Back to Journals » Infection and Drug Resistance » Volume 11
Insight into Acinetobacter baumannii: pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities
Authors Asif M, Alvi IA, Rehman SU ![]()
Received 27 February 2018
Accepted for publication 20 April 2018
Published 21 August 2018 Volume 2018:11 Pages 1249—1260
DOI https://doi.org/10.2147/IDR.S166750
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Muhammad Asif,1,2 Iqbal Ahmad Alvi,1,3 Shafiq Ur Rehman1
1Department of Microbiology and Molecular Genetics, University of the Punjab, Lahore, Pakistan; 2Department of Pathology, King Edward Medical University, Lahore, Pakistan; 3Department of Microbiology, Hazara University, Mansehra, Pakistan
Abstract: Acinetobacter baumannii, once considered a low-category pathogen, has emerged as an obstinate infectious agent. The scientific community is paying more attention to this pathogen due to its stubbornness to last resort antimicrobials, including carbapenems, colistin, and tigecycline, its high prevalence of infections in the hospital setting, and significantly increased rate of community-acquired infections by this organism over the past decade. It has given the fear of pre-antibiotic era to the world. To further enhance our understanding about this pathogen, in this review, we discuss its taxonomy, pathogenesis, current treatment options, global resistance rates, mechanisms of its resistance against various groups of antimicrobials, and future therapeutics.
Keywords: antibiotic resistance, colistin, tigecycline, phage therapy
Introduction
Acinetobacter baumannii, a non-fermenter Gram-negative coccobacillus, was considered a low-category pathogen in the past, but has now emerged as a leading cause of hospital- and community-acquired infections. It is a frequent cause of pneumonia and septicemia in immunocompromised patients. It resists many classes of antibiotics by virtue of chromosome-mediated genetic elements on one hand, while it can also persist for a prolonged period in harsh environments (walls, surfaces, and medical devices) in the hospital settings on the other hand.1,2
A. baumannii was isolated for the first time from soil by a Dutch bacteriologist Beijerinck in 1911 and was described as Micrococcus calcoaceticus.3 In succeeding 50 years, the same bacterium was isolated many times and reported with different names such as Moraxella lwoffi, Alcaligenes hemolysans, Mirococcuscalco-aceticus, and Herellea vaginicola. Four decades later, Brisou and Prevot purposed to include it in the genus Achromobacter, based on its inability to move and being non-pigmented.4 In 1968, Baumann et al placed all such isolates in one genus Acinetobacter, which was accepted by the committee on the taxonomy of Moraxella and Allied Bacteria 4 years later.5 Based on DNA similarity, Bouvet and Grimont further classified it into 12 groups in 1986.6 Currently, they are taxonomically classified as γ-proteobacteria, family Moraxellaceae and order Pseudomonadales.7
Acinetobacter calcoaceticus-baumannii complex is a group of aerobic, non-fermentative, gram-negative coccobacillus that encompasses four different Acinetobacteria, comprising A. baumannii, Acinetobacter pittii, Acinetobacter nosocomialis, and Acinetobacter calcoaceticus. The first three are implicated in infections, while the latter is rarely considered pathogenic.8 It appears as Gram-negative coccobacillus in pairs ranging from 1 to 1.5 µm when observed under the microscope after gram staining. It often resists complete decolorization and can deceive as Gram-positive cocci. Nutritionally, it is aerobic, non-fastidious, and a non-fermenter. It is a non-motile organism and does not produce cytochrome oxidase, urease, citrate, and indole; however, it produces catalase enzyme. A. calcoaceticus-baumannii complex nurtures well at 35°C–37°C; however, some environmental isolates grow well in the temperature range of 20°C–30°C. A. baumannii is the only bacterium in the genus that can grow at 44°C.6
It grows well on routine laboratory media such as blood agar, chocolate agar, and MacConkey agar. On blood agar, it forms colorless, non-hemolytic, shiny mucoid colonies, smooth in contexture with a diameter of 1–2 mm after 18–24 hours of incubation at 37°C. It produces colorless colonies on MacConkey agar which are shiny mucoid and tomb shaped, indicating its non-lactose fermenting ability. On selective agar, Leeds Acinetobacter Medium, it gives pink color colonies when grown in the presence of supplement.9
Acinetobacter spp. are free-living saprophytic organisms and widely distributed in different environments including soil, water, wastewater, vegetables, and skin of animals and humans.10 They have been isolated from various body parts of healthy individuals, including the nose, ear, throat, forehead, trachea, conjunctiva, vagina and perineum, axillae, groin, hands, and toe webs; however, most strains isolated were other than A. baumannii.11 In hospital environment, they reside on beds, curtains, walls, roofs, medical devices, and equipment, as well as on belongings of medical personnel, tap water sinks, telephones, door handles, hand sanitizers, dispensers, trolleys, bins, and even on computers. They have the capacity to survive for prolonged periods on inanimate objects. The factors that are responsible for their persistence in a hospital environment are resistance to key antimicrobial drugs and disinfectants and their ability to survive in desiccants.12
Pathogenicity of A. baumannii
A. baumannii has emerged as a major culprit involved in causing nosocomial infections, especially in intensive care units (ICUs) worldwide. The capability of this organism to pollute hospital surfaces for extended periods is linked with nosocomial outbreaks.13 It has gained the ability to infect not only hospitalized patients but also the general population. In hospital settings, it confers 26% mortality rate that goes up to 43% in ICUs.14 A. baumannii is a principal agent of ventilator-associated pneumonia, which accounts for nearly 15% of all hospital-acquired infections, with the highest morbidity and mortality in medical wards and especially in the ICUs. It accounts for ~50% of the total use of antibiotics in the ICUs.15
A. baumannii is not considered a community pathogen, but in immunocompromised individuals and in children, it populates tracheostomy sites and can cause community-acquired bronchiolitis and tracheobronchitis. It has also been implicated in community-acquired pneumonia with underlying conditions such as smoking, alcoholism, diabetes mellitus, and COPD in tropical regions of Asia and Australia.16 A. baumannii has been implicated in bloodstream infections in 10%–15% of cases due to invasive procedures (intravascular or respiratory catheters, tubes, or cannulas). In 20%–70% of A. baumannii infections, the origin of infection remains unknown.17
A. baumannii is an increasing threat to neurosurgery patients. It is responsible for 4% of all meningitis and shunt-related infections, with 70% mortality rates.18 It is responsible for 2.1% of ICU-acquired wound infections; however, its prevalence is more pronounced (32%) in casualties from battlefields of Afghanistan and Iraq. It is not a usual agent of urinary tract infections (UTIs); however, it can cause infection in debilitated elderly patients and in patients with prolonged indwelling catheter-related infections in the ICUs where it contributes 1.6% of the total UTIs. It may cause endocarditis, keratitis, and ophthalmitis following use of the contact lens and eye surgery.19
A. baumannii can be transmitted through the vicinity of affected patients or colonizers such as linens fomites, curtains, bed rails, tables, sinks, doors, feeding tubes, and even medical equipment. Contamination of respiratory support equipment, suction devices, and devices used for intravascular access is the key source of infection.20
A. baumannii is considered as a low-virulence pathogen, unless it is isolated from patients having comorbidities such as neonates with low birth weights and elderly patients with chronic illnesses such as malignancy. Major predisposing factors important in the acquisition of A. baumannii infection include prolonged hospital stay, mechanical ventilation, intravascular device, advanced age, immunosuppression, previous broad-spectrum antimicrobial therapy, previous sepsis, ICU stay, and enteral feedings.21
Pathogenesis of A. baumannii
The intrusion of a microorganism requires cell-to-cell adhesion to establish infection; however, the capability of A. baumannii to anchor with cells/mucosal cells is low as compared to other microorganisms such as Pseudomonas aeruginosa, Neisseria meningitides, Campylobacter, Yersinia enterocolitica, and Helicobacter pylori.22 The reduced adhesion and invasion of A. baumannii attribute to its low virulence; however, it possesses a hydrophobic ability that provides attachment to foreign materials such as plastics used in intravascular devices. It has been proven that surface hydrophobicity is highly expressed in strains isolated from patients as compared to normal flora of the skin.19
Outer membrane protein A (OmpA) is associated with improving adhesion, specifically to the epithelial cells of the respiratory tract. It localizes in the mitochondria and nuclei and induces expression of proapoptotic molecule cytochrome c, resulting in cell death.23 A. baumannii evades alternative complement pathway-mediated killing by neutralizing factor H, a key regulator of alternative complement pathway, with the help of OmpA. This phenomenon is known as serum resistance of A. baumannii.24 OmpA induces differentiation of CD4+, activation and maturation of dendritic cells, and causes their premature apoptosis.25
Secretion of outer membrane vesicles that contain different virulence-related proteins (proteases, phospholipases, superoxide dismutase, and catalase) at the infection site accelerates the local innate immune response and ultimately leads to tissue damage. The outer membrane vesicles also augment biofilm formation on abiotic surfaces.26 Polysaccharide capsule of Gram-negative rods is notorious as a virulence factor. It plays a central role in guarding bacteria against phagocytosis by the host innate immune system.27 Lipopolysaccharides (LPSs) of A. baumannii consist of an O-antigen, the carbohydrate core, and a lipid A moiety. LPS is a chemotactic agent that recruits inflammatory cells and compels them to release their cytotoxic material.28
Quorum sensing is the capability of bacteria to communicate with their neighboring counterparts to respond jointly to the changing environment. They produce small easily diffusible hormone-like molecules known as autoinducers, which are used to observe their population density and to adapt in an ever-changing environment.29 Like other Gram-negative rods, Acinetobacter yields acylhomoserine lactones as signaling molecules for interspecies and intraspecies communication. It also produces less-studied signaling molecules such as diketopiperazines, 2-heptyl-3-hydroxy-4-quinolone, and retention factor 1.30
Despite the abundance of iron in biological systems, availability of biologically active ferric iron is relatively low due to its decreased solubility in an aerobic environment and chelation by other compounds such as hemoglobin and ferric-binding protein called transferrin.31 A. baumannii is unable to acquire iron from transferrin or lactoferrin; however, it possesses siderophores, which have iron acquisition ability devoted to iron accumulation from heme.32
The ability of A. baumannii to form biofilms on biotic and abiotic surfaces is a well-studied mechanism of resistance. To survive in unfavorable conditions, it becomes metabolically inert in the deeper layers of biofilms. Poor penetration and the inability of antibiotics to act on metabolically inert bacteria augment its virulence.14 A. baumannii involved in epidemics shows a high-level desiccation resistance and biofilm-forming capability on biological surfaces. The property of A. baumannii to form pellicle by virtue of polysaccharide, poly-N-acetyl glucosamine, and csuA/B usher protein is a way to offshore antibiotic effect. Other virulence factors are also involved in biofilm evolution, including biofilm-associated protein (BAP), OmpA, BAP-like protein-1 (BLP-1), and BAP-like protein-2 (BLP-2).33
Antibiotic resistance and treatment options against A. baumannii
Discovery of antibiotics was a remarkable milestone in the history of modern medicine. The discovery of penicillin, followed by sulfonamides and aminoglycosides, urged scientists to speculate that a “magic bullet” to wipe out the infectious diseases has been found. However, unfortunately, the scenario is not as true as once thought. Fleming stated in his Nobel lecture that, in future, antibiotics will be easily available to everyone and quacks might undermine the positive role of antibiotics through exposing a persistent low regimen to bacteria that can result in the evolution of antibiotic resistance.34
Penicillin was administered for the first time in 1941, and penicillin-resistant isolates were detected in 1942. Similarly, methicillin was introduced in 1960 and methicillin-resistant strains were reported in 1961 and so on.35 Currently, the isolates of A. baumannii resistant to all available antimicrobials have been reported.36 The hard work of scientific community resulted in discovery of many antibiotics, but their misuse resulted in high degree of resistance. It can be said that pre-antibiotic era has started, where again microbes with greater killing capacity are in abundance.37 Acinetobacter has been endowed with the genetic setup for rapid development of antimicrobial resistance, and therefore, is known as a natural transformant. Scientific literature is full of reports stating it as one of the toughest bacteria.38
Until early 1970s, Acinetobacter infections were treatable with ampicillin, carbenicillin, gentamicin, and nalidixic acid, either as a monotherapy or combination therapy, but high rates of resistance were noticed after 1975.39 Presently, many valuable drugs such as ureidopenicillin, aminopenicillins, narrow-spectrum and even extended-spectrum cephalosporin, tetracycline, chloramphenicol, cephamycins such as cefoxitin, and most aminoglycosides have lost their efficacy against Acinetobacter.40
Carbapenems (imipenem and meropenem), aminoglycosides (amikacin and tobramycin), fluoroquinolones, (ciprofloxacin and levofloxacin), broad-spectrum cephalosporins (ceftazidime, cefotaxime, ceftriaxone, cefepime), and combinations of beta-lactamase inhibitors with antibiotic (ampicillin/sulbactam) are currently being used, provided that the organisms are susceptible; however, the minimum inhibitory concentrations (MICs) have substantially increased.1
Carbapenems
Carbapenems have been the mainstay of antimicrobial therapy against A. baumannii infections since 1990. Overwhelming resistance to carbapenems was first reported in 1985, the year of imipenem discovery, declaring that antibiotic resistance mechanisms existed even before their first use. Currently, about 8%–26% isolates are susceptible to imipenem, depending on the region of the world.41 North America and Europe harbor 13%–15% of carbapenem-resistant Acinetobacter, in comparison to Latin America, where 40% resistance has been reported.42 Another study reported 48% carbapenem resistance in the USA.43 Alarmingly, a recent review mentioned 50%, 85%, and 62%–100% as the frequency of carbapenem-resistant Acinetobacter in Singapore, India, and Pakistan, respectively. Likewise, the frequency of carbapenem-resistant A. baumannii was reported to be 70%, 92%, and 100% in Chile, Korea, and Portugal, respectively. The resistance to carbapenems also renders other beta-lactam drugs ineffective.44 Worse clinical outcomes with carbapenem-resistant Acinetobacter infections have been reported by many authors.44
Colistin
Colistin or polymyxin E is a bactericidal drug that disrupts cell membrane like a detergent. Its positively charged cationic region binds to negatively charged hydrophilic portion of LPSs. The resulting loss of integrity causes cell death.45 The current panic situation of antibiotic resistance in Acinetobacter infections had led to the use of historically discarded drug, colistin. Colistin has shown high nephrotoxicity, ranging from 11% to 76% in various retrospective and prospective studies. Therefore, its use was discontinued short after its discovery in late 1950s. However, recent studies do not advocate such higher incidence of nephrotoxicity in comparison to previous studies, if associated risk factors are kept in mind, such as dose, age of patient, duration, and existing comorbidities such as hypertension, obesity, and hypoalbuminemia.46,47 The mechanisms of colistin-induced nephropathy are not clearly understood; however, certain studies in this field suggested that accumulation of colistin in proximal renal tubules results in oxidative damage. Caspase-mediated apoptosis, inducible nitrous oxide synthase, and endothelial nitrous oxide synthase are also implicated in pathogenesis of nephrotoxicity.48 Colistin-driven neurotoxicity is infrequent and includes bronchoconstriction, cough, and chest tightness when administered via respiratory tract and results in chemical meningitis.49,50 Other dose-dependent reversible adverse effects include ataxia, apnea, paresthesias, delirium, visual disturbances, seizures, vertigo, and neuromuscular defects.51
Colistin is administered as a pro-drug in the form of colistin methanesulfonate; so, achievement of its critical levels in blood is difficult. Therefore, its use as monotherapy results in rapid emergence of regrowth. Heteroresistance (selective resistance of bacterial subpopulation followed by amplification) is another phenomenon that results in rapid emergence of resistant clones.52,53
Low plasma levels and heteroresistance of colistin raised serious concern on colistin monotherapy.54 Combination of colistin with other in vitro active agents that gives a synergistic effect is widely used by physicians in critically ill patients. However, debate is still open in literature about the advantages of combination versus monotherapy. Some studies suggest monotherapy as effective as combination.55,56 Extensive review of literature favored combination therapy in terms of microbiological clearance as well as clinical cure in A. baumannii infections.57,58 Colistin/carbapenem and colistin/rifampicin are the most studied combinations that have well-established in vitro and in vivo activities, and also, proved to be effective in the clinics.59,60 Other studied combinations include colistin/tigecycline, colistin/minocycline, colistin/aminoglycosides, colistin/ampicillin–sulbactam, colistin/trimethoprim– sulfamethoxazole, colistin/fosfomycin, colistin/daptomycin, and colistin/sulbactam.61–64
Unfortunately, resistance has emerged against this last resort antibiotic. Colistin-resistant Acinetobacter was first reported in the Czech Republic in 1999.65 Now it is increasingly being reported worldwide.66 Low levels of resistance (2.1%–7.1%) have been reported from the USA, while reports from Europe have documented 7%–11% resistance. Highest resistance has been reported from India (53%), followed by Iran (48%), Spain (40.7%), and Korea (30%)57,67 as shown in Table 1.
 | Table 1 Studies showing colistin resistance by Acinetobacter in different regions of the world |
Tigecycline
Tigecycline, being the first member of glycycline, is a novel drug approved by the US Food and Drug Administration in June 2005 for the treatment of complicated skin infections, community-acquired pneumonia, and intra-abdominal infections.68 It is also being used in the treatment of bacteremia and UTIs by multidrug-resistant (MDR) Gram-negative bacteria.69 It is active against a wide number of Gram-positive and Gram-negative bacteria including anaerobes.70 It has shown effectiveness against A. baumannii and other species of Acinetobacter in large number of studies.71
Testing the sensitivity of A. baumannii to tigecycline is not standardized yet. The European Committee on Antimicrobial Susceptibility Testing and Clinical and Laboratory Standards Institute still do not have established breakpoints for tigecycline sensitivity testing. However, many researchers use more flexible breakpoints, as reported by the US Food and Drug Administration (sensitive: ≤2 mg/L, resistant ≥8 mg/L).72 Therefore, interpretation of antimicrobial sensitivity in many studies has been controversial. The method of MIC determination also affects the results: E test gives somewhat higher MIC value than broth dilution method.73 The determination of MIC by Vitek 2 is reliable in 94% cases.74
The first case of tigecycline resistance was reported by Sader et al in 2005 and in 2007 Navon-Venezia et al reported 66% tigecycline resistance against A. baumannii in Israel.75,76 At times, varying percentages of resistance have been reported all over the world, with Turkey possessing the highest resistance rate (81%), as shown in Table 2.
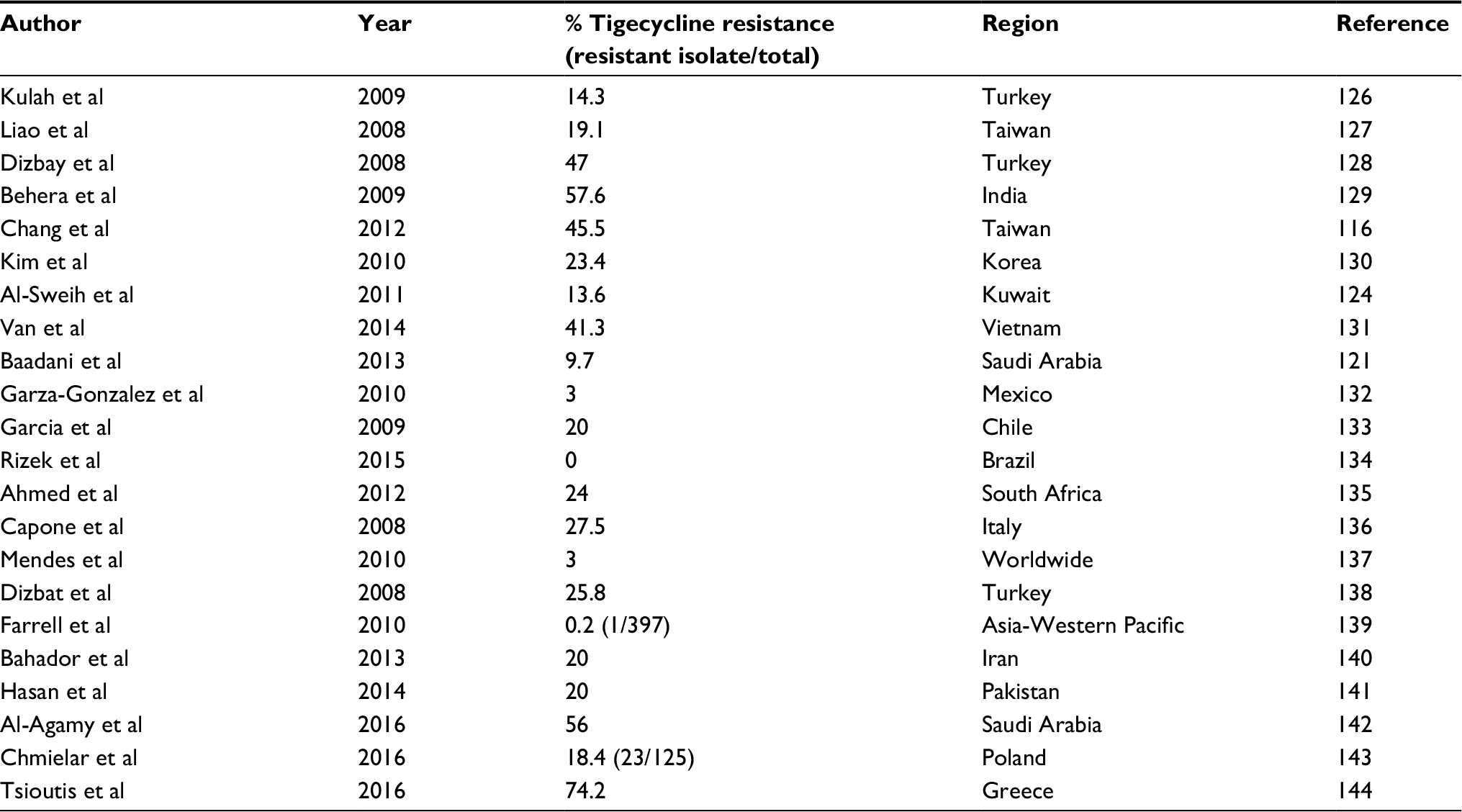 | Table 2 Studies showing tigecycline resistance by Acinetobacter in different regions of the world |
Mechanism of resistance
Enzyme-mediated degradation (beta-lactamases), genetic manipulations (mutations, acquiring or leaving a gene, upregulation or downregulation of gene expression), and efflux pumps are different strategies adopted by Acinetobacter to escape from destruction of antibiotics.77
Resistance to beta-lactams
Resistance to beta-lactam antibiotics is mediated through enhanced degradation by beta-lactamases, alteration in penicillin-binding proteins, changes in outer membrane porins for decreased permeability, and expulsion of antibiotics out of cell through efflux pump (Figure 1-I).78 Among beta-lactamases, ampC cephalosporinase or molecular class c beta-lactamase is more prevalent in A. baumannii.79 It is encoded by bla gene and confers resistance to penicillins and narrow- and extended-spectrum cephalosporins. Other beta-lactamases include class A beta-lactamases such as extended-spectrum beta-lactamases (PER-1, VEb-1, CTX-M, TEM, SHV), class B beta-lactamases such as metallo-beta-lactamases (MBLs; IMP, SIM, VIM), and class D beta-lactamases such as OXA.80 Acquired OXA-type carbapenemases are the mainstay against carbapenems, a treatment of choice, followed by MBLs.81 OXA23, OXA24, and OXA58 are plasmids that encode carbapenemases which are mainly responsible for carbapenem degradation. Coexistence of OXA23 and an MBL NDM-1, a nightmare in the history of antibiotic resistance, has been reported.82 All other types of OXA are chromosome mediated and include OXA25, OXA26, and OXA40.
 | Figure 1 Different mechanisms of resistance in A. baumannii: (I) beta-lactams; (II) aminoglycosides; (III) quinolones; (IV) colistin. Abbreviations: AME, aminoglycoside modifying enzyme; LPS, lipopolysaccharide; OMP, outer membrane porin; PBP, penicillin-binding protein. |
Reduced entry of drugs via outer membrane proteins (OMPs) or porins and modification of penicillin-binding proteins (PBPs) are implicated in resistance to beta-lactams. Many studies suggested reduced expression of OMPs and PBP2 results in carbapenem resistance. Among OMPs, a 43 kDa protein oprD and a 29 kDa protein CarO are the most studied porins that support the hypothesis of decreased expression. Probably, porins and beta-lactamase work collectively in conferring resistance.83
Presence of efflux pumps confers resistance to multiple classes of antibiotics. Six families of efflux pumps have been identified which include resistance nodulation cell division family, small multidrug resistance superfamily, ATP-binding cassette (ABC) family, major facilitator superfamily, multidrug toxic compound extrusion family, and recently identified proteobacterial antimicrobial compound efflux family.84 AdeABC efflux pump has been well characterized in A. baumannii and is a member of resistance nodulation cell division family that mediates resistance to many classes of antibiotics (cefotaxime, chloramphenicol, erythromycin, aminoglycosides, and fluoroquinolones). Overexpression of AdeABC also confers resistance to carbapenems.85 AbeS, a member of small MDR efflux pump, has also been identified in A. baumannii.86 AdeABC, AdeIJK, and AdeFGH are the major drug efflux pumps of ABC family. Other members of major facilitator superfamily include CraA, AmvA/AedF, and Tet(B). Members of recently discovered proteobacterial antimicrobial compound efflux family include AceI, while a representative of multidrug toxic compound extrusion family includes AbeM pump.87
Resistance to aminoglycoside
The most frequent mechanism of aminoglycoside resistance is the modification of amino or hydroxyl group by aminoglycoside modifying enzymes. All types of aminoglycoside modifying enzymes (adenylases, acetylases, methyltransferases, and phosphotransferases) have been identified in Acinetobacter. Reduced drug entry and alteration in target ribosomal protein are the other mechanisms involved in aminoglycoside resistance (Figure 1-II).88
Resistance to quinolones
Major mechanism for resistance to quinolones is mutations in the gyrA and parC genes, which results in phenotypic changes in DNA gyrase and topoisomerase IV, leading to reduced drug affinity.89 Drug influx and efflux system encoded by chromosomal DNA mediates reduced expression of OMPs involved in drug influx and increased expression of efflux proteins resulting in active drug expulsion; these are also responsible for quinolone resistance.90 Plasmid-encoded quinolone resistance determinants qnrA, qnrB, and qnrS have also been identified in A. baumannii that protect DNA by inhibiting binding of quinolones to DNA gyrase and topoisomerase (Figure 1-III).91,92
Mechanism of colistin resistance
The mechanisms of resistance to colistin are encoded by chromosomal DNA of the bacteria. Two major mechanisms have been reported. The first mechanism consists of mutations in lipid A encoding genes (lpxA, lpxC, and lpxD), resulting in loss of LPS, an outer part of Gram-negative organisms and an initial target of colistin.93 The second mechanism involves the two-component system of pmrAB, which is a response regulator and sensor kinase. It senses the environmental conditions (pH, Mg2+, and Fe3+) and, in response, regulates the expression of genes involved in lipid A synthesis. Point mutations in pmrA and pmrB upregulate their gene expression, resulting in remodeling of outer membrane (Figure 1-IV).94 Recently, a plasmid-mediated colistin resistance gene, mcr-1, has been reported in Escherichia coli.95,96 Although mcr-1 gene has not been identified in A. baumannii, still it is speculated that progression from MDR Acinetobacter to pan drug resistance is unavoidable due to the arrival of transmissible colistin resistance mechanisms.97
Alternate modalities
The dilemma of rapidly increasing antibiotic resistance with minimal options left in hand has steered the scientific community to think beyond antibiotics. In the last decade, renaissance of research in the field of different alternatives to antibiotics has intensified. Previously neglected modalities with therapeutic potential against MDR bacteria cannot be left apart. Bacteriophages are the best example of neglected modality. Bacteriophages and their encoded products such as lysins are extensively being studied as an alternative to antibiotics. The wild-type bacteriophages and their enzymatic products act in a manner like antibiotics and destroy target bacteria. The first report of isolation and characterization of phages against A. baumannii was published in 2010. The phages AB1 and AB2 have specifically shown lytic behavior against A. baumannii. Since then, many lytic phages have been isolated, characterized, and sequenced. The bulk of in vitro studies and characterization of phages against A. baumannii urged a dire need to test their in vivo efficacy and pharmacodynamics to fight back infectious diseases.98,99
Monoclonal antibodies are one alternative that can be used to treat A. baumannii infections. They bind to virulence factors of pathogens and neutralize them. It seems prudent to use them as an alternative due to their well-studied phenomena and clinical outcomes. However, their production is too expensive to be used for treating infections. Probiotics are live bacteria that exert a healthy effect on humans. They act by competing for the pathogen in the acquisition of nutrition and space for colonization; however, their exact mechanism of action is under study.100
Antimicrobial peptides (AMPs) or short AMPs are produced by various eukaryotic and prokaryotic organisms as their part of innate host immune response. They hold the potential to kill bacteria, so interest in AMPs as an alternative to antibiotic is increasing day by day. They are broad spectrum in nature, have low immunogenicity, low resistance, and carry a solution of antibiotic resistance for Gram-positive as well as Gram-negative bacteria. Several peptides having in vitro and in vivo activities against A. baumannii have been reported. A hybrid of cecropin A and melittin has shown activity in peritoneal sepsis by pan drug-resistant strain of A. baumannii in an animal model of infection. Brevinin 2, alyteserin 2, and catonic α-helical peptides have also demonstrated bactericidal activity against A. baumannii. A proline-rich peptide A3-APO has exhibited greater efficacy in controlling A. baumannii bacteremia in comparison to imipenem in a mice model. A short D-enantiomeric peptide D-RR4 protected the Caenorhabditis elegans model of infection from lethal infection by A. baumannii. Many successful reports exist in literature about potential of AMPs against such a robust organism, but factors such as cytotoxicity, moderate activity, enzymatic degradation, and high productivity cost need to be evaluated prior to concluding about their systemic use as an antibiotic.100,101
Gene editing technique by using clustered, regularly interspaced short palindromic repeat (Cas) system to knock out the resistance gene and make it labile to antimicrobial therapy is another possible way to nib such bugs.
Metal chelators that are essential in the expression of bacterial virulence factors, such as iron, zinc, and manganese, can be a promising target for designing newer antimicrobial drugs. Artificial nanoparticles made of lipids known as “liposomes” that closely resemble the membrane of host cells can act as decoys for bacterial toxins, and so are able to sequester and neutralize them.102,103
Conclusion
A. baumannii has emerged as an established nosocomial pathogen and exhibits a higher level of resistance to many antibiotics. Extensively drug resistant and pan drug resistant isolates are routinely being reported in various medical facilities. Carbapenems, the drug of choice to treat A. baumannii infections, are increasingly being ineffective due to higher resistance rates. Even resistance to newer antimicrobial tigecycline is emerging rapidly. Historically discarded drug colistin is left as the last resort antimicrobial, but resistance against this drug is also being reported all over the world at higher rates. Such vanishing treatment options have steered up the scientific community to look for an alternative to antibiotics. These alternatives are the dire need of the time and hopefully will be available in future.
Author contributions
All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.