Back to Journals » Neuropsychiatric Disease and Treatment » Volume 11
In vitro screening of major neurotransmitter systems possibly involved in the mechanism of action of antibodies to S100 protein in released-active form
Authors Gorbunov E, Ertuzun I, Kachaeva E, Tarasov S, Epstein O
Received 15 July 2015
Accepted for publication 10 August 2015
Published 3 November 2015 Volume 2015:11 Pages 2837—2846
DOI https://doi.org/10.2147/NDT.S92456
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Evgeniy A Gorbunov, Irina A Ertuzun, Evgeniya V Kachaeva, Sergey A Tarasov, Oleg I Epstein
OOO “NPF “MATERIA MEDICA HOLDING”, Moscow, Russian Federation
Abstract: Experimentally and clinically, it was shown that released-active form of antibodies to S100 protein (RAF of Abs to S100) exerts a wide range of pharmacological activities: anxiolytic, antiasthenic, antiaggressive, stress-protective, antihypoxic, antiischemic, neuroprotective, and nootropic. The purpose of this study was to determine the influence of RAF of Abs to S100 on major neurotransmitter systems (serotoninergic, GABAergic, dopaminergic, and on sigma receptors as well) which are possibly involved in its mechanism of pharmacological activity. Radioligand binding assays were used for assessment of the drug influence on ligand–receptor interaction. [35S]GTPγS binding assay, cyclic adenosine monophosphate HTRF™, cellular dielectric spectroscopy assays, and assays based on measurement of intracellular concentration of Ca2+ ions were used for assessment of agonist or antagonist properties of the drug toward receptors. RAF of Abs to S100 increased radioligand binding to 5-HT1F, 5-HT2B, 5-HT2Cedited, 5-HT3, and to D3 receptors by 142.0%, 131.9%, 149.3%, 120.7%, and 126.3%, respectively. Also, the drug significantly inhibited specific binding of radioligands to GABAB1A/B2 receptors by 25.8%, and to both native and recombinant human sigma1 receptors by 75.3% and 40.32%, respectively. In the functional assays, it was shown that the drug exerted antagonism at 5-HT1B, D3, and GABAB1A/B2 receptors inhibiting agonist-induced responses by 23.24%, 32.76%, and 30.2%, respectively. On the contrary, the drug exerted an agonist effect at 5-HT1A receptors enhancing receptor functional activity by 28.0%. The pharmacological profiling of RAF of Abs to S100 among 27 receptor provides evidence for drug-related modification of major neurotransmitter systems.
Keywords: dopamine agent, released-activity, serotonin agent, sigma1 receptor
Introduction
The aim of this study is to evaluate the mechanisms of action of the commercial drug Tenoten®, which has been used in the treatment of psychoneurotic and neurogenic disorders. The active ingredient of the drug is released-active form of antibodies to S100 protein (RAF of Abs to S100). Released-activity is defined as follows: pretreated ultrahigh dilutions cause modifying effect on the original substance.1 It has been shown that drugs of this class have a fundamentally novel pro-antigenic action (codirectional with antigen), since the released-active form of the antibody alters the interaction of the specific antigen (molecule) with its target by mechanism of conformation modification.2 The efficacy and safety of the drugs were intensively studied and proved in different experimental models and in clinical studies as well.3–11
RAF of Abs to S100 exert effects similar to those of the S100 protein. RAF of Abs to S100 possesses membranotropic effects: induced membrane depolarization, reduced the amplitude or completely blocked the action potential, accelerated the maximal rise of the action potential, reduced maximal conductance, and facilitated the steady-state inactivation of Ca2+ channels.12 These effects are unidirectional with a regulatory effect of the S100 protein on the electrical activity of the neuron membrane.13 RAF of Abs to S100 could modify S100 protein activity.14 Moreover, RAF of Abs to S100 without exerting direct influence on the induction of long-term post-tetanic potentiation prevents the blocking action of large doses of antibodies to S100 on long-term post-tetanic potentiation, which indicates its modifying effect.15 It has been previously shown during long-term sensitization conditioning that membrane structures may play a role in the effect of RAF of Abs to S100.16 S100 protein is a low-molecular weight protein (20 kDa), detected in the body of vertebrates, which has two calcium binding sites.17 S100 protein is involved in the regulation of such fundamental processes as generation and transmission of nerve impulses, protein phosphorylation, transcription factors, calcium homeostasis, and changes in cytoskeleton components, enzyme activity, cellular growth and differentiation, and in the development of inflammatory reaction.17 In the nervous system, S100 protein is involved in the tracking of synaptic vesicules, cellular survival, and differentiation.17–20
Experimentally and clinically, it was shown that RAF of Abs to S100 exerts unique psychotropic activity10 including anxiolytic, antiasthenic, antiaggressive, stress-protective, antihypoxic, antiischemic, neuroprotective, and nootropic effects. When evaluating the effects of RAF of Abs to S100 on rats, it was shown that blockage and stimulation of γ-aminobutyric acid (GABA) and serotoninergic receptors affected anxiolytic and antidepressant activity.21–23 The results from an in vivo study of anxiolytic and antidepressant activity of RAF of Abs to S100 have shown that preliminary ketanserin blockage of 5-HT2/1C serotonin receptors caused a reduction in intensity of the effects of RAF of Abs to S100. These data have proved the involvement of the serotoninergic system in the process, namely the involvement of 5-HT2/1C receptors in the realization of the effects of RAF of Abs to S100.21 Vogel conflict test results showed that GABAA receptor blockage induced by bicuculline and blockage of the chloride channel upon injection of picrotoxinin significantly reduced anti-conflict effect of RAF of Abs to S100.22 Additionally, simultaneous injection of GABAB receptor agonist baclofen with RAF of Abs to S100 reduced anxiolytic effect of the latter, while simultaneous injection of GABAB receptor antagonist phaclofen on the contrary enhanced anti-conflict effect of RAF of Abs to S100 providing further support for effecting this pathway.23
The purpose of this study was to evaluate the influence of RAF of Abs to S100 on major neurotransmitter systems which are possibly involved in its mechanism of pharmacological activity. For that purpose, we investigated the effect of RAF of Abs to S100 on dopaminergic, serotoninergic, and GABAergic receptors, different sites of N-methyl-d-aspartate (NMDA) receptors, as well as sigma1 and sigma2 receptors using radioligand binding and/or functional activity assays.
Methods
All the experiments were conducted using validated methods in accordance with standard operating procedures (SOPs) used by Eurofins Cerep (Paris, France) and Euroscreen S.A. (Brussels, Belgium). All cell membranes were provided by Eurofins Cerep (formerly Cerep) and Euroscreen S.A. All samples (positive controls, negative controls, and RAF of Abs to S100) were tested in the equal amount (v/v). Distilled water served as negative control for both RAF of Abs to S100 and placebo, while solvents or assay buffers - for positive controls. The number of replicas was in the range of two to six that corresponds to the number, which was ordinarily used in the similar in vitro experiments.24–26 Results analysis was performed as described in the respective chapters of the Assay Guidance Manual.27 The concentrations of all radioligands and nonspecific ligands used in radioligand binding assays were chosen in accordance with the results of the respective preliminary saturation binding assays and historical values determined at Eurofins Cerep or Euroscreen S.A.
The data from Eurofins Cerep and Euroscreen presented in the submitted manuscript was obtained from in vitro experiments that did not require approval from an ethical committee. Biological material originated from non-human cells expressing recombinant human proteins, immortalized cell lines and rodent tissues. In the latter case, rodents were bought from registered breeders providing us health monitoring reports and housed at Eurofins Cerep before tissue collection.
The Eurofins Cerep animal housing was approved by governmental institution in September 2011 for a 5 years period.
For tissue collection, Eurofins Cerep structure adheres to the external local ethics committee for animal experimentation, registered in December 13, 2012 by the Governmental Agency “Ministère de l’Enseignement Supérieur et de la Recherche” under the number 84. Euthanasia protocol was approved by Ethical Committee (COMETHEA PC 84) in December 2012.
Radioligand assays: 5-HT1F, 5-HT2B, 5-HT2Cedited, 5-HT3, and sigma1 receptors
Receptor binding assays for the human serotonin (5-hydroxytryptamine [5-HT]) receptors 5-HT1F, 5-HT2B, 5-HT2Cedited, and 5-HT3 were performed using membrane preparations from CHO-K1 cells (exception for 5-HT1F – Chinese hamster ovary [CHO] cells were used) expressing the respective human receptor. MCF-7 or Jurkat cells were used in the assays for human recombinant or native sigma1 receptors, respectively. Extracts of cell membranes were incubated with the appropriate radioligands in the presence or absence of RAF of Abs to S100 protein or placebo, diluted in assay buffer. Test compounds were added to the well at 50 μL (50%, v/v) for all 5-HT receptors and for recombinant sigma1 receptor, and at 20 μL (10%, v/v) or at the following range to generate a dose–response curve in the assay on the native sigma1 receptor: 20 μL, 10 μL, 4 μL, 2 μL, 1 μL, 0.4 μL, and 0.2 μL (placebo was tested at 20 μL or 10% [v/v] only). Protein concentration was 5 μg/well for 5-HT1F, 7 μg/well for 5-HT2B, 1 μg/well for 5-HT2Cedited, 2.5 μg/well for 5-HT3, 15 μg/well for recombinant sigma1, or 200 μg/well for native sigma1. The assay buffer containing 50 mM Tris (pH 7.4) and 0.1% ascorbic acid was supplied with the following: 4 mM CaCl2 for 5-HT1F, 4 mM CaCl2 and 10 μg/mL saponin for 5-HT2B, and 5 mM CaCl2 for 5-HT3. In case of the recombinant or native sigma1 receptors, 50 mM Tris (pH 8.0) or 50 mM Tris–HCl (pH 7.4) was used as the assay buffer. The following radioligands were used for each receptor binding assay: [3H]-LSD (7 nM) for 5-HT1F, [3H]-LSD (1.6 nM) for 5-HT2B, [3H]-mesulergin (1.85 nM) for 5-HT2Cedited, [3H]-BRL43694 (2.5 nM) for 5-HT3, and [3H](+)pentazocine for sigma1 receptors (61 nM or 8 nM for recombinant or native, respectively). Nonspecific binding was determined in the presence of a saturation concentration of the corresponding nonspecific ligand: BRL54443 (1 μM) for 5-HT1F, ritanserin (1 μM) for 5-HT2B, 5-HT (100 μM) for 5-HT2Cedited, MDL72222 (100 μM) for 5-HT3, and haloperidol for sigma1 receptors (5 μM or 10 μM for recombinant or native, respectively). All binding assays except those for recombinant sigma1 receptor were performed by a filtration method using Filtermate Harvester (PerkinElmer Inc., Waltham, MA, USA). Recombinant sigma1 binding assay was carried out using Filtermat A, Wallac. Filters were preliminary soaked in 0.5% polyethylenimine (PEI) for 5-HT receptors or in 0.1% and 0.3% PEI for recombinant or native sigma1 receptors, respectively.
For the rest of the receptors tested (for the human dopamine receptors D1, D2L, D2S, D4.4, D5; for the rat GABA receptors GABAA and the human GABAB(1b) receptors; for the rat glutamate receptors phencyclohexyl piperidine [PCP], α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid [AMPA], kainate, NMDA; for the human 5-HT receptors 5-HT1B, 5-HT1D [rat], 5-HT1E, 5-HT2A, 5-HT4e, 5-HT6, 5-HT7; and for the rat sigma2 receptors), Supplementary materials list the respective experimental conditions for the binding assays.
For radioligand binding experiments, the half maximal inhibitory concentration (IC50) and the half maximal effective concentration (EC50) values were determined (via computer software) by nonlinear regression analysis of the competition curves using Hill equation curve fitting. The inhibition constants (Ki) were calculated using the Cheng–Prusoff equation (Ki = IC50/(1+ (L/KD)), where L is the concentration of radioligand in the assay, and KD is the affinity of the radioligand for the receptor.
The results are expressed as a percent of control specific binding ([measured specific binding/control specific binding] ×100) and as a percent inhibition of control specific binding (100− [(measured specific binding/control specific binding) ×100]) obtained in the presence of the test compounds.
Functional assays: [35S]GTPγS binding assays on D3, GABAB1A/B2, 5-HT1A, and 5-HT1B receptors
Extracts of recombinant cell membranes of CHO expressing the respective human receptor were thawed on ice and diluted in assay buffer containing 20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES; pH 7.4), 100 mM NaCl, 10 μg/mL saponin, and 3 mM MgCl2 supplied with 0.1% bovine serum albumin for D3 and 5-HT1A receptors (for 5-HT1B, the following assay buffers were used: Krebs–Ringer HEPES, 1 mM 3-isobutyl-1-methylxanthine). They were kept on ice until assays were begun. Membranes were mixed with guanosine diphosphate (GDP) (v/v) diluted in assay buffer to reach the final concentration and incubated for at least 15 minutes on ice. In parallel, [35S]GTPγ (PerkinElmer, NEG030X) was diluted in assay buffer to reach a final concentration of 0.1 nM and was mixed with granules (PVT-WGA, Amersham, RPNQ0001) (v/v) directly prior to initiation of reaction.
The following compounds were successively added to the wells of an Optiplate (PerkinElmer) in order to determine agonistic activity: 50 μL of RAF of Abs to S100, or 50 μL placebo, or 50 μL of reference ligand (reference agonist – dopamine HCl for D3, SKF97541 for GABAB1A/B2, and 5-CT for 5-HT1A and for 5-HT1B – diluted in assay buffer in order to reach concentration from 0.01× EC50 to 100× EC50), 20 μL of membrane mixture with GDP, and 20 μL of mixture of [35S]GTPγ with beads.
The following compounds were successively added to the wells of an Optiplate (PerkinElmer) in order to determine antagonistic activity: 50 μL of RAF of Abs to S100, or 50 μL placebo, or 50 μL of reference ligand (reference antagonist – GR103691 for D3, CGP54626 for GABAB1A/B2, (S)-WAY100135 for 5-HT1A, and methiothepin for 5-HT1B – diluted in assay buffer in order to reach concentration from 0.01× IC50 to 100× IC50), 10 μL of assay buffer, and 20 μL of membrane mixture with GDP. After a 15-minute incubation, 10 μL of reference agonist (dopamine [3 nM] for D3 and SKF97541 [8 μM] for GABAB1A/B2) in concentration equal to historical EC80 and 20 μL of mixture of [35S]GTPγ with beads were added.
The plates were covered with a top seal, shaken on an orbital shaker for 2 minutes, and then incubated for 1 hour (for 5-HT1A and GABAB1A/B2 – 30-minute incubation) at 21°C. Then, the plates were centrifuged for 10 minutes at 2,000 rpm, incubated at 21°C for 1 hour for D3 or for 10 minutes for 5-HT1B (no incubation for 5-HT1A and GABAB1A/B2), and counted for 1 minute/well with a PerkinElmer TopCount™ reader.
For the rest of the receptors tested (for the human dopamine receptors D1, D2L, D2S, D4.4, D5; for the human 5-HT receptors 5-HT1D [rat], 5-HT1E, 5-HT2A, 5-HT2B, 5-HT2Cedited, 5-HT3, 5-HT4e, 5-HT6, 5-HT7), Supplementary materials list the respective experimental conditions for the functional assays.
For functional experiments, IC50 and EC50 values were determined (via computer software) by nonlinear regression analysis of the competition curves using Hill equation curve fitting. For the antagonists, the apparent dissociation constants (KB) were calculated using the modified Cheng–Prusoff equation (KB = IC50/(1+ (A/EC50A)), where A is the concentration of reference agonist in the assay, and EC50A is the EC50 value of the reference agonist.
Agonist activities of test compounds are expressed as a percentage of the activity of the reference agonist at its EC100 concentration. Antagonist activities of test compounds are expressed as a percentage of the inhibition of reference agonist activity at its EC80 concentration.
Test samples
The sample tested in this study was the active pharmaceutical ingredient of the commercial drug Tenoten® (OOO “NPF “MATERIA MEDICA HOLDING”, Moscow, Russian Federation), which corresponds to RAF of Abs to S100. RAF of Abs to S100 was manufactured in accordance with the technology for the respective commercial drug production and supplied by OOO “NPF “MATERIA MEDICA HOLDING” (Russian Federation) as a ready-to-use water solution. Technology is described previously in the US patent 8,535,66428 and meets applicable requirements of the European Pharmacopoeia (7th Edition, 2011). Briefly, RAF of Abs to S100 was manufactured by repeated consecutive dilution of the starting substance and external treatment during all technological steps. As a solvent, ethanol–water solution was used for intermediate dilutions, and purified water was used for preparation of the final dilution. All dilutions were prepared in glass vials in sterile conditions, avoiding direct intense light, and were stored at room temperature. Rabbit polyclonal antibodies to S100 protein (mixture of S100 protein isoforms used for the rabbit immunization), which had been concentrated approximately 2.5 mg/mL, were used as a starting substance (OOO “NPF “MATERIA MEDICA HOLDING”, Russian Federation). In the case of the placebo, the stock solution, which did not contain the rabbit polyclonal antibodies to S100 protein, was used as starting substance and underwent the same technological steps. The samples were coded by manufacturer and used blinded in the study. The remaining compounds and expendable materials used in the experiments were acquired.
Results
In all assays, the highest amount of RAF of Abs to S100, which could be added in the well in accordance with the SOPs used by Eurofins Cerep and Euroscreen S.A., was used. In the case of nonspecific effects of the test sample (eg, osmotic pressure cell bursts), the sample was added in the maximal possible amount, which did not cause the nonspecific effect. As a control, in parallel with the testing of RAF of Abs to S100 in each assay, vehicle (prepared using the similar dilution process) used for preparation of RAF of Abs to S100 was tested. In some cases (Tables 1 and 2) because of the high nonspecific effect of the vehicle, it was impossible to interpret the results obtained on RAF of Abs to S100, which were later considered as negative because of high nonspecific effect of vehicle. Only the results from the experiments which showed no nonspecific effect of the vehicle were analyzed.
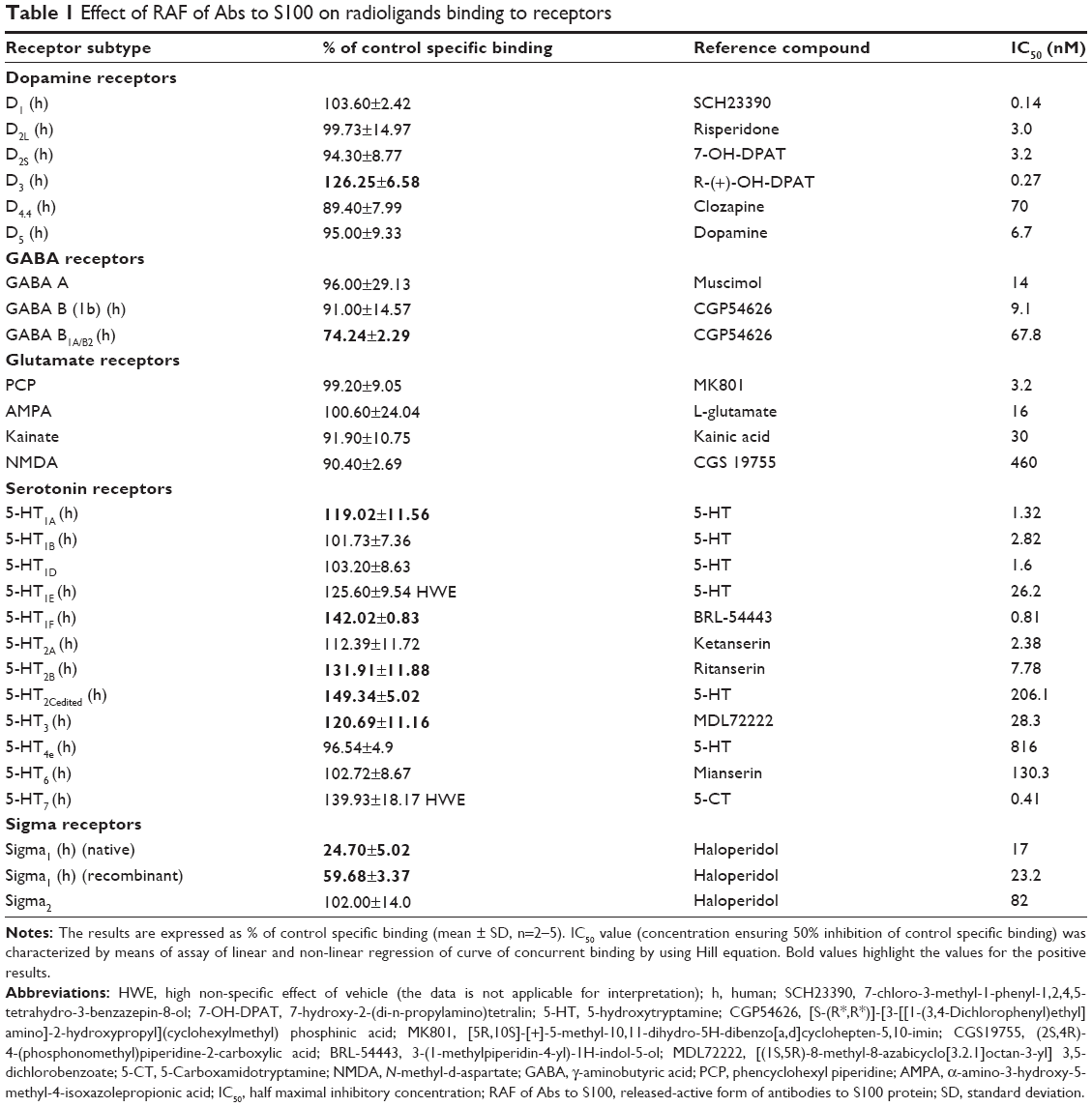 | Table 1 Effect of RAF of Abs to S100 on radioligands binding to receptors |
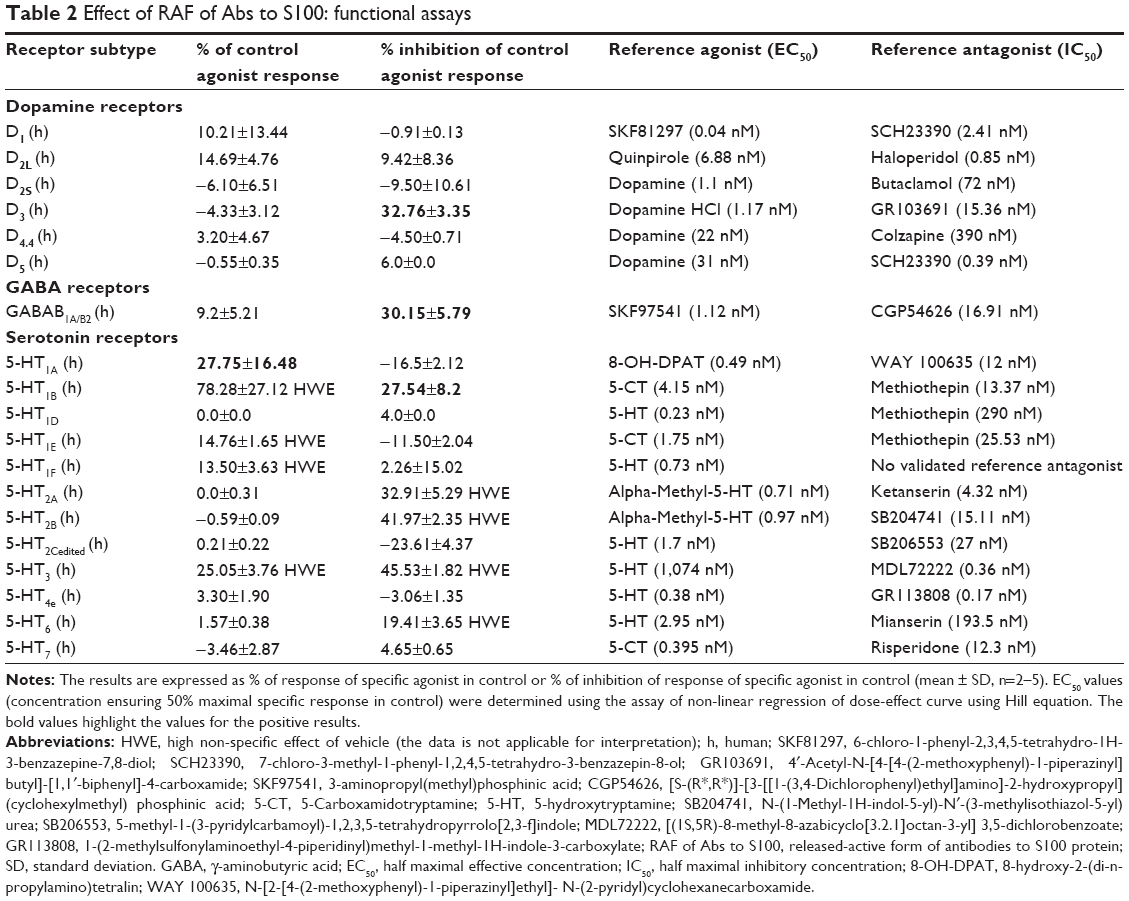 | Table 2 Effect of RAF of Abs to S100: functional assays |
Radioligand assays
RAF of Abs to S100 caused significant effect on the interaction of radioligands with seven receptors (Table 1). RAF of Abs to S100 increased radioligand binding to 5-HT1F, 5-HT2B, 5-HT2Cedited, and 5-HT3 receptors of serotoninergic system by 142.0%, 131.9%, 149.3%, and 120.7%, respectively. RAF of Abs to S100 enhanced radioligand binding to D3 receptors by 126.3% as well. The sample also inhibited specific binding of ([3,4-3H]-cyclohexylmethyl)phosphinic acid ([3H]-CGP54626) to GABAB1A/B2 receptors by 25.8%. RAF of Abs to S100 significantly inhibited specific binding of radioligand to both native and recombinant human sigma1 receptors, but the intensity of the drug effect was different. Thus, when analyzing the effect on ligand interaction with recombinant receptor, the effect of RAF of Abs to S100 was 40.32%, while with native receptor, the effect was 75.3% (Table 1). Testing of RAF of Abs to S100 in several concentrations made it possible to reveal the drug “dose–response” effect, which was manifested in different intensity of displacement of high-affinity ligand from its binding sites on native receptors (Figure 1). At the same time, placebo did not cause any effect on the ligand–receptor interaction. Under the experiments’ conditions, RAF of Abs to S100 did not cause significant effect on ligand–receptor interaction with the remaining receptors included in the screening.
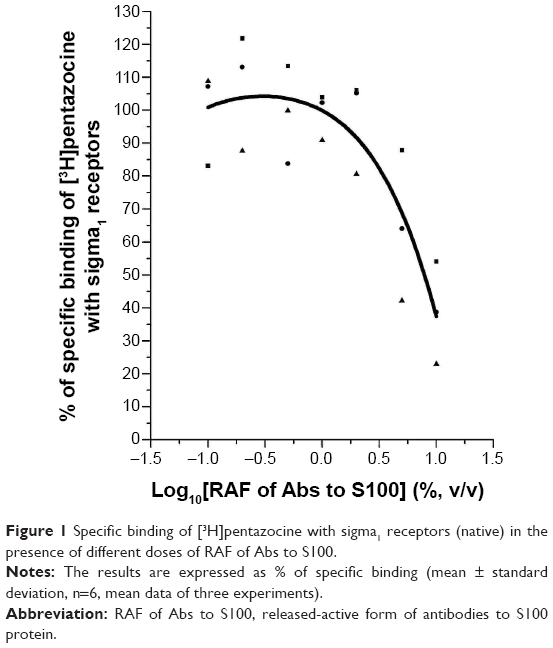 | Figure 1 Specific binding of [3H]pentazocine with sigma1 receptors (native) in the presence of different doses of RAF of Abs to S100. |
Functional assays
RAF of Abs to S100 exerted antagonist properties toward two receptors and agonist properties toward one receptor in the functional assays. It was shown that RAF of Abs to S100 exerted antagonism at 5-HT1B receptors, D3 receptors, and GABAB1A/B2 receptors inhibiting agonist-induced responses (respective agonist at EC80) by 23.24%, 32.76%, and 30.2%, respectively. And on the contrary, RAF of Abs to S100 exerted an agonist effect at 5-HT1A receptors enhancing receptor functional activity by 28% (Table 2). In the functional assays for the remaining tested receptors, RAF of Abs to S100 did not exert any effect. In some cases, for example, at GABAA, GABAB, glutamate receptors (PCP, AMPA, kainite, NMDA), and sigma1 and sigma2 receptors, the effect of RAF of Abs to S100 was not estimated due to lack of valid functional assays.
Discussion
The purpose of the current study was to determine the potential effects of RAF of Abs to S100 on the receptors of the major neurotransmitter systems which are possibly involved in its mechanism of pharmacological activity. The results provide direct evidence of drug influence on the serotonin, GABA, dopamine, and sigma receptors. These molecular interactions could explain the pharmaceutical action of RAF of Abs to S100 shown in previous experimental and clinical studies.10 With a wide range of pharmacological activities, together with the diversity of its clinical effects, it is difficult to understand the mechanism of its action. However, the deep interconnections between the various neurotransmitter systems, as well as the multireceptor nature of the mechanism of drug action, could be the basis of its diverse effects.
One of the major neurotransmitters in central nervous system, serotonin, mediates a wide range of physiological functions by interacting with 14 receptors grouped into seven families (from 5-HT1 to 5-HT7).29 In the present in vitro study, we have tested the effect of RAF of Abs to S100 on 12 5-HT receptors in both radioligand binding and functional assays. The data obtained in the cellular functional assays show that RAF of Abs to S100 exerted antagonist properties toward 5-HT1B and agonist properties toward 5-HT1A serotonin receptors. RAF of Abs to S100 stimulated radioligand binding with the 5-HT1F, 5-HT2B, 5-HT2Cedited, and 5-HT3 receptors, and it could be suggested to be an allosteric modulator of these receptors. This suggestion can be supported by earlier reports which showed that stimulated radioligand binding caused by positive allosteric modulator was typical.30,31 Additional studies would be worthwhile to evaluate the effect of RAF of Abs to S100 on radioligands affinity to the corresponding receptors in binding saturation assays as well as assays that evaluate receptor function under conditions appropriate for allosteric receptor modulation.
The obtained data are in agreement with previous in vivo studies which showed that RAF of Abs to S100 had anxiolytic activity in the Vogel conflict test, the elevated plus-maze test, and tail suspension test,32,33 and its antidepressant activity in forced swimming test can be reduced by preliminary ketanserin blockage of 5-HT2 receptors.21
In vitro data also clarify the molecular mechanisms of antidepressant and anxiolytic effects of RAF of Abs to S100. Activation of 5-HT1A receptors by agonists is associated with anxiolytic and antidepressant activity.34 Inhibition of 5-HT1B receptors by antagonists could be used for treatment of anxiety and depression.35,36 Stimulation of 5-HT2B receptors is reported to induce a serotonin-selective reuptake inhibitor-like response in behavioral and neurogenic assays37 as well as to possess anxiolytic effect in the Vogel conflict test in rats.38 5-HT2Cedited receptors modulate serotonin neurotransmission during stress, mediate anxiety in humans and animals,39 and are involved both in anxiolytic and anxiogenic effects of serotonin-selective reuptake inhibitors.40,41 The stress-induced increase in serotonin turnover and release can be inhibited by 5-HT2Cedited receptor activation.42
The anxiolytic effect of RAF of Abs to S100 is also mediated by GABAA receptors. Earlier in vivo studies demonstrated that blocking of GABAA receptor by bicuculline or its chloric channel by pyrotoxin significantly reduced anti-conflict effect of RAF of Abs to S100 in the Vogel conflict test.22 Additionally, according to unpublished data from an ex vivo study in mice, RAF of Abs to S100 significantly increased the number of benzodiazepine receptors expressed on the cellular membrane. In vitro evaluation of the drug effect on functional status of GABAA receptor was not conducted because of a lack of an appropriate method at the time of the initiation of the study.
Unlike benzodiazepine anxiolytics, RAF of Abs to S100 is also able to influence GABAB receptors in vivo23 and possesses properties of a GABAB1A/B2 receptor antagonist as well as a demonstrated ability to inhibit specific binding of radioligand with this receptor in the present study.
GABAB receptors are involved in the regulation of different central nervous system functions including anxiety, memory, and nociception.43 It is known that GABAB receptor antagonists possess antidepressant activity44 and are able to improve learning in rats.45
In vitro methods used in the present study revealed that RAF of Abs to S100 exerts its activity in binding assays only where RAF of Abs to S100 stimulated radioligand binding with the D3 receptors, whose ligands are known to have potential as antipsychotic and antiparkinson agents.46,47 Even though the received effect of RAF of Abs to S100 could be explained by an allosteric-like action, it should be demonstrated in additional experiments taking into account that the sample did not have any effect on functional status of the dopamine receptors used in the current study.
Taking into consideration the anti-cataleptic effect of RAF of Abs to S100, its effect on sigma receptors ensuring neuronal plasticity was analyzed. RAF of Abs to S100 reduced the number of radioligands specifically bound to the sigma1 receptors in a dose-dependent manner; however, it did not affect ligand–receptor interaction between sigma2 receptors and their radioligand. Additionally, according to unpublished data of an in vivo study, RAF of Abs to S100 under single dosing caused significant reduction in cataleptic activity of haloperidol, a ligand of the sigma1 receptor, on the Morpurgo’s model of catalepsy.
There are data proving that sigma1 receptors exert a modifying effect on all major neurotransmitter systems,48 including noradrenergic, serotoninergic, dopaminergic, and cholinergic system, and NMDA-regulated glutamate effects. Sigma1 receptors play an important role in the pathophysiology of neurodegenerative diseases (eg, Alzheimer disease, Parkinson disease), mental and affective disorders, and biological insult and are involved in the process of learning and memory. In this regard, the ability of RAF of Abs to S100 to affect the ligands’ interaction with sigma1 receptors might serve as one of the primary mechanisms for the realization of its pharmacological activity.
The key for understanding the possible mechanism of action on the receptor level is to demonstrate that S100 protein is the target for the RAF of Abs to S100. Thus, it was shown that RAF of Abs to S100 has similar effects as the S100 protein; direct action on membrane stabilization could modify S100 protein activity, and its effect has tight connections with membrane structures.12,14–16,49 Based on these observed effects, it could be assumed that RAF of Abs to S100 alters receptor binding and signaling through regulation of S100 cellular activities. S100 proteins regulate a diverse array of cellular activities, including membrane–cytoskeleton interactions, cytoskeleton dynamics,19 and membrane rearrangement,20 could affect the organization of lipid domains (rafts),50 and effect the modulation of the receptors function.18,51 The plasma membrane is a dynamic entity. It consists of a heterogeneous but regulated environment and includes discrete membrane domains that form through a combination of lipid and protein interactions.52 Highly ordered structures within the membrane that are rich in saturated lipids and scaffolding proteins are termed lipid rafts.53 Lipid rafts along with adaptor proteins regulate monoamine receptors, in particular serotonin and dopamine receptors.53 It was shown that lipid rafts can alter the conformation of the G-protein-coupled receptors, in particular acetylcholine and serotonin receptors, and thus can regulate neurotransmitter binding and signal transducing functions.53–55 In addition, lipid rafts also promote the function of GABAB receptors – lipid rafts affect cellular processes such as activity-dependent desensitization and receptor trafficking.56 Finally, the sigma1 receptor is enriched with lipid raft markers, and their functions are tightly connected.57 Directly, S100 as an adaptor protein has been found to interact with serotonin and dopamine receptors, causing an increased localization of the 5-HT1B and 5-HT4 receptors to the plasma membrane,19 modulates serotonin neurotransmission,58 and enhances D2 receptor signaling.59 Based on the current work, the exact mechanism should continue to be explored.
Conclusion
The received results provide with the first insight on the mechanism of the drug action on the receptor level. The pharmacological profiling of RAF of Abs to S100 and 27 receptor targets provides the direct evidence of a drug-related modification of the major neurotransmitter systems (serotoninergic, GABAergic, dopaminergic, and sigma1 receptors as well). Our results indicate that the effect of RAF of Abs to S100 on serotoninergic and dopaminergic receptors might have an allosteric nature which should be confirmed with further studies. Finding opportunities for optimization of in vitro techniques to overcome the limitations associated with the testing of water solutions will allow extending the understanding of mechanism of the drug action. Also, the demonstration of “in vitro–in vivo” correlation through further experiments including in vitro, in vivo, and ex vivo approaches should be done in order to confirm the connection between preclinical and clinical effects, which had been shown previously, with the receptor targets revealed within current work.
Acknowledgments
The authors thank members of Eurofins Cerep (France) and Euroscreen S.A. (Belgium) for carrying out the experiments, which were conducted using validated methods in accordance with the respective SOPs of the companies.
This work was funded by OOO “NPF “MATERIA MEDICA HOLDING”. All authors have an affiliation to the commercial funders of this research study.
Disclosure
Evgeniy A Gorbunov, Irina A Ertuzun, Evgeniya V Kachaeva, and Sergey A Tarasov are employees of the company, and Oleg I Epstein owns the company. The authors report no other conflicts of interest in this work.
References
Epstein OI. Release-activity: a long way from phenomenon to new drugs. Bull Exp Biol Med. 2012;154(1):54–58. | ||
Epstein OI. [The phenomenon of release activity and the hypothesis of “spatial” homeostasis]. Usp Fiziol Nauk. 2013; 44(3):54–76. Russian. | ||
Nicoll J, Gorbunov EA, Tarasov SA, Epstein OI. Subetta treatment increases adiponectin secretion by mature human adipocytes in vitro. Int J Endocrinol. 2013;2013:925874. | ||
Bailbé D, Philippe E, Gorbunov E, Tarasov S, Epstein O, Portha B. The novel oral drug Subetta exerts an antidiabetic effect in the diabetic Goto-Kakizaki rat: comparison with rosiglitazone. J Diabetes Res. 2013;2013:763125. | ||
Sakat SS, Mani K, Demidchenko YO, et al. Release-active dilutions of diclofenac enhance anti-inflammatory effect of diclofenac in carrageenan-induced rat paw edema model. Inflammation. 2014; 37(1):1–9. | ||
Sizova LV. Treatment of early arthritis using arthrofoon (ultra-low doses of antibodies to tumor necrosis factor-α). Indian J Pharmacol. 2011;43(6):724–725. | ||
Tarasov SA, Zarubaev VV, Gorbunov EA, Sergeeva SA, Epstein OI. Activity of ultra-low doses of antibodies to gamma-interferon against lethal influenza A(H1N1)2009 virus infection in mice. Antiviral Res. 2012;93(2):219–224. | ||
Chu X, Agmo A. Sexual incentive motivation in old male rats: the effects of sildenafil and a compound (Impaza) stimulating endothelial NO synthase. Pharmacol Biochem Behav. 2008;89(2):209–217. | ||
Dugina JL, Petrov VI, Babayeva AR, et al. A randomized, open-label, comparative, 6-month trial of oral ultra-low doses of antibodies to tumor necrosis factor-alpha and diclofenac in rheumatoid arthritis. Int J Tissue React. 2005;27(1):15–21. | ||
Epstein OI. Ultra-Low Doses (History of One Study). Moscow: Publishing House of RAMN; 2009. | ||
Karpova GV, Fomina TI, Vetoshkina TV, et al. Preclinical studied of general toxic properties of preparations containing ultralow doses of antibodies to endogenous regulators. Bull Exp Biol Med. 2009;148(3):543–546. | ||
Epshteĭn OI, Beregovoĭ NA, Sorokina NS, Starostina MV, Shtark MB. [Effect of various dilutions of the potentiated antibodies to the brain-specific protein S-100 on the post-titanic potentiation in surviving hippocampal slices]. Biull Eksp Biol Med. 1999;127(3):317–320. Russian. | ||
Kubista H, Donato R, Hermann A. S100 calcium binding protein affects neuronal electrical discharge activity by modulation of potassium currents. Neuroscience. 1999;90(2):493–508. | ||
Epshteĭn OI, Gaĭnutdinov KL, Shtark MB. [Effect of homeopathic doses of antibodies to antigen S-100 on the electric parameters of neuronal membranes]. Biull Eksp Biol Med. 1999;127(4):466–467. Russian. | ||
Epshtein OI, Gainutdinov KL, Shtark MB. Effect of homeopathic doses of antibodies to antigen S-100 on the electric parameters of neuronal membranes. Bull Exp Biol Med. 2003;127:111–113. | ||
Andrianov VV, Epstein OI, Gainutdinova TKH, Shtark MB, Timoshenko AKH, Gainutdinov KL. Antibodies to calcium-binding S100B protein block the conditioning of long-term sensitization in the terrestrial snail. Pharmacol Biochem Behav. 2009;94(1):37–42. | ||
Donato R. Intracellular and extracellular roles of S100 proteins. Microsc Res Tech. 2003;60(6):540–551. | ||
Donato R, Cannon BR, Sorci G, et al. Functions of S100 proteins. Curr Mol Med. 2013;13(1):24–57. | ||
Hermann A, Donato R, Weiger TM, Chazin WJ. S100 calcium binding proteins and ion channels. Front Pharmacol. 2012;3:67. | ||
Santamaria-Kisiel L, Rintala-Dempsey AC, Shaw GS. Calcium-dependent and -independent interactions of the S100 protein family. Biochem J. 2006;396(2):201–214. | ||
Kheifets IA, Dugina YL, Voronina TA, et al. Involvement of the serotoninergic system in the mechanism of action of ultralow dose antibodies to S-100 protein. Bull Exp Biol Med. 2007;143(5):598–600. | ||
Voronina TA, Molodavkin GM, Sergeeva SA, Epstein OI. GABAergic system in the anxiolytic effect of proproten: experimental study. Bull Exp Biol Med. 2003;135(suppl 7):125–127. | ||
Kheifez IA, Molodavkin GM, Voronina TA, Dugina YL, Sergeeva SA, Epstein OI. Involvement of the GABA-B system in the mechanism of action of ultralow-dose antibodies to S-100 protein. Bull Exp Biol Med. 2008;145(5):614–616. | ||
Ito H, Kiso T, Miyata K, et al. Pharmacological profile of YM-31636, a novel 5-HT3 receptor agonist, in vitro. Eur J Pharmacol. 2000;409(2):195–201. | ||
Mikami T, Komada T, Sugimoto H, et al. In vitro and in vivo pharmacological characterization of PF-01354082, a novel partial agonist selective for the 5-HT(4) receptor. Eur J Pharmacol. 2009;609(1–3):5–12. | ||
Zhang Y, Gilliam A, Maitra R, et al. Synthesis and biological evaluation of bivalent ligands for the cannabinoid 1 receptor. J Med Chem. 2010;53(19):7048–7060. | ||
Sittampalam G, Coussens N, Nelson H, et al. Assay Guidance Manual. Bethesda, MD: Eli Lilly & Company and the National Center for Advancing Translational Sciences; 2004. Available from: http://www.ncbi.nlm.nih.gov/books/NBK53196/. Accessed May 4th, 2012. | ||
Epstein OI, Shtark MB, Kolyadko TM, inventors; Oleg I. Epshtein, assignee. Method of treating a pathological syndrome and a pharmaceutical agent. United States patent US 8535664. 2013 Sep 17. | ||
Hayesa DJ, Greenshawa AJ. 5-HT receptors and reward-related behavior: a review. Neurosci Biobehav Rev. 2011;35(6):1419–1449. | ||
Im WB, Chio CL, Alberts GL, Dinh DM. Positive allosteric modulator of the human 5-HT2C receptor. Mol Pharmacol. 2003;64(1):78–84. | ||
de Oliveira-Pierce AN, Zhang R, Machu TK. Colchicine: a novel positive allosteric modulator of the human 5-hydroxytryptamine3A receptor. J Pharmacol Exp Ther. 2009;329(2):838–847. | ||
Castagne V, Lemaire M, Kheyfets I, Dugina JL, Sergeeva SA, Epstein OI. Antibodies to S100 proteins have anxiolytic-like activity at ultra-low doses in the adult rat. J Pharm Pharmacol. 2008;60(3):309–316. | ||
Loskutova LV, Shtark MB, Epstein OI. Efficiency of ultralow doses of antibodies to S100 protein and delta sleep-inducing peptide in rats with anxious depression. Bull Exp Biol Med. 2003;135(suppl 7):20–22. | ||
Banerjee P, Mehta M, Kanjilal B. The 5-HT1A receptor: a signaling hub linked to emotional balance. In: Chattopadhyay A, editor. Serotonin Receptors in Neurobiology. (Chap. 7). Boca Raton, FL: CRC Press; 2007. Available from: http://www.ncbi.nlm.nih.gov/books/NBK5212/ | ||
Moret C, Briley M. The possible role of 5-HT(1B/D) receptors in psychiatric disorders and their potential as a target for therapy. Eur J Pharmacol. 2000;404(1–2):1–12. | ||
Hudzik TJ, Yanek M, Porrey T, et al. Behavioral pharmacology of AR-A000002, a novel, selective 5-hydroxytryptamine(1B) antagonist. J Pharmacol Exp Ther. 2003;304(3):1072–1084. | ||
Diaz SL, Doly S, Narboux-Nême N, et al. 5-HT(2B) receptors are required for serotonin-selective antidepressant actions. Mol Psychiatry. 2012;17(2):154–163. | ||
Kennett GA, Trail B, Bright F. Anxiolytic-like actions of BW 723C86 in the rat Vogel conflict test are 5-HT2B receptor mediated. Neuropharmacology. 1998;37(12):1603–1610. | ||
Graeff FG. Serotonin, the periaqueductal gray and panic. Neurosci Biobehav Rev. 2004;28(3):239–259. | ||
Bristow LJ, O’Connor D, Watts R, Duxon MS, Hutson PH. Evidence for accelerated desensitisation of 5-HT(2C) receptors following combined treatment with fluoxetine and the 5-HT(1A) receptor antagonist, WAY 100,635, in the rat. Neuropharmacology. 2000;39(7):1222–1236. | ||
Bagdy G, Graf M, Anheuer ZE, Modos EA, Kantor S. Anxiety-like effects induced by acute fluoxetine, sertraline or m-CPP treatment are reversed by pretreatment with the 5-HT2C receptor antagonist SB-242084 but not the 5-HT1A receptor antagonist WAY-100635. Int J Neuropsychopharmacol. 2001;4(4):399–408. | ||
Mongeau R, Martin CB, Chevarin C, et al. 5-HT2C receptor activation prevents stress-induced enhancement of brain 5-HT turnover and extracellular levels in the mouse brain: modulation by chronic paroxetine treatment. J Neurochem. 2010;115(2):438–449. | ||
Varani AP, Balerio GN. GABA(B) receptors involvement in the effects induced by nicotine on anxiety-related behaviour in mice. Pharmacol Res. 2012;65(5):507–513. | ||
Nowak G, Partyka A, Pałucha A, et al. Antidepressant-like activity of CGP 36742 and CGP 51176, selective GABAB receptor antagonists, in rodents. Br J Pharmacol. 2006;149(5):581–590. | ||
Lasarge CL, Bañuelos C, Mayse JD, Bizon JL. Blockade of GABA(B) receptors completely reverses age-related learning impairment. Neuroscience. 2009;164(3):941–947. | ||
Joyce JN, Millan MJ. Dopamine D3 receptor antagonists as therapeutic agents. Drug Discov Today. 2005;10(13):917–925. | ||
Tadori Y, Forbes RA, McQuade RD, Kikuchi T. Functional potencies of dopamine agonists and antagonists at human dopamine D2 and D3 receptors. Eur J Pharmacol. 2011;666(1–3):43–52. | ||
Matsumoto RR, Bowen WD, Su T-P. Sigma Receptors, Chemistry, Cell Biology and Clinical Implication. Springer Science + Business Media, LLC: New York (NY), USA; 2007. | ||
Andrianov VV, Gainutdinov KHL, Gainutdinova TKH, Mukhamedshina DI, Shtark MB. Membranotropic effects of antibodies to S100 protein in ultralow doses. Bull Exp Biol Med. 2003;1:114–116. | ||
Drücker P, Pejic M, Galla HJ, Gerke V. Lipid segregation and membrane budding induced by the peripheral membrane binding protein annexin A2. J Biol Chem. 2013;288(34):24764–24776. | ||
Roux BT, Cottrell GS. G protein-coupled receptors: what a difference a ‘partner’ makes. Int J Mol Sci. 2014;15(1):1112–1142. | ||
Chichili GR, Rodgers W. Cytoskeleton-membrane interactions in membrane raft structure. Cell Mol Life Sci. 2009;66(14):2319–2328. | ||
Björk K, Svenningsson P. Modulation of monoamine receptors by adaptor proteins and lipid rafts: role in some effects of centrally acting drugs and therapeutic agents. Annu Rev Pharmacol Toxicol. 2011;51:211–242. | ||
Fantini J, Barrantes FJ. Sphingolipid/cholesterol regulation of neurotransmitter receptor conformation and function. Biochim Biophys Acta. 2009;1788(11):2345–2361. | ||
Preininger AM, Meiler J, Hamm HE. Conformational flexibility and structural dynamics in GPCR-mediated G protein activation: a perspective. J Mol Biol. 2013;425(13):2288–2298. | ||
Padgett CL, Slesinger PA. GABAB receptor coupling to G-proteins and ion channels. Adv Pharmacol. 2010;58:123–147. | ||
Li J, Liu B, Gao X, et al. Overexpression of sigma-1 receptor inhibits ADAM10 and ADAM17 mediated shedding in vitro. Protein Cell. 2012;3(2):153–159. | ||
Egeland M, Warner-Schmidt J, Greengard P, Svenningsson P. Co-expression of serotonin 5-HT(1B) and 5-HT(4) receptors in p11 containing cells in cerebral cortex, hippocampus, caudate-putamen and cerebellum. Neuropharmacology. 2011;61(3):442–450. | ||
Liu Y, Buck DC, Neve KA. Novel interaction of the dopamine D2 receptor and the Ca2+ binding protein S100B: role in D2 receptor function. Mol Pharmacol. 2008;74(2):371–378. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.