Back to Journals » Infection and Drug Resistance » Volume 10
In vitro activity of colistin mono- and combination therapy against colistin-resistant Acinetobacter baumannii, mechanism of resistance, and clinical outcomes of patients infected with colistin-resistant A. baumannii at a Thai university hospital
Authors Lertsrisatit Y ![]() , Santimaleeworagun W, Thunyaharn S, Traipattanakul J
, Santimaleeworagun W, Thunyaharn S, Traipattanakul J
Received 3 August 2017
Accepted for publication 18 September 2017
Published 20 November 2017 Volume 2017:10 Pages 437—443
DOI https://doi.org/10.2147/IDR.S148185
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Yongyut Lertsrisatit,1 Wichai Santimaleeworagun,2,3 Sudaluck Thunyaharn,4 Jantima Traipattanakul5
1College of Pharmacotherapy Thailand, Nonthaburi, Thailand; 2Department of Pharmacy, Faculty of Pharmacy, Silpakorn University, Nakorn Pathom, Thailand; 3Pharmaceutical Initiative for Resistant Bacteria and Infectious Diseases Working Group (PIRBIG) Faculty of Pharmacy, Silpakorn University, Nakorn Pathom, Thailand; 4Faculty of Medical Technology, Nakhonratchasima College, Nakhonratchasima, Thailand; 5Division of Infectious Disease, Department of Medicine, Phramongkutklao Hospital, Bangkok, Thailand
Purpose: Colistin is a drug of last resort for treating multidrug-resistant Acinetobacter baumannii infections. Unfortunately, colistin-resistant A. baumannii (CoR-AB) has been reported. Here, we examined the in vitro effect of mono- and combined antimicrobials against CoR-AB strains and their resistance mechanism, and evaluated the clinical outcomes of CoR-AB-infected patients.
Patients and methods: Seventeen clinical CoR-AB strains were isolated from patients at Phramongkutklao hospital, 2011–2015. The mono- and synergistic activities of colistin, tigecycline, sulbactam, imipenem, meropenem, amikacin, fosfomycin, and cotrimoxazole were examined by minimum inhibitory concentration (MIC) and fractional inhibitory concentration index. Clonal relationship and resistance genes were determined by repetitive extragenic palindromic polymerase chain reaction with specific primers. The effect of carbonyl cyanide 3-chlorophenylhydrazone combined with colistin was used to test efflux pump involvement. Patient treatment outcomes were also reported.
Results: The most prevalent infection in CoR-AB patients was pneumonia (35.3%), and all patients were administered colistin combined with another agent. The 30-day mortality was 70.6%, and the colistin MIC range and MIC50 was 16–512 µg/mL and 64 µg/mL, respectively. All CoR-AB strains were sensitive to tigecycline. Sporadic isolates were susceptible to sulbactam, imipenem, meropenem, and cotrimoxazole. A synergistic or additive effect was observed for colistin plus imipenem or meropenem (16.7%), sulbactam (66.7%), or tigecycline (66.7%). The CoR-AB isolates could be divided into four different clones (A–D) with a high prevalence of group B (47.1%). Eight isolates harbored blaOXA23, blaIMP, blaKPC, and blaNDM, and one contained blaOXA23, blaIMP, and blaKPC, while the eight remaining isolates carried only blaOXA23. The MIC values of all strains were greatly reduced for colistin plus carbonyl cyanide 3-chlorophenylhydrazone.
Conclusion: CoR-AB clinical isolates exhibited very high colistin resistance and a high frequency of resistance genes. The mechanism of colistin resistance appears to be mediated via an efflux pump. Thus, certain antimicrobials could be used as salvage therapy for CoR-AB infection.
Keywords: synergism, salvage therapy, clinical outcomes
Introduction
Acinetobacter baumannii, a gram-negative coccobacilli, is a pathogen causing major nosocomial infection, leading to a high level of unfavorable outcomes. Data from 49 US hospitals collected from 1995 through 2002 showed that A. baumannii bacteremia was the third highest cause of death in patients with bloodstream infections in intensive care units.1 In Thailand, A. baumannii is the most common causative pathogen of nosocomial pneumonia in tertiary care hospitals2 and the most abundant bacterium isolated from sputum cultures in a nationwide survey conducted in 55 hospitals in 2016.3
A. baumannii has been developing increasing resistance to a number of available antibiotics. Currently, the World Health Organization has defined carbapenem-resistant A. baumannii as one of the global priority superbugs.4 Data from National Antimicrobial Resistance Surveillance Thailand in 2016 have shown that ~70% of the clinical A. baumannii strains were resistant to carbapenems.3 However, no new antimicrobials are available for the treatment of carbapenem-resistant A. baumannii; thus, colistin and polymyxin B constitute the last active options in an era of multidrug-resistant A. baumannii.5
Colistin was initially used during the 1950s, but was therapeutically phased out in the 1980s because of increased nephrotoxicity. However, colistin is the most effective antibiotic against multidrug-resistant gram-negative bacilli and is therefore widely used nowadays as a salvage therapy.6 Unfortunately, colistin-resistant A. baumannii (CoR-AB) emerged in 2006 as a subpopulation of the heteroresistant strains with a minimal inhibitory concentration (MIC) ≤2 µg/mL.7 CoR-AB was first reported in the Czech republic.8 Data from a SENTRY study conducted across the USA, Europe, and Asia showed that CoR-AB prevalence was ~0.9%–3.3%,9 with levels as high as 30.6% in Korea.10
The mechanism of action of colistin resistance involves the disturbance of lipid A, a component of bacterial outer membrane lipopolysaccharide. The reduced negative charge of lipid A or loss of lipid A has been hypothesized as the cause of CoR-AB. Additionally, efflux pumps, proteinaceous transporters localized in the bacterial membrane, have also been shown to significantly contribute to colistin resistance. However, to date, there are no reports of mcr-1 in CoR-AB.6,11
To date, data regarding salvage therapy for CoR-AB infections and the clinical outcomes of patients infected with CoR-AB are limited. In addition, CoR-AB has been reported sporadically at our institute and is thus considered to be an urgent problem. Therefore, this study aimed to identify in vitro the mono- and combined antimicrobials against CoR-AB strains and their mechanism of resistance and to evaluate the clinical outcomes of patients infected with CoR-AB.
Patients and methods
All clinical strains and patient data were obtained from patients infected with CoR-AB admitted to Phramongkutklao Hospital, a 1,200-bed university hospital in Bangkok, Thailand, from January 2011 to December 2015. CoR-AB was defined as A. baumannii with a colistin MIC >2 µg/mL as determined using the broth microdilution method according to the Clinical and Laboratory Standards Institute guidelines, 2017.12
Ethical approval
As this study was conducted using archived bacterial isolates and involved a retrospective review of clinical outcomes, the study was given an exemption from having to obtain written informed consent from the patients by the ethical review committee of the Royal Thai Army Medical Department (approval no. Q016h/59). Following ethical approval, permission was obtained from the Director of the Phramongkutklao Hospital prior to data collection. To ensure confidentiality of patient information, anonymous typing was used and the data were maintained in a confidential manner.
Mono- and combination antimicrobial activity
The in vitro mono-activities of tigecycline, sulbactam, imipenem, meropenem, amikacin, and cotrimoxazole were evaluated as MIC using the E-test (Liofilchem, Teramo, Italy). Colistin (Sigma-Aldrich, St Louis, MO, USA; 12 twofold dilutions [0.25–512 µg/mL]) and fosfomycin (Wago Pure Chemical Industry, Osaka, Japan; eight concentrations [8–1,024 µg/mL]) were evaluated using the broth microdilution and agar dilution supplemented with 25 µg/mL of glucose-6-phosphate (Sigma-Aldrich) as the cofactor for fosfomycin transportation into the bacterial cell, respectively. Escherichia coli ATCC 25922 was used as the quality control strain based on Clinical and Laboratory Standards Institute guidelines.12
Checkerboard synergy testing was performed in a sterile 96-well U-bottom plate. Colistin combined with sulbactam, tigecycline, imipenem, meropenem, or cotrimoxazole (Wago Pure Chemical Industry) was examined. Each row well was inoculated with a twofold dilution of colistin (8–1,024 µg/mL), and then serial dilutions (0.25–512 µg/mL, except for tigecycline and cotrimoxazole 0.063–128 µg/mL) of the other antibacterials were added to each column well. The plates were incubated at 37°C for 18 h. MIC was defined as the lowest concentration that resulted in a clear well. Next, the fractional inhibitory concentration index (ΣFICI) of each combination was calculated; ΣFICI values ≤0.5, >0.5–1.0, >1.0–4.0, and >4.0 were interpreted as a synergistic, additive, indifferent, and antagonistic effect, respectively.
Molecular typing and detection of resistance genes
The genomic DNA of the CoR-AB strains was extracted using a commercial kit (Thermo Fisher Scientific, Waltham, MA, USA). The clonal relationship of the CoR-AB strains was evaluated using the repetitive extragenic palindromic polymerase chain reaction (REP-PCR) method. The PCR reaction mixture was set up as previously described using the primer set REP-forward: 5′-IIIGCforwardATCAGGC-3′ and REP-reverse: 5′-ACGTCTTATCAGGCCTAC-3′ to amplify the REP region.13 Thermocycling was conducted with a Biometra TGradient Thermocycler (Biometra, Gottingen, Germany). The REP-PCR products were visualized by electrophoresis on 1.2% agarose gels (Amresco, Solon, OH, USA) stained with ethidium bromide (Bio Basic, Markham, Ontario, Canada). Patterns that differed by more than three bands were classified as different clones.14
The primers, PCR reaction mixture, and conditions used for the detection of carbapenemase resistance genes blaOXA-23 (822 bp), blaOXA-40 (828 bp), and blaOXA-58 (843 bp) were as previously described.13 blaIMP (232 bp), blaVIM (390 bp), blaKPC (798 bp), and blaNDM (621 bp) were detected by multiplex PCR, as previously described.15 The PCR mixture, specific primers, and PCR conditions for mcr-1 (309 bp) detection were as previously described.16 All PCR products were separated by electrophoresis on 1% agarose gels (Amresco) stained with ethidium bromide (Bio Basic) and compared with the bands of known carbapenemase and mcr-1 genes (positive controls) along with the 1 kb plus DNA ladder (Thermo Fisher Scientific). PCR-grade water was used as a DNA-free negative control.
Detection of efflux pump mechanism
Efflux pumps were evaluated using the microdilution method.12,17 Mueller Hinton II broth (Beckton Dickinson Company, Baltimore, MD, USA) with and without carbonyl cyanide 3-chlorophenylhydrazone (CCCP; Sigma-Aldrich) was used as the efflux pump inhibitor (EPI). CCCP was added in twofold dilutions to a colistin 96-well plate at a final concentration of 20 µg/mL. The phenotypic determination of efflux pump activity was defined as a decreased MIC for colistin plus CCCP in more than fourfold dilution compared to the MIC of colistin alone.
Clinical outcomes
All patients with CoR-AB infection were reviewed for clinical information including age, sex, underlying diseases, admitting ward, mechanical ventilator use, shock, hepatic function, renal function, immunocompromised status, antimicrobial regimens, antimicrobial susceptibility of CoR-AB, source of infections, and clinical outcomes. Clinical outcomes were defined as follows: cure – clinical improvement and culture negative following treatment; clinical improvement – normal body temperature, normal level of white blood cells, and stable vital signs without microbiologic confirmation of a cure; treatment failure – worsening clinical symptoms or the requirement for different or additional antimicrobial therapy against CoR-AB; and death – in-hospital death following CoR-AB infection.
Results
Antimicrobial susceptibility
Seventeen cases infected with CoR-AB were identified over a 5-year period. The colistin MIC range and MIC50 of the 17 strains were 16–512 µg/mL and 64 µg/mL, respectively. The MIC distribution of sulbactam, tigecycline, imipenem, meropenem, trimethoprim–sulfamethoxazole, amikacin, and fosfomycin of the CoR-AB strains is detailed in Table 1.
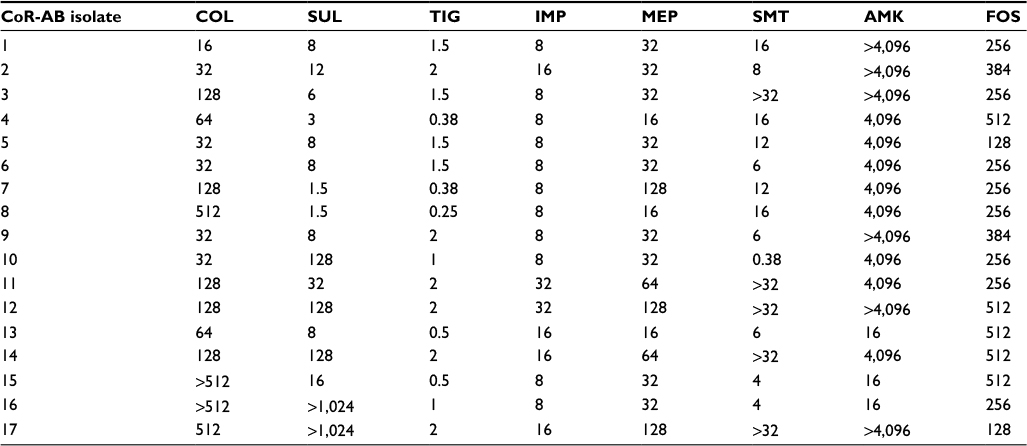 | Table 1 Antibiotic minimum inhibitory concentration (µg/mL) in colistin-resistant Acinetobacter baumannii Abbreviations: AMK, amikacin; COL, colistin; CoR-AB, colistin-resistant Acinetobacter baumannii; FOS, fosfomycin; IMP, imipenem; MEP, meropenem; SMT, trimethoprim–sulfamethoxazole; SUL, sulbactam; TIG, tigecycline. |
A synergistic or additive effect was observed for colistin and imipenem or meropenem in 1/6 isolates (16.7%) and for sulbactam and tigecycline in 4/6 isolates (66.7%). No antagonistic effects were observed for any of the antimicrobial agent combinations.
Molecular typing, resistance gene(s), and efflux pump detection
REP-PCR profiles revealed that the CoR-AB isolates could be classified into four clones, A–D. Most strains (n=8) belonged to clone B, followed by clone C (n=7), clone A (n=1), and clone D (n=1), as shown in Table 2.
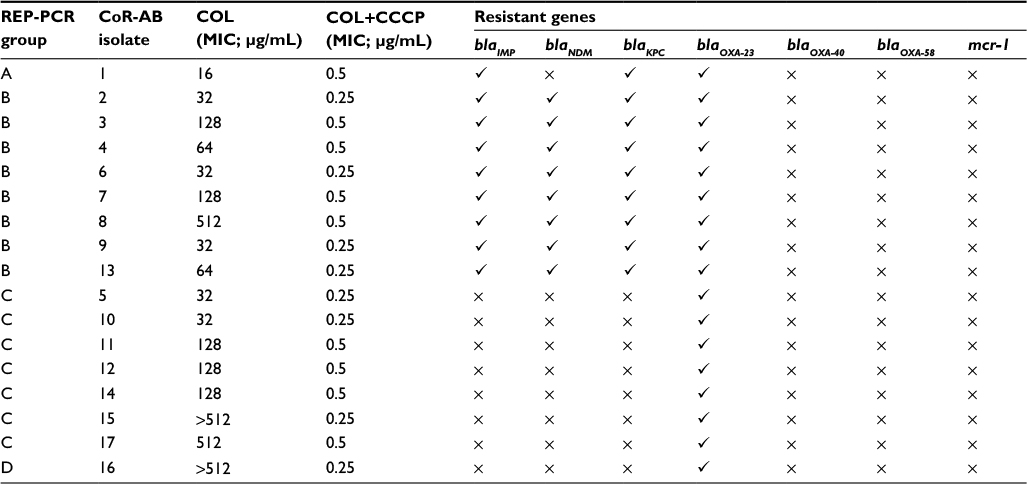 | Table 2 Clonal relationship, presence of carbapenemase genes and mcr-1, and efflux pump detection in colistin-resistant Acinetobacter baumannii Notes: Target gene present. ×Target gene absent. Abbreviations: CCCP, cyanide 3-chlorophenylhydrazone; COL, colistin; CoR-AB, colistin-resistant Acinetobacter baumannii; MIC, minimum inhibitory concentration; REP-PCR, repetitive element palindromic polymerase chain reaction. |
Carbapenemase genes were detected by PCR; all 17 CoR-AB isolates carried at least one gene (Table 2). Importantly, a number of CoR-AB isolates harbored more than one carbapenemase gene: eight isolates contained blaOXA23, blaIMP, blaKPC, and blaNDM, and one contained blaOXA23, blaIMP, and blaKPC, while the eight remaining isolates carried only blaOXA23. Interestingly, all of the isolates harboring blaOXA23, blaIMP, blaKPC, and blaNDM (n=8) and blaOXA23, blaIMP, and blaKPC (n=1) belonged to clones B and A, respectively, whereas the isolates carrying blaOXA23 belonged to clones C and D (Table 2).
The results of the efflux pump assay demonstrated that the colistin MIC values of the 17 CoR-AB isolates were greatly reduced (>32-fold) when combined with CCCP (Table 2).
Clinical outcomes
Of these 17 patients with CoR-AB infections, 11 patients (64.7%) were male, the median age was 77 years (interquartile range [IQR]: 62–82.5), and 11 patients (64.7%) had been admitted to the intensive care unit. The three highest ranking sources of infection were pneumonia (n=6; 35.3%), septicemia of unknown origin (n=5; 29.4%), and cholangitis (n=3; 17.6%), respectively (Table 3). All CoR-AB cases received combination antibacterial treatment, which mostly contained colistin as the backbone agent. The 30-day mortality, clinical improvement, and cure rates were 70.6%, 17.6%, and 11.8%, respectively.
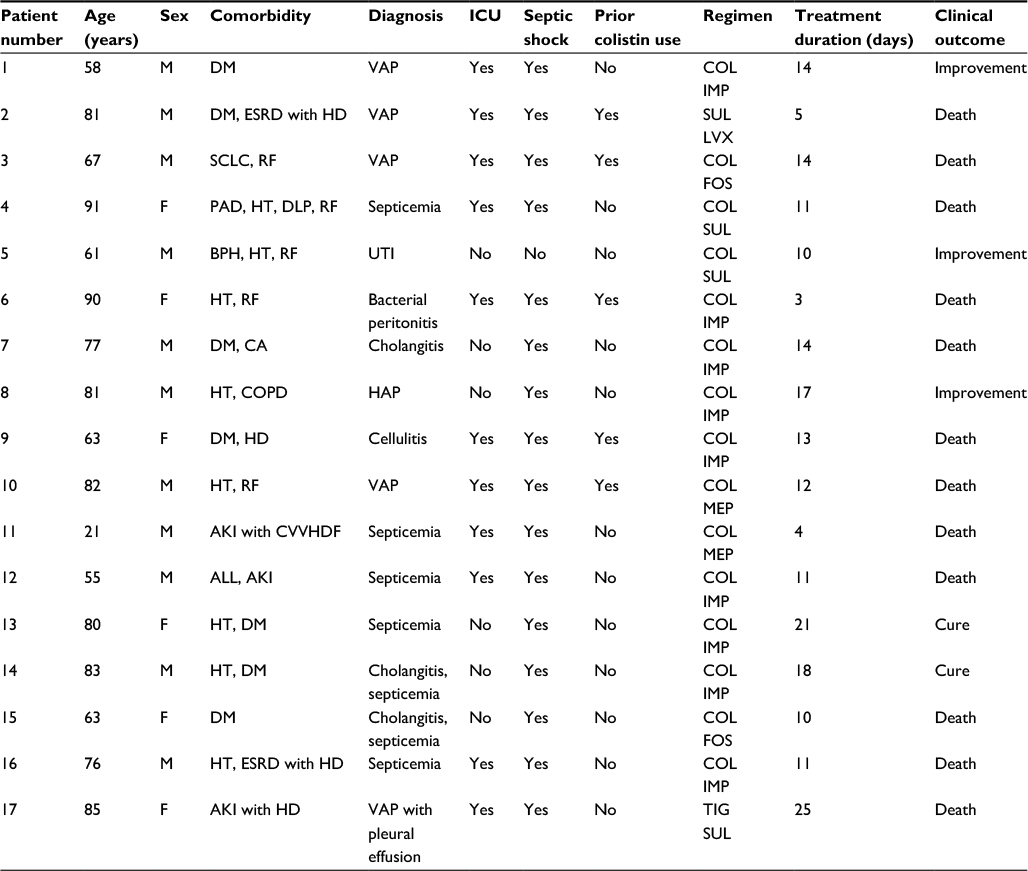 | Table 3 Characteristics and outcomes of patients infected with colistin-resistant Acinetobacter baumannii Abbreviations: AKI, acute kidney injury; ALL, acute lymphoblastic leukemia; BPH, benign prostatic hyperplasia; CA, cancer; COL, colistin; COPD, chronic obstructive pulmonary disease; CVVHDF, continuous veno-venous hemodiafiltration; DLP, dyslipidemia; DM, diabetes mellitus; ESRD, end-stage renal disease; FOS, fosfomycin; HAP, hospital-acquired pneumonia; HD, hemodialysis; HT, hypertension; ICU, intensive care unit; IMP, imipenem; LVX, levofloxacin; MEP, meropenem; PAD, peripheral artery disease; RF, renal failure; SCLC, small cell lung cancer; SUL, sulbactam; TIG, tigecycline; UTI, urinary tract infection; VAP, ventilator-associated pneumonia. |
Discussion
CoR-AB was first isolated in 1999 and is found mostly in carbapenem-resistant strains.5 Data from the EARS-Net collected from European countries revealed 5% resistance to polymyxins.11 However, another study conducted in Spain revealed polymyxin resistance rates as high as 40.6%.5 Fortunately, in Thailand, the National Antimicrobial Resistance Surveillance Thailand data in 2016 showed that the occurrence of CoR-AB is only 0.1%;3 however, this situation remains an urgent concern.
Our clinical outcome results indicate that the mortality rate was 70.6%, which is higher than the findings of Qureshi et al who showed a mortality rate of 30%.18 This high mortality might be due to the fact that our patients received less active antimicrobials, even in the form of combination therapy. In contrast, half of the CoR-AB patients in the previous study received at least two or more agents, with sulbactam as the treatment regimen against CoR-AB (sulbactam MIC90 of 8 µg/mL).18 However, data regarding the clinical outcomes of patients infected with CoR-AB are limited, and the impact of CoR-AB on morbidity and mortality is not well understood. Thus, further clinical studies are needed to evaluate the influence of CoR-AB infection.
Our results show that the CoR-AB strains exhibited high-level colistin resistance and that colistin as a monotherapy was ineffective; however, certain antimicrobials (e.g., tigecycline, sulbactam, or cotrimoxazole) remained active with the MIC values obtainable by standard dosages currently in clinical use. Moreover, a synergistic or additive effect was observed for colistin combined with imipenem, meropenem, sulbactam, or tigecycline. Thus, successful CoR-AB treatment could be accomplished using the remaining active agents or in combination with colistin.19
In this study, we found blaOXA23 in all clinical CoR-AB isolates, in agreement with previous studies conducted in Thailand.13,20,21 However, to the best of our knowledge, this is the first report on blaKPC identified in A. baumannii isolates in Thailand. Moreover, 52.4% of the CoR-AB isolates carried multiple carbapenemase genes. This prevalence is similar to our recent study;22 however, this is the first time we have identified the quadruple combination, blaOXA23, blaIMP, blaKPC, and blaNDM. To date, there are no existing data as to how multiple carbapenemase genes exist in A. baumannii. We hypothesize that the resistance genes might be spread via a mobile genetic element.23,24 However, the presence of such genes on plasmid needs to be confirmed by whole-genome or plasmid high-throughput sequencing.
A previous in vitro study has shown that CCCP significantly decreases the MIC values of colistin-resistant strains to susceptible levels (also termed colistin activity restoration). Our results suggest that colistin resistance in our CoR-AB isolates was mediated by an efflux pump. This mechanism is similar to the findings of Park and Ko;25 however, CCCP cannot be used clinically because of its intrinsic cytotoxicity. To date, no active EPI has been introduced into clinical practice; however, numerous EPIs from natural compounds and of synthetic origin have been developed. Newly developed EPIs could potentially be combined with colistin as standard treatment in our region.26 Additionally, to better understand the role of efflux pumps in CoR-AB, efflux pump mRNA transcription levels need to be determined.
Several limitations should be noted for our study. First, we gathered only 17 CoR-AB isolates; however, our isolates were from a 5-year period in a very low prevalence area. Second, we studied the colistin resistance via blamcr and efflux pump, but not by blapmrAB and blalpx.6 However, CCCP completely restored colistin activity, indicating that the resistance mechanism of all CoR-AB strains is mediated via an efflux pump. Finally, further studies are required to investigate the benefits of other active combinations against CoR-AB, such as tigecycline–sulbactam, sulbactam–imipenem, or cotrimoxazole. This in vitro study of synergism provides important information regarding the use of salvage regimens in cases of limited available antimicrobials.
Conclusion
CoR-AB clinical isolates showed a very high level of colistin resistance and had a high incidence of resistance genes. However, the mechanism of colistin resistance appears to be affected via an efflux pump. With high mortality rates, certain antimicrobials could be used as salvage treatments for CoR-AB infection.
Author contributions
All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Wisplinghoff H, Bischoff T, Tallent SM, Seifert H, Wenzel RP, Edmond MB. Nosocomial bloodstream infections in US hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin Infect Dis. 2004;39(3):309–317. | ||
Werarak P, Waiwarawut J, Tharavichitkul P, et al. Acinetobacter baumannii nosocomial pneumonia in tertiary care hospitals in Thailand. J Med Assoc Thai. 2012;95(Suppl 2):S23–S33. | ||
The National Antimicrobial Resistant Surveillance Thailand. Antibiogram; 2016. Available from: http://narst.dmsc.moph.go.th/. Accessed May 27, 2017 | ||
World Health Organization. WHO publishes list of bacteria for which new antibiotics are urgently needed: WHO Department of Communications. Available from: http://www.who.int/mediacentre/news/releases/2017/bacteria-antibiotics-needed/en/. Accessed May 27, 2017. | ||
Cai Y, Chai D, Wang R, Liang B, Bai N. Colistin resistance of Acinetobacter baumannii: clinical reports, mechanisms and antimicrobial strategies. J Antimicrob Chemother. 2012;67(7):1607–1615. | ||
Bialvaei AZ, Samadi Kafil H. Colistin, mechanisms and prevalence of resistance. Curr Med Res Opin. 2015;31(4):707–721. | ||
Gales AC, Jones RN, Sader HS. Contemporary activity of colistin and polymyxin B against a worldwide collection of Gram-negative pathogens: results from the SENTRY Antimicrobial Surveillance Program (2006–2009). J Antimicrob Chemother. 2011;66(9):2070–2074. | ||
Hejnar P, Kolar M, Hajek V. Characteristics of Acinetobacter strains (phenotype classification, antibiotic susceptibility and production of beta-lactamases) isolated from haemocultures from patients at the Teaching Hospital in Olomouc. Acta Univ Palacki Olomuc Fac Med. 1999;142:73–77. | ||
Yau W, Owen RJ, Poudyal A, et al. Colistin hetero-resistance in multidrug-resistant Acinetobacter baumannii clinical isolates from the Western Pacific region in the SENTRY antimicrobial surveillance programme. J Infect. 2009;58(2):138–144. | ||
Ko KS, Suh JY, Kwon KT, et al. High rates of resistance to colistin and polymyxin B in subgroups of Acinetobacter baumannii isolates from Korea. J Antimicrob Chemother. 2007;60(5):1163–1167. | ||
Giamarellou H. Epidemiology of infections caused by polymyxin-resistant pathogens. Int J Antimicrob Agents. 2016;48(6):614–621. | ||
Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Seventh Informational Supplement. Wayne, PA: Clinical and Laboratory Standards Institute; 2017:149. | ||
Santimaleeworagun W, Thathong A, Samret W, Preechachuawong P, Sae-lim W, Jitwasinkul T. Identification and characterization of carbapenemase genes in clinical isolates of carbapenem-resistant Acinetobacter Baumannii from general hospital in Thailand. Southeast Asian J Trop Med Public Health. 2014;45(4):874–880. | ||
Bou G, Cervero G, Dominguez MA, Quereda C, Martinez-Beltran J. PCR-based DNA fingerprinting (REP-PCR, AP-PCR) and pulsed-field gel electrophoresis characterization of a nosocomial outbreak caused by imipenem- and meropenem-resistant Acinetobacter baumannii. Clin Microbiol Infect. 2000;6(12):635–643. | ||
Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. | ||
Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. | ||
Ardebili A, Talebi M, Azimi L, Rastegar Lari A. Effect of Efflux Pump Inhibitor Carbonyl Cyanide 3-Chlorophenylhydrazone on the Minimum Inhibitory Concentration of Ciprofloxacin in Acinetobacter baumannii Clinical Isolates. Jundishapur J Microbiol. 2014;7(1):e8691. | ||
Qureshi ZA, Hittle LE, O’Hara JA, et al. Colistin-resistant Acinetobacter baumannii: beyond carbapenem resistance. Clin Infect Dis. 2015;60(9):1295–1303. | ||
Lenhard JR, Thamlikitkul V, Silveira FP, et al. Polymyxin-resistant, carbapenem-resistant Acinetobacter baumannii is eradicated by a triple combination of agents that lack individual activity. J Antimicrob Chemother. 2017;72(5):1415–1420. | ||
Niumsup PR, Boonkerd N, Tansawai U, Tiloklurs M. Carbapenem-resistant Acinetobacter baumannii producing OXA-23 in Thailand. Jpn J Infect Dis. 2009;62(2):152–154. | ||
Thapa B, Tribuddharat C, Srifuengfung S, Dhiraputra C. High prevalence of bla(OXA)-23 in oligoclonal carbapenem-resistant Acinetobacter baumannii from Siriraj Hospital, Mahidol University, Bangkok, Thailand. Southeast Asian J Trop Med Public Health. 2010;41(3):625–635. | ||
Santimaleeworagun W, Samret W, Preechachuawong P, Kerdsin A, Jitwasinkul T. Emergence of co-carbapenemase genes, blaOXA23, blaVIM, and blaNDM in carbapenem resistant Acinetobacter baumannii clinical isolates. Southeast Asian J Trop Med Public Health. 2016;47(5):1001–1007. | ||
Huang TW, Lauderdale TL, Liao TL, et al. Effective transfer of a 47 kb NDM-1-positive plasmid among Acinetobacter species. J Antimicrob Chemother. 2015;70(10):2734–2738. | ||
Martinez T, Martinez I, Vazquez GJ, Aquino EE, Robledo IE. Genetic environment of the KPC gene in Acinetobacter baumannii ST2 clone from Puerto Rico and genomic insights into its drug resistance. J Med Microbiol. 2016;65(8):784–792. | ||
Park YK, Ko KS. Effect of carbonyl cyanide 3-chlorophenylhydrazone (CCCP) on killing Acinetobacter baumannii by colistin. J Microbiol. 2015;53(1):53–59. | ||
Spengler G, Kincses A, Gajdacs M, Amaral L. New roads leading to old destinations: efflux pumps as targets to reverse multidrug resistance in bacteria. Molecules. 2017;22(3):E468. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.