Back to Journals » Psoriasis: Targets and Therapy » Volume 16
Inflammaging and Senescence-Associated Secretory Phenotype (SASP) in Psoriasis – A Narrative Review of Potential Mechanisms and Anti-Inflammaging Strategies
Authors Filipek K, Nowowiejska-Purpurowicz J, Flisiak I ![]()
Received 22 January 2026
Accepted for publication 30 April 2026
Published 28 May 2026 Volume 2026:16 598115
DOI https://doi.org/10.2147/PTT.S598115
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Mio Nakamura
Kinga Filipek, Julia Nowowiejska-Purpurowicz, Iwona Flisiak
Department of Dermatology and Venereology, Medical University of Bialystok, Bialystok, Poland
Correspondence: Kinga Filipek, Department of Dermatology and Venereology, Medical University of Bialystok, Bialystok, Poland, Email [email protected]
Abstract: Psoriasis is a common chronic dermatological disease, affecting approximately 1– 3% of the global population, and is associated with numerous comorbidities, impaired quality of life, and reduced life expectancy compared with the general population. The pathogenesis of psoriasis is complex and multifactorial, involving genetic susceptibility, external environmental factors, and immune system dysregulation. Psoriasis is perceived as a systemic disorder associated with a systemic inflammatory condition. In psoriasis, a chronically activated immune response results in the expression of numerous pro-inflammatory mediators, which enhance and sustain the inflammatory feedback loop, thereby exacerbating cell senescence. Senescent cells adopt a hypersecretory state called senescence-associated secretory phenotype (SASP). Multiple SASP-related mediators are dysregulated in psoriasis, including pro-inflammatory cytokines, chemokines, growth factors, proteases and regulators, soluble or shed receptors and ligands, some of which may serve as biomarkers or therapeutic targets. Therefore, psoriasis is closely associated with immune dysregulation and inflammaging, which refers to a chronic, low-grade inflammatory state with exacerbated cellular senescence. The relationship between psoriasis, inflammation, and cellular senescence is multidirectional, and might be considered in two hypothetical scenarios of cause-and-effect relationship. A persistent pro-inflammatory state might promote cellular senescence and SASP upregulation, which in turn may trigger immunological alterations characteristic of psoriasis, or psoriasis and associated dysregulation of the immune system leading to systemic inflammation and thereby senescence. Hence, it is essential to clarify the mechanisms of inflammaging and the role of SASP in psoriasis. It may support the development of the therapeutic strategies that take into consideration comorbidities and their shared inflammatory background with psoriasis, enabling more precise assessment of the disease. The image A showing inflammaging in psoriasis. Keratinocytes and immune cells are involved in chronic inflammation, mediated by pro-inflammatory mediators and SASP. The image B showing senescence-associated secretory phenotype (SASP). Senescent cells release cytokines, chemokines, growth factors, proteases, regulators and soluble or shed receptors or ligands. These contribute to autocrine/paracrine senescence, inflammation, tissue remodeling, angiogenesis and tumor promotion/suppression. The image C showing potential use in precision dermatology. Individual patient characteristics, external factors like environment and lifestyle and comorbidities are considered. SASP profiling aids in administering patient-tailored treatment based on data integration.Three panels on psoriasis: inflammaging, SASP and precision dermatology use.
Keywords: psoriasis, senescence-associated secretory phenotype, inflammaging
Introduction
Psoriasis is one of the most common chronic dermatological diseases, affecting approximately 1–3% of the global population,1,2 with particularly high prevalence in Western European countries.3,4 This implies that millions of people around the world struggle with the disease daily. Psoriasis is associated with several comorbidities, including cardiovascular and metabolic diseases, and it also affects patients’ mental health.5 Psoriasis is a chronic, lifelong inflammatory disease, and currently, there is no therapeutic strategy to definitively cure psoriasis or one that would act effectively for every patient.
The pathogenesis of psoriasis is complex, multifactorial, and still not fully elucidated. The development of the disease involves dysregulation of the immune response, abnormal keratinocyte differentiation and proliferation, and impaired angiogenesis.6,7 Genetic susceptibility8–10 and numerous environmental triggers, such as infections, stress, medications, and mechanical trauma, also contribute to disease onset and exacerbation.11,12
The function of numerous components of the immune system is significantly disrupted in psoriasis, which is reflected in abnormal levels of several regulatory molecules. Significantly increased concentrations of interleukins, chemokines, growth factors and other regulatory mediators are observed. Their interplay creates a complex signaling network that sustains chronic inflammation.
In recent years, increasing attention has been given to the role of cellular senescence and its contribution to psoriasis. Cellular senescence is a state of irreversible cell cycle arrest induced by stressors such as oxidative stress and DNA damage.13 Senescent cells remain metabolically active and adopt the senescence-associated secretory phenotype (SASP).14 Established markers of senescence include increased expression of cyclin-dependent kinase inhibitors, such as p16^INK4a and p21, as well as senescence-associated β-galactosidase (SA-β-gal) activity.15 Available evidence suggests that oxidative stress and DNA damage contribute to senescence-associated pathways in psoriasis.16 Increased reactive oxygen species (ROS) in psoriatic skin can lead to DNA damage and activation of pathways such as p53/p21 and p16^INK4a/Rb, promoting senescence.15 These senescent cells may further amplify inflammation via SASP. Cellular senescence is associated with markers of biological aging. Telomere shortening reflects cumulative cellular stress and can trigger DNA damage responses.13 Additionally, epigenetic age acceleration, assessed using DNA methylation-based clocks, provides a measure of biological aging independent of chronological age.17 These findings support the concept of inflammaging, which describes the interplay between chronic, low-grade inflammation and aging-related processes.
Epigenetic mechanisms, including DNA methylation, histone modifications, and non-coding RNAs, are known to regulate gene expression in psoriasis.18 However, their specific role in controlling SASP-related pathways in the context of psoriasis has not yet been clearly established and remains an emerging area of research.19
Keratinocytes, which play a crucial role in psoriasis pathology, are characterized by hyperproliferation and parakeratosis. Keratinocytes in psoriasis may show a senescent-like phenotype.20 Hyperproliferation may coexist with features of cellular senescence.20 Upper epidermal keratinocytes have been shown to exhibit a senescence-like state despite remaining metabolically active.21 Chronic inflammatory and stress-related signals, including oxidative stress and DNA damage, may induce cell-cycle arrest pathways.20 Pro-survival signalling mechanisms, such as activation of the PI3K/AKT pathway, may allow these senescent-like keratinocytes to persist within the psoriatic epidermis rather than being eliminated.21
Dermal fibroblasts are increasingly recognized as contributors to psoriasis pathogenesis by influencing keratinocytes and modulating local immune responses through the secretion of cytokines, growth factors, and extracellular matrix components.22,23 Although some features of activated fibroblasts overlap with characteristics associated with cellular senescence, direct evidence for a senescent phenotype in fibroblasts within psoriatic lesions remains limited.23
Endothelial cells also play an important role in psoriasis by contributing to vascular remodeling, angiogenesis, and leukocyte recruitment within lesional skin.24 Activated endothelial cells upregulate adhesion molecules, facilitating the infiltration of immune cells into the skin.24 Although chronic inflammatory and oxidative stress conditions present in psoriasis are known to promote endothelial senescence, direct evidence for a senescent phenotype in endothelial cells within psoriatic lesions remains limited.24
Among immune cells involved in psoriasis pathogenesis, senescence has been most clearly demonstrated in lesional CD4⁺ T cells, which exhibit increased expression of p16^INK4a^ and p21.25 Earlier studies have also suggested that chronic T-cell activation may contribute to disease persistence and cytokine dysregulation.26
The microscopic alterations translate into the clinical picture of psoriasis, which manifests with scaly plaques on the skin, erythematous-scaly lesions on the scalp, and nail lesions.27 Psoriasis significantly affects patients’ quality of life and may lead to work disability,28,29 deterioration of interpersonal relationships, and exclusion from many daily activities.30 Therefore, it is crucial to continue exploring the pathogenesis of the disease and to seek increasingly advanced, patient-tailored therapeutic strategies aimed at restoring normal functioning for millions of patients worldwide.
Senescence-Associated Secretory Phenotype (SASP)
Senescing cells undergo irreversible morphological and functional changes. As a result of factors such as DNA damage, telomere shortening, oncogene activation, or oxidative injury, the cell cycle becomes arrested,14,31,32 and the cells enter a state of hypersecretion characterized by persistent release of pro-inflammatory cytokines (eg, interleukin-6 (IL-6), interleukin-1 beta (IL-1β), interleukin-8 (IL-8)),14 chemokines (eg, C-C motif chemokine ligand 2 (CCL2), C-X-C motif chemokine ligand 1 (CXCL1), C-X-C motif chemokine ligand 5 (CXCL5), C-X-C motif chemokine ligand 8 (CXCL8)),14 growth factors (eg, vascular endothelial growth factor (VEGF), transforming growth factor beta (TGF-β), hepatocyte growth factor (HGF)), as well as extracellular matrix–degrading enzymes such as matrix metalloproteinases (eg, matrix metalloproteinase-1 (MMP-1), matrix metalloproteinase-3 (MMP-3), matrix metalloproteinase-10 (MMP-10))14 and serine proteases - collectively referred to as the senescence-associated secretory phenotype (SASP)33,34 (Figure 1). SASP is heterogeneous,35 depending on the cell type, the stimuli affecting the cell, its microenvironment, and the duration of exposure,36,37 often determining whether SASP has a beneficial or harmful impact.38 Initially described as a protective mechanism promoting tissue repair and tumor suppression, persistent SASP signaling can become damaging, leading to chronic inflammation and tissue remodeling.39 SASP factors secreted by senescing cells might have multimodal effects on their environment and other cells, promoting senescence, reinforcing senescence, and altering immunological processes.33,40,41 This indicates the complexity of the impact of SASP on biological processes leading to “inflammaging” and dysregulation of homeostasis, and may also contribute to the pathophysiology of skin diseases.20
 |
Figure 1 Cell in response to the aggravating factors develop a senescence-associated secretory phenotype (SASP), secreting numerous mediators e.g. cytokines, chemokines, growth factors, proteases and regulators, soluble or shed receptors or ligands. SASP components may affect numerous processes, including promoting senescence, sustaining inflammation, participating in angiogenesis and tissue remodeling, and promoting or suppressing tumor development. Created in BioRender. |
Inflammaging in Psoriasis
The concept of inflammaging refers to a chronic, low-grade inflammatory state that increases with age or pathological triggers like ultraviolet radiation (UVR), pollution, chronic inflammatory disorders, and in which senescent cells and their SASP play a central role.42 Senescent cells, which adopt a particular SASP profile, accelerate or sustain chronic inflammation, leading to progressive tissue dysfunction and the amplification of inflammatory pathways characteristic of psoriasis39 (Figure 2). Psoriasis is considered a disease linked to inflammaging. In psoriasis, the same molecular pathways - including interleukin-17 (IL-17), interleukin-23 (IL-23) and tumour necrosis factor alpha (TNF-α), oxidative stress, and endothelial dysfunction - are constitutively activated, resulting in cellular senescence features and an inflammaging-like profile.12 The chronic inflammatory condition in psoriasis is not only limited to the local skin inflammation, but rather systemic, which may account for the higher risk of comorbidities such as cardiovascular disease, obesity, metabolic syndrome, and others, in which the inflammatory background is also being explored.43 Patients with moderate to severe psoriasis have approximately a 5-year reduction in life expectancy compared with the general population,44,45 because of chronic systemic inflammation that increases the risk of comorbidities.43,46 Numerous SASP mediators in psoriasis demonstrate dysregulated expression, contributing to the development and persistence of inflammaging. The relationship between psoriasis, inflammation, and cellular senescence is multidirectional,47 and might be considered in two hypothetical scenarios of cause-and-effect relationship. A persistent pro-inflammatory state might promote cellular senescence and SASP upregulation, which in turn may trigger immune dysregulation characteristic of psoriasis, or psoriasis and associated dysregulation of the immune system leading to systemic inflammation and thereby senescence. The pathogenesis of psoriasis may be associated with impaired regulation of cellular senescence,48 which emphasizes the importance of clarifying the mechanisms of inflammaging and the role of SASP in psoriasis.
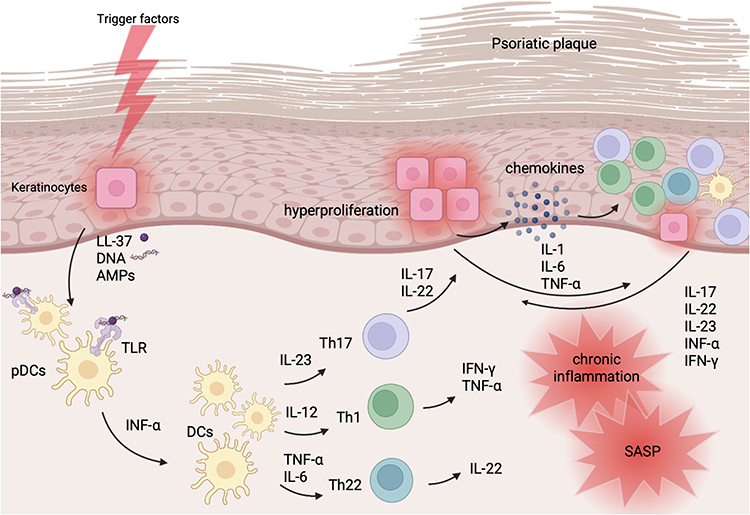 |
Figure 2 Schematic immunology of psoriasis. Trigger factor (e.g. injury, infection, drugs, toxic substances, genetics and others) cause damage-associated molecular patterns (DAMPs) release. Activated keratinocytes secrete antimicrobial peptides (AMPs), LL-37 and DNA of damaged cells, that form complexes and through the toll-like receptors (TLR) activate plasmacytoid dendritic cells (pDCs). Interferon alpha (IFN-α) participates in dendritic cells (DCs) maturation. DCs secrete interleukin-23 (IL-23), interleukin-12 (IL-12), interleukin-6 (IL-6), tumour necrosis factor alpha (TNF-α), which participate in the activation of lymphocytes. IL-23 induces Th17 lymphocytes to produce IL-17, IL-22. IL-17 affects keratinocytes, causing hyperproliferation, secretion of pro-inflammatory cytokines, secretion of chemokines that attract immune cells, therefore sustaining the inflammatory feedback loop. IL-22 impairs keratinocytes differentiation. IL-12 induces Th1 lymphocytes to produce TNF-α and IFN- γ, which in turn activates dendritic cells, keratinocytes, exaerbates inflammation. TNF-α and IL-6 induces Th22 lymphocytes to produce IL-22. This results in chronic inflammation and the transition of cells toward a senescence-associated secretory phenotype (SASP). Created in BioRender. |
Senescence-Associated Secretory Phenotype Factors and Their Involvement in Psoriasis
Cytokines
The secretory profile in psoriasis includes many cytokines released by fibroblasts, keratinocytes, immune cells such as dendritic cells and lymphocytes. Among the cytokines, whose role has been investigated extensively in the pathogenesis of psoriasis, there are IL-17, IL-23, IL-12, IL-22, TNF-α, and interferon gamma (IFN-γ).49 Some cytokines involved in the pathogenesis of psoriasis are also SASP components. Table 1 summarizes major SASP-cytokines and their contribution to the pathogenesis of psoriasis.
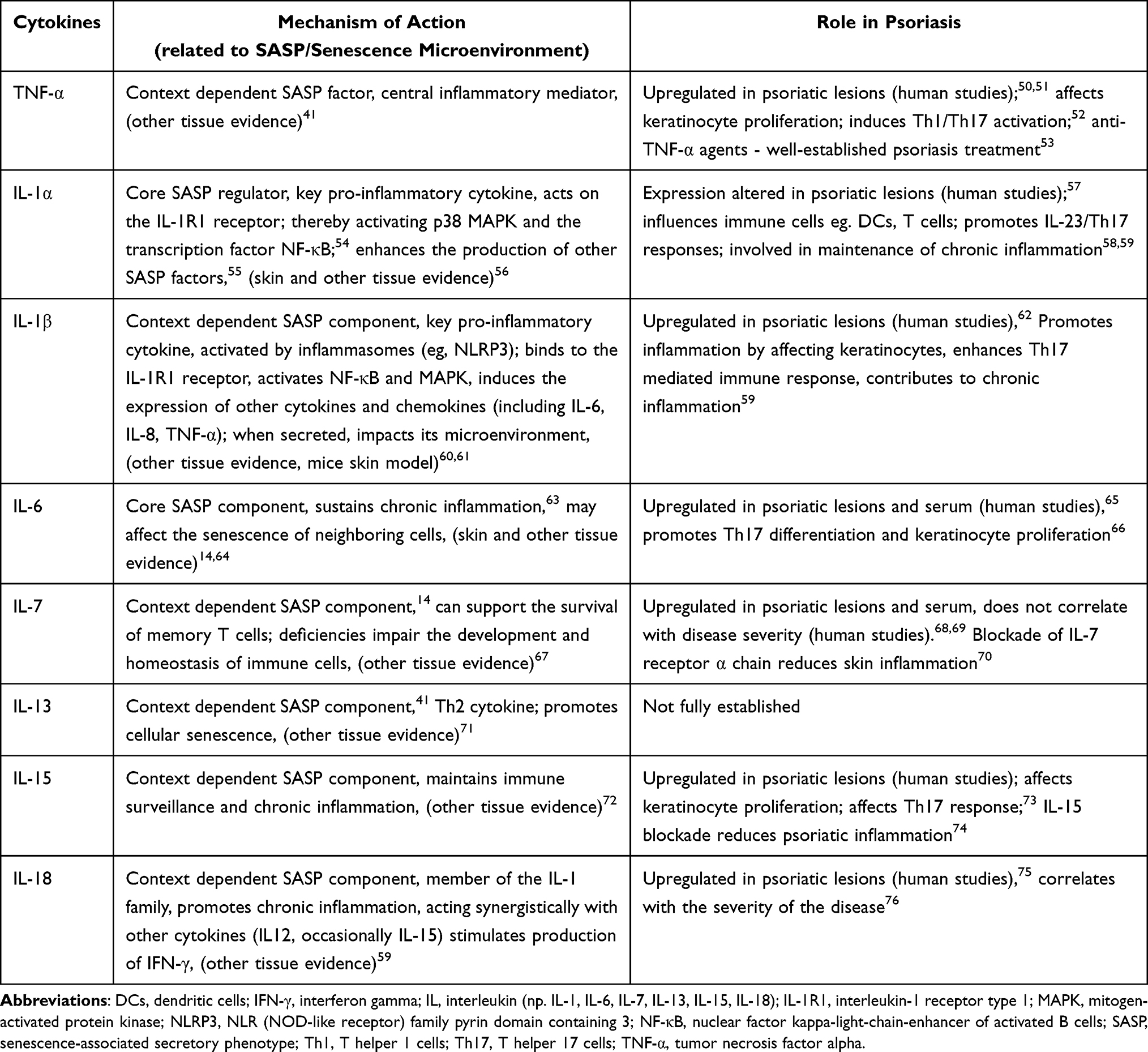 |
Table 1 The Table Summarizes Major SASP-Cytokines and Their Contribution to the Pathogenesis of Psoriasis |
One of the most important immune pathways in psoriasis is the interleukin-23/T helper 17 (IL-23/Th17) axis.77 IL-23, produced mainly by antigen-presenting cells (APCs), influences the expansion and stabilization of Th17 cells,77 which in turn secrete IL-17 and IL-22. IL-17 influences the proliferation and differentiation of keratinocytes and stimulates the production of pro-inflammatory cytokines (eg, IL-1, IL-6, TNF),78 chemokines (CXCL-1-3, CXCL-5, CXCL-8/IL-8, CCL-20),49 which in turn drives the pro-inflammatory cascade.79 IL-22, secreted mainly by Th17 and Th22, stimulates keratinocyte proliferation and influences their differentiation.80,81 In the context of psoriasis, pro-inflammatory cytokines such as IL-1α, IL-1β, IL-6, IL-7, IL-13, IL-15, IL-18, and TNF-α are components of the SASP, and their altered expression contributes to the maintenance and amplification of chronic inflammation.14
TNF-α is one of the main pro-inflammatory factors in psoriasis; it promotes dendritic cell maturation and IL-23 production, enhances IL-23/ Th17 response and NF-κB-dependent activation/proliferation of keratinocytes, and increases endothelial activation and leukocyte recruitment within plaques.52 TNF-α inhibitors are well-established and evidence-based therapeutic option in the management of moderate-to-severe psoriasis.53
IL-1α and IL-1β are pro-inflammatory cytokines involved in the pathogenesis of psoriasis. IL-1β is mainly circulating cytokine that contributes to the systemic inflammatory response. IL-1α is associated with local inflammatory processes due to its mainly cell membrane-bound form.82 The function of IL-1α and IL-1β is similar, targeting keratinocytes, fibroblasts, endothelial cells and lymphocytes. They contribute to increased production of cytokines, chemokines, and adhesion molecules, leading to enhanced recruitment and accumulation of inflammatory cells in the skin.61
IL-18 is a part of the IL-1 cytokine family. IL-18 acts mainly on NK cells and T lymphocytes. Together with IL-12 (secreted by dendritic cells and macrophages), it leads to increased production of IFN-γ by Th1 cells.61
IL-6 is a pro-inflammatory cytokine that has a significant function in acute inflammatory processes as well as in sustaining chronic inflammation.63 Therapies targeting IL-6, such as tocilizumab (an antibody against the IL-6 receptor) and siltuximab (an IL-6 antagonist), have been used in IL-6 associated diseases (eg. rheumatoid arthritis, cytokine release syndrome).83 Further research may help determine the full therapeutic potential, including psoriasis.
IL-7 is classified as part of the SASP profile in selected models depending on the context. It modulates the immune response by influencing the development and maintenance of numerous immune cells, including T cells.67
Typically described as a Th2 cytokine, IL-13 has complex mechanisms of action and its role in psoriasis is not fully established. It has been shown that IL-13 in psoriasis acts synergistically with IFN-γ to stimulate the production of chemokines CCL2 and CCL562. In patients with psoriasis, serum IL-13 levels were lower than in the control group and did not correlate with pruritus or disease severity.84 IL-13 may act as a context-dependent SASP factor, promoting cell aging, which leads to increased expression of SASP markers, and may itself belong to the cell SASP profile.14,85
IL-15 might play a role in pathogenesis of psoriasis. Experimental mouse models have shown that the blockade of IL-15 reduced the severity of psoriasis.74 IL-15, acting synergistically with IL-23, affects the Th17-dependent response.73
Chemokines
Chemokines, cytokines with chemotactic properties, are a large family of molecules responsible for directing the migration (chemotaxis) of cells.86 In psoriasis, keratinocytes, endothelial cells produce chemokines that regulate the influx of neutrophils, Th1/Th17 T cells, and other immune cells into lesional skin, thereby sustaining chronic inflammation.12,87,88 Cytokines, which play a key role in the pathogenesis of psoriasis, regulate the expression of chemokines and their receptors. Table 2 summarizes chemokines, their receptors, and their contribution to psoriasis pathogenesis, and their role as SASP. IL-17 upregulates expression of CXCL1, CXCL2, CXCL8, CCL2, CCL7 and CCL20, while IFN-γ upregulates expression of CXCL9, CXCL10, CXCL1112, thereby enhancing and sustaining the inflammatory feedback loop.88 In patients with psoriasis, the levels of several chemokines (including CCL2, CCL5, CCL20, CXCL8, CXCL9, CXCL10 and CXCL11) are elevated in lesional skin or serum.12 For some of these chemokines, serum or tissue concentrations show a positive correlation with disease severity. For example, CCL20 is being investigated as a potential biomarker in psoriasis,89 CXCL10 as a predictive biomarker for the development of psoriatic arthritis in patients with psoriasis.90 The expression of receptors for specific chemokines is crucial for the development of the immune response and the regulation of chemotaxis. In psoriasis, increased expression of these receptors on T cells is observed, which may intensify chemotaxis even at lower concentrations of chemokines.12 SASP of senescent keratinocytes and fibroblasts of the skin includes chemokines (eg, CCL2, CXCL8), some of these chemokines overlap with the chemokine secretion profile observed in psoriasis.14
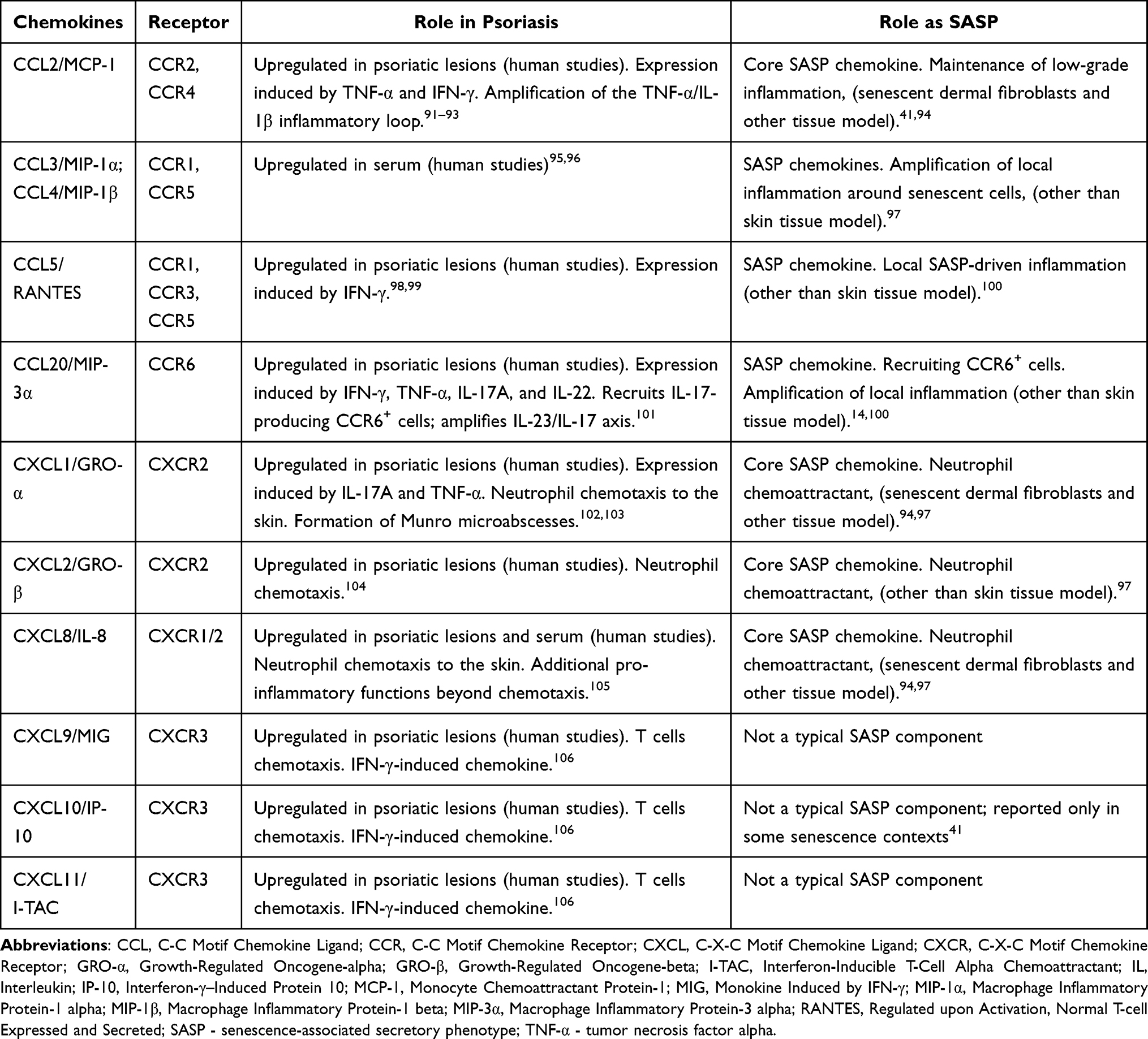 |
Table 2 The Table Summarizes Chemokines, Their Receptors, and Their Contribution to Psoriasis Pathogenesis, and Their Role as SASP |
Growth Factors
Growth factors (GFs) are a crucial component of the complex signaling system that regulates proliferation, differentiation, and cellular interactions within the skin. In psoriasis, their overexpression leads to a dysfunctional homeostasis contributing to chronic inflammation and abnormal tissue regeneration.107,108 Table 3 summarizes the role of growth factors in psoriasis.
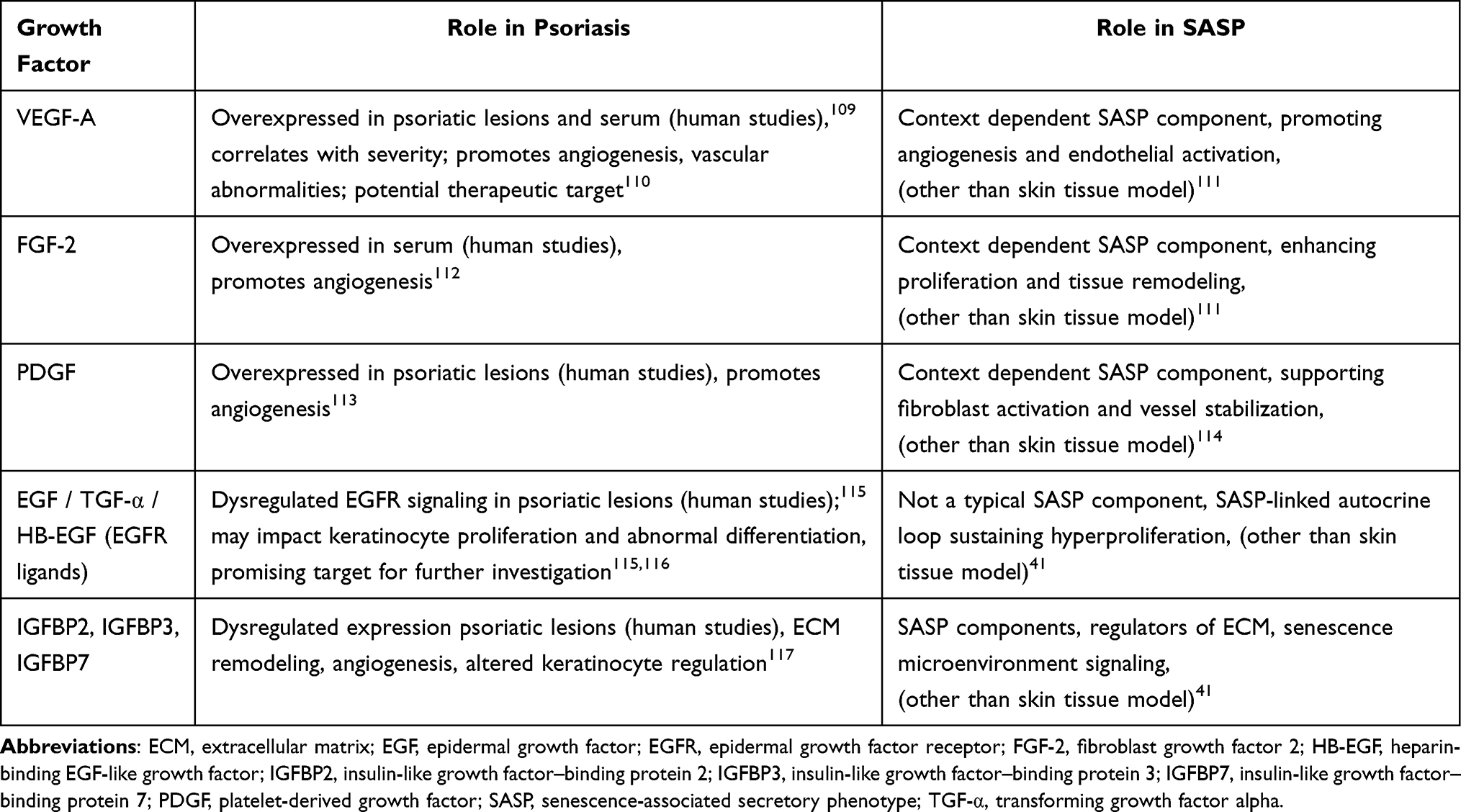 |
Table 3 The Table Summarizes Key Growth Factors Implicated in Psoriasis and Their Relevance to SASP-Related Processes Within the Inflammatory and Pro-Angiogenic Microenvironment |
Significantly increased VEGF expression was observed within plaques of psoriasis and in the serum of patients with psoriasis.109,118 A positive correlation between VEGF concentration in serum and psoriatic plaques and disease activity was also confirmed.119 VEGF represents a superfamily of molecules, among which VEGF-A is the main member, and the term VEGF usually refers to VEGF-A.120 VEGF molecules act through the receptors: vascular endothelial growth factor receptor 1 and 2 (VEGFR-1, 2), which are mainly present on vascular endothelial cells.121 Their activation leads to inflammatory angiogenesis. In psoriasis, this process causes vascular abnormalities such as increased endothelial cell proliferation, formation of new blood vessels, vessel enlargement, and increased endothelial permeability,122 which clinically contributes to the occurrence of the Auspitz sign.123 A significant decrease in serum VEGF concentration has been observed in patients treated with TNF-α inhibitors.124–126 This suggests that these drugs may indirectly reduce angiogenesis and indicates the need for further studies focusing on angiogenesis as a potential therapeutic target.110 There have also been single reports of patients in whom anti-VEGF therapy, initially used for oncological or ophthalmological conditions, resulted in a significant reduction in the severity of psoriatic lesions.127–130 Due to the role of VEGF in the pathogenesis of psoriasis, a study was conducted on transgenic mice to assess how overexpression of the VEGF gene in the skin affects vascular and immune processes.131 The study showed that increased local production of VEGF leads to abnormal angiogenesis and chronic skin inflammation resembling human psoriasis.131 VEGF may serve as a biomarker of psoriasis activity and as a potential therapeutic target; however, the efficacy and safety of direct inhibition of this pathway require further confirmation in clinical studies.110
Like VEGF, fibroblast growth factor 2 (FGF-2) and platelet-derived growth factor (PDGF) participate in the process of pathological angiogenesis in psoriasis, creating vessels with an unstable structure that promote chronic inflammation.132–134 Elevated concentration of FGF-2 has been observed in patients with psoriasis.112 Significantly increased expression of PDGF and its receptor has been demonstrated in skin lesions in psoriasis.113 However, their potential role in potential diagnostic/therapeutic solutions requires further investigation.
Another group of important factors is those from the epidermal growth factor (EGF) family, including epidermal growth factor (EGF), transforming growth factor alpha (TGF-α), and heparin-binding epidermal growth factor (HB-EGF),135 which activate the epidermal growth factor receptor/mitogen-activated protein kinase (EGFR/MAPK) pathway, intensifying keratinocyte proliferation and thickening of the epidermis.115 Overexpression of EGF family ligands has been observed in psoriatic skin and has been linked to the pathogenesis of the disease.116,136 The relationship between EGF concentration and clinical disease severity was observed and suggests that EGF might be investigated as a potential marker for assessing disease severity.136 EGFR overexpression in keratinocytes promotes inflammatory changes in psoriasis.115,137 Furthermore, individual clinical observations of cancer patients treated with EGFR inhibitors (eg, cetuximab, lapatinib, erlotinib, panitumumab) for cancer who also had psoriasis showed a reduction in psoriatic lesions.115
Insulin-like growth factor-binding proteins (IGFBPs) regulate insulin-like growth factor 1 and 2 (IGF-1, 2) by modulating their interaction with their receptor, and also influence the proliferation, differentiation, and inflammatory response of keratinocytes and fibroblasts.138 Among IGFBPs 1–7, the significance of IGFBP2, IGFBP3, and IGFBP7 in psoriasis has been investigated. It has been observed that IGFBP2 is aberrantly expressed in keratinocytes taken from skin lesions of patients with psoriasis, where this protein contributed to the reduction of keratinocyte apoptosis and the persistence of chronic inflammation.117 IGFBP3 shows increased expression in the epidermis from psoriasis lesions, but its function remains unclear.139,140 In psoriasis, IGFBP7 plays a complex role depending on the cell type – in keratinocytes, its reduced expression promotes excessive proliferation and epidermal differentiation dysfunction,141 while in endothelial cells, increased IGFBP7 expression escalates the inflammatory process.142 These data indicate that IGFBP7 participates in both the regulation of epidermal proliferation and the regulation of inflammation, making it a potential therapeutic target in psoriasis.142,143
Growth factors do not act independently, but form a complex communication system of interdependencies with inflammatory cytokines.7 IL-17A and TNF-α induce the secretion of VEGF-A, FGF-2, and HB-EGF in keratinocytes,144,145 and these in turn cause an increased lymphocyte influx by increasing the expression of adhesion molecules (ICAM-1, VCAM-1).24 This results in a self-perpetuating inflammatory cascade.7
Proteases and Regulators
CXCL13/B-Lymphocyte Chemoattractant
CXCL13/B-lymphocyte chemoattractant (BLC) is a chemokine that binds to the receptor CXCR5 and regulates the chemotaxis of B cells and T follicular helper cells, affecting the proper functioning and organization of secondary lymphatic organs.146,147 Disturbances of the CXCL13/CXCR5 axis are observed in several autoimmune and inflammatory diseases.148 In psoriasis, upregulation of CXCL13 is observed in lesional skin and peripheral blood and correlates with the disease severity. It has also been noted that CXCL13 levels in psoriatic lesions decrease after anti-IL-23 treatment, suggesting its potential role as a biomarker.148,149 Considered a SASP factor in certain tissue models, it contributes to immunosenescence by modulating the inflammatory microenvironment.150,151
Cathepsin B
Cathepsin B, a member of the cysteine cathepsin family, is a proteolytic enzyme found physiologically mainly in lysosomes that is involved in extracellular matrix (ECM) remodeling. Cathepsin B has been identified as a contributor to cancer progression, and its expression is elevated in several tumor types.152 Senescent cells show dysregulated expression and activity of cathepsin B, suggesting that this protease may influence senescence-related remodeling of the tissue microenvironment.41,153 In psoriatic skin, cathepsin B-positive senescent keratinocytes and mast cells may therefore contribute to the pathogenesis of psoriasis, and cathepsin B has been identified as a potential biomarker or therapeutic target.154
Cyclo-Oxygenase-1 and −2
Cyclo-oxygenase-1 and −2 (COX-1 and COX-2) are key enzymes in prostaglandin synthesis and therefore play a crucial role in regulating inflammatory processes.155,156 Both COX-1 and COX-2 show dysregulated expression in psoriatic skin, which leads to altered prostaglandin synthesis and contributes to the maintenance of chronic inflammation characteristic of psoriasis.157,158
CD26/Dipeptidyl Peptidase 4
CD26/dipeptidyl peptidase 4 (DPP4/DP4/DPPIV) molecule is involved in several complex processes. It performs an enzymatic function by degrading numerous peptides, including GLP-1 and GIP (incretins regulating insulin secretion), which is why DPP4 inhibitors are used in the treatment of type 2 diabetes. In addition, DPP4 is involved in immunological processes, participates in the activation of T lymphocytes, contributes to the degradation of chemokines, and modulates the inflammatory response.159 An 11-fold increase in the expression of the CD26/DPP4 mRNA in psoriatic epidermis has been demonstrated.160 Another study demonstrated elevated serum expression of DPP4 in individuals with psoriasis and proposed its potential role in the induction of pruritus.161
Granulocyte Colony-Stimulating Factor
Granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), macrophage colony-stimulating factor (M-CSF) play a crucial role in hematopoiesis, regulating the differentiation and development of neutrophils, granulocytes, and monocytes/macrophages.162 In contrast, aging cells secrete them along with other pro-inflammatory molecules, amplifying chronic inflammation in the tissue microenvironment.41 Increased expression of the GM-CSF was found in the skin and serum of patients with psoriasis.163 Clinical case reports document that administration of recombinant GM-CSF or G-CSF for neutropenia may induce or exacerbate psoriatic-like lesions.164–166 However, a randomized, controlled study demonstrated that namilumab, a specific human anti-GM-CSF monoclonal antibody, has not been effective enough to treat patients with psoriasis.167
Matrix Metalloproteinases
Matrix metalloproteinases (MMPs) are responsible for the degradation and remodeling of extracellular matrix components, regulating processes such as healing and cell migration. MMPs activity is regulated by their inhibitors eg. tissue inhibitor of metalloproteinases-1 (TIMP-1), tissue inhibitor of metalloproteinases-2 (TIMP-2).168 Several molecules from the MMPs family, which also belong to the classic SASP of skin cells, are involved in the regulation of inflammatory processes and in the pathogenesis of psoriasis.14 In psoriasis, MMP-1, MMP-2, MMP-3, MMP-9 and MMP-13 are most commonly reported as elevated in lesional skin and/or serum,169–172 whereas MMP-12 has been described as decreased in serum but increased in lesional skin,170,173,174 and MMP-3 shows both up- and down-regulation depending on the study.172,175 This indicates an overall dysregulation of MMPs in psoriasis. Disrupted balance between MMP/TIMP and their normalization after effective treatment has also been observed.171,176
Plasminogen Activator Inhibitor-, Plasminogen Activator Inhibitor-2
Plasminogen Activator Inhibitor-1 (PAI-1) is a member of the serine protease inhibitor (SERPIN) family. PAI-1 inhibits plasminogen activators, which convert plasminogen into plasmin, thereby regulating the process of fibrinolysis. PAI-1 is one of the most extensively investigated mediators of cellular senescence and a key component of the SASP, playing a crucial role in maintaining the senescent phenotype. Furthermore, inhibition of PAI-1 limits cellular senescence and modulates its pathological consequences.177–179 Elevated levels of PAI-1 have been observed in individuals with psoriasis and have been shown to correlate with disease activity180,181 and might influence pathological vascular remodeling,180,182 suggesting a role for PAI-1 in sustaining the inflammatory processes characteristic of psoriasis.183 Additionally, another member of the SERPIN family-Plasminogen Activator Inhibitor-2 (PAI-2) - has been reported to be elevated in psoriatic lesions, although its role remains much less studied.184,185
Macrophage Migration Inhibitory Factor
Macrophage Migration Inhibitory Factor (MIF) is a pro-inflammatory cytokine and is associated with senescence-related inflammation and the SASP.186,187 Elevated levels of MIF have been reported in the lesional skin and serum of patients with psoriasis activity.188,189
Soluble or Shed Receptors or Ligands
Soluble or shed receptors and ligands including annexin A3 (ANXA3), Fas/CD95 and Fas Ligand (FasL), intercellular adhesion molecule-1 and −3 (ICAM-1, ICAM-3), osteoprotegerin (OPG), soluble tumor necrosis factor receptors I and II (sTNFRI, sTNFRII), and TNF-related apoptosis-inducing ligand receptor-3 (TRAIL-R3) contribute to SASP inflammatory signalling. ICAM-1, ICAM-3 are responsible for recruitment of leukocytes to the inflammatory site, in psoriasis ICAM-1 and −3 expression is increased in psoriatic lesions and serum and correlates with disease severity.190,191 Recent study demonstrated that ANXA 3 is highly expressed in psoriatic skin, and its suppression reduced inflammation and keratinocyte hyperproliferation in experimental model, however the data remain limited.192 Fas/CD95-FasL signalling contributes to the induction of apoptosis, in psoriasis Fas is upregulated and its alternative pro-inflammatory pathways are predominant, while abnormal apoptosis of keratinocytes is observed.193 TRAIL-R3/decoy receptor 1 (DcR1) is a receptor for TRAIL, which induces apoptosis and contributes to inflammatory signalling. TRAIL-R3 does not induce apoptosis but rather is thought to inhibit TRAIL-mediated apoptosis.194 In psoriasis, increased expression of TRAIL and its receptors has been observed in psoriatic lesions, suggesting its dysregulated signalling.194,195 Increased levels of sTNFRI/II have been observed in serum of patients with psoriasis, they influence TNF-α activity, which is involved in pathogenesis of psoriasis.196,197
Current Intervention Strategies Against Aging and Their Potential Application in Psoriasis
A healthy lifestyle, including a well-balanced diet, regular physical activity, high-quality sleep, and minimal toxic exposure, is considered to have a significant impact on reducing cellular senescence and preserving healthy life expectancy (HALE). These fundamental measures are essential strategies for reducing inflammaging.46,198,199 A healthy lifestyle is considered the most essential intervention in maintaining health and preventing pathological, preterm aging.198
Senolytics are agents that aim to selectively eliminate senescent cells. Dasatinib and quercetin, considered as senolytics, have demonstrated lifespan extension and improvement of multiple organ functions, including skin, in animal models.200 Moreover, a pilot study involving senolytic therapy in older adults at risk for Alzheimer’s disease showed preliminary good tolerability and revealed a correlation between improvements in cognitive function and reductions in SASP.201 Dasatinib and quercetin were shown to decrease senescent cell levels and circulating SASP factors in Phase II clinical trials for chronic kidney disease, which involved patients with diabetic nephropathy.202 In a mouse model of imiquimod-induced psoriasis-like skin inflammation, quercetin reduced serum levels of TNF-α, IL-6, and IL-17 and Psoriasis Area and Severity Index (PASI). Dasatinib combined with quercetin eliminated senescent cells and reduced the expression of SASP, resulting in an improvement of aging-related changes in human skin.203 Fenofibrate is also considered a senolytic; its mechanism of action also includes the elimination of senescent cells.204 In keratinocyte models and an imiquimod-induced psoriasis mouse model, fenofibrate was shown to reduce inflammatory cytokines (IL-1β, IL-6, IL-17A, TNF-α), and enhance autophagy, presenting anti-inflammatory effects.205 Moreover, patients with psoriasis showed clinical improvement after treatment with fibrates, and relapse of lesions after discontinuation, but the group described in the study consisted of only 2 patients.206 Curcumin shows senolytic activity in senescent human intervertebral disc cells by reducing SASP expression associated with inflammatory responses.207 Curcumin administered topically in imiquimod-induced psoriasis-like inflammation in mice, was shown to reduce the production of pro-inflammatory cytokines IL-1β and IL-6.208 In another small clinical study, niosomes with curcumin, applied topically, demonstrated improvements in mild-to-moderate psoriasis, reducing lesion severity and reducing levels of the IL-17/IL-23 axis cytokines.209 Further research and ongoing clinical trials may lead to new therapeutic solutions, with senolytics as potential novel therapeutics in inflammatory skin diseases.20,210 Measurements of the safety of such interventions are crucial, given the physiological roles of senescence eg. in wound-healing processes and in tumor surveillance.
Senomorphics are molecules that do not eliminate senescent cells but instead suppress their SASP and its role in sustaining inflammaging. These include metformin, mTOR inhibitors (rapamycin and its derivatives), and JAK/STAT inhibitors, and others.211 In the clinical trial Metformin in Longevity Study (MILES), metformin was shown to modulate the expression of age-related genes,212 and it has been investigated whether metformin may slow the development of age-associated chronic diseases in the research program - TAME (Targeting Aging with Metformin).213 Metformin can reduce chronic inflammation and oxidative damage.214 It has been shown to be beneficial for patients with psoriasis and comorbidities such as diabetes, insulin resistance, metabolic syndrome and obesity, likely due to metabolic improvement.215–217 The beneficial effects of metformin in patients with psoriasis without comorbidities are rather controversial.216 Rapamycin (sirolimus) is approved for the treatment of lymphangioleiomyomatosis and for the prevention of organ rejection after kidney transplantation.218,219 Rapamycin, mTOR inhibitor, shows anti-inflammatory characteristics by attenuating senescent cells’ pro-inflammatory properties.220 The role of mTOR inhibitors in psoriasis is limited. Imiquimod-induced psoriasis-like inflammation treated with rapamycin shows improvement of skin lesions.221 A small trial with topically applied sirolimus showed a reduction in clinical score, although the thickness of the plaques and erythema showed no significant improvement.222 Application of topically administered mTOR inhibitors in psoriasis requires further investigation.223 Upadacitinib, a JAK inhibitor, is approved in Europe for treatment of several conditions including psoriatic arthritis and atopic dermatitis.224 It has been demonstrated that Upadacitinib, administered for atopic dermatitis with concomitant psoriasis, resulted in the resolution of skin lesions,225 it has been also observed that in patients with atopic dermatitis dupilumab-induced psoriasis was successfully treated with Upadacitinib.226,227 Upadacitinib administered for psoriatic arthritis resulted in improvement in PASI.228
The clinical application of senolytic therapies in psoriasis remains limited, as most available evidence comes from animal or experimental studies; clinical trials in humans are rare and involve small patient cohorts, and reliable data on long-term safety and efficacy are still lacking.
Current knowledge about the role of SASP in psoriasis remains limited, even though SASP is recognized in skin ageing and inflammation broadly.33,229 Further research is needed to determine whether targeting SASP components could offer therapeutic benefits or improve treatment outcomes in psoriasis.20
Conclusions
The pathogenesis of psoriasis is closely associated with impaired regulation of cellular senescence, which emphasizes the importance of clarifying the mechanisms of inflammaging and the role of SASP in psoriasis. Key knowledge gaps include the identification of cell-type–specific roles of SASP components in lesional and systemic contexts in psoriasis, and to determine possible clinical benefits of the SASP profile in psoriasis. Future research should focus on the identification of reliable senescence and SASP biomarkers, longitudinal studies to clarify causality, and the development of scalable methods for patient stratification. Such efforts may enable more precise, personalized therapeutic strategies. The development of research focused on determining an individual SASP profile in patients could lead to the creation of clinically groundbreaking diagnostic and therapeutic tools that would enable personalized analysis of molecular, clinical, and environmental data, thereby allowing the development of highly personalized therapeutic protocols (Figure 3). Such an approach, consistent with the concept of patient-tailored treatment, increases the possibility of optimizing therapeutic effectiveness, minimizing adverse effects, and adopting a more precise approach to the disease. The concept of individualized SASP profiling is a largely speculative therapeutic and diagnostic option at this moment. While advances in omics technologies support the theoretical feasibility of this approach, significant challenges remain. These include high inter-individual variability, lack of standardized biomarkers, technical complexity, and costs. Consequently, routine implementation of this solution in clinical practice is not yet possible.
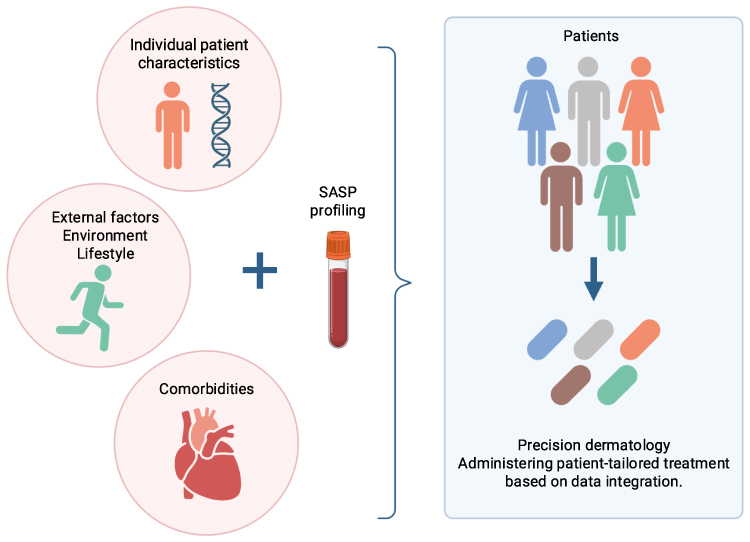 |
Figure 3 Potential development of patient-tailored treatment. Personalized analysis of patients’ individual characteristics, comorbidities, and external factors combined with individual SASP profiling might enable the development of highly personalized therapeutic protocols. Created in BioRender. |
Acknowledgments
Figures in graphical abstract created in BioRender.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no competing interests.
References
1. Parisi R, Iskandar IYK, Kontopantelis E, Augustin M, Griffiths CEM, Ashcroft DM. National, regional, and worldwide epidemiology of psoriasis: systematic analysis and modelling study. BMJ. 2020;369:m1590. doi:10.1136/bmj.m1590
2. Sewerin P, Brinks R, Schneider M, Haase I, Vordenbäumen S. Prevalence and incidence of psoriasis and psoriatic arthritis. Ann Rheum Dis. 2019;78(2):286–20. doi:10.1136/annrheumdis-2018-214065
3. Egeberg A, Andersen YMF, Thyssen JP. Prevalence and characteristics of psoriasis in Denmark: findings from the Danish skin cohort. BMJ Open. 2019;9(3):e028116. doi:10.1136/bmjopen-2018-028116
4. Damiani G, Bragazzi NL, Karimkhani Aksut C, et al. The Global, Regional, and National Burden of Psoriasis: results and Insights From the Global Burden of Disease 2019 Study. Front Med. 2021;8:743180. doi:10.3389/fmed.2021.743180
5. Daugaard C, Iversen L, Hjuler KF. Comorbidity in adult psoriasis: considerations for the clinician. Psoriasis Auckl NZ. 2022;12:139–150. doi:10.2147/PTT.S328572
6. Heidenreich R, Röcken M, Ghoreschi K. Angiogenesis drives psoriasis pathogenesis. Int J Exp Pathol. 2009;90(3):232–248. doi:10.1111/j.1365-2613.2009.00669.x
7. Zhou X, Chen Y, Cui L, Shi Y, Guo C. Advances in the pathogenesis of psoriasis: from keratinocyte perspective. Cell Death Dis. 2022;13(1):81. doi:10.1038/s41419-022-04523-3
8. Genome-wide scan reveals association of psoriasis with IL-23 and NF-κB pathways | Nature Genetics. Available from: https://www.nature.com/articles/ng.311.
9. RNF114 as a novel psoriasis susceptibility gene | Human Molecular Genetics | Oxford Academic. Available from: https://academic.oup.com/hmg/article-abstract/17/13/1938/2385713.
10. Cargill M, Schrodi SJ, Chang M, et al. A large-scale genetic association study confirms IL12B and leads to the identification of IL23R as psoriasis-risk genes. Am J Hum Genet. 2007;80(2):273–290. doi:10.1086/511051
11. Liu S, He M, Jiang J, et al. Triggers for the onset and recurrence of psoriasis: a review and update. Cell Commun Signal. 2024;22(1):108. doi:10.1186/s12964-023-01381-0
12. Sieminska I, Pieniawska M, Grzywa TM. The Immunology of Psoriasis—Current Concepts in Pathogenesis. Clin Rev Allergy Immunol. 2024;66(2):164–191. doi:10.1007/s12016-024-08991-7
13. von Zglinicki T, Saretzki G, Ladhoff J, d’Adda Di Fagagna F, Jackson SP. Human cell senescence as a DNA damage response. Mech Ageing Dev. 2005;126(1):111–117. doi:10.1016/j.mad.2004.09.034
14. Kumari R, Jat P. Mechanisms of cellular senescence: cell cycle arrest and senescence associated secretory phenotype. Front Cell Dev Biol. 2021;9. doi:10.3389/fcell.2021.645593
15. Shvedova M, Thanapaul RJRS, Thompson EL, Niedernhofer L, Roh DS. Cellular senescence in aging, tissue repair and regeneration. Plast Reconstr Surg. 2022;150:4S–11S. doi:10.1097/PRS.0000000000009667
16. Beranek M, Borsky P, Fiala Z, et al. Telomere length, oxidative and epigenetic changes in blood DNA of patients with exacerbated psoriasis vulgaris. An Bras Dermatol. 2023;98(1):68–74. doi:10.1016/j.abd.2022.01.008
17. Bell CG, Lowe R, Adams PD, et al. DNA methylation aging clocks: challenges and recommendations. Genome Biol. 2019;20:249. doi:10.1186/s13059-019-1824-y
18. Aslani S, Mirarefin SMJ, Zarredar H, et al. The epigenetic contribution to the pathogenesis of psoriasis: recent advances. Curr Med Chem. 2024;31(29):4621–4639. doi:10.2174/0929867330666230503143824
19. Dasgupta N, Arnold R, Equey A, Gandhi A, Adams PD. The role of the dynamic epigenetic landscape in senescence: orchestrating SASP expression. Npj Aging. 2024;10(1):48. doi:10.1038/s41514-024-00172-2
20. Thau H, Gerjol BP, Hahn K, et al. Senescence as a molecular target in skin aging and disease. Ageing Res Rev. 2025;105:102686. doi:10.1016/j.arr.2025.102686
21. Mercurio L, Bailey J, Glick AB, et al. RAS-activated PI3K/AKT signaling sustains cellular senescence via P53/P21 axis in experimental models of psoriasis. J Dermatol Sci. 2024;115(1):21–32. doi:10.1016/j.jdermsci.2024.03.002
22. Kovacs D, Falchi M, Cardinali G, et al. Immunohistochemical analysis of keratinocyte growth factor and fibroblast growth factor 10 expression in psoriasis. Exp Dermatol. 2005;14(2):130–137. doi:10.1111/j.0906-6705.2005.00261.x
23. Francis L, McCluskey D, Ganier C, et al. Single-cell analysis of psoriasis resolution demonstrates an inflammatory fibroblast state targeted by IL-23 blockade. Nat Commun. 2024;15(1):913. doi:10.1038/s41467-024-44994-w
24. Li Q, Pang B, Dang E, Wang G. Endothelial dysfunction in psoriasis: an integrative review. J Invest Dermatol. 2024;144(9):1935–1942. doi:10.1016/j.jid.2024.02.013
25. Zhu H, Jiang J, Yang M, et al. Topical application of a BCL-2 inhibitor ameliorates imiquimod-induced psoriasiform dermatitis by eliminating senescent cells. J Dermatol Sci. 2024;115(2):54–63. doi:10.1016/j.jdermsci.2024.06.002
26. Batista MD, Tincati C, Milush JM, et al. CD57 expression and cytokine production by T cells in lesional and unaffected skin from patients with psoriasis. PLoS One. 2013;8(2):e52144. doi:10.1371/journal.pone.0052144
27. Griffiths CE, Barker JN. Pathogenesis and clinical features of psoriasis. Lancet. 2007;370(9583):263–271. doi:10.1016/S0140-6736(07)61128-3
28. Orbai AM, Reddy SM, Dennis N, et al. Work absenteeism and disability associated with psoriasis and psoriatic arthritis in the USA-a retrospective study of claims data from 2009 TO 2020. Clin Rheumatol. 2021;40(12):4933–4942. doi:10.1007/s10067-021-05839-9
29. Armstrong AW, Schupp C, Wu J, Bebo B. Quality of life and work productivity impairment among psoriasis patients: findings from the National Psoriasis Foundation survey data 2003-2011. PLoS One. 2012;7(12):e52935. doi:10.1371/journal.pone.0052935
30. Bhosle MJ, Kulkarni A, Feldman SR, Balkrishnan R. Quality of life in patients with psoriasis. Health Qual Life Outcomes. 2006;4:35. doi:10.1186/1477-7525-4-35
31. Fitsiou E, Pulido T, Campisi J, Alimirah F, Demaria M. Cellular senescence and the senescence-associated secretory phenotype as drivers of skin photoaging. J Invest Dermatol. 2021;141(4, Supplement):1119–1126. doi:10.1016/j.jid.2020.09.031
32. Bellei B, Picardo M. Premature cell senescence in human skin: dual face in chronic acquired pigmentary disorders. Ageing Res Rev. 2020;57:100981. doi:10.1016/j.arr.2019.100981
33. Wang B, Han J, Elisseeff JH, Demaria M. The senescence-associated secretory phenotype and its physiological and pathological implications. Nat Rev Mol Cell Biol. 2024;25(12):958–978. doi:10.1038/s41580-024-00727-x
34. Ho CY, Dreesen O. Faces of cellular senescence in skin aging. Mech Ageing Dev. 2021;198:111525. doi:10.1016/j.mad.2021.111525
35. Hernandez-Segura A, de Jong TV, Melov S, Guryev V, Campisi J, Demaria M. Unmasking transcriptional heterogeneity in senescent cells. Curr Biol CB. 2017;27(17):2652–2660.e4. doi:10.1016/j.cub.2017.07.033
36. Sapieha P, Mallette FA. Cellular senescence in postmitotic cells: beyond growth arrest. Trends Cell Biol. 2018;28(8):595–607. doi:10.1016/j.tcb.2018.03.003
37. Cohn RL, Gasek NS, Kuchel GA, Xu M. The heterogeneity of cellular senescence: insights at the single-cell level. Trends Cell Biol. 2023;33(1):9–17. doi:10.1016/j.tcb.2022.04.011
38. Huang W, Hickson LJ, Eirin A, Kirkland JL, Lerman LO. Cellular senescence: the good, the bad and the unknown. Nat Rev Nephrol. 2022;18(10):611–627. doi:10.1038/s41581-022-00601-z
39. Lopes-Paciencia S, Saint-Germain E, Rowell MC, Ruiz AF, Kalegari P, Ferbeyre G. The senescence-associated secretory phenotype and its regulation. Cytokine. 2019;117:15–22. doi:10.1016/j.cyto.2019.01.013
40. Herranz N, Gil J. Mechanisms and functions of cellular senescence. J Clin Invest. 2018;128(4):1238–1246. doi:10.1172/JCI95148
41. Coppé JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol Mech Dis. 2010;5(5):2010):99–118. doi:10.1146/annurev-pathol-121808-102144
42. Pilkington SM, Bulfone-Paus S, Griffiths CEM, Watson REB. Inflammaging and the Skin. J Invest Dermatol. 2021;141(4):1087–1095. doi:10.1016/j.jid.2020.11.006
43. Garshick MS, Ward NL, Krueger JG, Berger JS. Cardiovascular risk in patients with psoriasis: JACC review topic of the week. J Am Coll Cardiol. 2021;77(13):1670–1680. doi:10.1016/j.jacc.2021.02.009
44. Mehta NN, Yu Y, Pinnelas R, et al. Attributable risk estimate of severe psoriasis on major cardiovascular events. Am J Med. 2011;124(8):775.e1–6. doi:10.1016/j.amjmed.2011.03.028
45. Gelfand JM, Troxel AB, Lewis JD, et al. The risk of mortality in patients with psoriasis: results from a population-based study. Arch Dermatol. 2007;143(12):1493–1499. doi:10.1001/archderm.143.12.1493
46. Zhao WQ, Gao R, Fan JY, Bu XL. Psoriasis and phenotypic age acceleration in relation to all-cause and cardiovascular disease mortality risks in US adults. Eur J Med Res. 2025;30(1):894. doi:10.1186/s40001-025-03176-0
47. Albanesi C, Pastore S. Pathobiology of chronic inflammatory skin diseases: interplay between keratinocytes and immune cells as a target for anti-inflammatory drugs. Curr Drug Metab. 2010;11(3):210–227. doi:10.2174/138920010791196328
48. Lin Z, fei WH, yan YL, et al. The relationship between biological aging and psoriasis: evidence from three observational studies. Immun Ageing. 2025;22(1):6. doi:10.1186/s12979-025-00500-4
49. Mahil SK, Capon F, Barker JN. Update on psoriasis immunopathogenesis and targeted immunotherapy. Semin Immunopathol. 2016;38(1):11–27. doi:10.1007/s00281-015-0539-8
50. Morar II, Tabăran FA, Mocan T, et al. Immunohistochemical study of psoriatic plaques and perilesional skin in psoriasis vulgaris patients: a pilot study. Exp Ther Med. 2019;18(2):888–894. doi:10.3892/etm.2019.7596
51. Ettehadi P, Greaves MW, Wallach D, Aderka D, Camp RDR. Elevated tumour necrosis factor-alpha (TNF-α) biological activity in psoriatic skin lesions. Clin Exp Immunol. 1994;96(1):146–151. doi:10.1111/j.1365-2249.1994.tb06244.x
52. Lowes MA, Suárez-Fariñas M, Krueger JG. Immunology of psoriasis. Annu Rev Immunol. 2014;32:227–255. doi:10.1146/annurev-immunol-032713-120225
53. Living EuroGuiDerm guideline for the systemic treatment of psoriasis vulgaris. Available from: https://guidelines.edf.one//guidelines/psoriasis-guideline.
54. Cavalli G, Colafrancesco S, Emmi G, et al. Interleukin 1α: a comprehensive review on the role of IL-1α in the pathogenesis and treatment of autoimmune and inflammatory diseases. Autoimmun Rev. 2021;20(3):102763. doi:10.1016/j.autrev.2021.102763
55. Ghosh K, Capell BC. The senescence-associated secretory phenotype: critical effector in skin cancer and aging. J Invest Dermatol. 2016;136(11):2133–2139. doi:10.1016/j.jid.2016.06.621
56. Jarrold BB, Tan CYR, Ho CY, et al. Early onset of senescence and imbalanced epidermal homeostasis across the decades in photoexposed human skin: fingerprints of inflammaging. Exp Dermatol. 2022;31(11):1748–1760. doi:10.1111/exd.14654
57. Debets R, Hegmans JP, Croughs P, et al. The IL-1 system in psoriatic skin: IL-1 antagonist sphere of influence in lesional psoriatic epidermis. J Immunol. 1997;158(6):2955–2963.
58. Baliwag J, Barnes DH, Johnston A. Cytokines in psoriasis. Cytokine. 2015;73(2):342–350. doi:10.1016/j.cyto.2014.12.014
59. Iznardo H, Puig L. The interleukin-1 family cytokines in psoriasis: pathogenetic role and therapeutic perspectives. Expert Rev Clin Immunol. 2021;17(2):187–199. doi:10.1080/1744666X.2021.1886081
60. Ge Y, Li M, Bai S, et al. Doxercalciferol alleviates UVB-induced HaCaT cell senescence and skin photoaging. Int Immunopharmacol. 2024;127:111357. doi:10.1016/j.intimp.2023.111357
61. Macleod T, Berekmeri A, Bridgewood C, Stacey M, McGonagle D, Wittmann M. The immunological impact of IL-1 family cytokines on the epidermal barrier. Front Immunol. 2021;12:808012. doi:10.3389/fimmu.2021.808012
62. Cai Y, Xue F, Quan C, et al. A critical role of the IL-1β-IL-1R signaling pathway in skin inflammation and psoriasis pathogenesis. J Invest Dermatol. 2019;139(1):146–156. doi:10.1016/j.jid.2018.07.025
63. Gabay C. Interleukin-6 and chronic inflammation. Arthritis Res Ther. 2006;8(Suppl 2):S3. doi:10.1186/ar1917
64. Pappalardo A, Kim JY, Abaci HE, Christiano AM. Restoration of hair follicle inductive properties by depletion of senescent cells. Aging Cell. 2025;24(1):e14353. doi:10.1111/acel.14353
65. Goodman WA, Levine AD, Massari JV, Sugiyama H, McCormick TS, Cooper KD. IL-6 signaling in psoriasis prevents immune suppression by regulatory T cells. J Immunol Baltim. 2009;183(5):3170–3176. doi:10.4049/jimmunol.0803721
66. Vičić M, Kaštelan M, Brajac I, Sotošek V, Massari LP. Current concepts of psoriasis immunopathogenesis. Int J Mol Sci. 2021;22(21):11574. doi:10.3390/ijms222111574
67. Chen D, Tang TX, Deng H, Yang XP, Tang ZH. Interleukin-7 biology and its effects on immune cells: mediator of generation, differentiation, survival, and homeostasis. Front Immunol. 2021;12:747324. doi:10.3389/fimmu.2021.747324
68. Szepietowski JC, Bielicka E, Nockowski P, Noworolska A, Wasik F. Increased interleukin-7 levels in the sera of psoriatic patients: lack of correlations with interleukin-6 levels and disease intensity. Clin Exp Dermatol. 2000;25(8):643–647. doi:10.1046/j.1365-2230.2000.00727.x
69. Bonifati C, Trento E, Cordiali-Fei P, et al. Increased interleukin-7 concentrations in lesional skin and in the sera of patients with plaque-type psoriasis. Clin Immunol Immunopathol. 1997;83(1):41–44. doi:10.1006/clin.1996.4313
70. Vranova M, Friess MC, Haghayegh Jahromi N, et al. Opposing roles of endothelial and leukocyte-expressed IL-7Rα in the regulation of psoriasis-like skin inflammation. Sci Rep. 2019;9(1):11714. doi:10.1038/s41598-019-48046-y
71. (PDF) Interleukin-13 promotes cellular senescence through inducing mitochondrial dysfunction in IgG4-related sialadenitis. ResearchGate; 2025. doi:10.1038/s41368-022-00180-6.
72. Fehniger TA, Caligiuri MA. Interleukin 15: biology and relevance to human disease. Blood. 2001;97(1):14–32. doi:10.1182/blood.v97.1.14
73. De Jesús-Gil C, Ruiz-Romeu E, Ferran M, et al. IL-15 and IL-23 synergize to trigger Th17 response by CLA+ T cells in psoriasis. Exp Dermatol. 2020;29(7):630–638. doi:10.1111/exd.14113
74. Villadsen LS, Schuurman J, Beurskens F, et al. Resolution of psoriasis upon blockade of IL-15 biological activity in a xenograft mouse model. J Clin Invest. 2003;112(10):1571–1580. doi:10.1172/JCI18986
75. Rasmy H, Mikhael N, Ismail S. Interleukin-18 expression and the response to treatment in patients with psoriasis. Arch Med Sci AMS. 2011;7(4):713–719. doi:10.5114/aoms.2011.24144
76. Lee JH, Cho DH, Park HJ, Lee JH, Cho DH, Park HJ. IL-18 and cutaneous inflammatory diseases. Int J Mol Sci. 2015;16(12):29357–29369. doi:10.3390/ijms161226172
77. Girolomoni G, Strohal R, Puig L, et al. The role of IL-23 and the IL-23/TH 17 immune axis in the pathogenesis and treatment of psoriasis. J Eur Acad Dermatol Venereol JEADV. 2017;31(10):1616–1626. doi:10.1111/jdv.14433
78. Volpe E, Servant N, Zollinger R, et al. A critical function for transforming growth factor-beta, interleukin 23 and proinflammatory cytokines in driving and modulating human T(H)-17 responses. Nat Immunol. 2008;9(6):650–657. doi:10.1038/ni.1613
79. Lynde CW, Poulin Y, Vender R, Bourcier M, Khalil S. Interleukin 17A: toward a new understanding of psoriasis pathogenesis. J Am Acad Dermatol. 2014;71(1):141–150. doi:10.1016/j.jaad.2013.12.036
80. Liang SC, Tan XY, Luxenberg DP, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J Exp Med. 2006;203(10):2271–2279. doi:10.1084/jem.20061308
81. Wolk K, Witte E, Wallace E, et al. IL-22 regulates the expression of genes responsible for antimicrobial defense, cellular differentiation, and mobility in keratinocytes: a potential role in psoriasis. Eur J Immunol. 2006;36(5):1309–1323. doi:10.1002/eji.200535503
82. Weber A, Wasiliew P, Kracht M. Interleukin-1 (IL-1) pathway. Sci Signal. 2010;3(105):cm1. doi:10.1126/scisignal.3105cm1
83. Uciechowski P, Dempke WCM. Interleukin-6: a Masterplayer in the Cytokine Network. Oncology. 2020;98(3):131–137. doi:10.1159/000505099
84. Bodoor K, Al-Qarqaz F, Heis LA, et al. IL-33/13 axis and IL-4/31 axis play distinct roles in inflammatory process and itch in psoriasis and atopic dermatitis. Clin Cosmet Invest Dermatol. 2020;13:419–424. doi:10.2147/CCID.S257647
85. Zhu M, Min S, Mao X, et al. Interleukin-13 promotes cellular senescence through inducing mitochondrial dysfunction in IgG4-related sialadenitis. Int J Oral Sci. 2022;14(1):29. doi:10.1038/s41368-022-00180-6
86. Hughes CE, Nibbs RJB. A guide to chemokines and their receptors. FEBS J. 2018;285(16):2944–2971. doi:10.1111/febs.14466
87. Zdanowska N, Kasprowicz-Furmańczyk M, Placek W, et al. The role of chemokines in psoriasis—an overview. Medicina. 2021;57(8). doi:10.3390/medicina57080754
88. Guo J, Zhang H, Lin W, Lu L, Su J, Chen X. Signaling pathways and targeted therapies for psoriasis. Signal Transduct Target Ther. 2023;8(1):437. doi:10.1038/s41392-023-01655-6
89. Elnabawi YA, Garshick MS, Tawil M, et al. CCL20 in psoriasis: a potential biomarker of disease severity, inflammation, and impaired vascular health. J Am Acad Dermatol. 2021;84(4):913–920. doi:10.1016/j.jaad.2020.10.094
90. Abji F, Pollock RA, Liang K, Chandran V, Gladman DD. Brief report: CXCL10 is a possible biomarker for the development of psoriatic arthritis among patients with psoriasis. Arthritis Rheumatol Hoboken NJ. 2016;68(12):2911–2916. doi:10.1002/art.39800
91. Samotij D, Nedoszytko B, Bartosińska J, et al. Pathogenesis of psoriasis in the “omic” era. Part I. Epidemiology, clinical manifestation, immunological and neuroendocrine disturbances. Adv Dermatol Allergol Dermatol Alergol. 2020;37(2):135–153. doi:10.5114/ada.2020.94832
92. Vestergaard C, Just H, Baumgartner Nielsen J, Thestrup-Pedersen K, Deleuran M. Expression of CCR2 on monocytes and macrophages in chronically inflamed skin in atopic dermatitis and psoriasis. Acta Derm Venereol. 2004;84(5):353–358. doi:10.1080/00015550410034444
93. Behfar S, Hassanshahi G, Nazari A, Khorramdelazad H. A brief look at the role of monocyte chemoattractant protein-1 (CCL2) in the pathophysiology of psoriasis. Cytokine. 2018;110:226–231. doi:10.1016/j.cyto.2017.12.010
94. Nan L, Guo P, Hui W, Xia F, Yi C. Recent advances in dermal fibroblast senescence and skin aging: unraveling mechanisms and pioneering therapeutic strategies. Front Pharmacol. 2025;16:1592596. doi:10.3389/fphar.2025.1592596
95. Purzycka-Bohdan D, Nedoszytko B, Zabłotna M, Gleń J, Szczerkowska-Dobosz A, Nowicki RJ. Chemokine profile in psoriasis patients in correlation with disease severity and pruritus. Int J Mol Sci. 2022;23(21):13330. doi:10.3390/ijms232113330
96. Dai YJ, Li YY, Zeng HM, et al. Effect of pharmacological intervention on MIP-1α, MIP-1β and MCP-1 expression in patients with psoriasis vulgaris. Asian Pac J Trop Med. 2014;7(7):582–584. doi:10.1016/S1995-7645(14)60098-5
97. Basisty N, Kale A, Jeon OH, et al. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol. 2020;18(1):e3000599. doi:10.1371/journal.pbio.3000599
98. de Groot M, Teunissen MBM, Ortonne JP, et al. Expression of the chemokine receptor CCR5 in psoriasis and results of a randomized placebo controlled trial with a CCR5 inhibitor. Arch Dermatol Res. 2007;299(7):305–313. doi:10.1007/s00403-007-0764-7
99. Ottaviani C, Nasorri F, Bedini C, de Pità O, Girolomoni G, Cavani A. CD56brightCD16(-) NK cells accumulate in psoriatic skin in response to CXCL10 and CCL5 and exacerbate skin inflammation. Eur J Immunol. 2006;36(1):118–128. doi:10.1002/eji.200535243
100. Hattangady NG, Carter K, Maroni-Rana B, et al. Mapping the core senescence phenotype of primary human colon fibroblasts. Aging. 2024;16(4):3068–3087. doi:10.18632/aging.205577
101. Harper EG, Guo C, Rizzo H, et al. Th17 cytokines stimulate CCL20 expression in keratinocytes in vitro and in vivo: implications for psoriasis pathogenesis. J Invest Dermatol. 2009;129(9):2175–2183. doi:10.1038/jid.2009.65
102. Nedoszytko B, Sokołowska-Wojdyło M, Ruckemann-Dziurdzińska K, Roszkiewicz J, Nowicki RJ. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: atopic dermatitis, psoriasis and skin mastocytosis. Postepy Dermatol Alergol. 2014;31(2):84–91. doi:10.5114/pdia.2014.40920
103. Uribe-Herranz M, Lian LH, Hooper KM, Milora KA, Jensen LE. IL-1R1 signaling facilitates Munro’s microabscess formation in psoriasiform imiquimod-induced skin inflammation. J Invest Dermatol. 2013;133(6):1541–1549. doi:10.1038/jid.2012.512
104. Lowes MA, Bowcock AM, Krueger JG. Pathogenesis and therapy of psoriasis. Nature. 2007;445(7130):866–873. doi:10.1038/nature05663
105. Russo RC, Garcia CC, Teixeira MM, Amaral FA. The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases. Expert Rev Clin Immunol. 2014;10(5):593–619. doi:10.1586/1744666X.2014.894886
106. Flier J, Boorsma DM, van Beek PJ, et al. Differential expression of CXCR3 targeting chemokines CXCL10, CXCL9, and CXCL11 in different types of skin inflammation. J Pathol. 2001;194(4):398–405.
107. Krueger JG, Krane JF, Carter DM, Gottlieb AB. Role of growth factors, cytokines, and their receptors in the pathogenesis of psoriasis. J Invest Dermatol. 1990;94(6 Suppl):135S–140S. doi:10.1111/1523-1747.ep12876121
108. Canavese M, Altruda F, Ruzicka T, Schauber J. Vascular endothelial growth factor (VEGF) in the pathogenesis of psoriasis--a possible target for novel therapies? J Dermatol Sci. 2010;58(3):171–176. doi:10.1016/j.jdermsci.2010.03.023
109. Bhushan M, McLaughlin B, Weiss JB, Griffiths CE. Levels of endothelial cell stimulating angiogenesis factor and vascular endothelial growth factor are elevated in psoriasis. Br J Dermatol. 1999;141(6):1054–1060. doi:10.1046/j.1365-2133.1999.03205.x
110. Luengas-Martinez A, Paus R, Young HS. Antivascular endothelial growth factor-A therapy: a novel personalized treatment approach for psoriasis. Br J Dermatol. 2022;186(5):782–791. doi:10.1111/bjd.20940
111. Xiong J, Dong L, Lv Q, et al. Targeting senescence-associated secretory phenotypes to remodel the tumour microenvironment and modulate tumour outcomes. Clin Transl Med. 2024;14(9):e1772. doi:10.1002/ctm2.1772
112. Yurekli UF, Akkus MR, Kirmit A. Serum fibroblast growth factor 2 level in psoriasis; relationship with disease severity. Ann Med Res. 2023;30(7):774–777.
113. Krane JF, Murphy DP, Gottlieb AB, Carter DM, Hart CE, Krueger JG. Increased dermal expression of platelet-derived growth factor receptors in growth-activated skin wounds and psoriasis. J Invest Dermatol. 1991;96(6):983–986. doi:10.1111/1523-1747.ep12476485
114. Kita A, Yamamoto S, Saito Y, Chikenji TS. Cellular senescence and wound healing in aged and diabetic skin. Front Physiol. 2024;15. doi:10.3389/fphys.2024.1344116
115. Wang S, Zhang Z, Peng H, Zeng K. Recent advances on the roles of epidermal growth factor receptor in psoriasis. Am J Transl Res. 2019;11(2):520–528.
116. Szterling-Jaworowska M, Flisiak I, Baran A, Chodynicka B. The role of epidermal growth factor in psoriasis. Dermatol Rev Dermatol. 2009;96(6):435–439.
117. Mercurio L, Lulli D, Mascia F, et al. Intracellular insulin-like growth factor binding protein 2 (IGFBP2) contributes to the senescence of keratinocytes in psoriasis by stabilizing cytoplasmic p21. Aging. 2020;12(8):6823–6851. doi:10.18632/aging.103045
118. Detmar M, Brown LF, Claffey KP, et al. Overexpression of vascular permeability factor/vascular endothelial growth factor and its receptors in psoriasis. J Exp Med. 1994;180(3):1141–1146. doi:10.1084/jem.180.3.1141
119. Flisiak I, Zaniewski P, Rogalska M, Myśliwiec H, Jaroszewicz J, Chodynicka B. Effect of psoriasis activity on VEGF and its soluble receptors concentrations in serum and plaque scales. Cytokine. 2010;52(3):225–229. doi:10.1016/j.cyto.2010.09.012
120. Gerkowicz A, Socha M, Pietrzak A, Zubilewicz T, Krasowska D. The role of VEGF in psoriasis: an update. Acta Angiol. 2018;24(4):134–140. doi:10.5603/AA.2018.0019
121. Veikkola T, Karkkainen M, Claesson-Welsh L, Alitalo K. Regulation of angiogenesis via vascular endothelial growth factor receptors. Cancer Res. 2000;60(2):203–212.
122. Gupta S, Kaur M, Gupta R, Singh S, Pant L, Singh PP. Dermal vasculature in psoriasis and psoriasiform dermatitis: a morphometric study. Indian J Dermatol. 2011;56(6):647–649. doi:10.4103/0019-5154.91820
123. Kaliyadan F. The dermoscopic Auspitz sign. Indian Dermatol Online J. 2018;9(4):290–291. doi:10.4103/idoj.IDOJ_309_17
124. Markham T, Mullan R, Golden-Mason L, et al. Resolution of endothelial activation and down-regulation of Tie2 receptor in psoriatic skin after infliximab therapy. J Am Acad Dermatol. 2006;54(6):1003–1012. doi:10.1016/j.jaad.2006.01.038
125. Campanati A, Goteri G, Simonetti O, et al. Angiogenesis in psoriatic skin and its modifications after administration of etanercept: videocapillaroscopic, histological and immunohistochemical evaluation. Int J Immunopathol Pharmacol. 2009;22(2):371–377. doi:10.1177/039463200902200214
126. Hanssen SCA, van der Vleuten CJM, van Erp PEJ, Seyger MMB, van de Kerkhof PCM. The effect of adalimumab on the vasculature in psoriatic skin lesions. J Dermatol Treat. 2019;30(3):221–226. doi:10.1080/09546634.2018.1506082
127. Narayanan S, Callis-Duffin K, Batten J, Agarwal N. Improvement of psoriasis during sunitinib therapy for renal cell carcinoma. Am J Med Sci. 2010;339(6):580–581. doi:10.1097/MAJ.0b013e3181dd1aa5
128. Fournier C, Tisman G. Sorafenib-associated remission of psoriasis in hypernephroma: case report. Dermatol Online J. 2010;16(2):17.
129. Datta-Mitra A, Riar NK, Raychaudhuri SP. Remission of psoriasis and psoriatic arthritis during bevacizumab therapy for renal cell cancer. Indian J Dermatol. 2014;59(6):632. doi:10.4103/0019-5154.143574
130. Akman A, Yilmaz E, Mutlu H, Ozdogan M. Complete remission of psoriasis following bevacizumab therapy for colon cancer. Clin Exp Dermatol. 2009;34(5):e202–204. doi:10.1111/j.1365-2230.2008.02991.x
131. Xia YP, Li B, Hylton D, Detmar M, Yancopoulos GD, Rudge JS. Transgenic delivery of VEGF to mouse skin leads to an inflammatory condition resembling human psoriasis. Blood. 2003;102(1):161–168. doi:10.1182/blood-2002-12-3793
132. Boonpethkaew S, Meephansan J, Ponnikorn S, et al. Exploring the role of growth factors as potential regulators in psoriatic plaque formation. Exp Dermatol. 2023;32(11):1924–1934. doi:10.1111/exd.14918
133. Richarz NA, Boada A, Carrascosa JM. Angiogenesis in dermatology – insights of molecular mechanisms and latest developments. Actas Dermo-Sifiliográficas. 2017;108(6):515–523. doi:10.1016/j.ad.2016.12.001
134. Matwiejuk M, Myśliwiec H, Chabowski A, et al. An overview of growth factors as the potential link between psoriasis and metabolic syndrome. J Clin Med. 2023;13(1). doi:10.3390/jcm13010109
135. Schneider MR, Wolf E. The epidermal growth factor receptor ligands at a glance. J Cell Physiol. 2009;218(3):460–466. doi:10.1002/jcp.21635
136. Flisiak I, Szterling-Jaworowska M, Baran A, Rogalska-Taranta M. Effect of psoriasis activity on epidermal growth factor (EGF) and the concentration of soluble EGF receptor in serum and plaque scales. Clin Exp Dermatol. 2014;39(4):461–467. doi:10.1111/ced.12356
137. Nanney LB, Stoscheck CM, Magid M, King LE. Altered [125I]epidermal growth factor binding and receptor distribution in psoriasis. J Invest Dermatol. 1986;86(3):260–265. doi:10.1111/1523-1747.ep12285389
138. Baxter RC. Signaling pathways of the insulin-like growth factor binding proteins. Endocr Rev. 2023;44(5):753–778. doi:10.1210/endrev/bnad008
139. Ozden MG, Ilter N, Suer OE, Solaktekin N, Gürer MA. Increased insulin-like growth factor binding protein-3 expression level in psoriatic tissue before and after systemic treatment with cyclosporine A and methotrexate. Int J Dermatol. 2008;47(11):1177–1183. doi:10.1111/j.1365-4632.2008.03810.x
140. Wraight CJ, Edmondson SR, Fortune DW, Varigos G, Werther GA. Expression of insulin-like growth factor binding protein-3 (IGFBP-3) in the psoriatic lesion. J Invest Dermatol. 1997;108(4):452–456. doi:10.1111/1523-1747.ep12289713
141. Nousbeck J, Sarig O, Avidan N, et al. Insulin-like growth factor-binding protein 7 regulates keratinocyte proliferation, differentiation and apoptosis. J Invest Dermatol. 2010;130(2):378–387. doi:10.1038/jid.2009.265
142. Li Q, Shao S, Zhu Z, et al. An IGFBP7hi endothelial cell subset drives T cell extravasation in psoriasis via endothelial glycocalyx degradation. J Clin Invest. 2023;133(9):e160451. doi:10.1172/JCI160451
143. Nousbeck J, Ishida-Yamamoto A, Bidder M, et al. IGFBP7 as a potential therapeutic target in psoriasis. J Invest Dermatol. 2011;131(8):1767–1770. doi:10.1038/jid.2011.108
144. Vidal S, Puig L, Carrascosa-Carrillo JM, González-Cantero Á, Ruiz-Carrascosa JC, Velasco-Pastor AM. From messengers to receptors in psoriasis: the role of IL-17RA in disease and treatment. Int J Mol Sci. 2021;22(13):6740. doi:10.3390/ijms22136740
145. Furue M, Furue K, Tsuji G, Nakahara T. Interleukin-17A and keratinocytes in psoriasis. Int J Mol Sci. 2020;21(4):1275. doi:10.3390/ijms21041275
146. Legler DF, Loetscher M, Roos RS, Clark-Lewis I, Baggiolini M, Moser B. B cell-attracting chemokine 1, a human CXC chemokine expressed in lymphoid tissues, selectively attracts B lymphocytes via BLR1/CXCR5. J Exp Med. 1998;187(4):655–660. doi:10.1084/jem.187.4.655
147. Schulz O, Hammerschmidt SI, Moschovakis GL, Förster R. Chemokines and chemokine receptors in lymphoid tissue dynamics. Annu Rev Immunol. 2016;34:203–242. doi:10.1146/annurev-immunol-041015-055649
148. Pan Z, Zhu T, Liu Y, Zhang N. Role of the CXCL13/CXCR5 axis in autoimmune diseases. Front Immunol. 2022;13. doi:10.3389/fimmu.2022.850998
149. Liu J, Chang HW, Huang ZM, et al. Single-cell RNA sequencing of psoriatic skin identifies pathogenic Tc17 cell subsets and reveals distinctions between CD8+ T cells in autoimmunity and cancer. J Allergy Clin Immunol. 2021;147(6):2370–2380. doi:10.1016/j.jaci.2020.11.028
150. Chen Y, Wu J, Wong C, Gao W, Qi X, Zhou H. Disturbed glycolipid metabolism activates CXCL13-CXCR5 axis in senescent TSCs to promote heterotopic ossification. Cell Mol Life Sci CMLS. 2024;81(1):265. doi:10.1007/s00018-024-05302-3
151. Zhao H, Liu Z, Chen H, et al. Identifying specific functional roles for senescence across cell types. Cell. 2024;187(25):7314–7334.e21. doi:10.1016/j.cell.2024.09.021
152. Aggarwal N, Sloane BF. Cathepsin B: multiple roles in cancer. Proteomics Clin Appl. 2014;8(5–6):427–437. doi:10.1002/prca.201300105
153. Oh SS, Park S, Lee KW, et al. Extracellular cystatin SN and cathepsin B prevent cellular senescence by inhibiting abnormal glycogen accumulation. Cell Death Dis. 2017;8(4):e2729–e2729. doi:10.1038/cddis.2017.153
154. West PW, Tontini C, Atmoko H, et al. Human mast cells upregulate Cathepsin B, a novel marker of itch in psoriasis. Cells. 2023;12(17):2177. doi:10.3390/cells12172177
155. Willoughby DA, Moore AR, Colville-Nash PR. COX-1, COX-2, and COX-3 and the future treatment of chronic inflammatory disease. Lancet Lond Engl. 2000;355(9204):646–648. doi:10.1016/S0140-6736(99)12031-2
156. Crofford LJ. COX-1 and COX-2 tissue expression: implications and predictions. J Rheumatol Suppl. 1997;49:15–19.
157. Garshick MS, Tawil M, Barrett TJ, et al. Activated platelets induce endothelial cell inflammatory response in psoriasis via COX-1. Arterioscler Thromb Vasc Biol. 2020;40(5):1340–1351. doi:10.1161/ATVBAHA.119.314008
158. Arasa J, Terencio MC, Andrés RM, et al. Defective induction of COX-2 expression by psoriatic fibroblasts promotes pro-inflammatory activation of macrophages. Front Immunol. 2019:10. doi:10.3389/fimmu.2019.00536
159. Klemann C, Wagner L, Stephan M, von Hörsten S. Cut to the chase: a review of CD26/dipeptidyl peptidase-4’s (DPP4) entanglement in the immune system. Clin Exp Immunol. 2016;185(1):1–21. doi:10.1111/cei.12781
160. Van Lingen RG, PCM VDK, Seyger MMB, et al. CD26/dipeptidyl-peptidase IV in psoriatic skin: upregulation and topographical changes. Br J Dermatol. 2008;158(6):1264–1272. doi:10.1111/j.1365-2133.2008.08515.x
161. Komiya E, Hatano R, Otsuka H, et al. A possible role for CD26/DPPIV enzyme activity in the regulation of psoriatic pruritus. J Dermatol Sci. 2017;86(3):212–221. doi:10.1016/j.jdermsci.2017.03.005
162. Clark SC, Kamen R. The human hematopoietic colony-stimulating factors. Science. 1987;236(4806):1229–1237. doi:10.1126/science.3296190
163. Bonifati C, Carducci M, Cordiali Fei P, et al. Correlated increases of tumour necrosis factor-alpha, interleukin-6 and granulocyte monocyte-colony stimulating factor levels in suction blister fluids and sera of psoriatic patients--relationships with disease severity. Clin Exp Dermatol. 1994;19(5):383–387. doi:10.1111/j.1365-2230.1994.tb02687.x
164. Alvarez-Ruiz S, Peñas PF, Fernández-Herrera J, Sánchez-Pérez J, Fraga J, García-Díez A. Maculopapular eruption with enlarged macrophages in eight patients receiving G-CSF or GM-CSF. J Eur Acad Dermatol Venereol JEADV. 2004;18(3):310–313. doi:10.1111/j.1468-3083.2004.00872.x
165. Kelly R, Marsden RA, Bevan D. Exacerbation of psoriasis with GM-CSF therapy. Br J Dermatol. 1993;128(4):468–469. doi:10.1111/j.1365-2133.1993.tb00218.x
166. Upasana M, Yin WY, Bo WJ, Quan SJ. Granulocyte-Colony stimulating factor induced early psoriasis - a case report and literature review. Our Dermatol Online. 2020;11(1):56–58. doi:10.7241/ourd.20201.14
167. Papp KA, Gooderham M, Jenkins R, et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) as a therapeutic target in psoriasis: randomized, controlled investigation using namilumab, a specific human anti-GM-CSF monoclonal antibody. Br J Dermatol. 2019;180(6):1352–1360. doi:10.1111/bjd.17195
168. Cabral-Pacheco GA, Garza-Veloz I, Castruita-De la Rosa C, et al. The roles of matrix metalloproteinases and their inhibitors in human diseases. Int J Mol Sci. 2020;21(24):9739. doi:10.3390/ijms21249739
169. Wagner MFMG, Theodoro TR, CDASM F, Oyafuso LKM, Pinhal MAS. Extracellular matrix alterations in the skin of patients affected by psoriasis. BMC Mol Cell Biol. 2021;22(1):55. doi:10.1186/s12860-021-00395-1
170. Mezentsev A, Nikolaev A, Bruskin S. Matrix metalloproteinases and their role in psoriasis. Gene. 2014;540(1):1–10. doi:10.1016/j.gene.2014.01.068
171. Flisiak I, Myśliwiec H, Chodynicka B. Effect of psoriasis treatment on plasma concentrations of metalloproteinase-1 and tissue inhibitor of metalloproteinases-1. J Eur Acad Dermatol Venereol JEADV. 2005;19(4):418–421. doi:10.1111/j.1468-3083.2005.01199.x
172. Michalak-Stoma A, Bartosińska J, Raczkiewicz D, Kowal M, Krasowska D, Chodorowska G. Assessment of selected Matrix Metalloproteinases (MMPs) and correlation with cytokines in psoriatic patients. Mediators Inflamm. 2021;2021:9913798. doi:10.1155/2021/9913798
173. Głażewska EK, Niczyporuk M, Przylipiak A, et al. Influence of narrowband ultraviolet-B phototherapy on plasma concentration of matrix metalloproteinase-12 in psoriatic patients. Postepy Dermatol Alergol. 2017;34(4):328–333. doi:10.5114/ada.2017.69312
174. Suomela S, Kariniemi AL, Snellman E, Saarialho-Kere U. Metalloelastase (MMP-12) and 92-kDa gelatinase (MMP-9) as well as their inhibitors, TIMP-1 and −3, are expressed in psoriatic lesions. Exp Dermatol. 2001;10(3):175–183. doi:10.1034/j.1600-0625.2001.010003175.x
175. Glazewska EK, Niczyporuk M, Lawicki S, et al. ROC analysis of selected matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs) in psoriatic patients. Adv Dermatol Allergol Dermatol Alergol. 2018;35(2):167–173. doi:10.5114/pdia.2017.66621
176. Głażewska EK, Niczyporuk M, Ławicki S, et al. Therapy of psoriasis with narrowband ultraviolet-B light influences plasma concentrations of MMP-2 and TIMP-2 in patients. Ther Clin Risk Manag. 2016;12:1579–1585. doi:10.2147/TCRM.S113769
177. Vaughan DE, Rai R, Khan SS, Eren M, Ghosh AK. PAI-1 is a marker and a mediator of senescence. Arterioscler Thromb Vasc Biol. 2017;37(8):1446–1452. doi:10.1161/ATVBAHA.117.309451
178. Khan SS, Shah SJ, Klyachko E, et al. A null mutation in SERPINE1 protects against biological aging in humans. Sci Adv. 2017;3(11):eaao1617. doi:10.1126/sciadv.aao1617
179. Sillen M, Declerck PJ, Sillen M, Declerck PJ. A narrative review on plasminogen activator inhibitor-1 and its (Patho)physiological role: to target or not to target? Int J Mol Sci. 2021;22(5). doi:10.3390/ijms22052721
180. Nielsen HJ, Christensen IJ, Svendsen MN, et al. Elevated plasma levels of vascular endothelial growth factor and plasminogen activator inhibitor-1 decrease during improvement of psoriasis. Inflamm Res off J Eur Histamine Res Soc Al. 2002;51(11):563–567. doi:10.1007/pl00012428
181. Teixeira GG, Mari NL, de Paula JCC, et al. Cell adhesion molecules, plasminogen activator inhibitor type 1, and metabolic syndrome in patients with psoriasis. Clin Exp Med. 2020;20(1):39–48. doi:10.1007/s10238-019-00595-2
182. Fujimura T, Asano Y. PAI-1 in skin malignancies: a central regulator of tumor progression and therapeutic resistance. Curr Treat Options Oncol. 2025;26(11):943–949. doi:10.1007/s11864-025-01357-x
183. Rubina KA, Sysoeva VY, Zagorujko EI, et al. Increased expression of uPA, uPAR, and PAI-1 in psoriatic skin and in basal cell carcinomas. Arch Dermatol Res. 2017;309(6):433–442. doi:10.1007/s00403-017-1738-z
184. Vaher H, Kivihall A, Runnel T, et al. SERPINB2 and miR-146a/b are coordinately regulated and act in the suppression of psoriasis-associated inflammatory responses in keratinocytes. Exp Dermatol. 2020;29(1):51–60. doi:10.1111/exd.14049
185. Wang J, Li J, Zhou L, Hou H, Zhang K. Regulation of epidermal barrier function and pathogenesis of psoriasis by serine protease inhibitors. Front Immunol. 2024;15. doi:10.3389/fimmu.2024.1498067
186. Barthelmess RM, Stijlemans B, Ginderachter JAV, Barthelmess RM, Stijlemans B, Ginderachter JAV. Hallmarks of cancer affected by the MIF cytokine family. Cancers. 2023;15(2). doi:10.3390/cancers15020395
187. Wu S, Ouyang Y, Hu Y, et al. Macrophage migration inhibitory factor mediates skin aging via CD74: insights from single-cell and bulk RNA sequencing data. Clin Immunol Orlando Fla. 2024;263:110199. doi:10.1016/j.clim.2024.110199
188. Bezdek S, Leng L, Busch H, et al. Macrophage Migration Inhibitory Factor (MIF) drives murine psoriasiform dermatitis. Front Immunol. 2018:9. doi:10.3389/fimmu.2018.02262
189. Shimizu T, Nakamura H, Abe R, et al. High macrophage Migration Inhibitory Factor (MIF) serum levels associated with extended psoriasis. J Invest Dermatol. 2001;116(6):989–990. doi:10.1046/j.0022-202x.2001.01366.x
190. Griffiths CE, Boffa MJ, Gallatin WM, Martin S. Elevated levels of circulating intercellular adhesion molecule-3 (cICAM-3) in psoriasis. Acta Derm Venereol. 1996;76(1):2–5. doi:10.2340/000155557625
191. Liu S, Xu J, Wu J. The role of co-signaling molecules in psoriasis and their implications for targeted treatment. Front Pharmacol. 2021;12. doi:10.3389/fphar.2021.717042
192. Li J, Ren F, Yuan H, Yan W. Knockdown of ANXA3 regulates NF-κB/STAT3 pathway to alleviate inflammation and hyperproliferation in psoriasis models. Allergol Immunopathol. 2025;53(2):32–41. doi:10.15586/aei.v53i2.1260
193. Gilhar A, Yaniv R, Assy B, Serafimovich S, Ullmann Y, Kalish RS. Fas Pulls the Trigger on Psoriasis. Am J Pathol. 2006;168(1):170–175. doi:10.2353/ajpath.2006.041354
194. Ryu S, Broussard L, Youn C, et al. Therapeutic effects of synthetic antimicrobial peptides, TRAIL and NRP1 blocking peptides in psoriatic keratinocytes. Chonnam Med J. 2019;55(2):75–85. doi:10.4068/cmj.2019.55.2.75
195. Caldarola G, Carbone A, Arena V, et al. Tumour necrosis factor-related apoptosis-inducing ligand (TRAIL): a possible pathogenic role in chronic plaque psoriasis. G Ital Dermatol E Venereol Organo Uff Soc Ital Dermatol E Sifilogr. 2016;151(1):17–24.
196. Gibellini L, Biasi SD, Bianchini E, et al. Anti-TNF-α drugs differently affect the TNFα-sTNFR system and monocyte subsets in patients with psoriasis. PLoS One. 2016;11(12):e0167757. doi:10.1371/journal.pone.0167757
197. Serwin AB, Mysliwiec H, Hukalowicz K, Porebski P, Borawska M, Chodynicka B. Soluble tumor necrosis factor-alpha receptor type 1 during selenium supplementation in psoriasis patients. Nutr Burbank Los Angel Cty Calif. 2003;19(10):847–850. doi:10.1016/s0899-9007(03)00165-5
198. Li X, Li C, Zhang W, Wang Y, Qian P, Huang H. Inflammation and aging: signaling pathways and intervention therapies. Signal Transduct Target Ther. 2023;8(1):239. doi:10.1038/s41392-023-01502-8
199. Weyh C, Krüger K, Strasser B. Physical activity and diet shape the immune system during aging. Nutrients. 2020;12(3):622. doi:10.3390/nu12030622
200. Novais EJ, Tran VA, Johnston SN, et al. Long-term treatment with senolytic drugs Dasatinib and Quercetin ameliorates age-dependent intervertebral disc degeneration in mice. Nat Commun. 2021;12(1):5213. doi:10.1038/s41467-021-25453-2
201. Millar CL, Iloputaife I, Baldyga K, et al. A pilot study of senolytics to improve cognition and mobility in older adults at risk for Alzheimer’s disease. eBioMedicine. 2025;113:105612. doi:10.1016/j.ebiom.2025.105612
202. Hickson LJ, Langhi Prata LGP, Bobart SA, et al. Senolytics decrease senescent cells in humans: preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine. 2019;47:446–456. doi:10.1016/j.ebiom.2019.08.069
203. Takaya K, Kishi K. Combined dasatinib and quercetin treatment contributes to skin rejuvenation through selective elimination of senescent cells in vitro and in vivo. Biogerontology. 2024;25(4):691–704. doi:10.1007/s10522-024-10103-z
204. Nogueira-Recalde U, Lorenzo-Gómez I, Blanco FJ, et al. Fibrates as drugs with senolytic and autophagic activity for osteoarthritis therapy. EBioMedicine. 2019;45:588–605. doi:10.1016/j.ebiom.2019.06.049
205. Park A, Heo TH. IL-17A-targeting fenofibrate attenuates inflammation in psoriasis by inducing autophagy. Life Sci. 2023;326:121755. doi:10.1016/j.lfs.2023.121755
206. Imamura T, Takata I, Ogasawara M, Matsutani Y, Yamamoto T, Asagami C. Clofibrate treatment of psoriasis with hypertriglycemia--clinical, histological and laboratory analysis. Nihon Hifuka Gakkai Zasshi Jpn J Dermatol. 1991;101(6):623–628.
207. Zia A, Farkhondeh T, Pourbagher-Shahri AM, Samarghandian S. The role of curcumin in aging and senescence: molecular mechanisms. Biomed Pharmacother Biomedecine Pharmacother. 2021;134:111119. doi:10.1016/j.biopha.2020.111119
208. Sun J, Zhao Y, Hu J. Curcumin inhibits imiquimod-induced psoriasis-like inflammation by inhibiting IL-1beta and IL-6 production in mice. PLoS One. 2013;8(6):e67078. doi:10.1371/journal.pone.0067078
209. Kolahdooz H, Khori V, Erfani-Moghadam V, et al. Niosomal curcumin suppresses IL17/IL23 immunopathogenic axis in skin lesions of psoriatic patients: a pilot randomized controlled trial. Life. 2023;13(5). doi:10.3390/life13051076
210. Mayo Clinic. MC230715 pilot study of the mechanistic feedback from CNS tumors with latent residual disease to guide individualized therapies. clinicaltrials.gov; 2025. Available from: https://clinicaltrials.gov/study/NCT07025226.
211. Zhang L, Pitcher LE, Prahalad V, Niedernhofer LJ, Robbins PD. Targeting cellular senescence with senotherapeutics: senolytics and senomorphics. FEBS J. 2023;290(5):1362–1383. doi:10.1111/febs.16350
212. Albert Einstein College of Medicine. Metformin in Longevity Study (MILES). clinicaltrials.gov; 2021. Available from: https://clinicaltrials.gov/study/NCT02432287.
213. Padki MM, Stambler I. Targeting Aging with Metformin (TAME). In: Encyclopedia of Gerontology and Population Aging. Cham: Springer;2021:4908–4910. doi:10.1007/978-3-030-22009-9_400
214. Mohammed I, Hollenberg MD, Ding H, Triggle CR. A critical review of the evidence that metformin is a putative anti-aging drug that enhances healthspan and extends lifespan. Front Endocrinol. 2021;12:718942. doi:10.3389/fendo.2021.718942
215. Singh S, Bhansali A. Randomized Placebo control study of metformin in psoriasis patients with metabolic syndrome (systemic treatment cohort). Indian J Endocrinol Metab. 2017;21(4):581–587. doi:10.4103/ijem.IJEM_46_17
216. Stanescu AMA, Simionescu AA, Florea M, Diaconu CC. Is metformin a possible beneficial treatment for psoriasis? A scoping review. J Pers Med. 2021;11(4):251. doi:10.3390/jpm11040251
217. Tam HTX, Thuy LND, Vinh NM, Anh TN, Van BT. The combined use of metformin and methotrexate in psoriasis patients with metabolic syndrome. Dermatol Res Pract. 2022;2022:9838867. doi:10.1155/2022/9838867
218. Saunders RN, Metcalfe MS, Nicholson ML. Rapamycin in transplantation: a review of the evidence. Kidney Int. 2001;59(1):3–16. doi:10.1046/j.1523-1755.2001.00460.x
219. Gao N, Zhang T, Ji J, Xu KF, Tian X. The efficacy and adverse events of mTOR inhibitors in lymphangioleiomyomatosis: systematic review and meta-analysis. Orphanet J Rare Dis. 2018;13(1):134. doi:10.1186/s13023-018-0874-7
220. Zhao P, Sui BD, Liu N, et al. Anti-aging pharmacology in cutaneous wound healing: effects of metformin, resveratrol, and rapamycin by local application. Aging Cell. 2017;16(5):1083–1093. doi:10.1111/acel.12635
221. Bürger C, Shirsath N, Lang V, et al. Blocking mTOR signalling with rapamycin ameliorates imiquimod-induced psoriasis in mice. Acta Derm Venereol. 2017;97(9):1087–1094. doi:10.2340/00015555-2724
222. Ormerod AD, A. A SS, Copeland P, Omar G, Winfield A. Treatment of psoriasis with topical sirolimus: preclinical development and a randomized, double-blind trial. Br J Dermatol. 2005;152(4):758–764. doi:10.1111/j.1365-2133.2005.06438.x
223. Buerger C. Epidermal mTORC1 signaling contributes to the pathogenesis of psoriasis and could serve as a therapeutic target. Front Immunol. 2018;9:2786. doi:10.3389/fimmu.2018.02786
224. EMA. RINVOQ summary of product characteristics. Available from: https://www.ema.europa.eu/en/medicines/human/EPAR/rinvoq.
225. Gargiulo L, Ibba L, Pavia G, et al. Upadacitinib for the treatment of concomitant psoriasis and atopic dermatitis: a case series. J Dermatol Treat. 2023;34(1):2183729. doi:10.1080/09546634.2023.2183729
226. Tsunoda M, Adachi T, Nakajima Y, Yatomi Y, Shimizu T, Nakasute K. Successful treatment of the dupilumab-induced psoriatic dermatitis/arthritis and atopic dermatitis with a JAK inhibitor: a case report and literature review. J Allergy Clin Immunol Glob. 2024;3(4):100323. doi:10.1016/j.jacig.2024.100323
227. Minghini M, Schettini N, Pacetti L, Bocchi C, Borghi A. Psoriasiform eruption induced by dupilumab successfully treated with upadacitinib. Dermatol Rep. 2025. doi:10.4081/dr.2025.10588
228. McInnes IB, Kato K, Magrey M, et al. Efficacy and safety of upadacitinib in patients with psoriatic arthritis: 2-year results from the phase 3 SELECT-PsA 1 study. Rheumatol Ther. 2023;10(1):275–292. doi:10.1007/s40744-022-00499-w
229. Strbo N, Younis S, Frasca D, Paganelli A. Skin aging and immunosenescence. Explor Immunol. 2025;5:1003218. doi:10.37349/ei.2025.1003218
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.