Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 15
Inducible Nitric Oxide Synthase iNOS-954-G>C and Ex16+14-C>T Gene Polymorphisms and Susceptibility to Vitiligo in the Saudi Population
Authors Al-Harthi F, Huraib GB, Mustafa M ![]() , Al-Qubaisy Y, Al-Nomair N, Abdurrahman N, Al-Asmari A
, Al-Qubaisy Y, Al-Nomair N, Abdurrahman N, Al-Asmari A
Received 2 December 2021
Accepted for publication 5 May 2022
Published 13 June 2022 Volume 2022:15 Pages 603—612
DOI https://doi.org/10.2147/PGPM.S344415
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Fahad Al-Harthi,1 Ghaleb Bin Huraib,2 Md Mustafa,2 Yasser Al-Qubaisy,1 Naif Al-Nomair,1 Nour Abdurrahman,1 Abdulrahman Al-Asmari2
1Deparment of Dermatology, Prince Sultan Military Medical City, Riyadh, Saudi Arabia; 2Scientific Research Center, Health service directorate, Riyadh, Saudi Arabia
Correspondence: Abdulrahman Al-Asmari, Scientific Research Center, Health service directorate, P.O. Box 8524, Riyadh, 11492, Saudi Arabia, Email [email protected]
Objective: Vitiligo is an acquired pigmentary skin disorder with regional disappearance of melanocytes. Multigenic inheritance has been proposed in the pathogenesis of vitiligo. The present study aimed to investigate the possible association of inducible nitric oxide synthase polymorphisms iNOS-954-G/C (rs1800482 G>C) and iNOS-Ex16+14-C/T (rs2297518 C>T) with vitiligo in the Saudi population, if any.
Methods: We included 120 vitiligo cases and an equal number of age matched healthy controls. Polymerase chain reaction with restriction fragment length polymorphism method was used for the analysis of genetic polymorphisms.
Results: The heterozygous (GC), (GC + CC) combined genotype and variant allele; C allele of rs1800482 G>C were associated significantly (p < 0.005, after Bonferroni correction) with increased risk of vitiligo (OR = 3.46, 95% CI = 1.99– 6.01, p = 0.001), (OR = 3.30, 95% CI= 1.93– 5.65, p = 0.001) and (OR = 1.94, 95% CI = 1.31– 2.87, p = 0.001) respectively. When GC genotype of rs1800482 G>C was co-inherited with common genotype (CC) and heterozygous genotype (CT) of rs2297518 C>T, the risk of vitiligo was significantly increased ((OR = 4.51, 95% CI = 2.18– 9.33, p = 0.001) and (OR = 3.60, 95% CI = 1.61– 8.01, p = 0.001)) respectively. None of the rs1800482 G>C and rs2297518 C>T genotypes and alleles have been associated with non-segmental vitiligo in terms of gender, age of onset, and types of vitiligo.
Conclusion: The heterozygous (GC), (GC+CC) combined genotype and variants allele; C allele of rs1800482 G>C, may cause overproduction of NO, which has been linked to melanocyte loss by increasing oxidative stress and decreasing melanocyte adhesion to the extracellular matrix components, and thus could be an associative risk factor for vitiligo.
Keywords: vitiligo, non-segmental, genetic polymorphism, rs1800482 G>C, rs2297518 C>T, nitric oxide, nitric oxide synthase
Introduction
Vitiligo is an acquired multifactorial progressive depigmentary disorder characterized by the appearance of defined white macules resulting from the progressive loss of functional melanocytes from the skin and mucous. It affects 0.1–2% of the world population without sex predilection.1,2 Although the precise etiology of vitiligo is still illusive the role of autoimmunity, genetics, oxidative stress, auto cytotoxic, neurogenic, viral, apoptotic, cell adhesion disorder, etc. have all been reported.3–5 The most widely accepted theory of its development is multifactorial according to which genetic changes predispose risk in the presence of favorable environmental conditions i.e. gene-environment interaction may be involved in melanocyte destruction.1
The high occurrence rate of vitiligo within the families suggested that genetic risk factors may be one of the predominant causes which is associated with this skin abnormality.6,7 Genome wide association studies confirmed that more than 40 robust genetic susceptible loci have been associated with vitiligo. Beside this, many more susceptible loci with unknown functions have also been identified in the pathogenesis of vitiligo.1,8,9 The inducible nitric oxide synthesize (iNOS) gene of 48Kb is located on the chromosome 17q11.2-q12, which encodes a protein of 131 kDa.10 Several SNPs in iNOS have been revealed, including the G>C in iNOS-954 (rs1800482) in the promoter region, which has been linked to increased iNOS expression, and the C>T substitution in iNOS exon 16 (rs2297518), which induces Ser608Leu amino acid substitution resulting in higher nitric oxide (NO) production11,12 Oxidative stress mediated toxicity in melanocyte destruction is another widely accepted hypothesis for the development of vitiligo.
The excessive generation of NO may cause increased oxidative stress and imbalance of the antioxidant system in melanocytes resulting in an accumulation of huge toxic intermediate of melanin, breaking the melanocyte redox homeostasis and reducing melanogenesis, all of which contribute to the onset of vitiligo.13,14 Polymorphism in oxidative stress related genes like inducible nitric oxide synthase (iNOS),15 glutathioneS-transferase,16 NF-E2-related factor2,17 catechol-O-methyltransferase, catalase,18 and angiotensinogen converting enzymes19 have previously been associated with a higher risk of vitiligo. The iNOS promoter polymorphisms have shown strong association with vitiligo in Chinese and Egyptian populations.15,20
Many factors, including lipopolysaccharides (LPO), cytokines, interleukins, and others, can influence the expression of iNOS in normal human melanocytes, affecting NO production.13,21 Previous studies from our laboratory have shown the association of inflammatory genes SNPs with vitiligo.22,23 So far, no studies have reported a genetic association of these genes with vitiligo in a Saudi population. Considering the importance of iNOS in NO production and its subsequent role in the pathogenesis of vitiligo, the present study intends to examine the potential association of functional polymorphisms in the iNOS rs1800482 G>C and rs2297518 C>T with vitiligo in the Saudi population.
Materials and Methods
In the present study, a total of a hundred and twenty (n = 120) vitiligo cases and an equal number of controls were recruited from the Dermatology Clinic of the Prince Sultan Military Medical City (PSMMC), Riyadh, Saudi Arabia. Patients who routinely visited the outpatient clinic without any known health risk were recruited as controls. All the subjects were age and sex matched and biologically unrelated Saudis. The ethical clearance was obtained from the institutional ethical clearance committee, PSMMC, Riyadh to conduct this research work. We collected required data of vitiligo in accordance with principles of the Declaration of Helsinki. The informed written consent was obtained from all the participants prior to the recruitment. The patient and control samples were collected after following strict exclusion and inclusion criteria. Only those vitiligo patients were included in this study who were not subject to any therapy in the last six months and had no history of autoimmune disorders. We recruited gender and age matched controls and excluded those with clinical evidence or a family history of a first- or second-degree relative with vitiligo or any autoimmune disorders to minimize genetic heterogeneity. All vitiligo cases recruited in the present study are non-segmental. The patient group consisted of n = 61 males and n = 59 females with age at diagnosis ranging from 6 to 79 years (mean ± SD = 27.04 ± 12.32). Venous blood (3 mL) was drawn in an EDTA coated vial from each subject, brought to the laboratory in ice boxes and stored at −80 °C before extraction of genomic DNA.
Genotyping Analysis
Genomic DNA for genotyping was isolated from whole blood by using commercially available QIAamp DNA mini kit (Qiagen, Venlo, Limburg, the Netherlands) as per the manufacturer's protocol. The quality of DNA was checked on agarose gel and quantitation was done by using Nano Drop-2000 at 260/280 nm (ThermoFisherScientific Inc, Waltham, MA, USA). The gradient PCR (Ependorf, Hamburg, Germany) was used to amplify the fragments that contained the selected iNOS polymorphic sites. The specific set of primers were used for the rs1800482 G>C amplification was 5’-CATATGTATGGGAATACTGTATTTCAG-3,5-TCTGAACTAGTCACTTGAGG-3’. DNA amplification was done in a 25 µL reaction mixture consisting of 50 ng of genomic DNA, 10 pM of each primer, 200 µM of dNTPs mixture, 1.5 mM MgCl2, 2.5 µL of 10x PCR buffer (10x 500 mM KCl, 100 mM TrisHCl, pH 9.0) and 1 U Taq DNA polymerase (New Englad Biolab Ipswich, MA, USA). Genomic DNA was amplified for 35 cycles. Initial denaturation was applied for 3 minutes at 95 °C. Each cycle consisted of: 95 °C for 30 sec, 59 °C for 30 sec, and 72 °C for 45 sec.
For rs2297518 C>T amplification, the following set of primers was used 5’-CATATGTAAACCAACTTCCGTG-3’,5’-GGCAGGGCTAGGAGTAGGAC-3’. The thermal cycler program was similar except the annealing temperature was kept at 60 °C. Both the PCR products obtained were separated by electrophoresis on 2% agarose gel in TBE buffer, visualized by ethidium bromide fluorescence. The PCR product of rs2297518 C>T was purified by gel band purification kit as per the manufacturer's protocol (GE Healthcare, Buckinghamshire, HP7 9NA, UK). Genotyping has been performed by previously standardized PCR-RFLP method by Shen et al.12 The PCR product of rs1800482 G>C was digested with BsaI enzyme and iNOS- rs2297518 C>T by MluCI (New England Biolab, Ipswich, MA, USA) for three hours at 37 °C. The restriction fragment was separated by 2.5% and 3% agarose gel electrophoresis to identify the genotypes of rs1800482 G>C and rs2297518 C>T respectively.
Statistical Analysis
The statistical analysis was carried out by using SPSS software, windows version 21.0 (Chicago, IL, USA). The age of patients and onset age (years) of disease were presented as mean ± SD, gender, types, stage, family history of vitiligo and autoimmune immune diseases are presented in frequency and percentage values. We computed odds ratio (OR) and 95% confidence interval (CI) for iNOS polymorphisms by univariable logistic regression taking the group as dependent and iNOS polymorphic variants as independent covariates. Bonferroni correction was applied to reduce the type-1 error due to multiple comparison testing. The corrected significant p value is <0.005.
Results
In our study, mean age of cases (27.04 ± 12.32 years) are matching with the mean age of controls (26.84 ± 4.89) years. The age of onset of vitiligo was (22.51 ± 15.40) years. The mean value of duration of disease was 6.2 (1–19) years. The proportion of male and female patients with vitiligo was almost the same. The family history of vitiligo and other autoimmune diseases were observed in 15% and 6.67% cases, respectively, the frequency of generalized vitiligo and focal were 33.34% (Table 1). The distribution of the genotypic frequencies of iNOS-954 were in Hardy-Weinberg equilibrium (HWE) in controls (p = 0.901) which implies similar genetic background to the Mendelian population. However, iNOS-954 G/C genotype (χ2 = 14.84, p < 0.001) in cases was not compatible with HWE (Table 2). In cases, the frequency of rs1800482 G>C common (GG), heterozygous (GC) and homozygous variants (CC) genotypes were 29.16%, 64.16% and 6.68%, while in controls 56.67%, 37.5% and 5.83% respectively. Calculated risk estimation revealed the GC genotype of rs1800482 was found to have a threefold risk of vitiligo in Saudis. However, the frequency of CC genotype was higher in cases but did not reach the level of significance. When, GC genotype was combined with CC genotype of rs1800482 G>C imposes significant risk (OR = 3.30, 95% CI = 1.93–5.65, p = 0.001) of vitiligo in cases as compared to controls. The variants allele; C allele, was found to be significantly (OR = 1.94, 95% CI = 1.34–2.87, p = 0.001) higher in cases as compared to controls (Table 2). Similarly, when rs2297518 C>T genetic polymorphism were evaluated between patients and controls using the CC genotype as a reference group, we observed that CT and TT genotypes were greater in cases but did not meet the level of significance (p > 0.005). Further, no significant difference was observed between cases and controls for the variant allele; T allele of rs2297518 C>T (Table 2). When GC genotype of rs1800482 G>C was co-inherited with the CT genotype of rs2297518 C>T, the risk of vitiligo was significantly increased (OR = 5.07, 95% CI = 2.46–10.43, p =< 0.001) in cases as compared to controls respectively. Similarly, when CC genotype of rs1800482 G>C was co-inherited with the CT genotype of rs2297518 C>T, the risk of vitiligo was also increased significantly (OR = 9.77, 95% CI = 1.02–93.6, p = 0.036) as shown in Table 3. None of the rs1800482 G>C and rs2297518 C>T genotypes have been associated with non-segmental vitiligo in terms of gender, age of onset, and types of vitiligo (Tables 4–6).
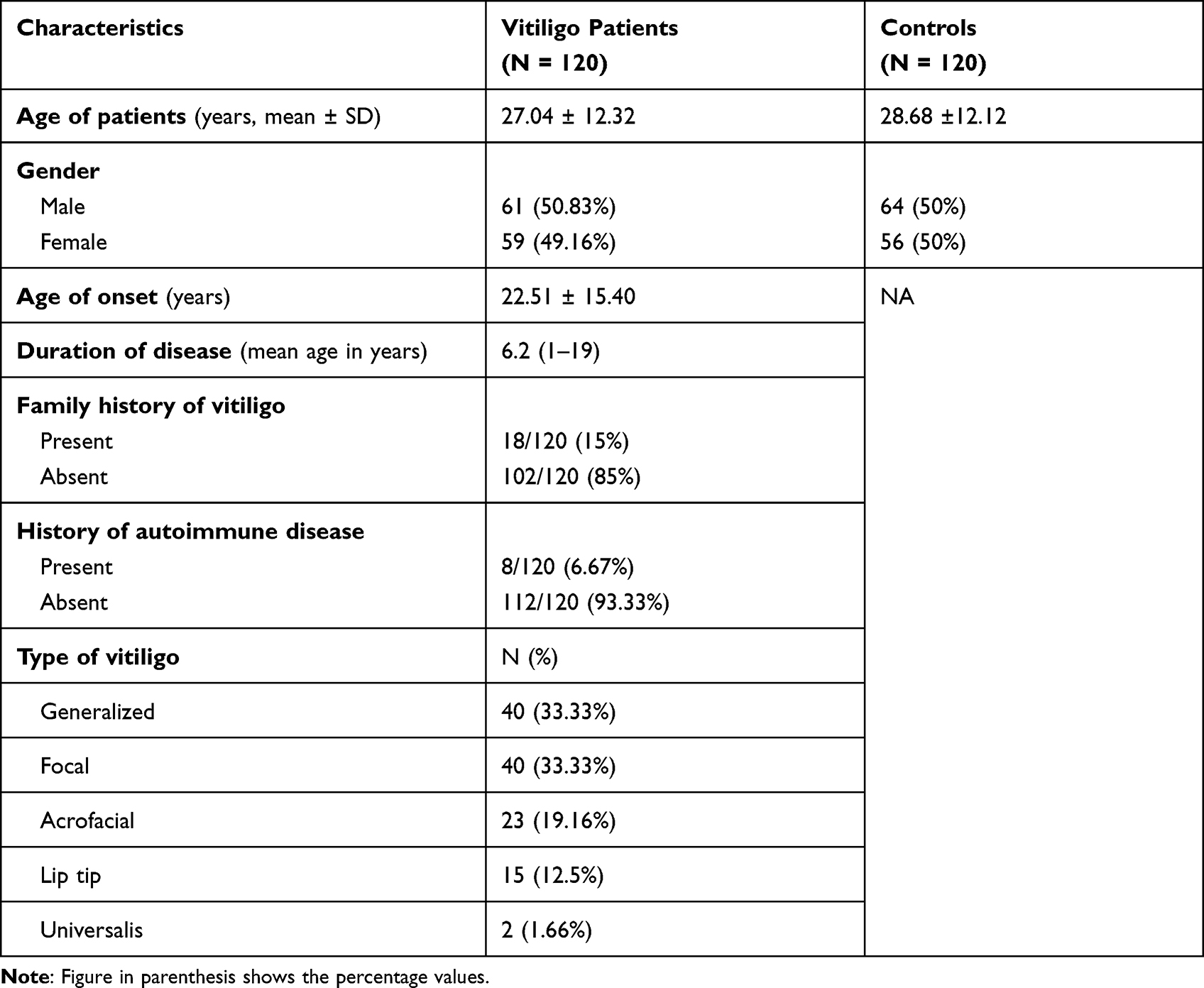 |
Table 1 Demographical Characteristics of Vitiligo Patients and Controls |
 |
Table 2 Genotypic Frequencies of rs1800482 G>C and rs2297518 C>T in Cases and Controls and Their Associations with Risk of Vitiligo |
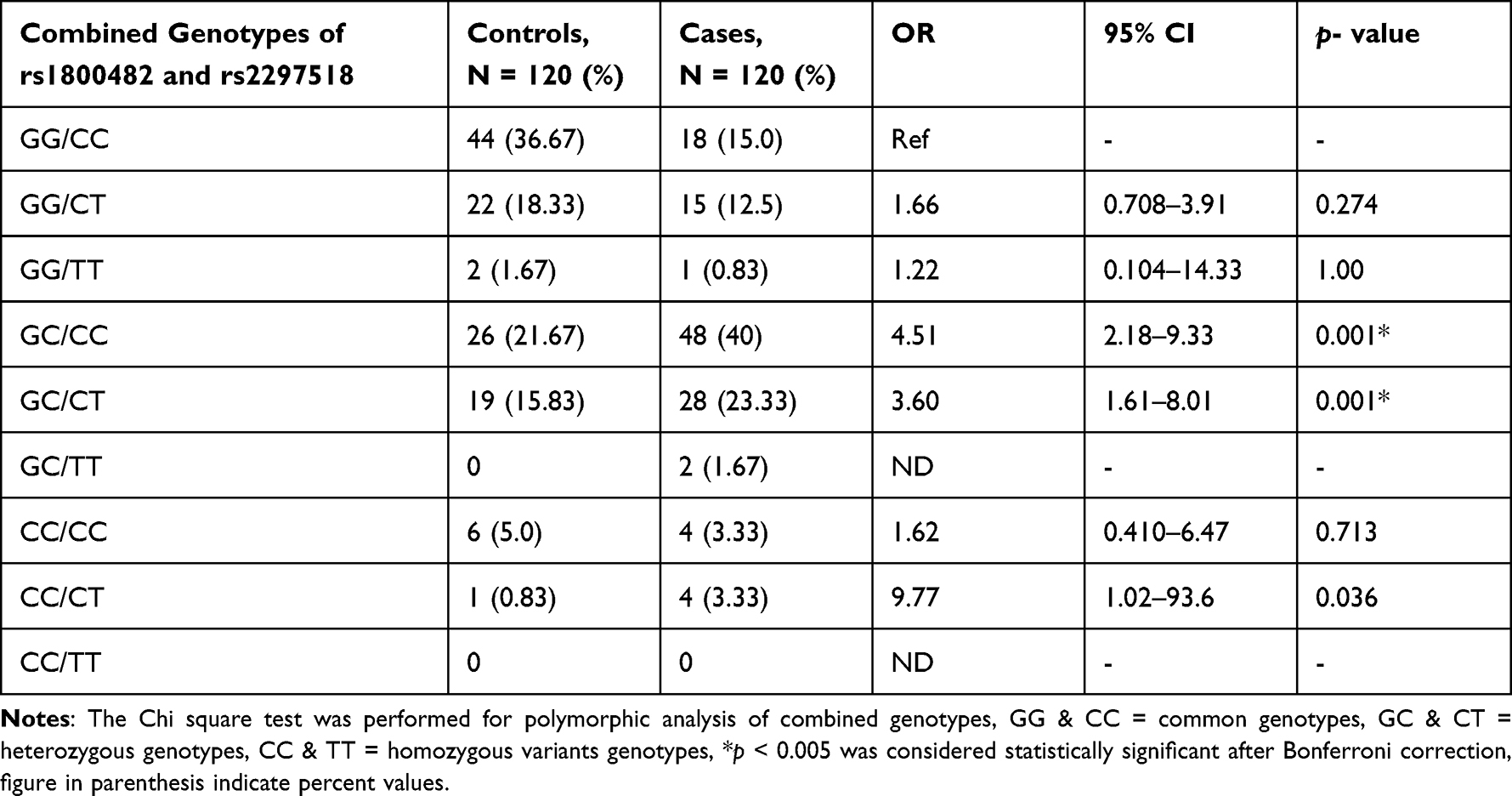 |
Table 3 Combined Genotypes Analysis of rs1800482 G>C and rs2297518 C>T in Cases and Controls |
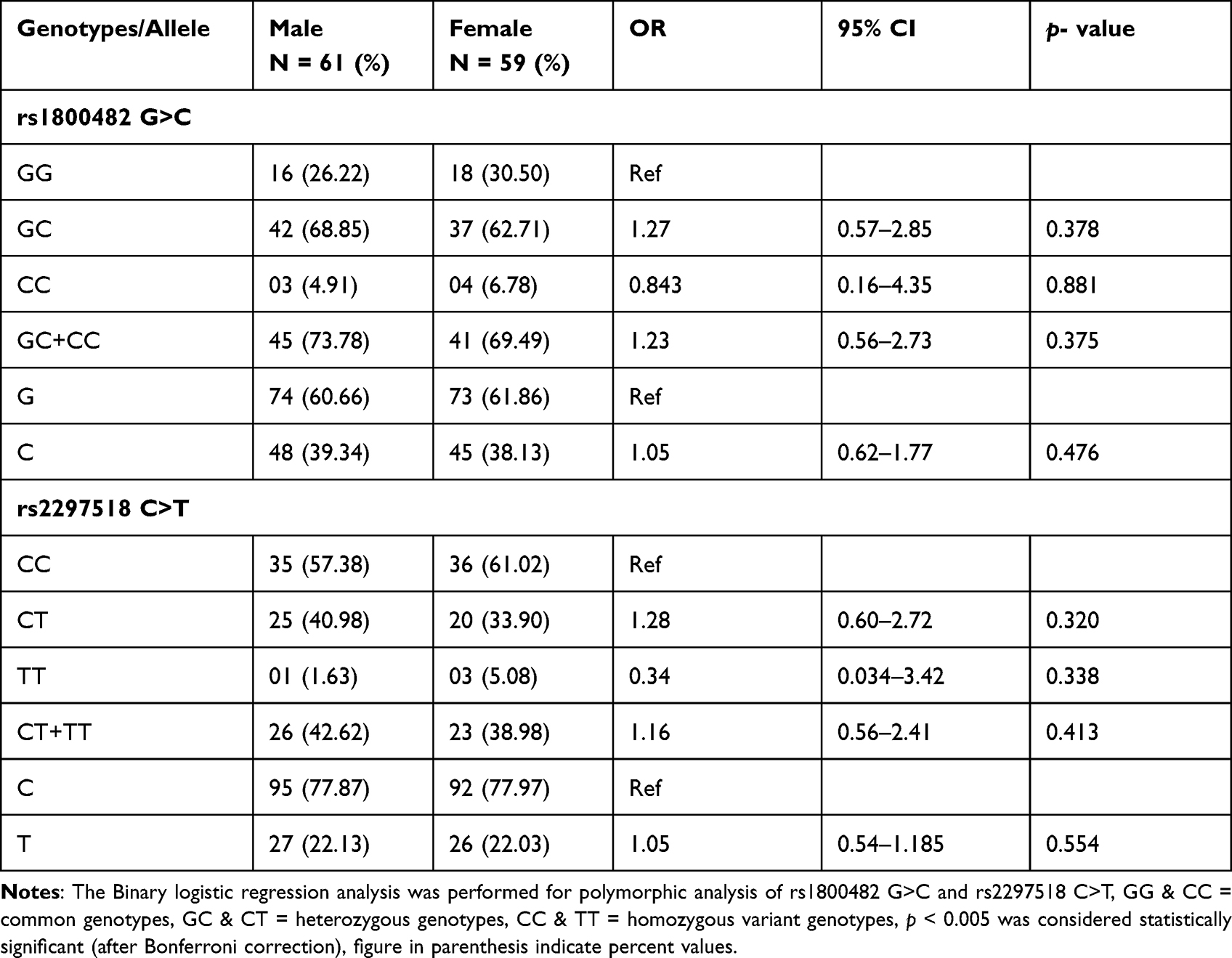 |
Table 4 Comparison of Genotypic Frequencies of rs1800482 G>C and rs2297518 C>T in Male and Female Cases of Vitiligo |
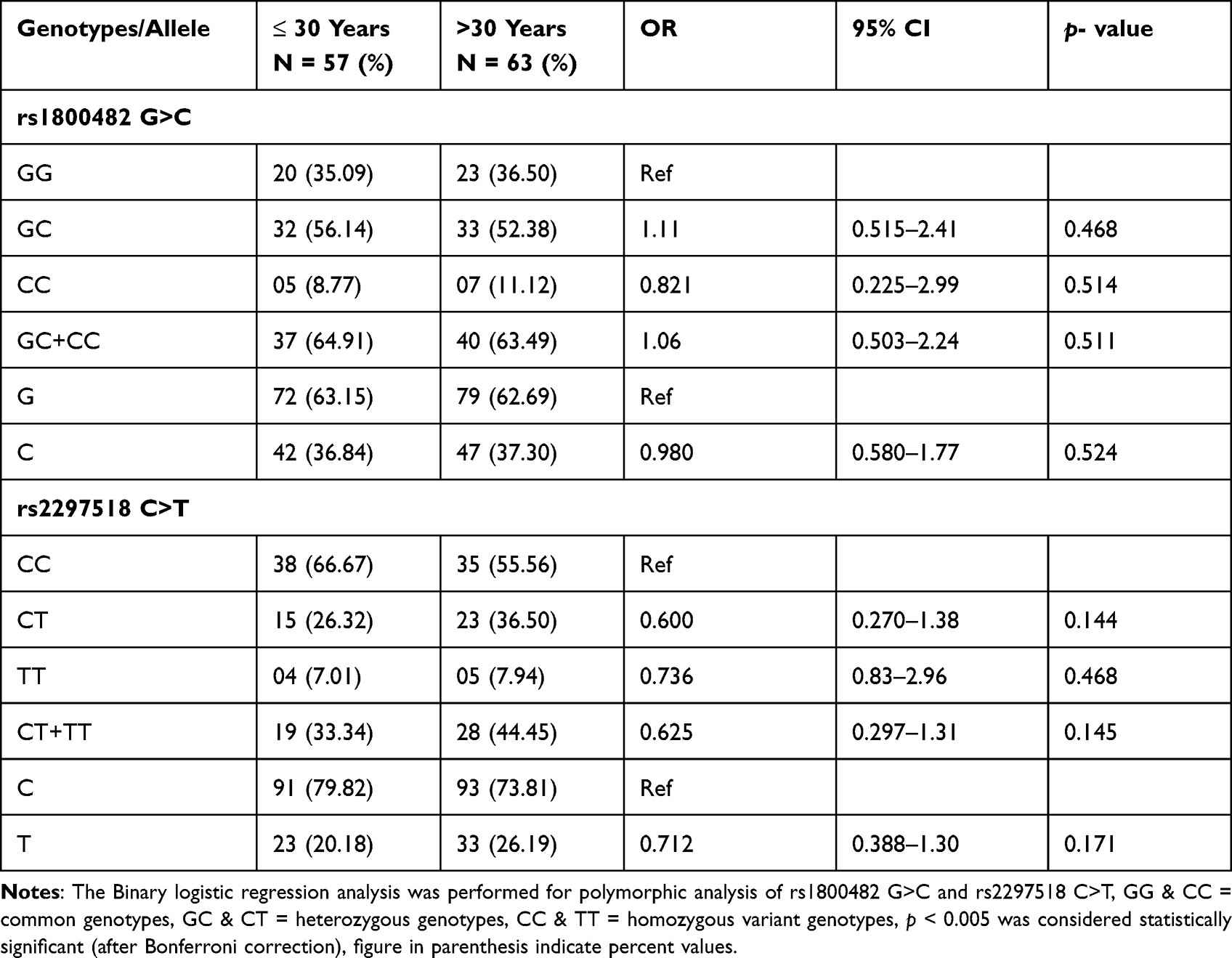 |
Table 5 Comparison of Genotypic Frequencies of rs1800482 G>C and rs2297518 C>T Between Two Groups of Onset Age in Vitiligo Patients |
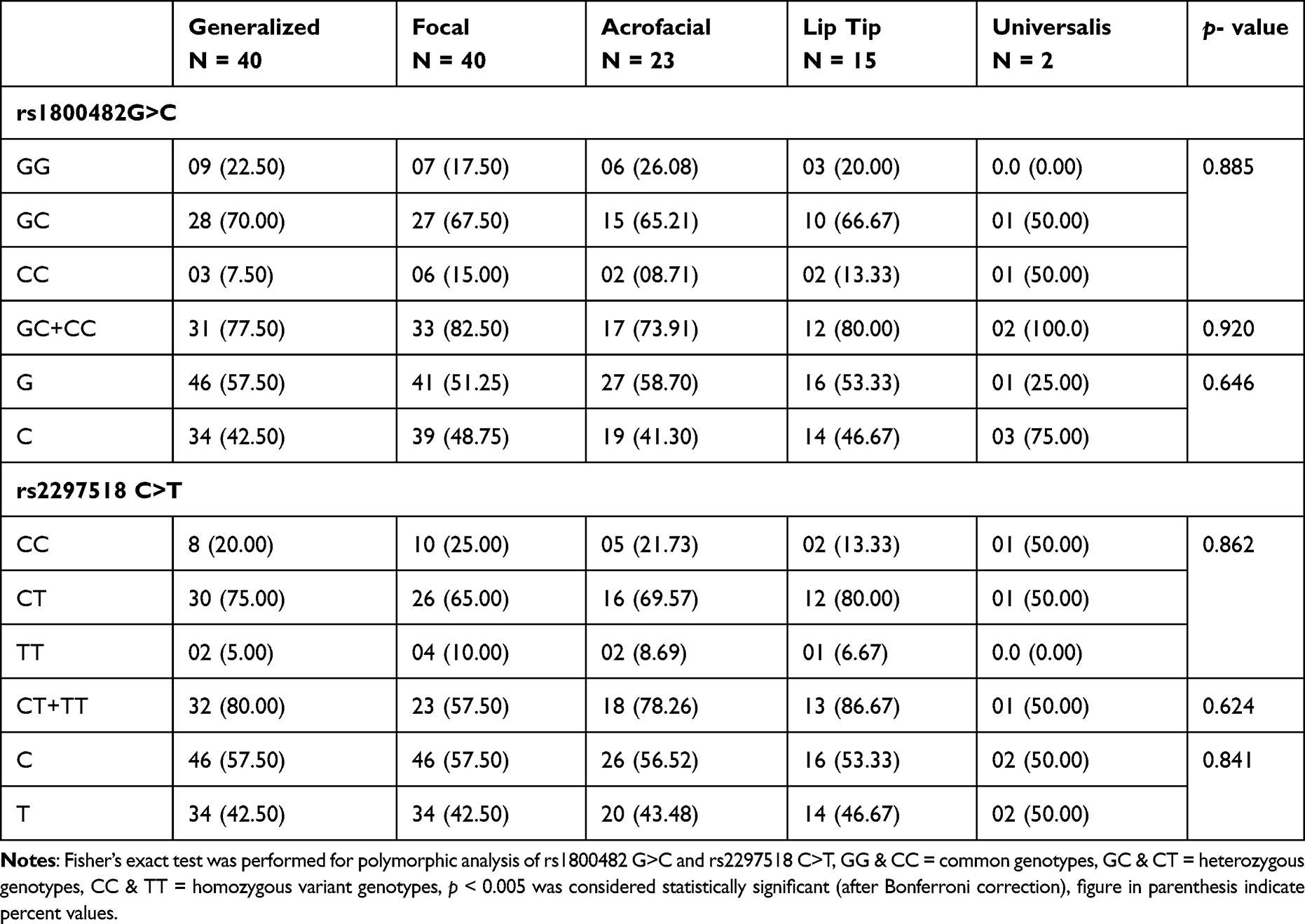 |
Table 6 Comparison of Genotypic Frequencies of rs1800482 G>C and rs2297518 C>T Among Different Types of Vitiligo Type of Vitiligo |
Discussion
The expression of the iNOS gene is regulated at the level of transcription. The presence of variation in the regulatory promoter region affects the gene product in the cell, while variation in the coding region bring structural changes in proteins and its activity. This change in gene product level and protein molecule structure could be the source of genetic predisposition in many human diseases.24,25 The promoter polymorphisms rs1800482 G>C have been shown to modify iNOS transcription and mRNA levels26,27 whereas the rs2297518 C>T exon 16 polymorphism, which causes an amino acid substitution from serine to leucine, may be of functional interest in vitiligo.28
In our study, when GG genotype of rs1800482 was considered as a reference group, the frequency of GC genotype was found significantly higher in cases as compared to controls. In addition, the combined (GC+CC) genotypes and variant allele; C allele of rs1800482 were also significantly associated with increased risk of vitiligo (Table 2). Our findings demonstrate that the risk factors aggregation may amplify the risk of vitiligo in the Saudi population. Similar to our study, Zayed et al,20 reported that GC genotype of rs1800482 may increase the risk of vitiligo by 3.35 times while combined (GC+CC) genotype of iNOS-954 may increase it by 3.02 times, when compared to controls in Egyptians. They also reported variants allele; C allele was significantly higher and a 2.21 fold risk of vitiligo. In another study, Zhang et al,15 did not find any significant association of GC genotype of rs1800482 with cases in the Chinese population while they have reported combined (GC+CC) genotype may increase by 1.36 times the risk of vitiligo. In their study, C allele was also reported to be significantly associated with the cases. The difference in genotypic frequency of rs1800482 in our study with Chinese and similarity with Egyptians populations, indicate that the genotypic distribution of the rs1800482 G>C polymorphism varies by ethnicity.
When we considered the CC genotype of rs2297518 C>T as a reference group, we did not find any significant difference for individuals with CT and TT or combined (CC+TT) genotypes between cases and controls (Table 2). Our observations are in accordance with the previous genotypic frequencies of rs2297518 reported among Chinese, Spanish and Japanese populations.12,15,27 In contrast to our findings, Zayed et al,20 found significant associations of CT and combined (CT+TT) genotypes of rs2297518 with vitiligo in Egyptian patients. Further, they have also reported a significantly high frequency of allelic variant; T allele in cases as compared to controls.
In the present study, when rs1800482 GC genotype was co-inherited with CC and CT genotypes of rs2297518, the risk of vitiligo was further increased significantly by 4.51 and 3.60 times in cases as compared to controls, respectively. When the CC genotype of rs1800482 was co-inherited with the CT genotype of rs2297518, the risk increased 9.77 times however, the data became non-significant after Bonferroni correction (p < 0.005), as shown in Table 3. Similar findings were reported by Zayed et al20 while Zhang et al15 identified a substantial difference only when the GC genotype of rs1800482 was co-inherited with the CC genotype of rs2297518. We found no significant association between non-segmental vitiligo and any genotype of rs1800482 G>C and rs2297518 C>T in terms of gender, onset age, and types of vitiligo. Zhang et al15 noticed a similar outcome in the Chinese population. In the Egyptian population, however, rs1800482 G>C were found to be significantly associated with female patients when compared to its counterpart in controls.20 Our data showed that there are no haplotypes or LD, and one of the markers did not follow the Hardy-Weinberg formula. The reported spacing between markers is approximately 31.9 kb.
The role of NO in the inhibition of cell proliferation, differentiation and death, and thus possible contribution in the pathogenesis of autoimmune disorders has been reported previously. In addition, as a regulatory mediator, NO has been linked to the onset of autoimmune disease.29 A study was carried out in the south Indian population to evaluate the association of the iNOS promoter polymorphism with rheumatoid arthritis (RA). They found that iNOS-277A/G and −1026 G/T promoter’s polymorphism may increase the risk of RA.30 Also, Sato et al, 2016 suggested the importance of the iNOS genotype in the clinical application of FeNO for asthmatic patients.31
The association of rs1800482 G>C promoter gene polymorphisms with vitiligo in our study may have a greater significance because variations in the regulatory promoter region of a gene can directly affect the level of gene product, subsequently it may play a causative role in the development of diseases. Several hypotheses have been proposed to understand the pathophysiology of vitiligo, however the actual mechanism underlying the onset of vitiligo remains unknown. The rs1800482 G>C promoter polymorphism is linked with the increased iNOS expression and also the C/T substitution in exon 16 causing an amino acid substitution (Ser608Leu), resulting in increased production of NO.32,33 Furthermore, C allele of iNOS may have a higher affinity to bind with DNA binding protein as compared to G allele therefore, the transcriptional activity and the mRNA expression will be higher for C allele than G allele which ultimately increases the iNOS activity and NO production.15 The increased iNOS activity triggers overproduction of NO and O−2 which leads to accumulation of hydrogen peroxide (H2O2) and exposure of melanocytes to H2O2 results in their destruction and depigmentation. Tetrahydrobiopterin, a cofactor, is essential for the production of NO by all nitric oxide synthase enzymes. Defective recycling of 6-tetrahydrobiopterin increases H2O2 production and decreases catalase levels.34 In vitiligo, an increase in iNOS activity caused by the overexpression of tetrahydrobiopterin or LPS/cytokines resulted in a large amount of NO production. Nitric oxide has been linked to melanocyte loss by increasing oxidative stress and decreasing melanocyte adhesion to the extracellular matrix component, which could be a risk factor for vitiligo.35,36 The identification of genetic susceptibility variants in vitiligo will lead to a better understanding of the pathophysiology of the disease and also enable us to screen the high risk patients at the early stages of the disease, development of new diagnostic markers, and treatment strategies.
Conclusion
Our findings supported the genetic basis of vitiligo. The present study clearly demonstrates the association of rs1800482 G>C polymorphism with vitiligo and it could be used as a genetic marker for screening vitiligo patients among Saudis. The functional polymorphisms could be an activator of gene expression therefore, our findings may be useful in exploring the molecular mechanism underlying the onset of vitiligo. Further study with a larger sample size, estimation of iNOS activity, oxidative stress parameters, and gene environment interaction will strengthen the current findings.
Ethical Approval
Institutional ethical approval has been taken to conduct this research study.
Acknowledgment
The authors are thankful to Mohammad Asmari and S. Sadaf Rizvi for their help with laboratory work.
Funding
No external grants or other support were received for the conduction of this study.
Disclosure
The authors declare that they have no conflict of interests in relation to this work.
References
1. Spritz RA. Recent progress in the genetics of generalized vitiligo. J Genet Genomics. 2011;38(7):271–278. doi:10.1016/j.jgg.2011.05.005
2. Strömberg S, Björklund MG, Asplund A, et al. Transcriptional profiling of melanocytes from patients with vitiligo vulgaris. Pigment Cell Melanoma Res. 2008;21:162–171. doi:10.1111/j.1755-148X.2007.00429.x
3. Taïeb A, Picardo M. Clinical practice. Vitiligo. N Engl J Med. 2009;360(2):160–169. doi:10.1056/NEJMcp0804388
4. Gavalas NG, Gottumukkala RV, Gawkrodger DJ, Watson PF, Weetman AP, Kemp EH. Mapping of melanin-concentrating hormone receptor 1 B cell epitopes predicts two major binding sites for vitiligo patient autoantibodies. Exp Dermatol. 2009;18:454–463. doi:10.1111/j.1600-0625.2008.00813.x
5. Xie H, Zhou F, Liu L, et al. Vitiligo: how do oxidative stress-induced autoantigens trigger autoimmunity? J Dermatol Sci. 2016;81:3–9. doi:10.1016/j.jdermsci.2015.09.003
6. Alkhateeb A, Fain PR, Thody A, et al. Epidemiology of vitiligo and associated autoimmune diseases in Caucasian probands and their families. Pigment Cell Res. 2003;16(3):208–214. doi:10.1034/j.1600-0749.2003.00032.x
7. Sun X, Xu A, Wei X, et al. Genetic epidemiology of vitiligo: a study of 815 probands and their families from south China. Int J Dermatol. 2006;45(10):1176–1181. doi:10.1111/j.1365-4632.2006.02907.x
8. Birlea SA, Gowan K, Fain PR, et al. Genome-wide association study of generalized vitiligo in an isolated European founder population identifies SMOC2, in close proximity to IDDM8. J Invest Dermatol. 2010;130(3):798–803. doi:10.1038/jid.2009.347
9. Cheong KA, Kim NH, Noh M, et al. Three new single nucleotide polymorphisms identified by a genome-wide association study in Korean patients with vitiligo. J Korean Med Sci. 2013;28(5):775–779. doi:10.3346/jkms.2013.28.5.775
10. Xu W, Charles IG, Liu L, et al. Molecular cloning and structural organization of the human inducible nitric oxide synthase gene (NOS2). Biochem Biophys Res Commun. 1996;219(3):784–788. doi:10.1006/bbrc.1996.0311
11. Kun JF, Mordmüller B, Lell B, et al. Polymorphism in promoter region of inducible nitric oxide synthase gene and protection against malaria. Lancet. 1998;351(9098):265–266. doi:10.1016/S0140-6736(05)78273-8
12. Shen J, Wang RT, Wang LW, et al. A novel genetic polymorphism of inducible nitric oxide synthase is associated with an increased risk of gastric cancer. World J Gastroenterol. 2004;10:3278–3283. doi:10.3748/wjg.v10.i22.3278
13. Custurone P, Di Bartolomeo L, Irrera N, et al. Role of cytokines in vitiligo: pathogenesis and possible targets for old and new treatments. Int J Mol Sci. 2021;22(21):11429. doi:10.3390/ijms222111429
14. Xuan Y, Yang Y, Xiang L, Zhang C, Rauf A. The role of oxidative stress in the pathogenesis of vitiligo: a culprit for melanocyte death. Oxid Med Cell Longev. 2022;2022:8498472. doi:10.1155/2022/8498472
15. Zhang Y, Li C, Li K, et al. Analysis of inducible nitric oxide synthase gene polymorphisms in vitiligo in Han Chinese people. PLoS One. 2011;6:e27077. doi:10.1371/journal.pone.0027077
16. Lu L, Wu W, Tu Y, et al. Association of glutathione S-transferase M1/T1 polymorphisms with susceptibility to vitiligo. Gene. 2014;535(1):12–16. doi:10.1016/j.gene.2013.11.024
17. Song P, Li K, Liu L, et al. Genetic polymorphism of the Nrf2 promoter region is associated with vitiligo risk in Han Chinese populations. J Cell Mol Med. 2016;20(20):1840–1850. doi:10.1111/jcmm.12874
18. Mehaney DA, Darwish HA, Hegazy RA, et al. Analysis of oxidative stress status, catalase and catechol-O-methyltransferase polymorphisms in Egyptian vitiligo patients. PLoS One. 2014;9(6):e99286. doi:10.1371/journal.pone.0099286
19. Rashed L, Abdel Hay R, Mahmoud R, et al. Association of Angiotensin-Converting Enzyme (ACE) gene polymorphism with inflammation and cellular cytotoxicity in vitiligo patients. PLoS One. 2015;10(7):e0132915. doi:10.1371/journal.pone.0132915
20. Zayed AA, Khorshied MM, Hussein MF. Inducible nitric oxide synthase promoter polymorphism: a molecular susceptibility marker for vitiligo in Egyptians. Int J Dermatol. 2015;54(6):675–679. doi:10.1111/ijd.12716
21. Fecker LF, Eberle J, Orfanos CE, et al. Inducible nitric oxide synthase is expressed in normal human melanocytes but not in melanoma cells in response to tumor necrosis factor-alpha, interferon-gamma, and lipopolysaccharide. J Invest Dermatol. 2002;118:1019–1025. doi:10.1046/j.1523-1747.2002.01744.x
22. Al-Harthi F, Zouman A, Arfin M, et al. Tumor necrosis factor-α and -β genetic polymorphisms as a risk factor in Saudi patients with vitiligo. Genet Mol Res. 2013;12(3):2196–2204. doi:10.4238/2013.July.8.1
23. Abanmi A, Al Harthi F, Zouman A, et al. Association of Interleukin-10 gene promoter polymorphisms in Saudi patients with vitiligo. Dis Markers. 2008;24(1):51–57. doi:10.1155/2008/210609
24. Qidwai T, Jamal F. Inducible nitric oxide synthase (iNOS) gene polymorphism and disease prevalence. Scand J Immunol. 2010;72(5):375–387. doi:10.1111/j.1365-3083.2010.02458.x
25. Zoghbi HY, Beaudet AL. Epigenetics and human disease. Cold Spring Harb Perspect Biol. 2016;8(2):a019497. doi:10.1101/cshperspect.a019497
26. Levesque MC, Hobbs MR, Anstey NM, et al. Nitric oxide synthase type 2 promoter polymorphisms, nitric oxide production, and disease severity in Tanzanian children with malaria. J Infect Dis. 1999;180(6):1994–2002. doi:10.1086/315140
27. Kun JF, Mordmuller B, Perkins DJ, et al. Nitric oxide synthase 2(Lambarene) (G-954C), increased nitric oxide production, and protection against malaria. J Infect Dis. 2001;184:330–336. doi:10.1086/322037
28. Johannesen J, Pie A, Pociot F, et al. Linkage of the human inducible nitric oxide synthase gene to type 1 diabetes. J Clin Endocrinol Metab. 2001;86(6):2792–2796. doi:10.1210/jcem.86.6.7559
29. Kolb H, Kolb-Bachofen V. Nitric oxide in autoimmune disease: cytotoxic or regulatory mediator? Immunol Today. 1998;19(12):556–561. doi:10.1016/S0167-5699(98)01366-8
30. Negi VS, Mariaselvam CM, Misra DP, et al. Polymorphisms in the promoter region of iNOS predispose to rheumatoid arthritis in south Indian Tamils. Int J Immunogenet. 2017;44(3):114–121. doi:10.1111/iji.12315
31. Sato S, Wang X, Saito J, et al. Exhaled nitric oxide and inducible nitric oxide synthase gene polymorphism in Japanese asthmatics. Allergol Int. 2016;65(3):300–305. doi:10.1016/j.alit.2016.02.007
32. Rocha IM, Guillo LA. Lipopolysaccharide and cytokines induce nitric oxide synthase and produce nitric oxide in cultured normal human melanocytes. Arch Dermatol Res. 2001;293(5):245–248. doi:10.1007/s004030100211
33. Ivanova K, Le Poole IC, Gerzer R, et al. Effect of nitric oxide on the adhesion of human melanocytes to extracellular matrix components. J Pathol. 1997;183(4):469–476. doi:10.1002/(SICI)1096-9896(199712)183:4<469::AID-PATH931>3.0.CO;2-T
34. Hasse S, Gibbons NC, Rokos H, et al. Perturbed 6-tetrahydrobiopterin recycling via decreased dihydropteridine reductase in vitiligo: more evidence for H2O2 stress. J Invest Dermatol. 2004;122(2):307–313. doi:10.1046/j.0022-202X.2004.22230.x
35. Morelli JG, Norris DA. Influence of inflammatory mediators and cytokines on human melanocyte function. J Invest Dermatol. 1993;100(2 Suppl):191S–195S. doi:10.1038/jid.1993.75
36. Schallreuter KU, Wood JM, Pittelkow MR, et al. Regulation of melanin biosynthesis in the human epidermis by tetrahydrobiopterin. Science. 1994;263(5152):1444–1446. doi:10.1126/science.8128228
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.