Back to Journals » Clinical Interventions in Aging » Volume 15
Influence of Ageing on Vascular Reactivity and Receptor Expression in Rabbit Aorta: A Complement to Elastocalcinosis and Smooth Muscle Mechanisms
Authors Cupitra NI ![]() , Calderón JC
, Calderón JC ![]() , Narvaez-Sanchez R
, Narvaez-Sanchez R
Received 26 October 2019
Accepted for publication 26 February 2020
Published 20 April 2020 Volume 2020:15 Pages 537—545
DOI https://doi.org/10.2147/CIA.S236173
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Richard Walker
Nelson Ivan Cupitra, Juan C Calderón, Raul Narvaez-Sanchez
Physiology and Biochemistry Research Group-PHYSIS, Faculty of Medicine, University of Antioquia, Medellin, Colombia
Correspondence: Raul Narvaez-Sanchez Carrera 51 D #62-29, Office 203, Medellin, Colombia
Tel +57 42196030
Email [email protected]
Aim: To contribute to the knowledge about the mechanisms involved in aortic stiffness due to ageing.
Materials and Methods: Aortic rings from young (1.5± 0.5 months, 0.8± 0.2 kg), adult (6± 0.5 months, 2.7± 0.5 kg) and old (28± 8 months, 3.2± 0.8 kg) male New Zealand rabbits were used to evaluate: 1) intima-media thickness by optical microscopy; 2) vascular reactivity (VR) in terms of sensitivity (pD2) and efficacy (Emax) to KCl; phenylephrine (PE); U-46619, a thromboxane A2 receptor agonist, TXA2; carbachol (CCh), isoproterenol and sodium nitroprusside (SNP), using organ bath experiments; and 3) the expression of receptors α 1, β 2 and thromboxane-prostanoids (TP), by immunofluorescence.
Results: Ageing 1) did not change the thickness of tunica; 2) significantly reduced the pD2 to KCl, increased the pD2 to PE and reduced both the pD2 and Emax to TXA2, CCh and isoproterenol, and reduced the pD2 to SNP; and 3) significantly increased the expression of α 1 and β 2 receptors in the intima and adventitia, and the expression of TP only in the adventitia.
Conclusion: Our results suggest that ageing makes the aorta more reactive to α 1 adrenergic contraction, and it could be a compensation for lower responsiveness to prostanoids. The aged aorta is less reactive to endothelium-dependent and non-dependent relaxation, and the vessel seems to try to compensate for that stiffness increasing β 2 receptors, although probably less functional. These results complement the proposed mechanisms of elastocalcinosis and smooth muscle rigidity, expanding the vision that should guide the treatment of aortic stiffness due to aging.
Keywords: ageing, aortic rings, vascular reactivity, vascular smooth muscle, vascular stiffness
Introduction
Ageing is a natural process that predisposes individuals to cardiovascular disease (CVD), which is responsible for the highest percentage of deaths in the world, reaching 31%.1,2 As the population of older adults expands, elucidating the mechanisms of vascular dysfunction with age is critical to better direct appropriate and measured pharmacological and lifestyle interventions against CVD risk. Increased aortic stiffness is an important feature of aging and of CVD and is mainly attributed to changes in the extracellular matrix (ECM), sometimes called elastocalcinosis.3,4 Complementarily, Qiu et al proposed a mechanism due to smooth muscle stiffness.5 But ageing affects vascular reactivity (VR) too, by altering the sensitivity and maximal effect of stimulating contractile agonists and endogenous vasodilators.6,7 It is feasible that the vascular stiffness is due not only to one mechanism but also to the complementarity of several, and one very important to take into account would be the change in the VR. Further, in diagnosing or research, it is important to differentiate the possible changes in the blood vessel caused by a disease from those caused by the ageing of the vessels. Moreover, VR is a more plastic factor, while ECM changes require more time and could be less responsive to treatment,3 which has an impact on therapeutic strategies.
Since a study on vascular ageing in humans is challenging, the rabbit is an interesting model frequently used in research focused on cardiovascular pathophysiologies8 such as heart damage,9 hypertension10 and ageing.11 This is so because the rabbit has an over 90% homology with humans, is readily available, can be grown in controlled conditions and has a short life span. The rabbit aorta is widely used to quantify the influence of possible drugs on VR and as an initial approach that allows VR to be extrapolated to humans.12,13 As for ageing, it has been shown in rabbits that it decreases aortic relaxation by reducing β-adrenergic receptors activity.14 This decrease in vascular function with ageing has also been described in the aorta of male and female rats.15 We consider important to contribute about the mechanisms and the expression of the receptors involved in the changes of the VR evidenced by the research in rabbits and other models such as mice and rats.
Then, the goal of this study is to characterize the changes in the kinetics of rabbit aortic ring response as a function of ageing and to clarify the mechanisms involved, using one receptor-independent contraction inducer (potassium chloride, KCl), two receptor-dependent contraction inducers (phenylephrine (PE) and U-46619, a thromboxane A2 agonist, TXA2), two receptor-dependent relaxation inducers (carbachol (CCh) and isoproterenol) and independent relaxation inducer (sodium nitroprusside dihydrate, SNP). Complementarily, the wall thickness and expression of the receptors involved in some pathways were measured.
Materials and Methods
Animal Model and Reagents
Ten young (“YO”; 1.5±0.5 months, 0.8±0.2 kg), 10 adult (“AD”; 6±0.5 months, 2.7±0.5 kg) and 7 old (“OL”; 28±8 months, 3.2±0.8 kg) male New Zealand rabbits were obtained from the cuniculture facility of the National Learning Service at La Salada (Antioquia, Colombia). The animals were housed in metal cages, with a room temperature of 21±2°C, under 12 hours of light and with free access to normal chow and water. The use of rabbits was approved by the Animal Experimentation Committee of the University of Antioquia (Medellin, Colombia), act 122 of February 5, 2019 in accordance with law 84 of 1989 “National Statute for Animal Protection” and resolution 8430 of 1993 “Scientific, technical and administrative standards for health research”.
Carbamoylcholine chloride (212385-M), phenylephrine hydrochloride (P6126), and isoproterenol bitartrate (I2760), sodium nitroprusside dihydrate (71778) were from Sigma–Aldrich Co. (St. Louis, MO, USA). U-46619 (ab144540) was from Abcam (Cambridge, USA). Stocks were prepared in distilled water with 10 µM ascorbic acid in order to prevent oxidation. All antibodies were from Abcam (see below).
Dissection
Each animal was anaesthetized with intraperitoneal sodium thiopental (60 mg/kg) with subsequent cervical dislocation. The descending aorta was rapidly dissected, without the branching zones, and conserved in Krebs-Henseleit (KH) solution, with the following composition (mM): NaCl, 118; KCl, 4.7; KH2PO4, 1.2; MgSO4, 1.2; NaHCO3, 25.0; glucose, 11.1; CaCl2, 2.5; pH 7. Under magnification, excess peripheral tissue was removed from the aortic segment which was cut into 10 rings, 3–5 mm in length. Four rings were used for relaxation and contraction experiments that began within the first hour after the sacrifice. Four rings were preserved for 24 hours in KH solution previously bubbled with carbonated gas (95% O2–5% CO2) inside a cooler at 4°C and were used only for contraction studies. Other studies have shown that there are no differences in vascular reactivity at 24 hours of preservation in KH at 4°C.16 The remaining two rings were embedded in Shandon Cryomatrix (Thermo Fisher Scientific, USA) and frozen in liquid nitrogen for immunohistochemistry. Previous results from our laboratory showed that in this type of assay, there is no significant difference with respect to fresh samples.
Vascular Reactivity
The rings were placed in an LE13206 PANLAB organ bath (Barcelona, Spain) with 10 mL of KH at 37°C and bubbled with carbogen, as described.17 The contractile response was digitized with a Powerlab PL-3504 (Sydney, Australia) and analysed with the dose–response module of LabChart 8 software. Each preparation was stabilized at 2 g of resting tension for 90 minutes before initiating the agonist stimuli; during this period, the KH solution was replaced every 20 minutes. After stabilization, the arterial ring was stimulated twice with 4×10−2 M KCl, recording the response for 10 minutes, and washed for 20 minutes between each stimulus. Then, 1×10−6 M PE and 1×10−5 M CCh were applied to the bath to confirm the presence of the endothelium; this was performed for all arterial rings. Then, the evaluation was started by applying a single dose of 4×10−2 M KCl; a cumulative dose of KCl ranging from 5 to 60 mM; a cumulative dose of PE ranging from 1×10−9 M to 1×10−5 M; and a cumulative dose of U-46619 ranging from 1×10−10 M to 1×10−6 M. Relaxation studies were performed with 1×10−6 M PE precontracted rings, using CCh at doses ranging from 1×10−9 M to 1×10−5 M, isoproterenol at doses ranging from 1×10−9 M to 1×10−5 M, SNP at doses ranging from 1×10−10 M to 1×10−5 M. The VR to isoproterenol was measured only in relaxation and was suspended at the onset of contraction because precontraction with an α1 agonist could have affected this contractile VR.
Histology and Immunofluorescence
Frozen arteries were cut in 10-μm thick sections. For histology, they were fixed in Carnoy’s solution (ethanol, chloroform and acetic acid, 6:3:1) and stained with haematoxylin and eosin to evaluate changes in morphology, and with Van Gieson and a modified Van Gieson (with orcein) to qualitatively evaluate collagen and elastic fibers, respectively. Evaluation of the images was performed under a 20× objective. ImageJ v1.52a was used to process the images by quantifying the thickness of the intima-media in 2 random regions for each image.
For immunofluorescence, the sections were fixed in acetone, permeabilized with 0.1% Triton X-100 and blocked with 5% bovine serum albumin in phosphate-buffered saline. Different sections were incubated with the following primary antibodies at 4°C overnight (all diluted 1:1000 in blocking solution; Abcam): anti-α1 adrenergic receptor (ab3462, Abcam), anti-β2 adrenergic receptor (ab61778, Abcam), anti-thromboxane A2 (TP) receptor (ab233288, Abcam). After washing, the rings were incubated with goat anti-rabbit secondary antibody conjugated to Alexa Fluor 594 (red, 1:800 in blocking solution; ab150080, Abcam) at room temperature for 2 hours. As a control, one section from each experimental group was not incubated with a primary antibody but was incubated with a secondary antibody. Images were obtained using an inverted fluorescence microscope (Axio Observer A1, Carl Zeiss), a mercury lamp (HXP120, Carl Zeiss) and a filter (64 HE MPLUM, Carl Zeiss; emission window 647/70 nm). Evaluation of the images was performed under a 20× objective. ImageJ v1.52a was used to process the images using the Split channels tool in RGB images, quantifying the average intensity of red in 3 regions of interest of the intima, media and adventitia for each image.
Statistical Analysis
The results are presented as the mean ± SEM of the maximum contraction (Emax) and the negative logarithm of the molar concentration of agonist producing 50% of the maximum response (pD2). pD2 was calculated by nonlinear regression of the concentration–response curves. The groups were compared using two-way ANOVA followed by post hoc Tukey’s test, considering a significance level of P<0.05. The statistical package GraphPad Prism 6 Statistics, version 6.01, was used.
Results
Intima-Media Thickness
In the quantification of vascular thickness, no differences were found between YO, AD and OL (YO, 126±6 µM; AD, 125.7±7.2 µM; and OL, 133±5.5 µM; n=5 for all the experiments. Figure 1; p=0.62). In addition, in the histology results by Van Gieson and modified Van Gieson (with orcein), no qualitative differences were found between the groups.
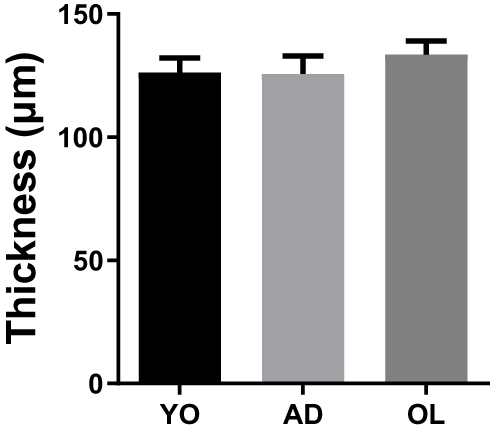 |
Figure 1 Quantification of the thickness of the vascular wall of aortic rings of YO, AD and OL rabbits. Results on average ± SEM. The number of experiments is 4 in all cases. |
Contraction
Upon stimulation with KCl, VR in the aorta showed a decreased sensitivity in AD and OL (p=0.01). However, no significant differences were found in the maximal response (Figure 2A, p=0.1). VR with PE showed significant differences in sensitivity between YO and AD compared with OL (Figure 2B; p=0.002), together with an increase in the Emax of OL versus YO (Figure 2B; p=0.01). The stimulus with U-46619 resulted in a decrease in the pD2 of OL compared with that in YO and AD (p=0.001). The maximum response to the stimulus with U-46619 was higher in YO than in AD and OL (Figure 2C; p=0.001). A summary and number of experiments can be found in Table 1.
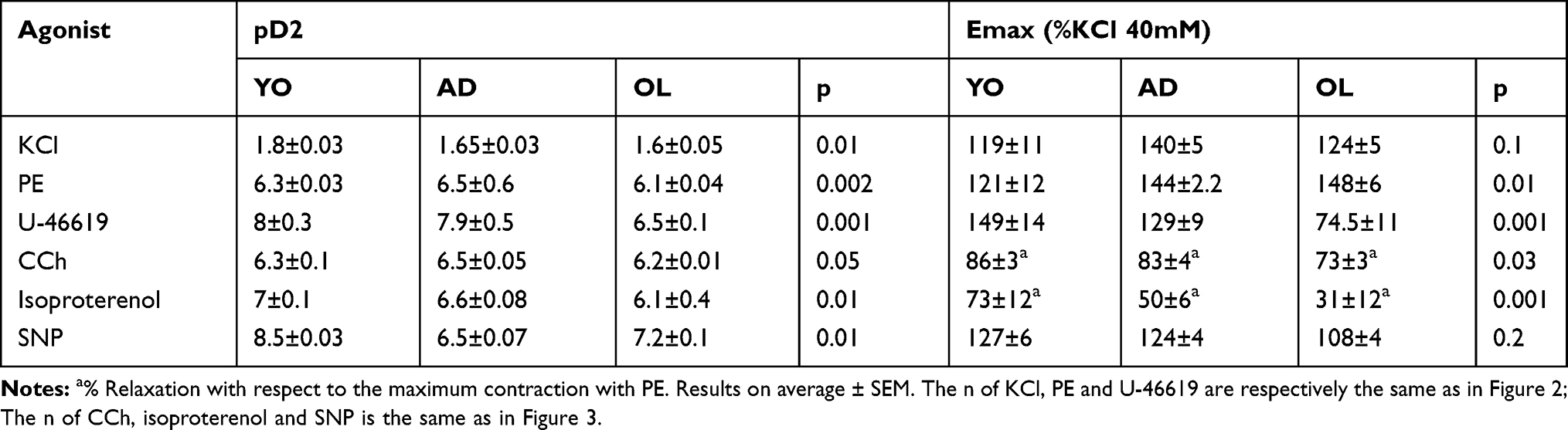 |
Table 1 pD2 and Emax to KCl, FE, U-46619, CCh Isoproterenol and SNP in Aortic Rings in YO, AD, and OL Rabbits |
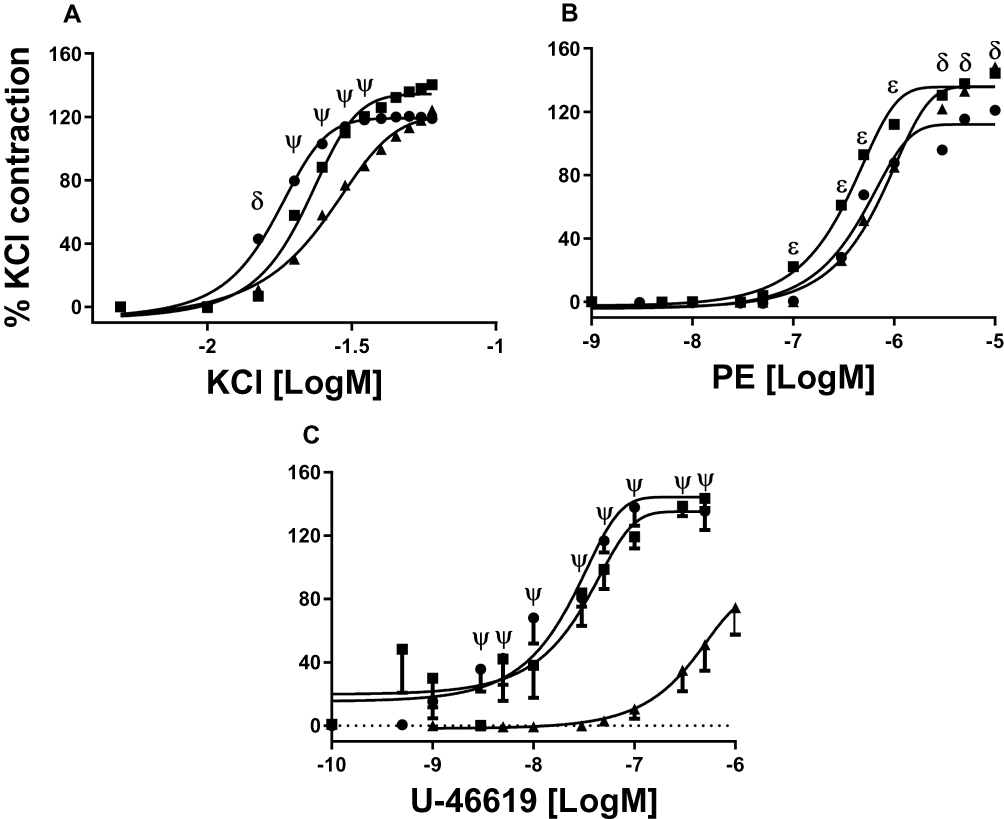 |
Figure 2 Concentration–response curves to KCl (A), PE (B) and U-46619 (C). YO (●, 4 animals, 8 rings), AD (■, 4,8) and OL (▲, 4,8). δ indicates a higher sensitivity to KCl and lower Emax to PE in YO arteries, compared to AD and OL. ψ indicates a higher sensitivity to KCl and to U-46619, and a higher Emax to U-46619, in YO and AD arteries compared with OL. In panel B, ε indicates a higher sensitivity to PE in AD, compared to YO and OL. pD2 and Emax values in all conditions can be seen in Table 1. Significance level of P<0.05. |
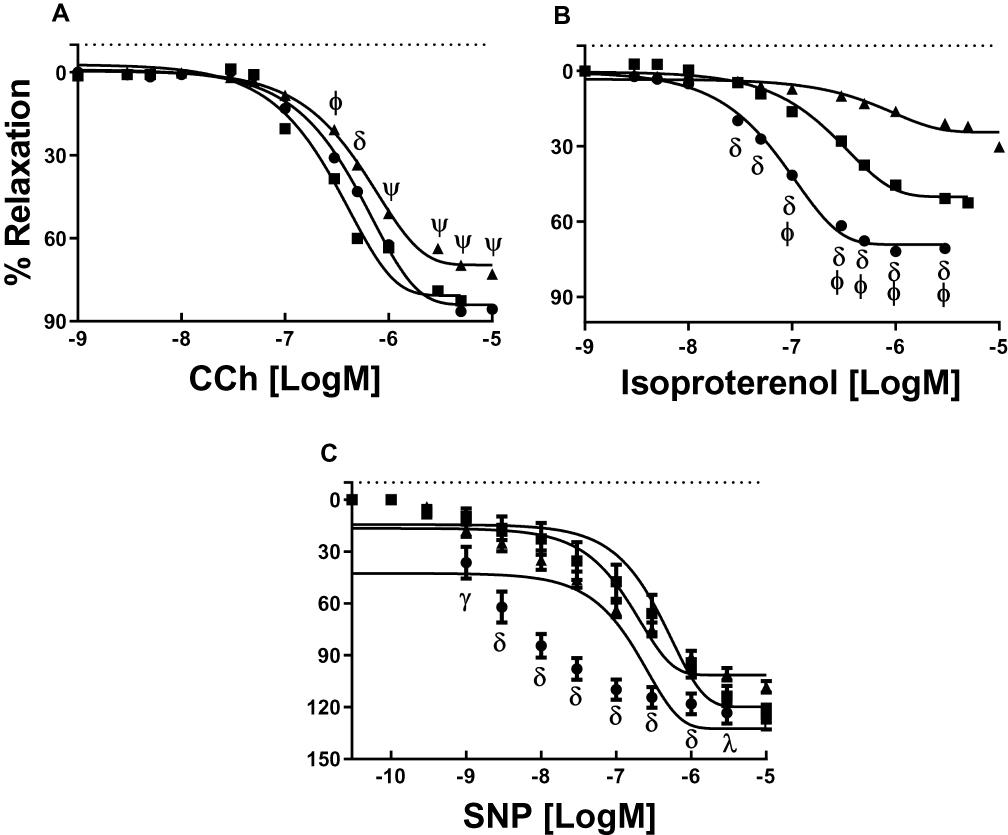 |
Figure 3 Concentration–response curves to CCh (A), isoproterenol (B) and SNP (C). YO (●, 4 animals, 8 rings), AD (■, 4,8) and OL (▲, 4,8). δ indicates a lower sensitivity in YO to CCh than AD and higher than OL, and a higher sensitivity to isoproterenol and SNP and higher Emax to isoproterenol in YO compared with AD and OL. ψ indicates a higher sensitivity to CCh in YO and AD compared with OL. ϕ indicates a higher sensitivity to CCh and isoproterenol and higher Emax to isoproterenol in AD compared with YO and OL. γ indicates a lower sensitivity in AD to SNP than YO. λ indicates a lower relaxation in OL to SNP than YO. pD2 and Emax values in all conditions can be seen in Table 1. Significance level of P<0.05. |
Relaxation
In the CCh-induced endothelium-dependent relaxation showed a decreased sensitivity (Table 1; p=0.05) and maximal response in OL (Figure 3A; p=0.03). When stimulated with isoproterenol showed significant differences in sensitivity between YO compared with AD and OL. The maximal relaxation was found decreased in OL (Emax: YO, 73±12%; AD, 50±6%; and OL, 30±3% of maximum relaxation) compared to the other groups (Figure 3B, p=0.001). VR with SNP showed significant differences in sensitivity between YO compared with AD and OL (Figure 3C; p=0.01). However, no significant differences were found in the maximal response (Figure 3C; p=0.2). A summary and number of experiments can be found in Table 1.
Immunofluorescence
The expression of receptors quantified by fluorescence in rabbit aorta was normalized with respect to YO (Figure 4A). For the α1 receptor, the values for the intima were 1±0.2 for YO (n=4 for all the experiments), 2.3±0.9 for AD and 3±0.5 for OL, with no significant differences among the groups (Figure 4B, p>0.05); for the media, the values were 1±0.5 for YO, 3.44±1.3 for AD and 4.04±2.4 for OL, without significant differences among the groups (Figure 4B, p>0.05); and for the adventitia, the values were 1±0.3 for YO, 20.7±10.2 for AD and 13.4±5.8 for OL, with significant differences between YO and AD and between YO and OL (Figure 4B, p<0.01). For the β2 receptor, the values for the intima were 1±0.6 for YO, 1.9±0.55 for AD and 5.2±1.4 for OL, with significant differences between YO and OL and between AD and OL (Figure 4C, p<0.05); for the media, the values were 1±0.7 for YO, 0.65±0.13 for AD and 1.08±0.4 for OL, with no significant differences among groups (Figure 4C, p>0.05); and for the adventitia, the values were 1±0.8 for YO, 1.4±0.2 for AD and 4.3±1.2 for OL, with significant differences between YO and OL and between AD and OL (Figure 4C, p<0.01). For the TP receptor, the values for the intima were 1±0.2 for YO, 1.7±0.8 for AD and 0.75±0.3 for OL, without significant differences among the groups (Figure 4D, p>0.05); for the media, the values were 1±0.4 for YO, 1.75±0.6 for AD and 1.2±0.4 for OL, without significant differences among the groups (Figure 4D, p>0.05); and for the adventitia, the values were 1±0.5 for YO, 5.7±1.9 for AD and 7.6±2 for OL, with significant differences between YO and AD and between YO and OL (Figure 4D, p<0.01).
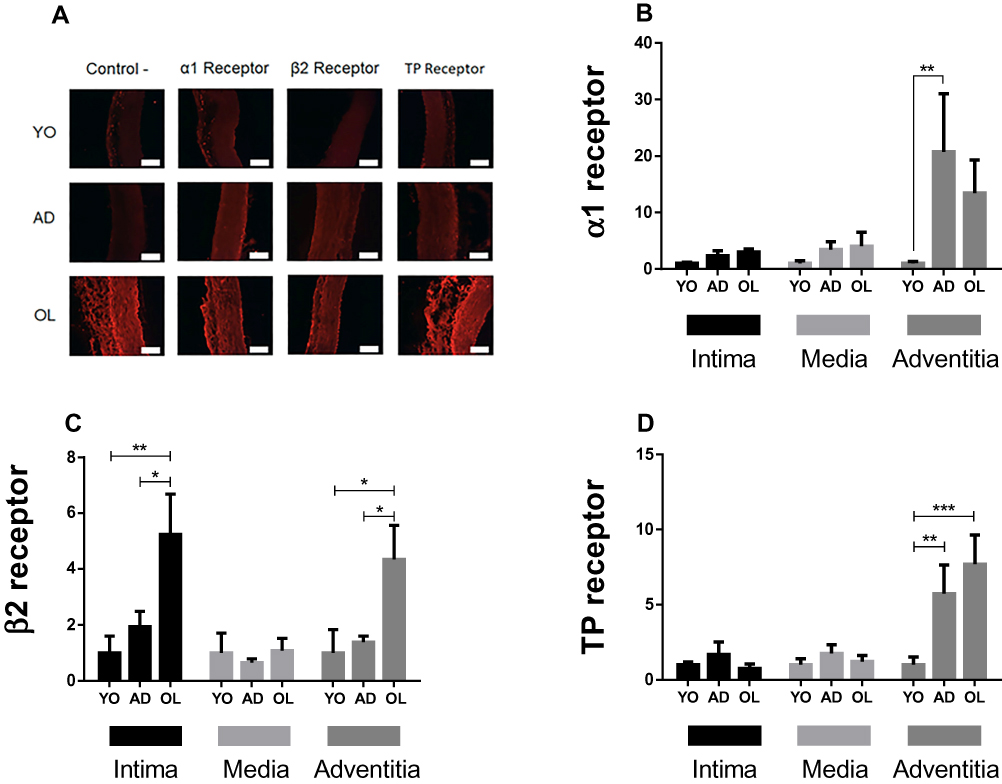 |
Figure 4 Quantification of protein expression of α1, β2 and thromboxane A2 (TP) receptors in the aortic wall of YO, AD, and OL rabbit. (A) representative images of immunofluorescence to identify α1, β2 and thromboxane A2 (TP) receptors. (B–D) Quantification of the fluorescence of the α1, β2 and TP receptors, respectively, in the tunica of the aortic wall. The number of experiments is 4 in all cases. (A) The length of the white calibration bars are equivalent to 100 μm. *Indicates p<0.05. **p<0.01. ***p<0.001. The expression was always normalized to YO. |
Discussion
We assessed the effect of ageing on structure, VR and receptor expression in rabbit aorta. Our main findings were as follows: 1) there were no structural changes in the ageing aorta, 2) ageing causes a decrease in the sensitivity of the aorta to KCl, U-46619 and isoproterenol, and 3) there was an increase in the adrenergic response with ageing, associated to an increased α1 receptor expression.
Structural Vascular Changes with Ageing
We performed a histological approach in order to establish whether there was vascular remodeling attributable to ageing, but we found no differences in intima-media thickness, as showed in Figure 1. We stained with Van Gieson and a modified Van Gieson (with orcein) to qualitatively evaluate collagen and elastic fibers, respectively, without finding a difference either.
Functional Vascular Changes with Ageing
We demonstrated that ageing causes a decrease in the sensitivity of aortic smooth muscle to receptor-independent agonists; however, Emax did not change. This result contrasts with,6 which showed an increase in sensitivity and Emax to KCl in SAMR1 and SAMP8 mouse aorta (senescence model). This difference in results is expected because6 used female mice in an accelerated senescence model, that could not exactly meet natural ageing processes. Another important finding of the present study is the lack of differences between adult and old subjects, in agreement with18 and,19 who showed identical results in the aorta of healthy young vs old Sprague-Dawley rats; however, they did find differences in Emax at 18 months in SHR rats (hypertensive model).
We observed an increase in the adrenergic response with ageing, consistent with the increased expression of the α1 receptor in the tunica intima and media (observed as a trend) and in the adventitia (significant increase); however, cellular diversity of this external layer makes it difficult to establish how this increase in α1 expression contributes to the changes seen in VR. The increase in adrenergic response that we observed contrasts with,20 which found decreased sensitivity to stimulation with norepinephrine (NE) in adult rat aorta, but it is probably due to NE has an α and β effect, while the PE used in the present study has specific affinity for α1. Our results also differed from,21 which did not find differences in VR to PE in rat coronary artery, but agreed with,22 which showed an increase in the α1 receptor in the human mammary artery in relation to age, and with,23 which observed an increase in the reactivity of the renal artery to the sympathetic stimulus in humans over 50 years of age. Notably, the results involve different vascular beds and different species. The diversity of results justifies new complementary studies in arteries of distribution of various species, for example, in the mesenteric arteries of humans, rabbits and rats, to further clarify this scenario.
The VR to the TXA2 analogue, normalized to KCl, showed a marked difference in contraction between young and old rabbits and between adult and old rabbits, suggesting that sensitivity to prostanoids remains until six months of development in rabbits, which would be equivalent to approximately 14 human years.13 However, the sensitivity deteriorates in old age. This result is reinforced by the tendency to decrease in the expression of the TP receptor in the intima and media after adulthood, and coincides with that reported in the female SAMR1 and SAMP8 mouse aorta, attributing this increase in sensitivity and maximum effect to the stimulus with U-44619 to a decrease in the availability of nitric oxide and increase in contractile prostanoid synthesis.6 Strikingly, there is a significant increase in the expression of the TP receptor in the vascular adventitia, but again, the cellular diversity of this external layer makes it difficult to define how this increase in TP contributes to the changes seen in VR. Our results for the intima match those reported in,24 which did not find differences in the expression of the TP receptor in endothelial cells of SHR and WKY rats between 36 and 72 weeks, and contrasts with those reported in,25 which did not find differences to the stimulus with U-46619 in male 6 and 22 months old Fischer F344 rat aorta, and with those reported in,26 which did not find differences with the same agonist in rat cerebral arterioles. These results suggest that this contrast may be due to the animals and vessels used, in addition to different ages, but could also be related to the amount of perivascular tissue removed during the cleaning and mounting procedure.
We add that such changes in VR, at least in the aorta, may have other origins, for example, in the dynamics of phosphorylation at the cellular level, the availability of second messengers and other downstream mechanisms, which we did not explore but are reviewed in.3
It is important to take into account that the gain in contraction could be accentuated by the decrease in endothelial compensation.24 In our relaxation results, VR with CCh showed a tendency to decrease sensitivity to stimulation in old rabbits, coinciding with studies that showed decreased endothelial-dependent vasodilatory response to stimulation with acetylcholine or CCh in coronary21 and mesenteric27 arteries related to ageing in rats. This deterioration of relaxation is usually attributed to oxidative processes in the vascular wall.
In the present study, when we applied isoproterenol, a vascular relaxant that acts through the β2 receptor, we found a significant decrease in sensitivity and a maximum effect that was accentuated as age progressed, in contrast with the increase in β2 receptor expression in the endothelium and adventitia, suggesting that there is less β-adrenergic receptor activity as an individual ages, as discussed in14 and.22 It is feasible that the vessel responds by increasing the number of β receptors in adults and older patients especially in the endothelium and adventitia; however, each unit is relatively less functional, at least in this vascular bed. These results are similar to those reported in,28 which showed a decrease in the vasorelaxant response in the human coronary artery in relation to ageing, although this decrease is much more pronounced in response to an endothelium-dependent vasodilator than to an endothelium-independent vasodilator. Additionally, our results would contribute to understanding the reduction of vascular stiffness when sympathetic denervation is made29 although apparently not when baroreflex activation therapy is performed,30 suggesting that vascular stiffness does not depend exclusively on sympathetic control.
When testing the non-endothelium-dependent relaxation response, the present work evidenced a marked decrease in SNP sensitivity in AD and OL with respect to YO, suggesting a reduction in the ability of vascular smooth muscle to relax in aging, complemented by a structural change of the vessel that makes it more rigid, known as elastocalcinosis.3,4 These results contrast with studies that did not find such differences in the SNP response between young and old animals, in soleus feed arteries of Fisher 344 rats,31 in aorta of Wistar rats32 and in carotid of Albino rats.33 The evidence strongly suggests that stiffening due to aging occurs differently depending on the vascular territory, as,34 co-authored by the same author of,31 showed in aorta, iliac, femoral and gastrocnemius feed arteries of Fisher 344 rats, that deterioration of the dilatation to the SNP did occur in large/elastic arteries, but not in resistance arteries. According to this, and to our results, vascular stiffening due to aging is not only due to endothelial changes but to a structural and functional changes of the different layers of the artery. Additional studies are required to characterize the involved mechanisms.
Conclusion
In a rapidly ageing world, it is critical to know more about the mechanisms by which blood vessels harden with age. Our results suggest that ageing makes the aorta more rigid not only mechanically but, in addition, more reactive to α1 adrenergic contraction, perhaps as compensation for lower responsiveness to the prostanoid contractile pathway. In the same sense, the aorta is less reactive to endothelium-dependent and non-dependent relaxation, which could not be attributable only to oxidative stress. The vessel seems to try to compensate for that stiffness with increased receptors, with little success due to their probable lower functionality. The evidence here presented complements the view that there is a deterioration of the ECM (elastocalcinosis) and an increase in the rigidity of vascular smooth muscle. In this way, we hope to contribute to a better rationale of lifestyle and pharmacological interventions which attempt to treat/prevent CVD. Our quantification could also contribute to improving the comparability of vascular outcomes between groups of people with different ages. In addition, if experimental animals are used, our results insist on the necessity to ensure that the age of the animals used is equivalent to the age range of incidence of the disease in humans, to decrease a probable bias when extrapolating the findings.
Acknowledgments
We want to thank the project “High Level Human Talent Training” approved by the Science, Technology and Innovation Fund (CTeI) of the General Royalty System (SGR)-BPIN 2013000100103, Tolima Governorate and University of Tolima, Colombia; the CODI grants, 2014-University of Antioquia; and the Foundation for the Promotion of Research and Technology, 2017-Bank of the Republic of Colombia.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Childs BG, Durik M, Baker DJ, Van Deursen JM. Cellular senescence in aging and age-related disease: from mechanisms to therapy. Nat Med. 2015;21(12):1424. doi:10.1038/nm.4000
2. WHO. WHO | cardiovascular diseases (CVDs). 2018. Available from: https://www.who.int/cardiovascular_diseases/en/.
3. Donato AJ, Machin DR, Lesniewski LA. Mechanisms of dysfunction in the aging vasculature and role in age-related disease. Circ Res. 2018;123(7):825–848. doi:10.1161/CIRCRESAHA.118.312563
4. Dao HH, Essalihi R, Bouvet C, Moreau P. Evolution and modulation of age-related medial elastocalcinosis: impact on large artery stiffness and isolated systolic hypertension. Cardiovasc Res. 2005;66(2):307–317. doi:10.1016/j.cardiores.2005.01.012
5. Qiu H, Zhu Y, Sun Z, et al. Short Communication: vascular smooth muscle cell stiffness as a mechanism for increased aortic stiffness with aging. Circ Res. 2010;107(5):615–619. doi:10.1161/CIRCRESAHA.110.221846
6. Novella S, Dantas AP, Segarra G, et al. Aging enhances contraction to thromboxane A 2 in aorta from female senescence-accelerated mice. Age. 2013;35(1):117–128. doi:10.1007/s11357-011-9337-y
7. Hongo K, Nakagomi T, Kassell N, et al. Effects of aging and hypertension on endothelium-dependent vascular relaxation in rat carotid artery. Stroke. 1988;19(7):892–897. doi:10.1161/01.STR.19.7.892
8. Kokozidou M, Katsargyris A, Verhoeven EL, Schulze-Tanzil G. Vascular access animal models used in research. Ann Anat. 2019;225:65–75. doi:10.1016/j.aanat.2019.06.002
9. Lapenna D, Ciofani G, Pierdomenico SD, Giamberardino MA, Porreca E. Iron status and oxidative stress in the aged rabbit heart. J Mol Cell Cardiol. 2018;114:328–333. doi:10.1016/j.yjmcc.2017.11.016
10. Ning B, Chen Y, Waqar AB, et al. Hypertension enhances advanced atherosclerosis and induces cardiac death in Watanabe heritable hyperlipidemic rabbits. Am J Pathol. 2018;188(12):2936–2947. doi:10.1016/j.ajpath.2018.08.007
11. Orlandi A, Francesconi A, Marcellini M, Ferlosio A, Spagnoli LG. Role of ageing and coronary atherosclerosis in the development of cardiac fibrosis in the rabbit. Cardiovasc Res. 2004;64(3):544–552. doi:10.1016/j.cardiores.2004.07.024
12. Russell JC, Proctor SD. Small animal models of cardiovascular disease: tools for the study of the roles of metabolic syndrome, dyslipidemia, and atherosclerosis. Cardiovasc Pathol. 2006;15(6):318–330. doi:10.1016/j.carpath.2006.09.001
13. Woodruff-Pak DS. Aging and classical conditioning: parallel studies in rabbits and humans. Neurobiol Aging. 1988;9:511–522. doi:10.1016/S0197-4580(88)80108-8
14. Fleisch JH, Hooker CS. The relationship between age and relaxation of vascular smooth muscle in the rabbit and rat. Circ Res. 1976;38(4):243–249. doi:10.1161/01.RES.38.4.243
15. Sullivan JC, Loomis ED, Collins M, Imig JD, Inscho EW, Pollock JS. Age-related alterations in NOS and oxidative stress in mesenteric arteries from male and female rats. J Appl Physiol. 2004;97(4):1268–1274. doi:10.1152/japplphysiol.00242.2004
16. Buchinger-Kähler V, Stoldt VR, Muth T, Schipke JD. Function and viability of vessels in different preservation solutions-an experimental study on human great saphenous veins. J Angiol Vasc Surg. 2016;1(003):1–7. doi:10.24966/AVS-7397/100003
17. Estrada O, González-Guzmán JM, Salazar-Bookaman M, Fernández AZ, Cardozo A, Alvarado-Castillo C. Pomolic acid of Licania pittieri elicits endothelium-dependent relaxation in rat aortic rings. Phytomedicine. 2011;18(6):464–469. doi:10.1016/j.phymed.2010.10.008
18. Soltis EE. Effect of age on blood pressure and membrane-dependent vascular responses in the rat. Circ Res. 1987;61(6):889–897. doi:10.1161/01.RES.61.6.889
19. Ponte A, Sánchez-Ferrer CF, Hernández C, Alonso MJ, Marín J. Effect of ageing and hypertension on endothelial modulation of ouabain-induced contraction and sodium pump activity in the rat aorta. J Hypertens. 1996;14(6):705–712. doi:10.1097/00004872-199606000-00005
20. Feletou M, Köhler R, Vanhoutte PM. Nitric oxide: orchestrator of endothelium-dependent responses. Ann Med. 2012;44(7):694–716. doi:10.3109/07853890.2011.585658
21. Su N, Narayanan N. Age related alteration in cholinergic but not α adrenergic response of rat coronary vasculature. Cardiovasc Res. 1993;27(2):284–290. doi:10.1093/cvr/27.2.284
22. Rudner XL, Berkowitz DE, Booth JV, et al. Subtype specific regulation of human vascular α1-adrenergic receptors by vessel bed and age. Circulation. 1999;100(23):2336–2343. doi:10.1161/01.CIR.100.23.2336
23. Boddi M, Sacchi S, Lammel RM, Mohseni R, Gastone G, Serneri N. Age-related and vasomotor stimuli-induced changes in renal vascular resistance detected by Doppler ultrasound. Am J Hypertens. 1996;9(5):461–466. doi:10.1016/0895-7061(96)00027-1
24. Tang EH, Vanhoutte PM. Gene expression changes of prostanoid synthases in endothelial cells and prostanoid receptors in vascular smooth muscle cells caused by aging and hypertension. Physiol Genomics. 2008;32(3):409–418. doi:10.1152/physiolgenomics.00136.2007
25. Shams G, Wallace LJ, Miller DD, Feller DR. Effects of thromboxane A2 on thoracic aorta of young and old rats: use of selective thromboxane receptor antagonists. Pharmacology. 1990;40(1):27–32. doi:10.1159/000138635
26. Mayhan WG, Faraci FM, Baumbach GL, Heistad DD. Effects of aging on responses of cerebral arterioles. Am J Physiol Heart Circ Physiol. 1990;258(4):H1138–H1143. doi:10.1152/ajpheart.1990.258.4.H1138
27. Atkinson J, Tatchum‐Talom R, Capdeville‐Atkinson C. Reduction of endothelial function with age in the mesenteric arterial bed of the normotensive rat. Br J Pharmacol. 1994;111(4):1184–1188. doi:10.1111/j.1476-5381.1994.tb14870.x
28. Egashira K, Inou T, Hirooka Y, et al. Effects of age on endothelium-dependent vasodilation of resistance coronary artery by acetylcholine in humans. Circulation. 1993;88(1):77–81. doi:10.1161/01.CIR.88.1.77
29. Brandt MC, Reda S, Mahfoud F, Lenski M, Böhm M, Hoppe UC. Effects of renal sympathetic denervation on arterial stiffness and central hemodynamics in patients with resistant hypertension. J Am Coll Cardiol. 2012;60(19):1956–1965. doi:10.1016/j.jacc.2012.08.959
30. Gronda E, Brambilla G, Seravalle G, et al. Effects of chronic carotid baroreceptor activation on arterial stiffness in severe heart failure. Clin Res Cardiol. 2016;105(10):838–846. doi:10.1007/s00392-016-0992-y
31. Woodman CR, Price EM, Laughlin H. Selected contribution: aging impairs nitric oxide and prostacyclin mediation of endothelium-dependent dilation in soleus feed arteries. J Appl Physiol. 2003;95(5):2164–2170. doi:10.1152/japplphysiol.01073.2002
32. Kim SY, Park JT, Park JK, Lee JS, Choi JC. Aging impairs vasodilatory responses in rats. Korean J Anesthesiol. 2011;61(6):506–510. doi:10.4097/kjae.2011.61.6.506
33. Omar NM, Abbas AM, Abdel-Malek H, Suddek GM. Effect of age on the contractile response of the rat carotid artery in the presence of sympathetic drugs and L-NAME. Acta Physiol Hung. 2013;100(3):266–279. doi:10.1556/APhysiol.100.2013.3.3
34. Luttrell M, Kim H, Shin SY, Holly D, Massett MP, Woodman CR. Heterogeneous effect of aging on vasorelaxation responses in large and small arteries. Physiol Rep. 2020;8(1):e14341. doi:10.14814/phy2.14341
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.