Back to Journals » Psoriasis: Targets and Therapy » Volume 9
Increased dermal expression of chromatin-associated protein HMGB1 and concomitant T-cell expression of the DNA RAGE in patients with psoriasis vulgaris
Authors Strohbuecker L ![]() , Koenen H, van Rijssen E, van Cranenbroek B, Fasse E
, Koenen H, van Rijssen E, van Cranenbroek B, Fasse E ![]() , Joosten I, Körber A, Bergmann C
, Joosten I, Körber A, Bergmann C ![]()
Received 11 October 2018
Accepted for publication 11 January 2019
Published 12 February 2019 Volume 2019:9 Pages 7—17
DOI https://doi.org/10.2147/PTT.S190507
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Uwe Wollina
Lisa Strohbuecker,1 Hans Koenen,2 Esther van Rijssen,2 Bram van Cranenbroek,2 Esther Fasse,2 Irma Joosten,2 Andreas Körber,1 Christoph Bergmann3
1Department of Dermatology, University Hospital Essen, 45147 Essen, Germany; 2Department of Laboratory Medicine, Laboratory of Medical Immunology, Radboud University Medical Center, Nijmegen, the Netherlands; 3Department of Otorhinolaryngology, University Hospital Essen, 45147 Essen, Germany
Purpose: Psoriasis vulgaris (PV) is an autoimmune-related chronic inflammatory disease of the skin, with both vascular and metabolic effects. Aggravating factors have been identified that initiate and maintain inflammation, including expression of Th1-, Th17-, and Th22-cell derived cytokines. Recently, we showed that the evolutionarily ancient and highly conserved damage-associated molecular pattern molecule “high mobility group box 1 (HMGB1)” is significantly increased in the serum of PV patients with disease progression and is decreased under standard therapies.
Materials and methods: To better understand the role of HMGB1 in the pathogenesis of PV, we recruited 22 untreated psoriatic patients with either mild or severe disease, defined by the Psoriasis Area Severity Index. We assessed HMGB1 and receptor for advanced glycation end products (RAGE) expression in the skin by immunohistochemistry and analyzed the immune-phenotype of Treg and Th17 cells by flow cytometry.
Results: We found increased staining for HMGB1 in the dermis of psoriatic plaques in comparison to uninvolved skin of patients with PV. In addition, the major histocompatibility complex class III-encoded DNA and HMGB1 RAGE, induced by HMGB1, were highly expressed on psoriatic CD8+ T cells and CD4+ Treg. High expression of HMGB1 in the lesional skin was associated with even higher expression of its receptor, RAGE, on the cell surface of keratinocytes in patients with severe PV.
Conclusion: The presence of HMGB1 and RAGE signaling may impact orchestration of chronic inflammation in PV which might have implications for Treg and Th17 cells.
Keywords: HMGB1, RAGE, psoriasis vulgaris, Th17
Introduction
Psoriasis vulgaris (PV) is a chronic, immune-mediated disease that affects the skin and joints with a complex multigenic genetic architecture defined by genome-wide association studies.1 PV is one of the most common skin diseases with a prevalence of ~2% within the general population. It is associated with numerous comorbidities including increased cardiovascular risk or psoriatic arthritis in up to 25% of patients.2 These comorbidities influence patients’ health and quality of life (CLCI), and contribute to a 3- to 7-year reduction in life expectancy when severe.3 Thus, psoriasis is a systemic, inflammatory disease in which increased release of pro-inflammatory cytokines from immune-related cells associated with chronic activation of the innate and adaptive immune systems are mechanisms that mediate long-term damage to multiple tissues and organ.1 Several key mechanisms have been proposed to participate in initiating and maintaining psoriasis, including activation of dendritic cells by self-DNA along with LL37, putative auto-antigens, or the release of pro-inflammatory mediators such as IL-17A, IL-23, or tumor necrosis factor.4 Neither the role of the DNA binding protein RAGE nor the chromatin-associated protein HMGB1 have been extensively studied in this disease.
The activation of keratinocytes leads to an increased production of antimicrobial peptides including the beta defensins, LL-37, and several skin homing chemokines, as well as DNA, perpetuating the inflammatory loop within the skin. However, the precise mechanism underlying the activation of keratinocytes in psoriasis is not fully clarified yet. The T-helper type-17 subset (Th17) produces, in addition to IL-17A, other inflammatory cytokines including IL-23, IL-6, IL-21, IL-1, or TGF-β that can interact with resident dermal cells including keratinocytes, dendritic cells, and endothelial cells.5 Monoclonal antibodies or soluble receptors for IL-17 or IL-23 are promising modalities for targeted psoriasis therapy, as Th17 cells themselves are likely responsible for the chronic course of psoriasis.6
Th17 cells have key functions in several mouse autoimmune disease models and are thought to be similarly involved in human diseases.7,8 The mechanisms leading to the differentiation of Th17 cells is still poorly understood in humans. Differentiated CD4+ T-cell subpopulations display a high grade of plasticity. Their initial differentiation along an individual pathway is not necessarily a terminal end point in T-cell development. In particular, FOXP3+ regulatory T cells (Treg) and Th17 cells demonstrate a high grade of plasticity. This allows for a functional adaptation to various physiological situations during an ongoing immune response.9 In psoriasis vulgaris, Bovenschen et al showed that Treg can differentiate into Th17 cells, particularly when stimulated by IL-23.10 In other autoimmune diseases, including rheumatoid arthritis or graft-versus-host disease, HMGB1 modulates the Treg/ Th17 ratio toward IL-17-producing cells.11,12
The high mobility group nuclear proteins were discovered in 1973 in an effort to understand chromatin organization and later as specific regulators of gene expression.13 HMGB1, the predominant and most abundant member of this family, is a non-histone, chromatin-associated protein present in all metazoans within most eukaryotic cells, helping to stabilize and compact nucleosomes and acting to promote several transcriptional complexes including nuclear hormones, p53/p73 nuclear complexes, and others.14 HMGB1 within the cytosol promotes mitochondrial quality control and autophagy, but extracellular HMGB1 serves as the prototypic damage-associated molecular pattern (DAMP), functioning as a cytokine-like molecule, interacting promiscuously with several cytosolic and cell surface receptors.15 It is released passively during heightened autophagy or cellular necrosis as well as being secreted actively by immune cells including monocytes, macrophages, and dendritic cells.16 We recently demonstrated that serum levels of HMGB1 correlate with disease progression of PV and are decreased following treatment with standard therapies.17 HMGB1 has been shown to interact with toll-like receptors 2 (TLR2) and 4 (TLR4) on keratinocytes in an imiquimod-induced psoriasis like mouse model.18 Following blockade of HMGB1, reduced numbers of infiltrating CD3+ T cells and CD4+ RORγt+Th17 cells in psoriasiform lesions, as well as reduced serum levels of IL-17, could be demonstrated.19
Here, we focused on expression of HMGB1 within the skin, and analyzed in those same patients, peripheral blood immune cells, especially Treg and Th17 cells, as potential sequelae of heightened HMGB1 release and signaling in patients.
Materials and methods
Study participants
The study group used for flow cytometric analysis consisted of 22 patients with PV, who were recruited at the Department of Dermatology, University of Essen, Germany, between 2013 and 2014. Ten healthy subjects were recruited from current or retired members of the clinic staff. All controls and patients were medically healthy, except for the patients who suffered from PV. All participants provided written informed consent according to the Declaration of Helsinki and obtained approval of medical ethics committee of the University Hospital Essen.
Psoriasis disease severity of the patients was measured by the Psoriasis Area Severity Index (PASI). The PASI includes the percentage area affected by psoriasis, which is evaluated in the following four regions of the body (head and neck, upper limbs, trunk, and lower limbs), and the intensity of redness, thickness, and scaling of the plaques. Medium to severe disease was defined by a PASI score <15 and mild disease by a PASI score <5. Eleven patients with mild PV, eleven patients with severe PV, and ten healthy controls were included in this study (Table 1). The mean age was 46, 41, and 31 years for the two patient groups and individual controls, respectively.
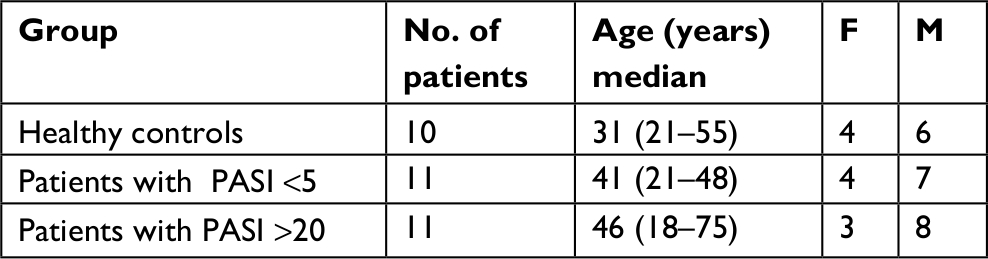 | Table 1 Demographics of the study subjects for evaluation of peripheral blood Abbreviation: PASI, Psoriasis Area Severity Index. |
Skin samples were obtained from five patients with severe PV, five patients with mild PV, and four healthy controls (Table 2). Two skin samples were taken from each of these PV patients: one of a psoriatic plaque (lesional) and one of unaffected/ healthy skin (uninvolved). All biopsies were taken from the upper limb. The mean age was 48, 42, and 23 years for the patients and controls, respectively.
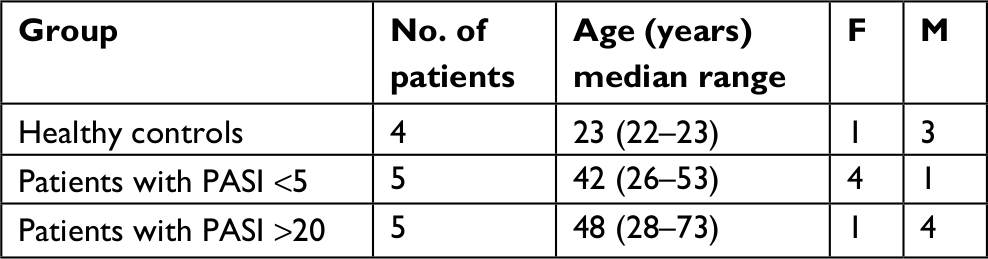 | Table 2 Demographics of the study subjects for tissue samples Abbreviation: PASI, Psoriasis Area Severity Index. |
The patients had no local or systemic therapy for at least 4 weeks. A systemic treatment for PV had been terminated for at least 3 months before study inclusion.
Immunohistochemistry
Tissue sections of 5 µm were generated after embedding the skin biopsies in paraffin. To analyze the presence of human T cells and IL-17 expressing cells in the skin samples, a double immunostaining was conducted according to the manufacturer’s protocol using antihuman primary antibodies directed against CD3 (general T-cell marker, F7.2.38; Abcam, Cambridge, UK) and IL-17A (polyclonal goat; R&D Systems, Minneapolis, MN, USA), respectively. In this double immunostaining, first the presence of IL-17 expressing cells was established; to this end, antigens were retrieved by boiling the sections in Tris/EDTA buffer (50 mM Tris, 2 mM EDTA, pH 9.0). The sections were washed, preincubated for 15 minutes with 20% normal swine serum, and incubated overnight with anti-IL-17A antibody (1:500). To detect IL-17A-producing cells, the sections were stained using the labeled streptavidin-biotin (LSAB) method (Universal LSAB Kit/AP; Dako, Jena, Germany) and visualized using permanent red chromogen (Dako). Finally, for the second immunostaining procedure, the sections were preincubated as described earlier and incubated overnight with anti-CD3 antibody (1:50). CD3 expressing cells were detected by the LSAB method and visualized using vector blue (Vector Laboratories, Philadelphia, PA, USA).
To detect the expression of HMGB1 in the skin samples, single immunostaining was conducted using anti-HMGB1 antibody according to the manufacturer’s protocol (Novus Biologicals, Littleton, CO, USA). After deparaffinization and rehydration, heat-induced antigen retrieval with citrate buffer (pH 6) was performed for 30 minutes. After washing and applying the endogenous peroxidase block for 15 minutes, sections were incubated with 1% bovine serum albumin (BSA; Dako). The antibody was applied (anti-HMGB1-Ab, 1:200) dissolved in 1% BSA, and sections were incubated overnight. The sections were sequentially washed and incubated with Envision+ System HRP labeled polymer anti-rabbit (Dako; for HMGB1) and visualized with DAB cobalt. HMGB1 was counterstained with Mayer’s hematoxylin (Sigma-Aldrich Co., St Louis, MO, USA).
All sections were washed in tap water, dehydrated, and mounted using Permount (Thermo Fisher Scientific, Waltham, MA, USA). Sections were photographed at an objective magnification of 20× or 40× using a microscope (Axioskop2 MOT; Zeiss, Sliedrecht, the Netherlands), digital camera (Axiocam MRc5; Zeiss), and AxioVision software (Zeiss).
For quantification of positive cells, images were saved at 10× objective magnification. The total epidermal area and positive respective markers area were measured. To quantify the cell counts in the dermis, a representative the region of interest (ROI) was drawn from the lowest epidermal papilla till 300 µm depth in the dermis using ImageJ and counted as the number of cells per mm.2 Cell numbers were independently analyzed by two researchers. The presence of HMGB1, IL-17, and CD3 was measured in the dermis of the skin samples.
Flow cytometry and antibodies
Peripheral blood cells were phenotypically analyzed by four- and ten-color immunofluorescence. Briefly, peripheral blood mononuclear cells were washed twice with buffer (PBS containing 0.2% BSA) and labeled with the antibodies of interest conjugated with FITC, PE, ECD, PE-CY5/APC, or PE-CY7. Cells were incubated for 20 minutes in the dark at room temperature, washed twice, and analyzed on a Coulter Epics XL flow cytometer (Beckman Coulter, Fullerton, CA, USA). A total of 5,000–10,000 living cells were collected and analyzed. Isotype-matched antibodies were used to define marker settings. The following conjugated antibodies were used: CD4 (UCHT1; Beckman Coulter), CD4 (MT310; Dako), CD4 (38261; BioLegend, San Diego, CA, USA), CD8 (SFCI21Thy2D3; Beckman Coulter), CD25 (M-A251; BD, Franklin Lakes, NJ, USA), CD127 (R34.34; Beckman Coulter), TLR4 (HTA125; eBioscience, San Diego, CA, USA), TLR2 (TL2.1; eBioscience), RAGE (polyclonal; Biologo, Kronshagen, Germany), IL23R (218213; RD Systems), and CCR6 (11A9; BD).
To detect the intra-cytoplasmic cytokines or the transcription factors FOXP3 (PCH101; eBioscience) and RORγt (AFKJS-9; eBioscience), the cells of interest were fixed and permeabilized (Fix and Perm reagent A and B; Caltag Laboratories, Vienna, Austria) according to the instructions of the manufacturer and, thereafter, labeled with the monoclonal antibody of interest. After 45-minute incubation at room temperature, the cells were washed three times and analyzed by flow cytometry. Appropriate isotype control monoclonal antibodies were used for marker settings. Data were analyzed by Kaluza Flow Cytometry Analysis Software (Beckman Coulter).
Statistical analyses
The data of flow cytometry and immune histochemistry of the three different groups (patients with mild and severe PV and healthy controls) were compared using the nonparametric Mann–Whitney U test. Statistical analyses were performed using the GraphPad Prism 5 software. A value of P<0.05 was considered statistically significant.
Results
Increased HMGB1 expression in the dermis of patients with psoriasis vulgaris
We analyzed the expression of HMGB1 in lesional and non-lesional skin biopsies of recently recruited psoriatic patients and healthy controls, using immunohistochemistry. The histopathological features of the two biopsy locations of a psoriatic patient are illustrated in Figure 1C and D. With respect to the general histology, lesional skin (Figure 1C) revealed the characteristic psoriatic features of thickened epidermis, parakeratosis, elongated rete ridges, and a mixed cellular infiltrate. A detailed view of lesional skin (Figure 1D) shows the accumulation of HMGB1 expressing cells in the dermis along with cytosolic and extracellular HMGB1. The distant uninvolved skin (Figure 1B) did not reveal any pathological changes and lacks HMGB1 extranuclear expressing cells.
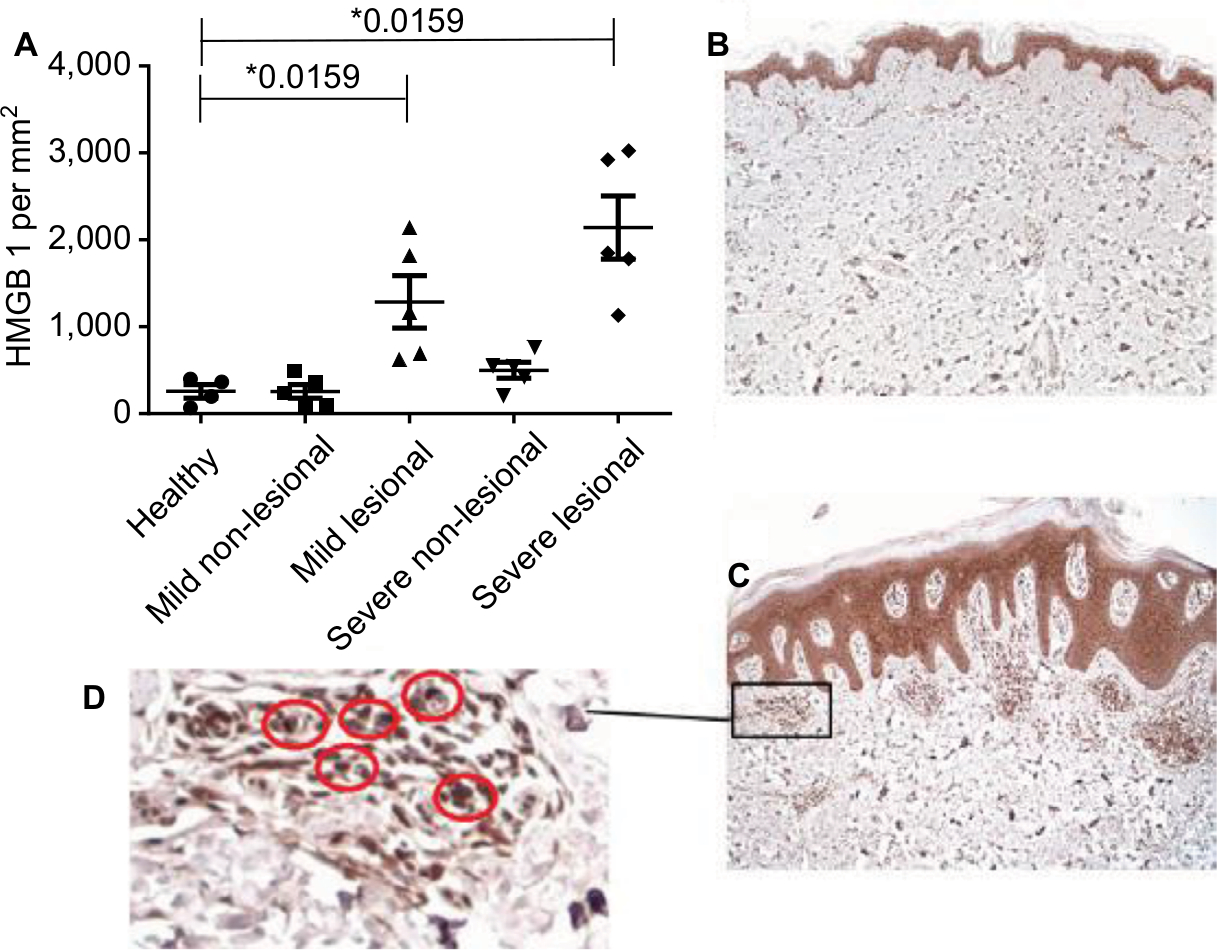 | Figure 1 HMGB1 staining is enhanced in the dermis of psoriatic skin compared with uninvolved skin. Notes: (A) HMGB1 in the dermis of healthy controls (n=4) and patients with mild (n=5) and severe (n=5) psoriasis vulgaris was assessed using immunostaining. Cutaneous cytokine levels in individual patients as well as mean levels are shown. *P<0.05, Mann–Whitney U test. (B) Immunostaining of HMGB1 of uninvolved skin in a psoriatic patient. (C) Immunostaining of HMGB1 of a skin biopsy out of a psoriatic plaque (lesional skin). (D) Detailed view of HMGB1 molecules in dermal immunostaining. Red circles indicate HMGB1-positive cells. |
In the lesional dermal skin of mild and severe psoriatic patients, we found a significant higher number of HMGB1 extranuclear positive cells as compared with healthy controls (mean ± SD; 1,164±672.9 HMGB1 expressing cells/mm2, n=5; 2,142±809.4, n=5; 257±154.2, n=4; P=0.0159; respectively; Figure 1A). The percentage of extranuclear HMGB1 expressing cells in the lesional dermis related with disease progression (Figure 1A).
Within psoriatic patients, the uninvolved dermis revealed fewer numbers of extranuclear HMGB1 expressing cells/mm2 dermis than the lesional skin samples. In the uninvolved dermis of the mild psoriatic patients, the number of extranuclear HMGB1 expressing cells was similar to that found in the dermis of the healthy controls (mean ± SD; 256±173.5 HMGB1 expressing cells/mm2, n=5; 257±154.2, n=4; Figure 1A). Interestingly, in the uninvolved dermis of the severe psoriatic patients, we measured approximately double the amount of extranuclear HMGB1 expressing cells as compared to patients with mild psoriasis and healthy individuals (mean ± SD; 499±203.4 extranuclear HMGB1 expressing cells/mm2, n=5; 256±173.5, n=5; 257±154.2, n=4; respectively) (Figure 1A).
Lesional skin, thus, reveals the highest levels of extranuclear HMGB1 expressing cells, suggesting a correlation with disease activity; HMGB1 expressing cells in non-lesional skin were only increased in severe PV patients.
IL-17A and CD3 expression in the dermis
Th17 cells and their signature cytokine IL-17A are known to play a key role in the pathogenesis of psoriasis vulgaris.20 Also, in psoriatic patients, phenotypic Tregs are found to secrete IL-17.10 HMGB1 seems to influence the balance of Th17 cells and Tregs toward IL-17-producing cells.11,12,21,22 To characterize the IL-17-producing T cells, we analyzed the presence of CD3 and/or IL-17-producing cells in skin biopsies of psoriasis patients and healthy controls by immunostaining for both CD3 and IL-17A (Figure 2A–E).
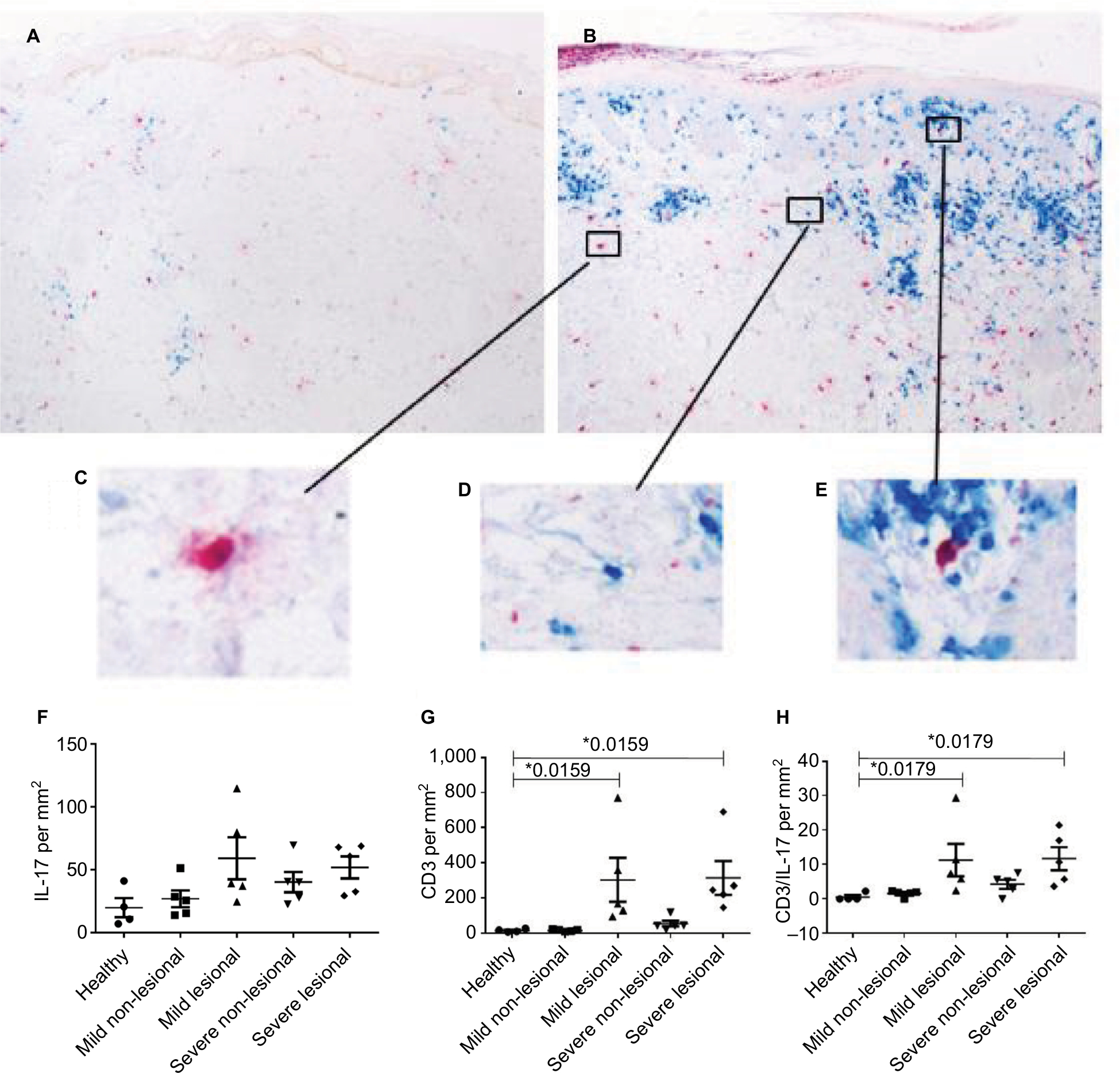 | Figure 2 Marked increases in IL-17 expression and CD3+ T cells within the dermis of affected skin. Notes: (A) Immunostaining of IL-17 (red) and CD3 (blue) of an uninvolved skin biopsy of a psoriatic patient. (B) Immunostaining of IL-17 (red) and CD3 (blue) of a skin biopsy out of a psoriatic plaque (lesional skin). (C) Detailed view of IL-17 staining in immunohistochemical analysis. (D) Detailed view of a CD3+ cell in the immunostaining. (E) Detailed view of a IL-17+ CD3+ cell in the double immunostaining. (F–H) IL-17, CD3, and IL-17+ CD3 double staining in the dermis of healthy controls (n=4) (F) and patients with mild (n=5) (G) and severe (n=5) (H) psoriasis vulgaris assessed with immunostaining. Cutaneous IL-17 expressing cells/mm2 in individual patients as well as the arithmetic mean are shown. *P<0.05, Mann–Whitney U test. |
As expected in lesional dermis of both patients with severe or mild PV, the number of CD3 expressing cells was higher as compared to that in the dermis of healthy controls (mean ± SD; 314±215 CD3+ cells/mm2, n=5; 303±279.4, n=5; 15±9, n=4; P=0.0179, Figure 2G). This implies on average a 20-fold, significant increase in T cells in the lesional dermis of psoriatic patients as compared to the dermis of healthy controls, and this was observed irrespective of disease severity, confirming the results of others.23
The uninvolved skin of mild psoriatic patients showed similar CD3 expression levels, as found in the dermis of healthy controls, whereas the severe psoriatic group showed a higher number of CD3 expressing cells (55±37, n=5; 15.8±6.1, n=5; and 15.3±9, n=4 in severe PV vs mild PV, and healthy controls, respectively) (Figure 2G).
Lesional skin of both mild and severe PV patients revealed strongly increased levels of CD3+ T-cell expression, while an increase of CD3 in non-lesional skin was only observed in severe PV patients.
The presence of IL-17-producing T cells, as indicated by IL-17 and CD3 coexpression, shows a trend toward higher numbers in the transition from uninvolved to lesional skin. In the uninvolved dermis of PV patients and healthy controls, the numbers of CD3+ IL-17+ cells were very low (mean ± SD; 1.7±0.9 CD3+ IL-17+ cells / mm2, n=5; 4±2.9, n=5; 0.5±1.1, n=4; respectively) (Figure 2H). Conversely, in the lesional dermis of mild and severe psoriatic patients, we detected the presence of CD3+ IL-17+ cells (mean ± SD; 11±10.6, n=5; 12±7.5, n=5; 0.5±1.1, n=4; P=0.0179). We observed a significantly higher number of IL-17+ T cells in the lesional dermis of psoriatic patients compared to healthy controls (Figure 2H). Most of the IL-17A found in the skin of psoriatic patients and healthy controls was not associated with CD3 (Figure 2F), and could well be expressed by neutrophils or mast cells, as has been previously reported for psoriatic skin.24–26 In the lesional dermis of psoriatic patients, a trend to higher numbers of single IL-17A expressing cells was found as compared to the dermis of healthy donors (mean ± SD; 52±19.3, n=5; 59±37.3, n=5; 20±15.2, n=4; P=0.11 and P=0.064, in severe, mild patients, and healthy controls, respectively) (Figure 2F). In the uninvolved dermis of the severe psoriatic patients, as compared to the dermis of mild psoriatic patients or healthy controls, we observed a trend toward higher expression of single IL-17A expressing cells (40±18.2, n=5; 25±15, n=5; 20±15.3, n=4 in severe PV vs mild PV, and healthy controls, respectively) (Figure 2F).
Immune cell surface expression of RAGE, TLR2, and TLR4
After having observed increased numbers of IL-17-producing CD3+ cells and elevated levels of HMGB1 molecules in the psoriatic skin, we examined peripheral blood mononuclear cells for the expression of receptors for HMGB1. TLR2, TLR4, and RAGE are defined signaling receptors for HMGB1. Hence, we measured the expression of these receptors on the surface of CD4+ and CD8+ cells, as well as on CD4+ Treg, defined by CD4+ CD25highCD127low surface expression as previously described by Liu et al, in the peripheral blood of patients with psoriasis vulgaris and healthy controls using flow cytometry.27
There was no difference in the expression level of the receptors TLR2 and TLR4 and RAGE between healthy donors, mild or severe psoriatic patients (data not shown and Figure 3A). Subgroup analysis revealed that the percentage of RAGE expressing CD4+ cells in the peripheral blood of psoriatic patients, however, decreased with disease progression (8.9%, 11.2%, and 13.1% of all CD4+ cells were positive for RAGE in severe PV (n=11), mild PV (n=11), and healthy controls (n=10), respectively) (Figure 3B).
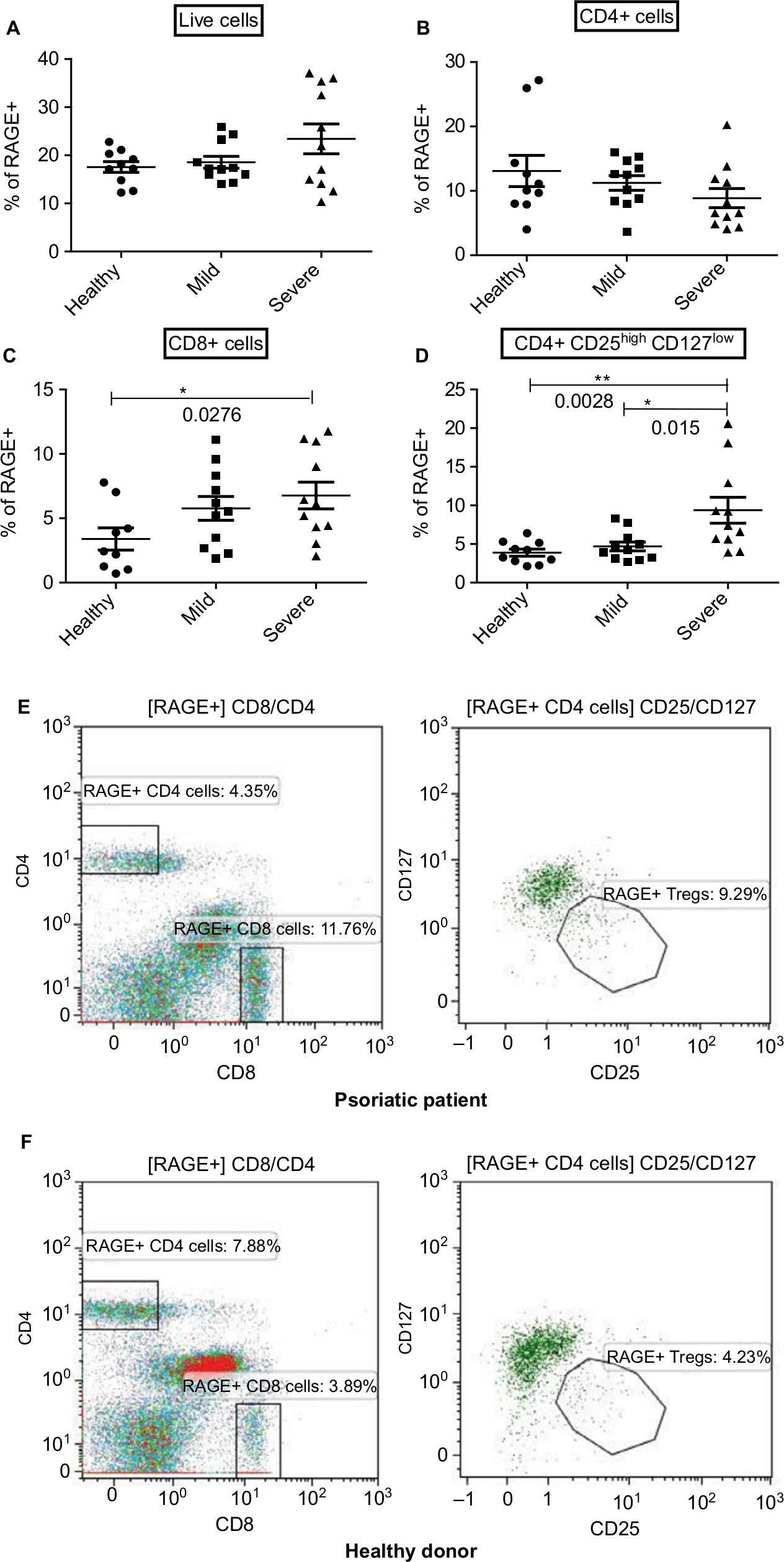 | Figure 3 Expression of RAGE on the surface of CD8+ and CD4+ T cells as well as CD4+ regulatory T cells correlates with disease severity. Notes: Flow cytometry of RAGE expression on all live immune cells (A), CD8+ T Cells (B), CD4+ T Cells (C), and CD4+ Treg (D) of healthy controls (n=10) and patients with mild (n=11) and severe (n=11) psoriasis vulgaris. Representative flow plots are displayed for RAGE+ Tregs in patients (E) and controls (F). RAGE expressions in individual patients as well as arithmetic means are shown. *P<0.05, **P<0.01, Mann–Whitney U test. |
Increased percentages of RAGE expressing CD8+ cells were found in PV patients as compared to the healthy controls (6.8%, 5.8%, and 3.4% in severe PV, mild PV, and healthy controls, respectively; P=0.0276) (Figure 3C) with expression positively correlating with disease progression. The percentage of CD4+ Treg that expressed RAGE on their surface was also significantly higher in the peripheral blood of psoriatic patients as compared with healthy controls (9.3%, 4.25%, and 3.9% of the gated CD4+ Treg were positive for RAGE in severe PV, mild PV, and healthy controls, respectively; P=0.0195) (Figure 3D–F). Of note, increased numbers of RAGE expressing CD8+ cells and CD4+ Treg are paralleled by increased serum levels of HMGB1, as we previously reported, in severe psoriatic patients, suggesting a possible HMGB1–RAGE relationship that is consistent with HMGB1’s ability to upregulate and induce RAGE expression.17
RORγt, CCR6, and IL-23R expression on CD4+ cells and CD4+ Treg in patients with PV
HMGB1 decreases the Treg/Th17 ratio in individual autoimmune diseases.11,21,22,28,29 Thus, we examined whether increased HMGB1 and RAGE expression in PV was associated with increased Th17 levels in the peripheral blood of PV patients and healthy controls using flow cytometry. We examined both CCR6 and IL-23-receptor (IL-23R) surface expression, and the intracellular expression of the transcription factor RORγt as markers for Th17 cells.30
CD4+ lymphocytes expressing CCR6 and IL-23R on their surface were significantly increased in the peripheral blood of severe psoriatic patients (0.7%, 0.35%, and 0.4% in severe PV, mild PV, and healthy controls; P=0.0385) (Figure 4A). Also, the number of CCR6 and IL-23R expressing CD4+ Treg was increased in severe psoriatic patients (1.2%, 0.5%, and 0.6% in severe PV, mild PV, and healthy controls; P=0.0195) (Figure 4B).
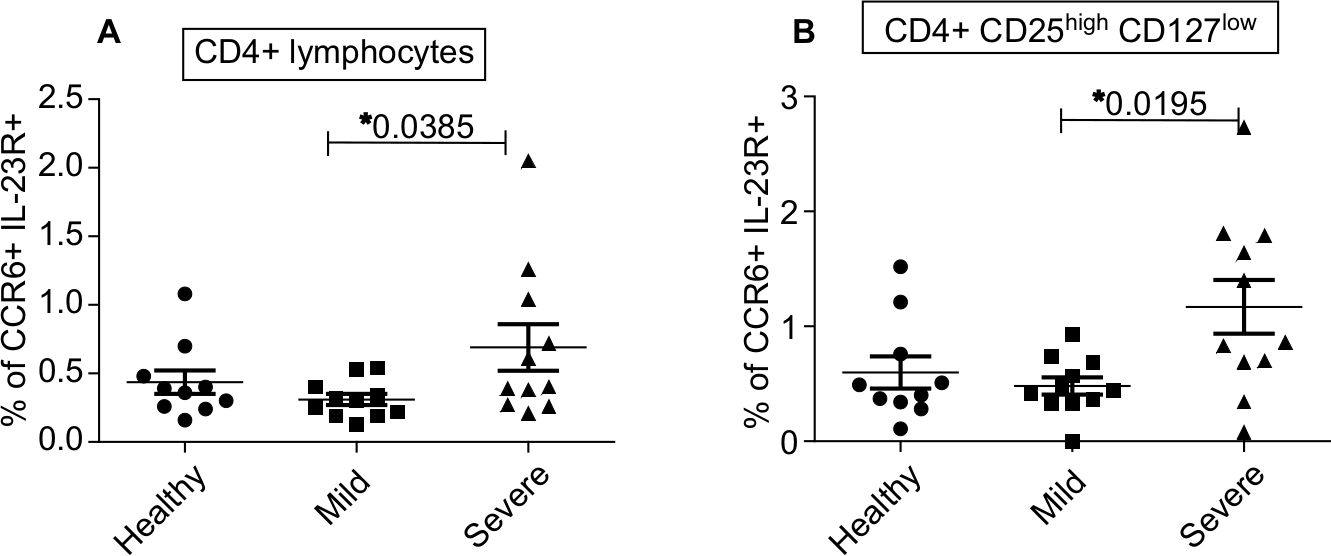 | Figure 4 Markedly enhanced coexpression of CCR6 and the IL-23 receptor on the surface of CD4+ Tregs and CD4+ T cells within affected skin. Notes: Flow cytometric assessment of CCR6 and IL-23 receptor on CD4+ T Cells (A) and CD4+ Treg (B) of healthy controls (n=10) and patients with mild (n=11) or severe (n=11) psoriasis vulgaris. CCR6 and IL-23R expressions in individual patients as well as the arithmetic mean are shown. *P<0.05, Mann–Whitney U test. |
In contrast, we observed a trend toward reduced percentages of RORγt+cells in PV patients as compared to healthy controls in both the CD4+ CD25highFOXP3high Treg (2.2%, 3.3%, and 4% in severe PV, mild PV, and healthy controls) (Figure 5A) and total CD4+ population (1.7%, 1.6%, and 1.0% in severe PV, mild PV, and healthy controls) (Figure 5B). The percentages of thymic-derived FOXP3+ CD4+ Treg in the peripheral blood of PV patients and healthy controls were similar (data not shown).
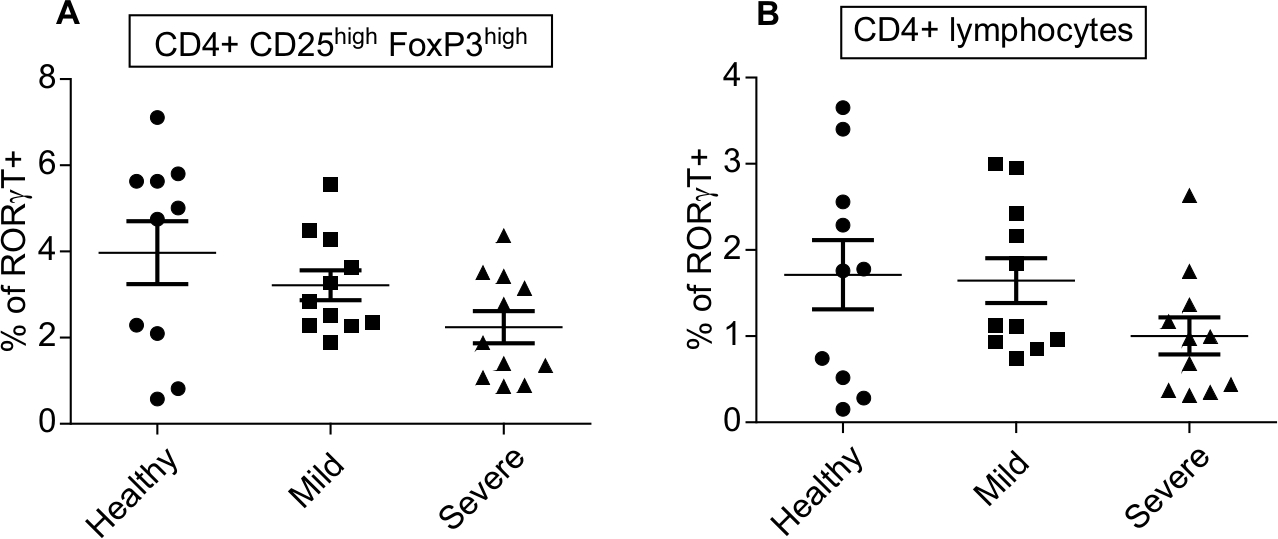 | Figure 5 Reduced expression of the transcription factor RORγt in CD4+ T cells and CD4+ regulatory T cells. Notes: Intracellular flow cytometry of RORγt in CD4+ Treg (A) and CD4+ T cells (B) of healthy controls (n=10) and patients with mild (n=11) and severe (n=11) psoriasis vulgaris. RORγt expressions in individual patients as well as mean are shown. |
Discussion
HMGB1 is emerging as a centrally important factor found in the serum of patients with trauma, sepsis, cancer, and various inflammatory disorders.31–33 We previously demonstrated that HMGB1 levels were increased in the sera of psoriatic patients, and these levels related to disease progression. Moreover, upon standard psoriasis therapy, we found a reduction in these HMGB1 serum levels.17 These previous finding suggests a role for HMGB1 and by extension, its receptors TLR2, TLR4, and/or RAGE in the immune pathology of psoriasis.
In the present work, we show that HMGB1 expressing cells are significantly increased in the skin of psoriatic patients and that its receptor RAGE is highly expressed on the cell surface of both circulating CD8+ cells and CD4+ Treg in patients with psoriasis vulgaris, which is linked to disease progression. Furthermore, we measured a higher number of IL-17-producing cells in the lesional skin of psoriatic patients in comparison to the uninvolved skin of the same patient, or the skin of healthy controls. We also detected an increased expression of IL-23-receptor and CCR6 on CD4+ cells, as putative Th17 cells, and CD4+ Treg.
HMGB1 is a chromatin-associated molecule that acts to promote access to various transcription factors, stabilizing nucleosomes and further compacting the DNA.34 During stress, it is released by macrophages or by necrotic cells, acting as a pro-inflammatory molecule to promote a wound healing response, recruiting mesangioblasts, neutrophils, platelets, macrophages, and ultimately adaptive immune cells in several autoimmune diseases and cancer.35–37 HMGB1 is thus the prototypic DAMP molecule or an endogenous danger signal.16,38 Our findings deepen and extend this work to demonstrate its presence in the skin of patients with psoriasis as well as peripheral blood serum and cellular changes, correlating with disease severity.
Several studies have highlighted involvement of HMGB1 in individual autoimmune diseases. For example, in skin lesions of patients with lupus erythematosus, in the sera of patients with systemic sclerosis, or in the minor salivary glands of patients with Sjogren’s syndrome elevated concentrations of HMGB1 are found.39–41 The role of HMGB1 in these autoimmune diseases has not yet been fully elucidated. An emergent finding in the last 5 years has been that HMGB1 signaling promotes conversion of Treg into IL-17-producing cells. This differentiation has been shown in graft-versus-host disease, hepatitis B, rheumatic arthritis, autoimmune myocarditis, and atherosclerosis.11,12,21,22,29 This shift away from Tregs promotes tissue destruction and production of pro-inflammatory Th17 cells. Uncontrolled differentiation of Treg into Th17 cells could be an important step in the onset and maintenance of autoimmune diseases. In PV, Treg readily differentiate into IL-17-producing cells. This differentiation was promoted by the cytokine IL-23 and correlated with the loss of FOXP3 and enhancement of RORγt expression.10
We analyzed the expression level of these three receptors of HMGB1 on the surface of peripheral blood immune cells and found a significant average twofold increase of RAGE expression on the surface of Treg and CD8+ cells of severe psoriatic patients as compared to healthy donors. The expression levels of TLR2 and TLR4 showed no difference in our psoriasis and healthy control groups. HMGB1 in the serum likely promotes the expression of RAGE as the predominant altered receptor (when compared with TLR2 and TLR4) in the peripheral blood. HMGB1 and its receptor RAGE on Treg and on CD8+ cells play a role in the pathogenesis of psoriasis, suggesting that efforts to target these molecules could indeed be part of the armamentarium for treating patients.42,43 We described earlier that the differentiation of Treg into IL-17-producing cells is accompanied by a higher expression of the transcription factor RORγt, a marker for Th17 cells.44 Higher expression levels of RORγt and a decrease of FOXP3 following in vitro stimulation with HMGB1 of peripheral mononuclear cells of patients with hepatitis B and atherosclerosis was found.21,29 Our analysis revealed that peripheral blood T cells showed decreased RORγt expression in Treg and CD4+ cells with disease progression in patients with PV. IL-17-producing cells, which express higher percentages of RORγt, might be recruited to the location of primary inflammation in the skin or are induced at site. Indeed, we observed that Th17 cells were overrepresented in lesional psoriatic skin, representing 49%–93% of CD4+ cells compared with 3%–18% in peripheral blood of these patients and increased following in vitro– activation to a greater extent than cells in the periphery.45 Immunostaining of CD3 and IL-17 revealed higher numbers of CD3+ IL-17+ cells in the lesional skin when compared to uninvolved skin of a psoriatic patient or healthy controls, indicating the possible relevance of these cells at site and for disease progression.
We showed a significant increase of Treg and CD4+ cells expressing CCR6 and IL-23R in the peripheral blood of severe psoriatic patients. The combined expression of these two receptors is a useful marker to identify (precursor) Th17 cells. Expression of CCR6 precisely identified all IL-17-producing T cells with differing levels of RORγt expression. Th17 clones derived from these cells not only expressed RORγt, but also remarkable amounts of the Th1 transcription factor T-bet.30 These results highlight that the initial apparent differentiation of T cells is contradicted by a high grade of plasticity of T cells, allowing functional adaptation to various physiological situations during an ongoing immune response.9
Following injection of HMGB1-neutralizing antibody in the peritoneal cavity of mice with psoriatic lesions, reduced numbers of infiltrating CD3+ T cells and CD4+ RORγt+Th17 cells in psoriatic lesions were found, as well as reduced serum levels of IL-17.19 HMGB1 itself promotes the Th17 immune response in PV via caspase 1 cleavage and subsequent release of IL-18 from keratinocytes.19 We found that HMGB1 could also interact directly with CD4+ Treg and CD8+ cells via binding to RAGE.17 Inhibition of HMGB1 suppresses immigration of immune cells in the skin and secretion of pro-inflammatory cytokines in the peripheral blood of psorasiform mice.19 Targeting HMGB1 seems a rational target for therapy of patients with psoriasis vulgaris.
Conclusion
The role of HMGB1 in autoimmune diseases and especially PV is likely due to secretion by keratinocytes and several types of immune cells including activated macrophages, mature dendritic cells, and natural killer cells in response to injury, infection, or other inflammatory stimuli.37,46,47 It plays an important role in the pathogenesis of PV by influencing homeostasis, limiting the natural immunosuppressive phenotype and promoting pro-inflammatory conditions. We speculate that the interaction of HMGB1 promotes RAGE expression, thereby driving a Th17 shift in patients with PV and promoting persistence of the inflammatory state. More detailed functional assays will be needed to fully evaluate the association of serum and skin cytokine levels of HMGB1, the expression of RAGE on psoriatic Treg, and the shift from regulatory T cells to Th17-like cells. Our studies promote a subsequent study with larger patient cohort and further defined immune subsets to evaluate targeting HMGB1 as a biomarker or as a potential therapeutic target during disease progression of PV.
Acknowledgment
This study received institutional support from the Department of Dermatology, University Hospital Essen, Essen, Germany.
Author contributions
LS performed experiments and wrote manuscript. HK analyzed and interpreted the data and wrote manuscript. EVR, BVC, and EF performed the experiments. IJ analyzed and interpreted the data and wrote manuscript. AK provided patients’ material and wrote manuscript. CB concepted and supervised the study, and wrote manuscript. All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Greb JE, Goldminz AM, Elder JT, et al. Psoriasis. Nat Rev Dis Primers. 2016;2:16082. | ||
Mahil SK, Capon F, Barker JN. Update on psoriasis immunopathogenesis and targeted immunotherapy. Semin Immunopathol. 2016;38(1):11–27. | ||
Augustin M. Cumulative life course impairment: identifying patients at risk. Curr Probl Dermatol. 2013;44:74–81. | ||
Reich K, Papp KA, Matheson RT, et al. Evidence that a neutrophil-keratinocyte crosstalk is an early target of IL-17A inhibition in psoriasis. Exp Dermatol. 2015;24(7):529–535. | ||
Fc E, Bruck J, Holstein J, Hirahara K, Ghoreschi K. Recent advances in understanding psoriasis. F1000Res. 2016;5. pii: F1000 Faculty Rev-770. | ||
Kim J, Krueger JG. Highly effective new treatments for psoriasis target the IL-23/Type 17 T cell autoimmune axis. Annu Rev Med. 2017;68(1):255–269. | ||
Bettelli E, Korn T, Kuchroo VK. Th17: the third member of the effector T cell trilogy. Curr Opin Immunol. 2007;19(6):652–657. | ||
Stockinger B, Veldhoen M. Differentiation and function of Th17 T cells. Curr Opin Immunol. 2007;19(3):281–286. | ||
Kleinewietfeld M, Hafler DA. The plasticity of human Treg and Th17 cells and its role in autoimmunity. Semin Immunol. 2013;25(4):305–312. | ||
Bovenschen HJ, van de Kerkhof PC, van Erp PE, Woestenenk R, Joosten I, Koenen HJ. Foxp3+ regulatory T cells of psoriasis patients easily differentiate into IL-17A-producing cells and are found in lesional skin. J Invest Dermatol. 2011;131(9):1853–1860. | ||
Xu YJ, Li L, Chen Y, et al. Role of HMGB1 in regulation of STAT3 expression in CD4(+) T cells from patients with aGVHD after allogeneic hematopoietic stem cell transplantation. Clin Immunol. 2015;161(2):278–283. | ||
Shi Y, Sandoghchian Shotorbani S, Su Z, et al. Enhanced HMGB1 expression may contribute to Th17 cells activation in rheumatoid arthritis. Clin Dev Immunol. 2012;2012(3):1–8. | ||
Goodwin GH, Sanders C, Johns EW. A new group of chromatin-associated proteins with a high content of acidic and basic amino acids. Eur J Biochem. 1973;38(1):14–19. | ||
Stros M, Ozaki T, Bacikova A, Kageyama H, Nakagawara A. HMGB1 and HMGB2 cell-specifically down-regulate the p53- and p73-dependent sequence-specific transactivation from the human Bax gene promoter. J Biol Chem. 2002;277(9):7157–7164. | ||
Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol. 2005;5(4):331–342. | ||
Scaffidi P, Misteli T, Bianchi ME. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature. 2002;418(6894):191–195. | ||
Bergmann C, Strohbuecker L, Lotfi R, et al. High mobility group box 1 is increased in the sera of psoriatic patients with disease progression. J Eur Acad Dermatol Venereol. 2016;30(3):435–441. | ||
van der Fits L, Mourits S, Voerman JS, et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J Immunol. 2009;182(9):5836–5845. | ||
Zhang W, Guo S, Li B, et al. Proinflammatory effect of high-mobility group protein B1 on keratinocytes: an autocrine mechanism underlying psoriasis development. J. Pathol. 2017;241(3):392–404. | ||
Di Cesare A, Di, di Meglio P, Nestle FO. The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J Invest Dermatol. 2009;129(6):1339–1350. | ||
Li J, Wang FP, She WM, et al. Enhanced high-mobility group box 1 (HMGB1) modulates regulatory T cells (Treg)/T helper 17 (Th17) balance via toll-like receptor (TLR)-4-interleukin (IL)-6 pathway in patients with chronic hepatitis B. J Viral Hepat. 2014;21(2):129–140. | ||
Su Z, Sun C, Zhou C, et al. HMGB1 blockade attenuates experimental autoimmune myocarditis and suppresses Th17-cell expansion. Eur J Immunol. 2011;41(12):3586–3595. | ||
Lowes MA, Kikuchi T, Fuentes-Duculan J, et al. Psoriasis vulgaris lesions contain Discrete populations of Th1 and Th17 T cells. J Invest Dermatol. 2008;128(5):1207–1211. | ||
Res PC, Piskin G, de Boer OJ, et al. Overrepresentation of IL-17A and IL-22 producing CD8 T cells in lesional skin suggests their involvement in the pathogenesis of psoriasis. PLoS One. 2010;5(11):e14108. | ||
Lin AM, Rubin CJ, Khandpur R, et al. Mast cells and neutrophils release IL-17 through extracellular trap formation in psoriasis. J Immunol. 2011;187(1):490–500. | ||
Schön MP, Erpenbeck L. The Interleukin-23/Interleukin-17 axis links adaptive and innate immunity in psoriasis. Front Immunol. 2018;9:1323. | ||
Liu W, Putnam AL, Xu-Yu Z, et al. CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4 + T reg cells. J Exp Med. 2006;203(7):1701–1711. | ||
Shi Y, Sandoghchian Shotorbani S, Su Z, et al. Enhanced HMGB1 expression may contribute to Th17 cells activation in rheumatoid arthritis. Clin Dev Immunol. 2012;2012(3):1–8. | ||
Ding JW, Zheng XX, Zhou T, Tong XH, Luo CY, Wang XA. HMGB1Modulates the Treg/Th17 ratio in atherosclerotic patients. J Atheroscler Thromb. 2016;23(6):737–745. | ||
Acosta-Rodriguez EV, Rivino L, Geginat J, et al. Surface phenotype and antigenic specificity of human interleukin 17–producing T helper memory cells. Nat Immunol. 2007;8(6):639–646. | ||
Wang H, Yang H, Tracey KJ. Extracellular role of HMGB1 in inflammation and sepsis. J Intern Med. 2004;255(3):320–331. | ||
Sims GP, Rowe DC, Rietdijk ST, Herbst R, Coyle AJ. HMGB1 and RAGE in inflammation and cancer. Annu Rev Immunol. 2010;28(1):367–388. | ||
Harris HE, Andersson U, Pisetsky DS. HMGB1: a multifunctional alarmin driving autoimmune and inflammatory disease. Nat Rev Rheumatol. 2012;8(4):195–202. | ||
Walker JM, Gooderham K, Hastings JR, Mayes E, Johns EW. The primary structures of non-histone chromosomal proteins HMG 1 and 2. FEBS Lett. 1980;122(2):264–270. | ||
Zhang Q, O’Hearn S, Kavalukas SL, Barbul A. Role of high mobility group box 1 (HMGB1) in wound healing. J Surg Res. 2012;176(1):343–347. | ||
Straino S, Di Carlo A, Mangoni A, et al. High-mobility group box 1 protein in human and murine skin: involvement in wound healing. J Invest Dermatol. 2008;128(6):1545–1553. | ||
Bonaldi T, Talamo F, Scaffidi P, et al. Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. Embo J. 2003;22(20):5551–5560. | ||
Lotze MT, Deisseroth A, Rubartelli A. Damage associated molecular pattern molecules. Clin Immunol. 2007;124(1):1–4. | ||
Popovic K, Ek M, Espinosa A, et al. Increased expression of the novel proinflammatory cytokine high mobility group box chromosomal protein 1 in skin lesions of patients with lupus erythematosus. Arthritis Rheum. 2005;52(11):3639–3645. | ||
Yoshizaki A, Komura K, Iwata Y, et al. Clinical significance of serum HMGB-1 and sRAGE levels in systemic sclerosis: association with disease severity. J Clin Immunol. 2009;29(2):180–189. | ||
Ek M, Popovic K, Harris HE, Nauclér CS, Wahren-Herlenius M, Erlandsson Harris H. Increased extracellular levels of the novel proinflammatory cytokine high mobility group box chromosomal protein 1 in minor salivary glands of patients with Sjögren’s syndrome. Arthritis Rheum. 2006;54(7):2289–2294. | ||
Elangovan I, Thirugnanam S, Chen A, et al. Targeting receptor for advanced glycation end products (RAGE) expression induces apoptosis and inhibits prostate tumor growth. Biochem Biophys Res Commun. 2012;417(4):1133–1138. | ||
Radia ALM, Yaser ALM, Ma X, et al. Specific siRNA targeting receptor for advanced glycation end products (RAGE) decreases proliferation in human breast cancer cell lines. Int J Mol Sci. 2013;14(4):7959–7978. | ||
Koenen HJ, Smeets RL, Vink PM, van Rijssen E, Boots AM, Joosten I. Human CD25highFoxp3pos regulatory T cells differentiate into IL-17-producing cells. Blood. 2008;112(6):2340–2352. | ||
Lewis BJ, Rajpara S, Haggart AM, Wilson HM, Barker RN, Ormerod AD. Predominance of activated, clonally expanded T helper type 17 cells within the CD4+ T cell population in psoriatic lesions. Clin Exp Immunol. 2013;173(1):38–46. | ||
Wang H, Bloom O, Zhang M, et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science. 1999;285(5425):248–251. | ||
Semino C, Angelini G, Poggi A, Rubartelli A. NK/iDC interaction results in IL-18 secretion by DCs at the synaptic cleft followed by NK cell activation and release of the DC maturation factor HMGB1. Blood. 2005;106(2):609–616. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.