Back to Journals » Journal of Inflammation Research » Volume 8
Inflammation-related cytokine gene polymorphisms in Behçet’s disease
Authors Al-Okaily F, Arfin M, Al-Rashidi S, Al-Balawi M, Al-Asmari A, Al-Otaibi M
Received 26 May 2015
Accepted for publication 3 July 2015
Published 28 September 2015 Volume 2015:8 Pages 173—180
DOI https://doi.org/10.2147/JIR.S89283
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Ning Quan
Fahda Al-Okaily,1 Misbahul Arfin,2 Seham Al-Rashidi,1 Maysoon Al-Balawi,1 Abdulrahman Al-Asmari2
1Department of Rheumatology, 2Division of Molecular Biology and Genetics, Research Center, Prince Sultan Military Medical City, Riyadh, Saudi Arabia
Abstract: Behçet’s disease (BD) is a complex, multisystemic inflammatory disorder of unclear etiology. Single nucleotide polymorphisms in tumor necrosis factor (TNF) and interleukin (IL)-10 genes have been implicated in susceptibility to BD with inconsistent results in several ethnic populations. The aim of this case-control study was to evaluate the association of TNF-α (−308G/A), TNF-β (+252A/G), and IL-10 (−1082G/A, −819C/T, and −592 C/A) polymorphisms with susceptibility of BD in Saudi patients. Molecular genotyping of TNF-α, TNF-β, and IL-10 gene polymorphisms was performed to analyze the alleles and genotypes distribution in 272 Saudi subjects, including BD patients (61) and healthy controls (211). The frequencies of allele A and genotype GA of TNF-α (−308G/A) were significantly higher, whereas those of allele G and genotypes GG were significantly lower in BD patients than controls, indicating that A allele and GA genotype are susceptible, while G allele and GG genotype may be refractory to BD. The distribution of frequencies of alleles and genotype of TNF-β (+252A/G) promoter polymorphism was not significantly different between BD patients and healthy controls. Genotypes 1082GG, −819TT, and 592AA of IL-10 polymorphisms are significantly associated with susceptibility risk of BD, while genotypes 1082AA, 1082GA, 819CC, 819CT, 592CC, and 592CA are resistant to BD. This study indicates that TNF-α (−308G/A) and IL-10 (−1082G/A, −819C/T, and −592C/A) polymorphisms are associated with risk of BD susceptibility in Saudi patients. However, larger scale studies in Saudi population as well as in other ethnicities are needed to confirm this association.
Keywords: tumor necrosis factor, interleukin-10, polymorphism, Saudis, Behçet's disease
Background
Behçet’s disease (BD; MIM 109650) is a multisystemic inflammatory disorder of unclear etiology, characterized by recurrent oral aphthous ulcers, ocular symptoms, skin lesions, and genital ulcerations. This systemic vasculitis is more prevalent in countries along the ancient Silk Route, mainly in the Far East, the Mediterranean, and the Middle Eastern countries.1–3 Its prevalence is highest in Turkey (80–420 cases per 100,000), with 7.6, 15, 17, 20, 22, 30.2, 80, 110, 146.4 cases per 100,000 in Egypt, Morocco, Iraq, Saudi Arabia, Japan, Korea, Iran, People’s Republic of China, and Israel, respectively.1–4
BD is a complex disease and its pathogenesis/etiology is not well established; however, environmental factors, genetic determinants, and diverse immunological responses have been implicated and studied extensively.5,6 Inflammation, a characteristic feature in BD, is thought to be mediated by cytokines.7,8 Decreased levels of interleukin (IL)-10 and increased levels of tumor necrosis factor (TNF)-α have been observed in the serum and active lesions of BD patients and has been indicated to play a significant role in the immune response, pathogenesis, and activity in BD.9–15
IL-10 and TNF-α production may be regulated at the transcriptional level, and several single nucleotide polymorphisms (SNPs) at the promoter region of IL-10 and TNF-α gene have been shown to be associated with changes in the expression levels of IL-10 and TNF-α production.16,17 On the other hand, numerous recent studies have demonstrated an association between BD and SNPs of IL1018–22 and TNF-α22–25 in different ethnic groups.
Three polymorphisms −1082 A/G (rs1800896), −819 T/C(rs1800871), and −592 A/C (rs1800872) in the promoter region of the IL-10 gene are correlated to the expression level of IL-10. Similarly, several polymorphisms within the promoter region of TNF-α and the intron 1 polymorphism of TNF-β, in particular, have been associated with altered levels of circulating TNF-α.26,27 TNF-α (−308G/A) polymorphism (rs1800629) affects a consensus sequence for the binding site of transcription factor AP-2 and leads to a less common allele-A (allele 2), which has been associated with increased TNF-α production in vitro28 and higher rate of TNF-α transcription than wild-type allele G (allele 1). Allele A has been shown to produce six- to sevenfold higher levels of TNF-α transcription.17,29,30
TNF-β, which is closely linked to TNF-α, has also been shown to contribute to the susceptibility of several inflammatory/autoimmune diseases.31–35 Higher level of TNF-β is produced by the γδ T-cells of BD patients than by those of healthy controls.36,37 A polymorphism TNF-β (+252A/G) (rs909253) has been reported to consist of a guanine (TNF-β +252G) on one allele and an adenine (TNF-β +252A) on the alternate allele. The presence of G at this position defines the mutant allele known as TNF-β*1 (allele-1) which is the less frequent allele in white subjects and is associated with higher TNF-α and TNF-β production.38,39
The candidate gene approach has been useful in identifying susceptibility and severity genes in BD.40 Besides HLA-B51 molecules, SNPs in TNF and IL-10 genes have been implicated in susceptibility to BD.24,41–43 However, the results are inconsistent and variations are found among several ethnic populations. So in this study, the association of five polymorphisms in TNF-α, -β, and IL-10 genes with BD susceptibility risk in Saudi patients was evaluated.
Methods
Subjects
A total of 272 Saudi subjects visiting Prince Sultan Military Medical City (PSMMC), Riyadh, Saudi Arabia, were recruited for this study. Sixty-one unrelated patients with BD, ranging from 20 to 64 years, and 211 unrelated healthy matched voluntary Saudi blood donors, age ranging from 20 to 60 years, were studied for polymorphisms in TNF-α, -β, and IL-10 genes. Patients with any other inflammatory/autoimmune diseases were excluded from the study. Power was calculated online (http://www.stat.ubc.ca/~rollin/stats/ssize/caco.html).
This study was approved by the research and ethical committee of PSMMC, and written informed consent was obtained from each subject before recruitment. All patients were diagnosed according to the diagnostic criteria prepared by the international study group for BD.44
Detailed information including demographics, disease duration, treatment duration, and clinical features were obtained from a review of medical records and an interview at the time of enrollment. We evaluated the clinical features such as oral ulcers, genital ulcers, skin lesions (including papulopustular and erythema nodosum-like), pathergy response, ocular inflammation, gastrointestinal lesions, articular manifestations, vascular lesions (including deep vein thrombosis, superficial thrombophlebitis, and aneurysm), central nervous system involvement, and epididymitis. Active or inactive form of BD was determined by clinical parameters.44
PCR amplification
Genomic DNA was extracted from the peripheral blood of BD patients and controls using QIAamp® DNA mini kit (Qiagen, Valencia, CA, USA). TNF-α, TNF-β, and IL-10 genes were amplified using amplification refractory mutation systems (ARMS)–polymerase chain reaction (PCR) methodology45 to detect polymorphisms at position −308 of TNF-α, +252 in Intron1 of TNF-β, and at positions −1082, −819, and −592 of IL-10 genes. PCR amplification was carried out using Ready-To-Go™ PCR Beads (GE Healthcare UK Ltd, Little Chalfont, UK). PCR consisted of ten temperature cycles of denaturation for 15 seconds at 94°C, annealing for 50 seconds at 65°C, and extension for 40 seconds at 72°C. This was followed by 25 cycles of denaturation for 20 seconds at 94°C, annealing for 50 seconds at 59°C, and extension for 50 seconds at 72°C. Final extension was performed at 72°C for 7 minutes. A positive control was included in the PCR assay by amplification of the human growth hormone gene. The amplified product for various samples were separated on a 1.5% agarose gel, stained with ethidium bromide, and then photographed. For quality control, 25% of the random blind samples were repeated for genotyping and positive and negative controls were also used in the PCR.
Statistical analysis
The differences in allele/genotype frequencies between patients and controls were analyzed by the Fisher’s exact test. P-values ≤0.05 were considered significant. The strength of the association of disease with respect to a particular allele/genotype is expressed by odds ratio, interpreted as relative risk (RR) following the Woolf’s method as out lined by Schallreuter et al.46 It was calculated only for those alleles/genotypes which were increased or decreased in BD patients as compared to control group. The RR was calculated for all the subjects using the formula given below:
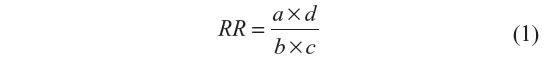
where, a = number of patients with expression of allele or genotype, b = number of patients without expression of allele or genotype, c = number of controls with expression of allele or genotype, and d = number of controls without expression of allele or genotype.
Etiologic fraction (EF) indicates the hypothetical genetic component of the disease. The values 0.0–0.99 are of significance. EF was calculated for positive association only, where RR >1 using the following formula:47
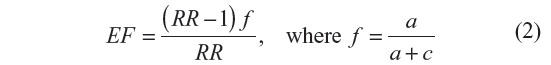
Preventive fraction (PF) indicates the hypothetical protective effect of one specific allele/genotype for the disease. PF was calculated for negative association only, where RR <1 using the following formula:47
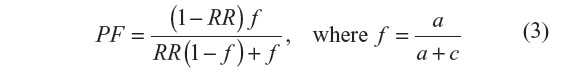
Values <1.0 indicate the protective effect of the allele/genotype against the manifestation of disease.
Results
Demographic data and clinical characteristics of patients with BD are presented in Tables 1 and 2. The number of controls per case was 3.4, which yielded a power of 90%. The most frequently found clinical symptoms in BD patients were oral, genital, ocular, musculoskeletal, and cutaneous followed by gastrointestinal, nervous, renal, vascular, and pulmonary symptoms, whereas cardiovascular symptoms were absent in our BD patients. Among BD patients, the male to female ratio was 43:18 (2.4:1). There was no significant difference in clinical manifestation or prognosis comparing men to women in our study.
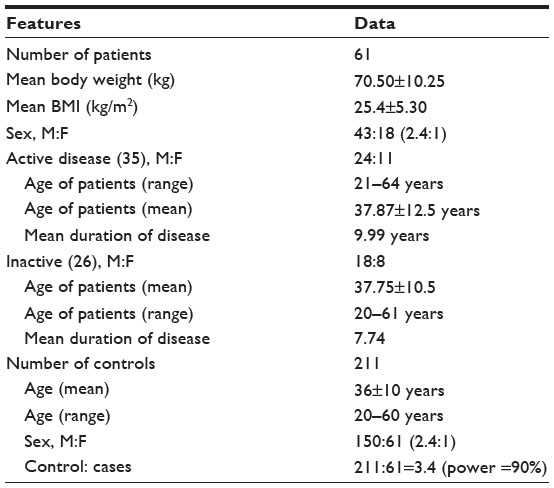 | Table 1 Demographic features of Saudi patients with Behçet’s disease and controls from same ethnic population |
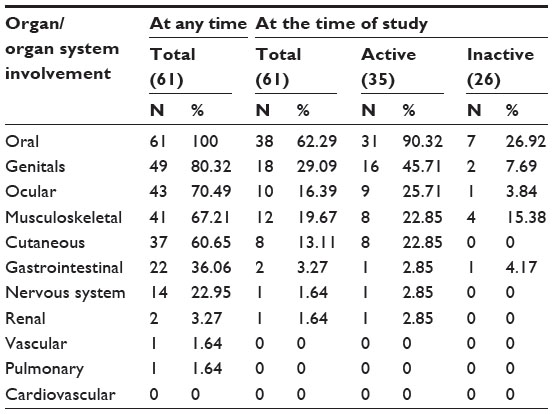 | Table 2 Clinical manifestations in patients with Bahcet’s disease |
The genotype and allele frequencies of TNF-α (−308G/A) and promoter polymorphism are presented in Table 3. For quality control, 25% of the random blind samples were repeated and genotyping results were compared with 100% success rate.
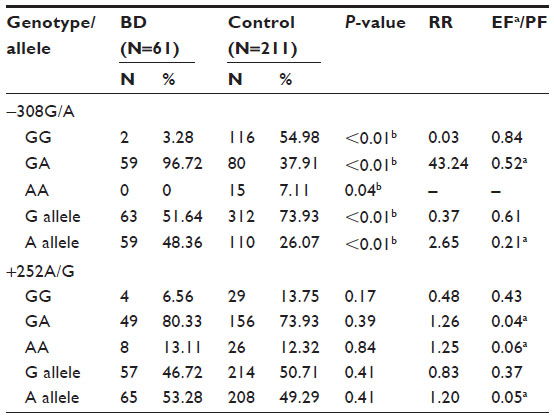 | Table 3 Genotype and allele frequencies of TNF-α and TNF-β variants in BD patients and matched controls |
The frequency of heterozygous genotype GA was significantly higher in BD patients than controls (P<0.01), whereas the frequency distribution of homozygous genotypes GG and AA was significantly lower in BD than controls (P<0.05). The frequency of allele A was significantly higher in BD patients than control subjects (P<0.01). On the other hand, allele G was significantly lower in BD patients as compared to controls (P<0.01).
The distribution of frequencies of alleles and genotype of TNF-β (+252A/G) promoter polymorphism did not differ significantly between BD patients and healthy controls (Table 3). The frequencies of allele A and genotypes GA and AA were slightly higher, whereas those of allele G and genotype GG were lower in BD patients than control subjects.
The frequency of −1082GG genotype was found to be significantly higher (P=0.04) in BD patients as compared to controls subjects. On the contrary, the frequency of heterozygous genotype GA was significantly lower (P=0.03) in patients as compared to control subjects. The frequency of homozygous AA genotype was higher in patients compared to controls, but the difference was not statistically significant (Table 4).
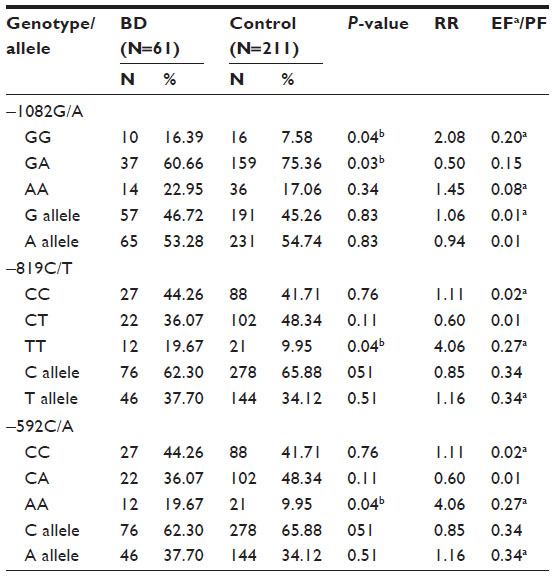 | Table 4 Genotype and allele frequencies of IL-10 variants in BD patients and matched controls |
The frequencies of −819CC and −819CT genotypes do not differ significantly between BD patients and controls, while the frequency of −819TT genotype was significantly higher in BD patients as compared to controls (P=0.04). Similarly, the frequency of homozygous −592AA genotype was significantly higher in BD patients as compared to controls (P=0.04), while CC and CA genotypes frequencies varied insignificantly in patient and control groups (Table 4).
Discussion
Clinical manifestations in our BD patients were almost similar to those reported earlier from Saudi BD patients by Al-Dalaan et al.48 Men were more affected than women. The male preponderance has also been reported in the Middle Eastern countries earlier by Kaklamani et al.49 Influence of sex on BD is well-known, and several reports have suggested a more severe course of disease among males.50 However, recently Davatchi et al51 found no strong association between the male sex and major organ involvement, except for vascular lesions. In contrast, females were more affected with BD than males in Japan and Korea.52 In our study, although no significant difference in clinical manifestation or prognosis between men and women was noticed, however, results should be interpreted with caution as the number of male and female patients is very low.
Significantly higher frequencies of allele A and genotype GA of TNF-α (308) polymorphism in patient group than controls (P<0.001) indicated that allele A and genotype GA are associated with the BD susceptibility risk in Saudi patients. The RR in GA carriers for the disease was very high (RR =43.236). On the other hand, the homozygous genotypes GG and AA might be protective for BD as their frequencies were significantly lower in patient groups than controls (P<0.001 and P=0.04, respectively). Our results are in agreement with a recent meta-analysis which showed higher frequency of allele A in Asian BD population.42
Our results clearly suggested that A allele and GA genotype confer susceptibility to BD and might be responsible for higher levels of TNF-α in BD patients as TNF-α (308) allele A has been associated with increased TNF-α expression.53 Moreover, patients with ocular manifestation have statistically significant higher serum levels of TNF-α than those without ocular manifestations.
Significantly higher TNF-α level has been reported in BD patients with active disease compared to those with inactive disease.14,54 Contrary to these, the G allele and GG genotype of TNF-α (308) polymorphism have been associated with increased risk of BD in Koreans,55 while no association of TNF-α (308) polymorphism with BD was reported in Turkish,22,56,57 Korean,58 Tunisian,23 Lebanes,59 Iranian,60 and Moroccan population.25
Though several individual studies from various geographical regions or ethnicities showed no association between TNF-α (308) polymorphism and BD, when pooled together in meta-analysis, a significant association of TNF-α polymorphism with BD was observed.42,43 Zhang et al43 indicted that the TNF-α (−308G/A) and TNF-α (−857T/C) polymorphisms are linked with BD only in Asians, while the TNF-α (−238A/G) and TNF-α (−1031C/T) polymorphisms are associated with BD in Caucasians. The other meta-analysis of 12 studies by Liang et al42 using the fixed-effect model also showed an association of the TNF-α (−308G/A) polymorphism with the susceptibility to BD. The discrepancy between the individual studies and meta-analysis has been explained by the facts that the sample size of individual studies is relatively small and that the allelic frequencies of genes often differ substantially in different ethnic groups as shown in various reports.42,43
In our study, the frequency of TNF-α (308) AA genotype was zero in Saudi BD patients. These results are in agreement with several other findings which also reported the absence of AA genotype in Lebanese, Iranian, and Moroccan BD patients.25,59,60 Absence of AA genotype in BD patients indicates that AA genotype might be protective for BD. Similarly, lower frequency of GG genotype in Saudi BD suggests its protective role in the Saudi cohort.
Our results of TNF-β (+252A/G) polymorphism showed no significant association with BD in Saudi patients. Other available reports also indicated that there is no association of TNF-β polymorphism with BD in Korean and Tunisian populations.23,58 In contrast, Verity et al61 reported a higher TNF-β*2 allele (A allele) frequency among BD cases compared to controls in Palestinian and Jordanian populations. They found strong linkage disequilibrium between HLA-B*51 and allele A of TNF-β (+252A/G) and suggested that both the alleles contribute to disease risk and that their coexpression leads to severe blinding in BD patients. Mizuki et al62 also reported significantly lower frequency of TNF-β1 (G allele) homozygote in Japanese ocular BD patients than controls. These variations in the association between TNF-β (+252) polymorphism and BD may reflect the heterogeneity in the genetic susceptibility to this disorder.
Our results indicated that genotype-1082GG of IL-10 (−1082G/A) is susceptible to BD, while genotype AA and GA are resistant to BD. Similar to our results, Talaat et al63 also reported increased frequency of genotype-1082GG and decreased frequency of genotype GA in Egyptian BD patients as compared to controls. However, Dilek et al64 found lower frequency of AA genotype in Turkish BD patients compared to the control group, but a higher frequency of −1082GA genotype. Contrary to this, Wallace et al20 reported from two populations (UK and Middle Eastern cohort) that the −1082AA genotype was weakly associated with BD when all patients were analyzed as a group, but not in the UK or Middle Eastern cohorts of patients alone compared to local controls.
Our results also showed that −819TT and 592 AA genotypes of IL-10 are significantly associated with susceptibility risk of BD in Saudi patients. IL-10 (−819T/A) polymorphism has been indicated to be significantly associated with BD.42 Wu et al65 reported that −819 T/A polymorphism is associated with BD in Chinese Han population. An association with −819T was also reported in UK and Middle Eastern BD patients, and this was because of an association in the UK but not Middle Eastern patients.20 Another meta-analysis with results from Turkish and Korean cohorts also showed genome-wide significant associations with −819T of IL10.19 A meta-analysis showed that IL-10 (–1082G/A, −819C/T, −592C/A) polymorphisms are significantly associated with BD,42 while Ates et al22 found no significant associations between IL10–1082G/A, −819C/T, −592C/A polymorphisms and Turkish BD.
Recently, emphasis has been given to address and clarify the functional relevance of the different genes found to be associated with BD susceptibility and the potential interactions between genes located within and outside the major histocompatibility complex region.66 Our results, suggesting an association of allele A and genotype GA of TNF-α (308) with BD in Saudi patients, indicate the functional relevance of the allele A as it is linked with higher levels of TNF-α reported in active BD patients as compared to controls.14,54 The proinflammatory cytokines, including TNF-α, are potent inducers of inflammation, and elevated levels of these cytokines are frequently associated with the activation of macrophages, thereby influencing the severity of inflammatory responses. These cytokines induce local inflammatory responses and also exert systemic effects. The overexpression of these cytokines may be responsible for the pathogenesis of recurrent BD.55 TNF-α has been suggested to be directly responsible for activation of T-cells and neutrophils, the main pathogenic changes in BD.67 Moreover, low producers (1082GG genotype) of IL-10, an anti-inflammatory cytokine, may not suppress the activity of TNF-α, resulting in increased inflammatory responses in BD patients. IL-10 limits the secretion of proinflammatory cytokines, such as TNF-α and IL-12.68 It is possible that the pathophysiology of a deficiency in the IL-10 involves undue and prolonged activation of mononuclear cells, resulting in an augmented efflux of inflammatory cytokines including TNF-α. IL-10 is a multifunctional cytokine with profound involvement in diverse areas of the human immune system.69
These results might have prognostic value for future clinical observations. Further, TNF-α (−308) polymorphism may serve as a guideline in determining the response to anti-TNF-α therapy, as patients with GG genotype are better responders to anti-TNF-α treatment than those with AA or GA.70,71 However, such associations need further validation and investigation in more patients with BD, as they may have implications for the development of novel therapies as suggested by Xavier et al.72 The main weakness of the study is the small sample size, while the strengths of the study lies in the fact that it is the first report from Saudi patients with BD, showing association of five polymorphisms in IL-10 and TNF-α and -β genes. The errors in genotyping both cases or controls have been avoided carefully by using a standard protocol with positive and negative controls. Statistical analysis was performed to get P-values, RR, etiologic fraction and preventive fraction, and power.
Conclusion
This study, though based on a small number of cases, clearly shows that TNF-α (−308) and IL-10 (−1082, −819, and −592) polymorphisms are significantly associated with BD susceptibility. This is the first report showing association of TNF-α and IL-10 polymorphisms with BD in Saudi Arabian patients.
Acknowledgments
The authors thank PSMMC for providing all facilities and S Sadaf Rizvi and Mohammad Al-Asmari for their help with laboratory work.
Disclosure
The authors report no conflicts of interest in this work.
References
Davatchi F, Shahram F, Chams-Davatchi C, et al. Behçet’s disease in Iran: analysis of 6500 cases. Int J Rheum Dis. 2010;13:367–373. | |
Kapsimali VD, Kanakis MA, Vaiopoulos GA, Kaklamanis PG. Etiopathogenesis of Behcet’s disease with emphasis on the role of immunological aberrations. Clin Rheumatol. 2010;29:1211–1216. | |
Yurdakul S, Yazici Y. Epidemiology of Behçet’s syndrome and regional differences in disease expression. In: Yazici Y, Yazici H, editors. Behçet’s Syndrome. 1st ed. New York, NY: Springer; 2010:35–52. | |
Cho SB, Cho S, Bang D. New insights in the clinical understanding of Behçet’s disease. Yonsei Med J. 2012;53(1):35–42. | |
Kaya TI. Genetics of Behcet’s disease. Patholog Res Int. 2012; 2012:912589. | |
Pineton de Chambrun M, Wechsler B, Geri G, Cacoub P, Saadoun D. New insights into the pathogenesis of Behcet’s disease. Autoimmun Rev. 2012;11:687–698. | |
Gul A. Behcet’s disease: an update on the pathogenesis. Clin Exp Rheumatol. 2001;19:6–12. | |
Takeno M, Shimoyama Y, Kashiwakura J, Nagafuchi H, Sakane T, Suzuki N. Abnormal killer inhibitory receptor expression on natural killer cells in patients with Behcet’s disease. Rheumatol Int. 2004;24: 212–216. | |
Raziuddin S, al-Dalaan A, Bahabri S, Siraj AK, al-Sedairy S. Divergent cytokine production profile in Behçet’s disease. Altered Th1/Th2 cell cytokine pattern. J Rheumatol. 1998;25:329–333. | |
Hamzaoui K, Hamzaoui A, Guemira F, Bessioud M, Hamza M, Ayed K. Cytokine profile in Behçet’s disease patients. Relationship with disease activity. Scand J Rheumatol. 2002;31:205–210. | |
Ben Ahmed M, Houman H, Miled M, Dellagi K, Louzir H. Involvement of chemokines and Th1 cytokines in the pathogenesis of mucocutaneous lesions of Behçet’s disease. Arthritis Rheum. 2004;50:2291–2295. | |
Akdeniz N, Esrefoglu M, Keles MS, Karakuzu A, Atasoy M. Serum interleukin-2, interleukin-6, tumor necrosis factor-α and nitric oxide levels in patients with Behcet’s disease. Ann Acad Med Singapore. 2004;33:596–599. | |
Oztas MO, Onder M, Gurer M, Bukan AN, Sancak B. Serum interleukin 18 and tumour necrosis factor-levels are increased in Behcet’s disease. Clin Exp Dermatol. 2005;30:61–63. | |
Akman A, Sallakci N, Coskun M, et al. TNF-α gene 1031 T/C polymorphism in Turkish patients with Behcet’s disease. Br J Dermatol. 2006;155:350–356. | |
Sugita S, Kawazoe Y, Imai A, et al. Role of IL-22- and TNF-α-producing Th22 cells in uveitis patients with Behcet’s disease. J Immunol. 2013;190:5799–5808. | |
Turner DM, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson IV. An investigation of polymorphism in the interleukin-10 gene promoter. Eur J Immunogenet. 1997;24:1–8. | |
Wilson AG, Symons JA, McDowell TL, McDevitt HO, Duff GW. Effects of a polymorphism in the human tumor necrosis factor α promoter on transcriptional activation. Proc Natl Acad Sci U S A. 1997;94: 3195–3199. | |
Remmers EF, Cosan F, Kirino Y, et al. Genome-wide association study identifies variants in the MHC class I, IL10, and IL23R-IL12RB2 regions associated with Behçet’s disease. Nat Genet. 2010;42:698–702. | |
Mizuki N, Meguro A, Ota M, et al. Genome-wide association studies identify IL23R-IL12RB2 and IL10 as Behçet’s disease susceptibility loci. Nat Genet. 2010;42:703–706. | |
Wallace GR, Kondeatis E, Vaughan RW, et al. IL-10 genotype analysis in patients with Behçet’s disease. Hum Immunol. 2007;68:122–127. | |
Khaib Dit Naib O, Aribi M, Idder A, et al. Association analysis of IL10, TNF-α, and IL23R-IL12RB2 SNPs with Behçet’s disease risk in Western Algeria. Front Immunol. 2013;4:342. | |
Ates O, Dalyan L, Hatemi G, Hamuryudan V, Topal-Sarikaya A. Analyses of functional IL10 and TNF-α genotypes in Behcet’s syndrome. Mol Biol Rep. 2010;37:3637–3641. | |
Kamoun M, Chelbi H, Houman MH, Lacheb J, Hamzaoui K. Tumor necrosis factor gene polymorphisms in Tunisian patients with Behcet’s disease. Hum Immunol. 2007;68:201–205. | |
Touma Z, Farra C, Hamdan A, et al. TNF polymorphisms in patients with Behcet’s disease: a meta-analysis. Arch Med Res. 2010;41:142–146. | |
Radouane A, Oudghiri M, Chakib A, Bennani S, Touitou I, Barat-Houari M. SNPs in the TNF-α gene promoter associated with Behcet’s disease in Moroccan patients. Rheumatology (Oxford). 2012;51:1595–1599. | |
Sharma S, Ghosh B, Sharma SK. Association of TNF polymorphisms with sarcoidosis, its prognosis and tumour necrosis factor(TNF)-α levels in Asian Indians. Clin Exp Immunol. 2008;151:251–259. | |
Sharma S, Sharma A, Kumar S, Sharma SK, Ghosh B. Association of TNF haplotypes with asthma, serum IgE levels, and correlation with serum TNF-α levels. Am J Respir Cell Mol Biol. 2006;35:488–495. | |
Abraham LJ, Kroeger KM. Impact of the –308 TNF promoter polymorphism on the transcriptional regulation of the TNF gene: relevance to disease. J Leukoc Biol. 1999;66:562–566. | |
Abdallah AN, Cucchi-Mouillot P, Biteau N, Cassaigne A, Haras D, Iron A. Analysis of the polymorphism of the tumour necrosis factor (TNF) gene and promoter and of circulating TNF-α levels in heart-transplant patients suffering or not suffering from severe rejection. Eur J Immunogenet. 1999;26:249–255. | |
Jeong P, Kim EJ, Kim EG, Byun SS, Kim CS, Kim WJ. Association of bladder tumors and GA genotype of –308 nucleotide in tumor necrosis factor-α promoter with greater tumor necrosis factor-α expression. Urology. 2004;64:1052–1056. | |
Kula, D, Jurecka-Tuleja B, Gubala E, Krawczyk A, Szpak S, Jarzab M. Association of polymorphism of LT α and TNF genes with Grave’s disease. Folia Histochem Cytobiol. 2001;39:77–78. | |
Lu LY, Cheng HH, Sung PK, Tai MH, Yeh JJ, Chen A. Tumor necrosis factor-β +252A polymorphism is associated with systemic lupus erythematosus in Taiwan. J Formos Med Assoc. 2005;104:563–570. | |
Panoulas VF, Nikas SN, Smith JP, et al. Lymphotoxin 252 A>G polymorphism is common associates with myocardial infarction in patients with rheumatoid arthritis. Ann Rheum Dis. 2008;67:1550–1556. | |
Boraska V, Zeggini E, Groves CJ, et al. Family-based analysis of tumor necrosis factor and lymphotoxin-α tag polymorphism with type 1 diabetes in the population of South Croatia. Hum Immunol. 2009;70:195–199. | |
Al-Rayes H, Al-Swailem R, Albelawi M, Arfin M, Asmari A, Tariq M. TNF-α and TNF-β gene polymorphism in Saudi rheumatoid arthritis patients. Clin Med Insights Arthritis Musculoskelet Disord. 2011;4: 55–63. | |
Yamashita N, Kaneoka H, Kaneko S, et al. Role of gamma delta T lymphocytes in the development of Behcet’s disease. Clin Exp Immunol. 1997;107:241–247. | |
Triolo G, Accardo-Palumbo A, Dieli F, et al. Vgamma9/Vdelta2 T lymphocytes in Italian patients with Behçet’s disease: evidence for expansion, and tumour necrosis factor receptor II and interleukin-12 receptor β1 expression in active disease. Arthritis Res Ther. 2003;5:R262–R268. | |
Messer G, Spengler U, Jung MC, et al. Polymorphic structure of the tumor necrosis factor (TNF) locus: an Ncol polymorphism in the first intron of the human TNF-β gene correlates with a variant amino acid in position 26 and a reduced level of TNF-β production. J Exp Med. 1991;173:209–219. | |
Abraham LJ, French MA, Dawkins RL. Polymorphic MHC ancestral haplotypes affect the activity of tumor necrosis factor α. Clin Exp Immunol. 1993;92:14–18. | |
Wallace G. Novel genetic analysis in Behçet’s disease. Arthritis Res Ther. 2009;11:123. | |
Song YW, Kang EH. Behçet’s disease and genes within the major histocompatibility complex region. Mod Rheumatol. 2012;22:178–185. | |
Liang Y, Xu WD, Zhang M, et al. Meta-analysis of association between cytokine gene polymorphisms and Behcet’s disease risk. Int J Rheum Dis. 2013;16:616–624. | |
Zhang M, Xu WD, Wen PF, et al. Polymorphisms in the tumor necrosis factor gene and susceptibility to Behcet’s disease: an updated meta-analysis. Mol Vis. 2013;19:1913–1924. | |
International study group for Behcet’s disease. Criteria for diagnosis of Behcet’s disease. Lancet. 1990;335:1078–1080. | |
Perrey C, Turney S, Pravica V, Howell WM, Hutchinson IV. ARMS-PCR methodologies to determine IL-10, TNF-α, TNF-β and TGF-β1 gene polymorphisms. Transpl Immunol. 1999;7:127–128. | |
Schallreuter KU, Levenig C, Kuhnl P, Loliger C, Hohl-Tehari M, Berger J. Histocompatability antigens in vitiligo: Hamburg study on 102 patients from Northern Germany. Dermatology. 1993;187:186–192. | |
Svejgaard A, Platz P, Ryder LP. HLA and disease 1982-A survey. Immunol Rev. 1983;70:193–218. | |
Al-Dalaan AN, Al Balaa SR, El Ramahi K, et al. Behçet’s disease in Saudi Arabia. J Rheumatol. 1994;21:658–661. | |
Kaklamani VG, Vaiopoulos G, Kaklamanis PG. Behçet’s disease. Semin Arthritis Rheum. 1998;27:197–217. | |
Yazici H, Tüzün Y, Pazarli H, et al. Influence of age of onset and patient’s sex on the prevalence and severity of manifestations of Behçet’s syndrome. Ann Rheum Dis. 1984;43:783–789. | |
Davatchi F, Shahram F, Chams-Davatchi C, et al. Behçet’s disease: is there a gender influence on clinical manifestations? Int J Rheum Dis. 2012;15:306–314. | |
Bang D, Lee JH, Lee ES, et al. Epidemiologic and clinical survey of Behcet’s disease in Korea: the first multicenter study. J Korean Med Sci. 2001;16:615–618. | |
Grove J, Daly AK, Bassendine MF, Day CP. Association of a tumor necrosis factor promoter polymorphism with susceptibility to alcoholic steatohepatitis. Hepatology. 1997;26:143–146. | |
El-Menyawi M, Fawzy M, Al-Nahas Z, et al. Serum tumor necrosis factor α (TNF-α) level in patients with Behçet’s disease: relation to clinical manifestations and disease activity. Egypt Rheumatologist. 2014;36:139–143. | |
Park K, Kim N, Nam J, Bang D, Lee ES. Association of TNFA promoter region haplotype in Behcet’s disease. J Korean Med Sci. 2006;21: 596–601. | |
Duymaz-Tozkir J, Gul A, Uyar FA, Ozbek U, Saruhan-Direskeneli G. Tumor necrosis factor-α gene promoter region –308 and –376 G>A polymorphisms in Behcet’s disease. Clin Exp Rheumatol. 2003;21: S15–S18. | |
Ates A, Kinikli G, Duzgun N, Duman M. Lack of association of tumor necrosis factor-α gene polymorphisms with disease susceptibility and severity in Behcet’s disease. Rheumatol Int. 2006;26:348–353. | |
Lee EB, Kim JY, Lee YJ, Park MH, Song YW. TNF and TNF receptor polymorphisms in Korean Behcet’s disease patients. Hum Immunol. 2003;64:614–620. | |
Arayssi TK, Hamdan AR, Touma Z, et al. TNF polymorphisms in Lebanese patients with Behçet’s disease. Clin Exp Rheumatol. 2008;26: S130–S131. | |
Amirzargar A, Shahram F, Nikoopour E, et al. Proinflammatory cytokine gene polymorphisms in Behcet’s disease. Eur Cytokine Netw. 2010;21:292–296. | |
Verity DH, Wallace GR, Vaughan RW, et al. HLA and tumour necrosis factor (TNF) polymorphisms in ocular Behcet’s disease. Tissue Antigens. 1999;54:264–272. | |
Mizuki N, Inoko H, Sugmura K, et al. RFLP analysis in the TNF-β gene and the susceptibility to alloreactive NK cells in Behcet’s disease. Invest Ophthalmol Vis Sci. 1992;33:3084–3090. | |
Talaat RM, Ashour ME, Bassyouni IH, Raouf AA. Polymorphisms of interleukin 6 and interleukin 10 in Egyptian people with Behcet’s disease. Immunobiology. 2014;219:573–582. | |
Dilek K, Ozçimen AA, Saricaoglu H, et al. Cytokine gene polymorphisms in Behçet’s disease and their association with clinical and laboratory findings. Clin Exp Rheumatol. 2009;27(2 Suppl 53):S73–S78. | |
Wu Z, Zheng W, Xu J, et al. IL10 polymorphisms associated with Behçet’s disease in Chinese Han. Hum Immunol. 2014;75:271–276. | |
Piga M, Mathieu A. Genetic susceptibility to Behcet’s disease: role of genes belonging to the MHC region. Rheumatology (Oxford). 2011;50: 299–310. | |
Balkwill F. TNF-α in promotion and progression of cancer. Cancer Metastasis Rev. 2006;25:409–416. | |
Fiorentino DF, Zlotnik A, Vieira P, et al. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J Immunol. 1991;146:3444–3451. | |
Glocker EO, Kotlarz D, Boztug K, et al. Inflammatory bowel disease and mutations affecting the interleukin-10 receptor. N Engl J Med. 2009;361:2033–2045. | |
Guis S, Balandraud N, Bouvenot J, et al. Influence of –308A/G polymorphism in the tumor necrosis factor α gene on etanercept treatment in rheumatoid arthritis. Arthritis Rheum. 2007;57:1426–1430. | |
Seitz M, Wirthmuller U, Moller B, Villiger PsM. The –308 tumor necrosis factor-α gene polymorphism predicts therapeutic response to TNFα-blockers in rheumatoid arthritis and spondyloarthritis patients. Rheumatology (Oxford). 2007;46:93–96. | |
Xavier JM, Shahram F, Davatchi F, et al. Association study of IL10 and IL23R-IL12RB2 in Iranian patients with Behçet’s disease. Arthritis Rheum. 2012;64:2761–2772. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.