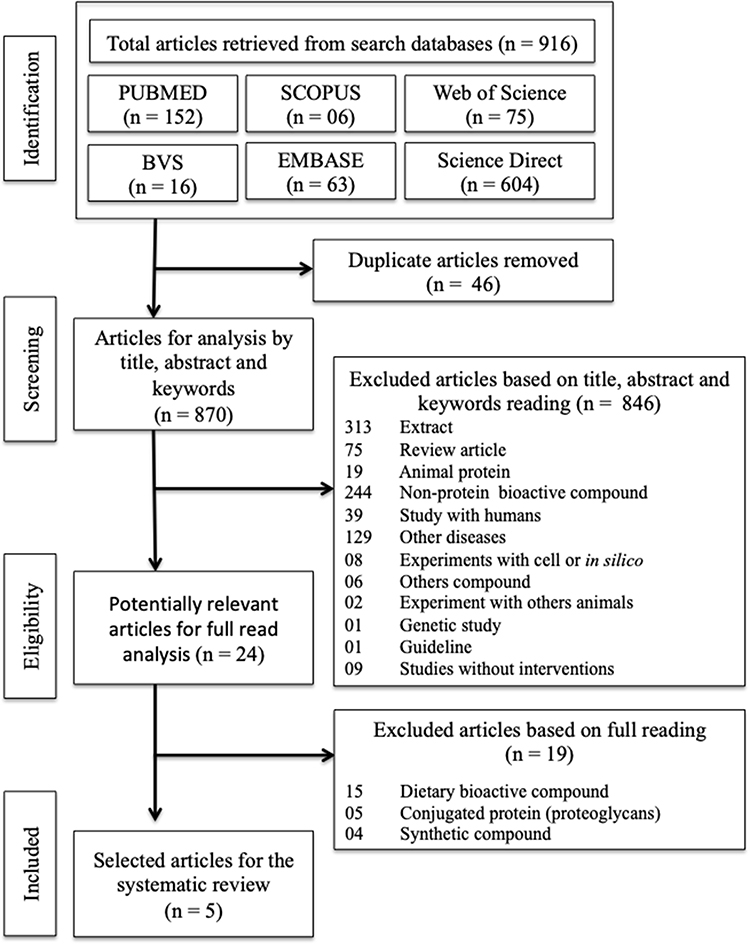 |
Figure 1 Flowchart for selection of study articles based on PRISMA-P. |
Risk of Bias and Quality Evaluation
The risk of bias and the methodological quality of the studies were evaluated in the five articles chosen for the construction of the study. The individual scores of the SYRCLE tool and the methodological quality indicators of each included study are provided, as shown in Table 2.
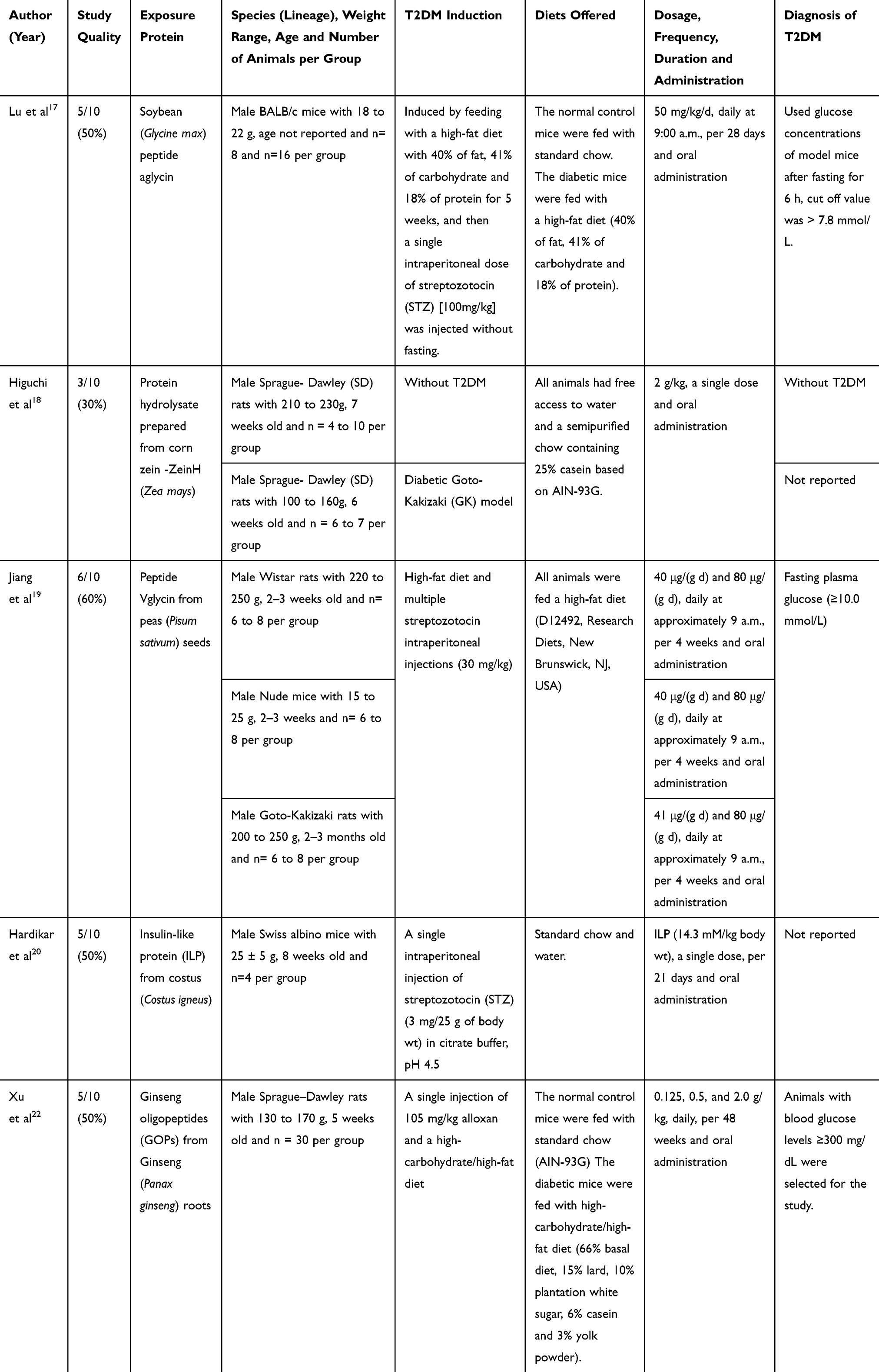 |
Table 2 Characteristics of the Studies Selected for the Systematic Review |
Considering selection bias, no study presented a method for generating sequences (0%; Q1). Although some studies mention the animals were randomly assigned to specific groups, the randomization method was not reported. All studies showed that there was pairing in relation to the characteristics of the animals in specific groups (100%; Q2). However, there was no blinding in order to avoid predictions of the results (0%, Q3). Three studies showed random allocation during the experiment (80%, Q4), but no study showed that there was blinding of the caregivers and researchers regarding the treatment for each group (0%, Q5).
Regarding the selection bias (Q6 and Q7), only 1 study described that there was a random selection of animals to evaluate the results (20%, Q6). However, in none of the articles the evaluator’s blinding in relation to the results was reported. (0%, Q7). Incomplete results data were adequately treated in almost all studies (80%, Q8). Regarding the reporting of biases, no risk was identified in the selected studies (100%, Q9). In addition, other potential sources of bias were not identified in any of the articles (100%, Q10).
Bioactive Protein and Peptide Mechanism of Action on T2DM
In the selected studies, different experimental designs were identified, with rats or mice of different strains. We could find studies that seek to identify metabolic pathways conditioning proteins and peptides of plant sources to improve the profile of diabetic conditions. In the study by Lu et al,17 the study peptide was aglycin - whose molecular mass is 3742.3 Da and amino acid sequence of 37 residues (ASCNGVCSPFEMPPCGSSACRCIPVGLVVGYCRHPSG). This peptide was purified from soybean (Glycine max) by high performance liquid chromatography (HPLC). T2DM was induced in BALB/c mice mixtures by feeding with a high fat content and a single dose of streptozotocin. The experimental model allowed the evaluation of gene and protein expression IR, IRS1, Akt and glucose transporter 4 (GLUT4), after administration of aglycin. The results showed that aglycin acted by restoring the expression of p-IR (forward, 5′GCCGCTCCTATCTCTGGTAT3′; reverse, 5′GAGTGATGGTGAGGTTGTGTTTG3′), p-IRS1 (forward, 5′AGTGGTGGAGTGGTAGGG3′), p-Akt (forward, 5′TAGGCATCCCTTCCTTACG3′; reverse, 5′GACACAATCTCCGCACCATA3′) and the membrane protein GLUT4 (forward, 5′TTCCTTCTATTTGCCGTCCTC3′; reverse, 5 ′CTGTTTTGCCCCTCAGTCATT3′), since in the condition of T2DM the expression of these proteins are reduced. In addition, aglycin was completely absorbed, detected intact in the animals’ plasma, 60 min after oral administration.
In the study with protein hydrolyzate prepared from corn zein (Zea mays), ZeinH - the amino acid sequence deposited in the NCBI contains 841 amino acid residues. However, this sequence was removed from the deposit because the gene was discontinued. Higuchi et al,18 used normal male Sprague-Dawley (SD) rats and the diabetic Goto-Kakizaki (GK) rats model, and found that the plasma glucose concentrations of the GK rats reduced significantly under treatment with ZeinH, when administered at 2 g/kg. They also observed that the serum concentrations of GLP-1 were significantly higher after 15 minutes of oral administration, in the treated rats, when compared to those that did not undergo the treatment. Regarding incretin, GIP, no stimulatory effect was observed in GK rats, but it was seen that in SD rats, without T2DM, there was a significant increase. Because the study showed a path of improvement in the glycemic profile in the diabetic model, by increasing the secretion of GLP-1, ZeinH was identified as an incretinotropic peptide.18
Jiang et al19 analyzed the peptide derived from peas (Pisum sativum) seeds, called Vglycin - whose molecular mass is of 3786.4 Da and the amino acid sequence of 37 residues (VSCNGVCSPFEMPPCGSSACRCIPYGLVVGNCRHPSG). This peptide also presents resistance to the digestive process, similarly to aglycin.17 In the experimental model, induction of T2DM in Wistar rats was done through a high-fat diet and multiple intraperitoneal injections of streptozotocin. Glucose and insulin tolerance tests were performed.
The group of diabetic rats that received Vglycin showed a reduction in serum insulin and glucose concentrations, compared to the group of diabetic rats that did not receive treatment. Thus, the authors inferred that there was an improvement in the resistance profile, characteristic in T2DM. In addition, the p-AKT and p-IR concentrations in the liver tissue of the animals were verified. The group treated with Vglycin presented similar results to the animals treated with insulin, in the physiological dosage, as a positive control. These results lead the authors to conclude that glycemia has a mimetic effect to insulin.19
Another study used an insulin-like protein (insulin-like protein - ILP) from costus (Costus igneus), which presents the amino acid sequence of two fragments: GLFAPIVVIAR and TCAAFTNEGSLIR, with molecular masses of 1155,725 and 1552,779 Da, respectively.20 However, it is not defined which sequences, or what pathways of action occur for the bioactive activity of such a molecule. Given these facts, they continued with the study published by Hardikar et al,20 in which the experimental model with male albino mice was used. T2DM was induced with a single injection of streptozotocin (STZ) in Swiss mice, and the animals were treated for 2 weeks with a single dose of ILP (14.3 mM/kg). The results indicated that the group treated with ILP did not show a reduction in the pancreatic islets, suggesting the pancreatic function was preserved, differently from the groups of untreated diabetic animals. This analysis indicated that ILP not only presents an insulin mimetic effect, but also protected the islets from further damage in diabetic Swiss mice.
Corroborating these findings, Joshi et al21 analyzed the oral activity and the insulin-like potential of ILP purified from the costus (Costus igneus) extract. Increasing concentrations of ILP were used to test the hypoglycemic property of the purified protein, with human insulin as a control, in RIN5f cell lines that are responsive to insulin (National Center for Cell Science, Pune). The amount of glucose that was not in the extracellular medium was dosed every 30 min for 180 minutes using the DNSA method. They identified that there was a rapid reduction in the concentrations of glucose in the supernatant in the wells. When using ILP it was observed that it had a better response in mediating glucose transport when compared to insulin. The authors concluded this result indicates an efficient uptake of glucose by the cells and insulin-like activity of ILP.
A study of the signaling pathway of ILP was developed in a cellular model using L6 myotubes.20 The findings indicated that ILP causes accumulation of IRS-1 and translocation to the plasma membrane of GLUT-4, facilitating the uptake and use of glucose, and promoting the reduction of blood glucose concentrations. These findings, in cellular models, corroborate with the experimental studies of Jiang et al19 and Lu et al.17
Xu et al22 conducted a study with the ginseng oligopeptides (GOPs) of Panax ginseng Meyer. The article did not present data on GOPs mass or its amino acid sequence, but sought to analyze its antidiabetic effect. They used Sprague-Dawley (SD) rats, and induced T2DM with alloxan injected intraperitoneally and a high-carbohydrate, high-fat diet. The authors analyzed plasma glucose and insulin concentrations of diabetic animals treated with GOPs and compared them to untreated rats. They found that the glucose concentrations of those treated with GOPs were lower compared to those not treated. In addition, an improvement in glucose resistance and insulin was also observed.
When analyzing the effect of GOPs on the expression of NF-kB, Bax, cleaved caspase-3 and Bcl-2 by Western-Blot, there was a significant difference in the expression of NF-kB, Bax, and caspase-3 when cleaved, while increasing Bcl-2 expression in the treated group at the end of the treatment for seven weeks. The histopathology and immunohistochemistry revealed a regeneration in the pancreatic islets, which were quite atrophied in the diabetic group without treatment. According to the study, GOPs seem to attenuate the diabetic condition by significantly inhibiting inflammation through the negative regulation of NF-kB expression and by having a potential effect in inhibiting oxidative stress and increasing the expression of the Bcl-2 family.22
Finally, the present study was able to produce a relevant contribution towards the organization and systematization on the understanding and interpretation regarding the metabolic pathways that are shown in Figure 2. This figure presents the metabolic pathways mentioned by each author and illustrates the regulatory pathways of these different proteins and bioactive peptides of plant origin found by the present review on the modulation of insulin resistance and in T2DM.
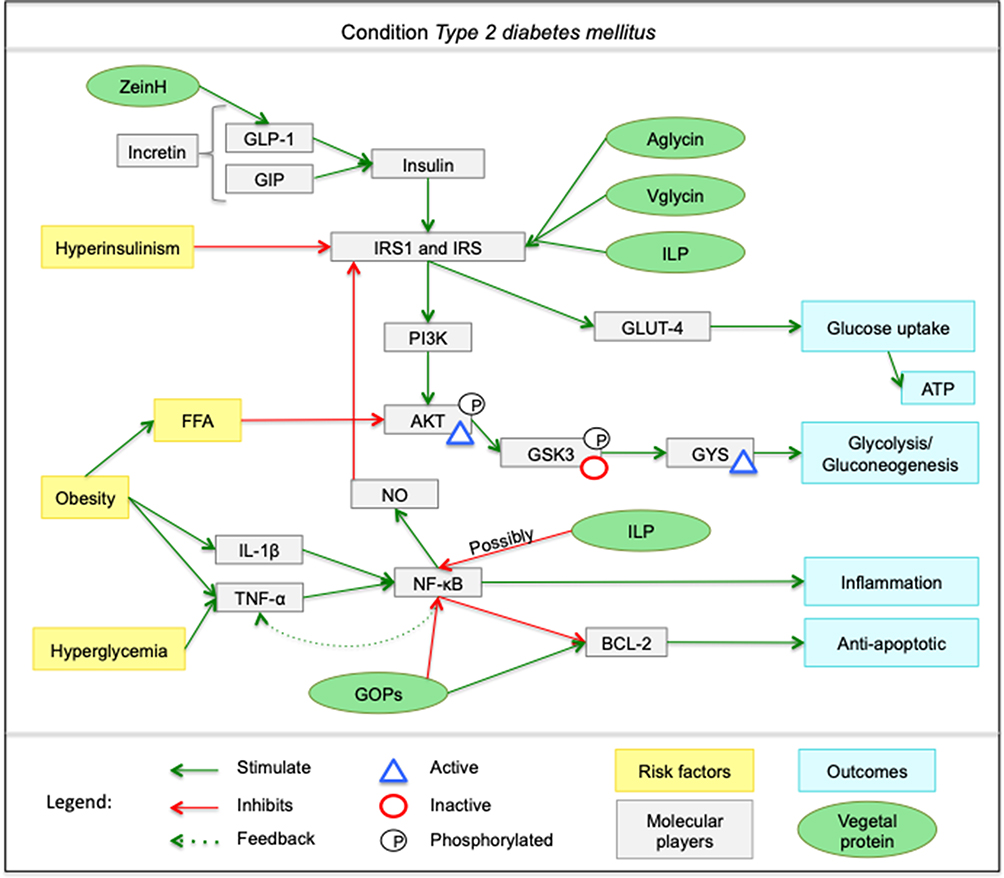 |
Figure 2 Mechanisms of action of proteins and peptides from selected articles focusing on T2DM. The ZeinH peptide acts by stimulating GLP-1, which signals increased insulin synthesis. The Aglycin, Vglycin and ILP peptides stimulate the IRS1 and IRS, which favors the serum glucose uptake cascade. In addition, ILP possibly acts by inhibiting the NF-κB signaling pathway. The GOPs peptide acts by inhibiting NF-κB, which is responsible for increasing inflammation signaling, and stimulates the expression of the BCL-2 family, which has anti-apoptotic activity. |
Discussion
The present systematic review study retrieved 5 experimental studies developed in animals. The analysis of the studies allowed the description of the mechanisms of action of isolated or purified proteins and peptides extracted from vegetables in the control of glycemia in T2DM.
Glucose metabolism involves several signaling pathways and is intrinsically related to the action of insulin. The entire process begins in the gastrointestinal tract, where nutrients are digested and absorbed, and, as this process occurs, there is hormonal and neuronal signaling for regulation of glucose concentrations, gastric motility and satiety. Among several hormones are glucose-dependent insulinotropic polypeptide (GIP) and glucagon-like peptide-1 (GLP-1), also called INCRETIN (Intestine Secretion Insulin), which are stimulated by the increase in plasma glucose to increase the secretion of insulin by insulin secreting granules in the pancreatic β cells.23
T2DM is a multifactorial metabolic disease characterized by hyperinsulinemia, hyperglycemia, insulin resistance (IR), hyperlipidemia and hypertension.24–26 IR implies a decrease in insulin sensitivity, decreasing glucose uptake, interfering with GLUT function, resulting in increased neoglycogenesis in the liver, and postprandial hyperglycemia. This metabolic condition is directly associated with obesity and cardiometabolic diseases, in which there is an increase in lipolysis and consequently high circulating concentrations of free facts acids (FFAs) leading to an increase in the synthesis of triglycerides and overproduction of lipoproteins from very low density (LDL).25
Inflammation is another pathway that negatively influences insulin activity. Increased concentrations of FFA have anti-insulin action and are directly proportional to the plasma concentrations of inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), interleukin-1β (IL-1β), resistin and others.25 These inflammatory cytokines, produced by adipocytes and immunocytes, trigger the development of insulin resistance and DM through mechanisms involving various kinases and signaling pathways.27–31
The inflammatory cytokines are involved in the activation of the NF-κB pathway, leading to the serine phosphorylation of the insulin receptor substrate (IRS), resulting in resistance to the action of insulin.29,30 Also, DM occurs due to pancreatic islet β cell dysfunctions caused by excess IL-6 and TNF-α.28 Compounds that can act in this way modulate the effect of resistance to the action of insulin and consequently, T2DM.
These metabolic changes can lead to oxidative stress, negatively affecting insulin activity, due to the formation of reactive oxygen species (ROS): such as hydrogen peroxide and superoxide anions.32 These species can damage the β cells from the pancreas islets, resulting in reduced insulin release.33
Some limitations of the present systematic review should be mentioned. The present study intended to be a meta-analysis, and this was not performed because: 1) Bioactives were different from each other, although all from vegetable origin. Not being the same molecules, the amino acid sequences reported were different. Aglycin and vglycin were the only very similar, but still distinct from one another; 2) Studies were conducted using different animals, some with a lineage without genetic alterations and others with genetic alterations for diabetes, which lead to greater heterogeneity of the animals; 3) Different blood glucose measurement protocols – some studies dosed for weeks and others for minutes; 4) Data were presented in graphic formats, without the appropriate standard deviation values, which made the meta-analysis unfeasible.
Nevertheless, the present study contributed to the understanding that bioactives in protein sources can change biochemical pathways involved in the development of T2DM. The ZeinH peptide, acts on the increase of GLP-1 expression, stimulating the insulin release. At this point, it is possible to list the Aglycin, Vglycin and ILP that influence IRS. These peptides act increasing the expression of IRS, improving GLUT translocation and consequent glucose uptake and plasmatic maintenance of glucose and insulin. Besides, IRS also acts by activating the PI3K/AKT cascade, culminating in glycolysis and gluconeogenesis.
The GOPs peptide is related to the reduction of NF-κB signaling, helping to decrease the inflammatory profile and nitric oxide synthesis, which is associated with less IRS signaling. GOP also acts by increasing the expression of BCL-2, which has an anti-apoptotic function. ILP also appears to reduce NF-κB expression, according to experiments with cells.22
Furthermore, clinical trials using the extract of ginseng (Panax ginseng), the root source of GOP,22 are in research, as reported in the systematic review and meta-analysis of researchers Gui et al,34 which aimed to search the scientific literature for randomized clinical trials that assessed the effect of ginseng (Panax ginseng) or ginsenoside Re versus control, in patients with T2DM or with impaired glucose tolerance. The research shows the effectiveness of therapy related to ginseng (Panax ginseng) supplementation in the concentrations of postprandial insulin, HOMA-IR, triglycerides, total cholesterol and LDL-cholesterol in patients with T2DM or impaired glucose tolerance, with no effect on HbA1c levels or HDL-cholesterol. Thus, revealing a possible research path for application in human health.
It is known that the pathogenesis for the appearance of T2DM involves several metabolic pathways.35 This study identified that bioactive proteins and peptides of plant origin also act by several biochemical pathways, modulating the effect of insulin resistance and consequently, the maintenance of the hyperglycemic state and progression of T2DM.
These proteins and peptides of plant origin, mainly Aglycin, Vglycin and ILP, act similarly to human insulin. This finding shows the importance of studies in the area to test their safety and applicability in humans since they present great potential as hypoglycemic agents. Thus, these bioactives are likely to meet the public health global demand for alternative treatments for diabetes mellitus.
Conclusion
Bioactive proteins and peptides of plant origin control T2DM acting on the insulin signaling pathway. Three peptides - Aglycin, Viglycin and ILP act stimulating the IRS. ZeinH stimulates GLP-1 incretin, while GOPs downregulate NF-kB. The studies found in the present systematic review suggest that these bioactive proteins and peptides act as insulin-like molecules and thus promote secondary stimuli in the insulin signaling cascade until glucose uptake. In addition to concluding the mechanism of action, in this review, the perception of fragility that the experimental articles are subject to was notorious. Therefore, it is suggested that future studies should be more careful and precise about the methodological procedures to meet with quality the requirements assessed by the risk of bias tools such as SYRCLE. Therefore, these bioactives are promising for research, because they are naturally found in vegetables, and might represent alternative efforts in the diabetes mellitus treatment.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This work received the financial support from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES - finance code 001) and the Conselho Nacional de Desenvolvimento Cientítico e Tecnológico (CNPq - Award Number: 426116/2018-6) research promotion agencies.
Disclosure
The authors report no conflicts of interest for this work.
References
1. American Diabetes Association. Classification and diagnosis of diabetes: standards of medical care in diabetes-2019. Diabetes Care. 2019;42:S13–S28. doi:10.2337/dc19-S002
2. Bertuzzi A, Conte F, Mingrone G, Papa F, Salinari S, Sinisgalli C. Insulin signaling in insulin resistance states and cancer: a modeling analysis. PLoS One. 2016;11(5):1–26. doi:10.1371/journal.pone.0154415
3. Boucher J, Kleinridders A, Kahn CR. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb Perspect Biol. 2014;6(a009191):1–23. doi:10.1101/cshperspect.a009191
4. Aronoff SL, Berkowitz K, Shreiner B, Want L. Glucose metabolism and regulation: beyond insulin and glucagon. Diabetes Spectr. 2004;17(3):183–190. doi:10.2337/diaspect.17.3.183
5. Shaw LM. The insulin receptor substrate (IRS) proteins: at the intersection of metabolism and cancer. Cell Cycle. 2011;10(11):1750–1756. doi:10.4161/cc.10.11.15824
6. Belgi A, Akhter Hossain M, W Tregear G, D Wade J. The chemical synthesis of insulin: from the past to the present. Immun Endoc Metab Agents Med Chem. 2011;11(1):40–47. doi:10.2174/187152211794519412
7. Prasad VSS, Adapa D, Vana DR, Choudhury A, Jahangir MA, Chatterjee A. Nutritional components relevant to Type-2-diabetes: dietary sources, metabolic functions and glycaemic effects. J Res Med Dent Sci. 2018;6(5):52–75.
8. Carvalho FMCC, Lima VCOO, Costa IS, et al. A trypsin inhibitor from tamarind reduces food intake and improves inflammatory status in rats with metabolic syndrome regardless of weight loss. Nutrients. 2016;8(544):1–14. doi:10.3390/nu8100544
9. Kitamura K, Takamura Y, Iwamoto T, et al. Dammarane-type triterpene extracts of Panax notoginseng root ameliorates hyperglycemia and insulin sensitivity by enhancing glucose uptake in skeletal muscle. Biosci Biotechnol Biochem. 2017;81(2):335–342. doi:10.1080/09168451.2016.1246173
10. Choi J, Kim KJ, Koh EJ, Lee BY. Gelidium elegans extract ameliorates type 2 diabetes via regulation of MAPK and PI3K/Akt signaling. Nutrients. 2018;10(1):51. doi:10.3390/nu10010051
11. Bem GF, Costa CA, Santos IB, et al. Antidiabetic effect of euterpe oleracea mart. (açaí) extract and exercise training on high-fat diet and streptozotocin-induced diabetic rats: A positive interaction. PLoS One. 2018;13(6):1–19. doi:10.1371/journal.pone.0199207
12. Lo HY, Ho TY, Li CC, Chen JC, Liu JJ, Hsiang CY. A novel insulin receptor-binding protein from Momordica charantia enhances glucose uptake and glucose clearance in vitro and in vivo through triggering insulin receptor signaling pathway. J Agric Food Chem. 2014;62(36):8952–8961. doi:10.1021/jf5002099
13. Lo HY, Ho TY, Lin C, Li CC, Hsiang CY. Momordica charantia and its novel polypeptide regulate glucose homeostasis in mice via binding to insulin receptor. J Agric Food Chem. 2013;61(10):2461–2468. doi:10.1021/jf3042402
14. Moher D, Shamseer L, Clarke M, et al.; PRISMA-P Group. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst Rev. 2015;207:1–9. doi:10.1186/2046-4053-4-1.
15. Hooijmans CR, Rovers MM, de Vries RB, Leenaars M, Ritskes-Hoitinga M, Langendam MW. SYRCLE’s risk of bias tool for animal studies. BMC Med Res Methodol. 2014;14(10):1281–1285. doi:10.7507/1672-2531.20140206
16. Medeiros AF, Costa IS, Piuvezam G, Medeiros GCBS, Maciel BLL, Morais AHA. Mechanism of the action of bioactive proteins of vegetables in diabetes mellitus type 2. Medicine. 2019;98(39):e17326. doi:10.1097/MD.0000000000017326
17. Lu J, Zeng Y, Hou W, et al. The soybean peptide aglycin regulates glucose homeostasis in type 2 diabetic mice via IR/IRS1 pathway. J Nutr Biochem. 2012;23(11):1449–1457. doi:10.1016/j.jnutbio.2011.09.007
18. Higuchi N, Hira T, Yamada N, Hara H. Oral administration of corn zein hydrolysate stimulates GLP-1 and GIP secretion and improves glucose tolerance in male normal rats and Goto-Kakizaki rats. Endocrinology. 2013;154(9):3089–3098. doi:10.1210/en.2012-2275
19. Jiang H, Feng J, Du Z, et al. Oral administration of soybean peptide Vglycin normalizes fasting glucose and restores impaired pancreatic function in Type 2 diabetic Wistar rats. J Nutr Biochem. 2014;25(9):954–963. doi:10.1016/j.jnutbio.2014.04.010
20. Hardikar MR, Varma ME, Kulkarni AA, Kulkarni PP, Joshi BN. Elucidation of hypoglycemic action and toxicity studies of insulin-like protein from Costus igneus. Phytochemistry. 2016;124:99–107. doi:10.1016/j.phytochem.2016.02.001
21. Joshi BN, Munot H, Hardikar M, Kulkarni AA. Orally active hypoglycemic protein from Costus igneus N. E. Br.: an in vitro and in vivo study. Biochem Biophys Res Commun. 2013;436(2):278–282. doi:10.1016/j.bbrc.2013.05.093
22. Xu M, Sun B, Li D, et al. Beneficial effects of small molecule oligopeptides isolated from Panax ginseng meyer on pancreatic beta-cell dysfunction and death in diabetic rats. Nutrients. 2017;9(10):1061. doi:10.3390/nu9101061
23. Ezcurra M, Reimann F, Gribble FM, Emery E. Molecular mechanisms of incretin hormone secretion. Curr Opin Pharmacol. 2013;13:922–927. doi:10.11569/wcjd.v23.i16.2582
24. Ma Q, Li Y, Li P, et al. Research progress in the relationship between type 2 diabetes mellitus and intestinal flora. Biomed Pharmacother. 2019;117:109138. doi:10.1016/j.biopha.2019.109138
25. Genser L, Rossario J, Mariolo C, Castagneto-gissey L, Panagiotopoulos S. Obesity, Type 2 diabetes, and the metabolic syndrome pathophysiologic relationships and guidelines for surgical intervention. Surg Clin North Am. 2016;96:681–701.
26. Halim M, Halim A. The effects of in fl ammation, aging and oxidative stress on the pathogenesis of diabetes mellitus (type 2 diabetes). Diabetes Metab Syndr. 2019;13(2):1165–1172. doi:10.1016/j.dsx.2019.01.040
27. Crook M. Type 2 diabetes mellitus: a disease of the innate immune system? An update. Diabet Med. 2004;21:203–207. doi:10.1046/j.1464
28. Donath MY. Targeting inflammation in the treatment of type 2 diabetes. Diabetes Obes Metab. 2013;15(S3):193–196. doi:10.1111/dom.12172
29. Mahmoud F, Al-Ozairi E. Inflammatory cytokines and the risk of cardiovascular complications in type 2 diabetes. Dis Markers. 2013;35(4):235–241. doi:10.1155/2013/931915
30. Patel PS, Buras ED, Balasubramanyam A. The role of the immune system in obesity and insulin resistance. J Obes. 2013;2013. doi:10.1155/2013/616193
31. Gratas-Delamarche A, Derbré F, Vincent S, Cillard J. Physical inactivity, insulin resistance, and the oxidative-inflammatory loop. Free Radic Res. 2014;48(1):93–108. doi:10.3109/10715762.2013.847528
32. Rösen P, Nawroth PP, King G, Möller W, Tritschler HJ, Packer L. The role of oxidative stress in the onset and progression of diabetes and its complications: A summary of a congress series sponsored by UNESCO-MCBN, the American diabetes association and the German diabetes society. Diabetes Metab Res Rev. 2001;17(3):189–212. doi:10.1002/dmrr.196
33. Evans JL, Goldfine ID, Maddux BA, Grodsky GM. Perspectives in diabetes: are oxidative stress - activated signaling pathways mediators of insulin resistance and B-cell dysfunction? Diabetes. 2003;52:1–8. doi:10.2337/diabetes.52.1.1
34. Gui QF, Xu ZR, Xu KY, Yang YM. The efficacy of ginseng-related therapies in Type 2 diabetes mellitus. Medicine (United States). 2016;95(6):1–10. doi:10.1097/MD.0000000000002584
35. Hameed I, Masoodi SR, Mir SA, Nabi M, Ghazanfar K, Ganai BA. Type 2 diabetes mellitus: from a metabolic disorder to an inflammatory condition. World J Diabetes. 2015;6(4):598. doi:10.4239/wjd.v6.i4.598
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.