Back to Journals » Infection and Drug Resistance » Volume 12
In vitro activity of cefoperazone and cefoperazone-sulbactam against carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa
Authors Lai CC ![]() , Chen CC
, Chen CC ![]() , Lu YC
, Lu YC ![]() , Chuang YC, Tang HJ
, Chuang YC, Tang HJ
Received 23 July 2018
Accepted for publication 7 November 2018
Published 20 December 2018 Volume 2019:12 Pages 25—29
DOI https://doi.org/10.2147/IDR.S181201
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Chih-Cheng Lai,1 Chi-Chung Chen,2,3 Ying-Chen Lu,3 Yin-Ching Chuang,2,4 Hung-Jen Tang5,6
1Department of Intensive Care Medicine, Chi Mei Medical Center, Liouying, Tainan, Taiwan; 2Department of Medical Research, Chi Mei Medical Center, Tainan, Taiwan; 3Department of Food Science, National Chiayi University, Chiayi, Taiwan; 4Department of Internal Medicine, Chi Mei Medical Center, Liouying, Tainan, Taiwan; 5Department of Medicine, Chi Mei Medical Center, Tainan, Taiwan; 6Department of Health and Nutrition, Chia-Nan University of Pharmacy and Science, Tainan, Taiwan
Background: This study aimed to investigate the in vitro activity of cefoperazone–sulbactam against carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa, and to evaluate the antibiotic resistance mechanisms of these bacteria.
Materials and methods: In total, 21 isolates of carbapenem-resistant P. aeruginosa and 15 isolates of carbapenem-resistant A. baumannii with different pulsed-field gel electrophoresis types were collected for assessment of the in vitro antibacterial activities of cefoperazone and cefoperazone–sulbactam and the associated resistance mechanisms of the bacteria.
Results: For carbapenem-resistant P. aeruginosa, the minimum inhibitory concentration (MIC) value and antibiotic susceptibility rate were similar for cefoperazone and cefoperazone–sulbactam (at 1:1 and 2:1 ratios). In contrast, for carbapenem-resistant A. baumannii, the MIC values, including the MIC range, MIC that inhibited 50% of isolates (MIC50) and MIC that inhibited 90% of isolates (MIC90), were reduced after treatment with sulbactam and cefoperazone. We screened resistance genes, including VIM-2, OXA-2 and OXA-10, in 21 carbapenem-resistant P. aeruginosa isolates. Only one (4.8%) of the isolates showed expression of VIM-2, and neither the OXA-2 nor the OXA-10 gene was detected. However, 20 (95.2%) isolates among the carbapenem-resistant P. aeruginosa isolates selected for oprD sequencing showed the phenomenon of nucleotide substitution or deletion. Among 15 carbapenem-resistant A. baumannii isolates, we found that ten (66.7%) isolates had concomitant expression of the OXA-23 and ISAba1-OXA-23 genes, and six (40.0%) isolates had expression of the OXA-24-like gene. All 15 isolates had OXA-51-like gene expression, and only 1 (6.7%) isolate had ISAba1-OXA-51-like gene expression. None of the isolates contained the IMP-1, IMP-8, KPC, NDM, VIM-1 or OXA-48 genes.
Conclusion: The in vitro antibacterial activity of cefoperazone against carbapenem-resistant A. baumannii can be enhanced by adding sulbactam to cefoperazone, but the addition does not affect carbapenem-resistant P. aeruginosa. This significant difference can be explained by the different resistance mechanisms of carbapenem-resistant A. baumannii and P. aeruginosa.
Keywords: cefoperazone–sulbactam, Acinetobacter baumannii, Pseudomonas aeruginosa
Introduction
Both the Acinetobacter baumannii complex and Pseudomonas aeruginosa belong to the non-fermentative Gram-negative bacilli, and they are common causes of clinical infection. Although carbapenems, including imipenem, meropenem and doripenem, have broad-spectrum antibacterial activity and are commonly used to treat infections caused by A. baumannii and P. aeruginosa, carbapenem resistance among these two pathogens is gradually increasing.1,2 The list of antibiotics that are effective against these carbapenem-resistant organisms is becoming limited. In vitro studies3,4 have shown that the addition of sulbactam to cefoperazone can help enhance its antibacterial activity against carbapenem-resistant Enterobacteriaceae or A. baumannii. In contrast, two studies4,5 did not show an additive effect of sulbactam on cefoperazone against carbapenem-resistant P. aeruginosa. Therefore, we urgently need a more comprehensive evaluation of the activity of cefoperazone–sulbactam against different carbapenem-resistant organisms. However, knowledge about the antimicrobial activity of cefoperazone–sulbactam against the carbapenem-resistant A. baumannii complex and P. aeruginosa and their associated antibiotic resistance mechanisms are limited. Therefore, we conducted this study to investigate the in vitro activity of cefoperazone–sulbactam against carbapenem-resistant A. baumannii and P. aeruginosa and to evaluate their antibiotic resistance mechanisms.
Materials and methods
Collection of clinical isolates
In total, 21 isolates of carbapenem-resistant P. aeruginosa and 15 isolates of carbapenem-resistant A. baumannii with different pulsed-field gel electrophoresis (PFGE) types were collected for study. Carbapenem resistance is defined as resistance to imipenem, meropenem or doripenem, and the carbapenem-resistant phenotypes of the P. aeruginosa and A. baumannii isolates were confirmed by the modified Hodge test. Species confirmation was performed by standard biochemical methods on a VITEK 2 automated system (bioMérieux, Marcy l’Etoile, France).
In vitro susceptibility
The minimum inhibitory concentrations (MICs) of the drugs were measured by broth microdilution in prepared Mueller–Hinton broth (Oxoid, Basingstoke, UK) supplemented with 25 µg/mL of calcium and 12.5 µg/mL of magnesium (CAMHB). All experiments were performed in triplicate, and the microdilution trays were incubated at 35°C for 16–20 hours.6 Standard powders of cefoperazone and sulbactam were provided by TTY (TTY Biopharm, Taipei, Taiwan), and MIC determinations and susceptibility interpretation criteria followed the Clinical and Laboratory Standards Institute guideline.7 Antimicrobial susceptibilities were determined using broth microdilution MIC tests with a standard inoculum (5×105 CFU/mL). For both microorganisms, doubling dilutions of cefoperazone ranged from 0.25 to 64 µg/mL, and three different sets of dilutions were prepared. The first series of cefoperazone dilutions was created without adding sulbactam. The second series contained cefoperazone combined with sulbactam in a 2:1 ratio (two parts cefoperazone and one part sulbactam). The third series contained cefoperazone combined with sulbactam in a 1:1 ratio (one part cefoperazone and one part sulbactam). Susceptibilities to cefoperazone alone and cefoperazone–sulbactam at 1:1 and 2:1 ratios were classified according to the MIC of cefoperazone ≤16 µg/mL.7 We used an ELISA plate reader for reading. Bacterial growth was detected by OD (ELISA plate reader, Epoch™ Microplate spectrophotometer; BioTek Instruments, Winooski, VT, USA). Escherichia coli ATCC 25922 and Kp ATCC 700603 were used as quality-control strains.
Detection of β-lactamase genes
Gene detection was modified as described previously.6 In brief, DNA was extracted to be used as a template, and PCR was used to amplify IMP-1, IMP-8, KPC, NDM, OXA-2, OXA-10, OXA-23, ISAba1-OXA-23, OXA-24-like, OXA-48, OXA-51-like, ISAba1-OXA-51-like, oprD, VIM-1 and VIM-2 using specific primers.6,8,9 PCR products were purified with PCR clean-up kits (Roche Diagnostics, Penzberg, Germany) and sequenced on an ABI Prism 3730 sequencer analyzer (Applied Biosystems, Foster City, CA, USA).
Results
The MIC values of cefoperazone alone and in combination with sulbactam against carbapenem-resistant P. aeruginosa and carbapenem-resistant A. baumannii are shown in Table 1. For carbapenem-resistant P. aeruginosa, the MIC range, MIC50 and MIC90 values were similar for cefoperazone and cefoperazone–sulbactam (at 1:1 and 2:1 ratios). In contrast, for carbapenem-resistant A. baumannii, the MIC values, including the MIC range, MIC50 and MIC90, were reduced after adding sulbactam with cefoperazone. The susceptibility rates of carbapenem-resistant P. aeruginosa to cefoperazone, cefoperazone–sulbactam (1:1) and cefoperazone–sulbactam (2:1) were 23.8%, 28.6% and 33.3%, respectively. However, the susceptibility rates of carbapenem-resistant A. baumannii to cefoperazone, cefoperazone–sulbactam (1:1) and cefoperazone–sulbactam (2:1) were 0.0%, 80.0% and 40.0%, respectively.
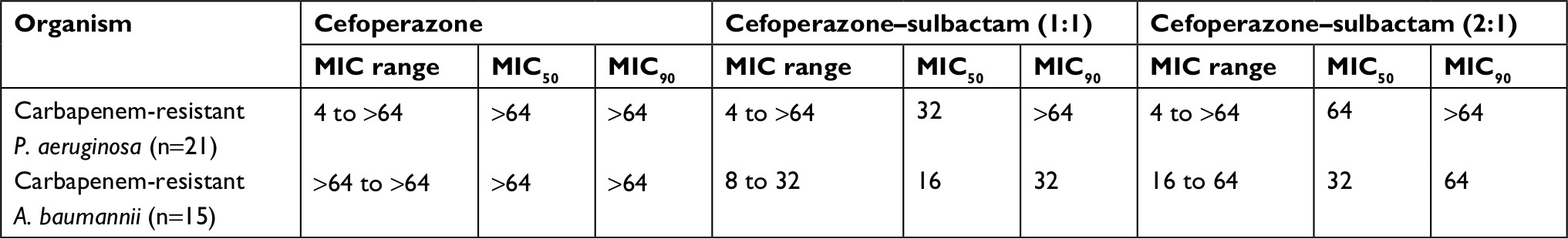 | Table 1 MIC range, MIC50 and MIC90 of cefoperazone alone, cefoperazone–sulbactam (1:1) and cefoperazone–sulbactam (2:1) against different organisms Abbreviations: MIC, minimum inhibitory concentration; MIC50, MIC that inhibited 50% of isolates; MIC90, MIC that inhibited 90% of isolates; A. baumannii, Acinetobacter baumannii; P. aeruginosa, Pseudomonas aeruginosa. |
We found that the resistance mechanisms of carbapenem-resistant P. aeruginosa and carbapenem-resistant A. baumannii isolates were different (Tables 2 and 3). Among 21 carbapenem-resistant P. aeruginosa isolates, we screened resistance genes, including VIM-2, OXA-2 and OXA-10. Only one (4.8%) of the isolates had expression of VIM-2, and neither the OXA-2 nor the OXA-10 gene was detected. However, 20 (95.2%) isolates among the carbapenem-resistant P. aeruginosa isolates selected for oprD sequencing showed the phenomenon of nucleotide substitution or deletion. Among 15 carbapenem-resistant A. baumannii isolates, we found that 10 (66.7%) isolates had concomitant expression of the OXA-23 and ISAba1-OXA-23 genes, and 6 (40.0%) isolates had expression of the OXA-24-like gene. All of the 15 isolates had OXA-51-like expression, and only 1 (6.7%) isolate had ISAba1-OXA-51-like expression. None of them had detectable IMP-1, IMP-8, KPC, NDM, VIM-1 or OXA-48 genes.
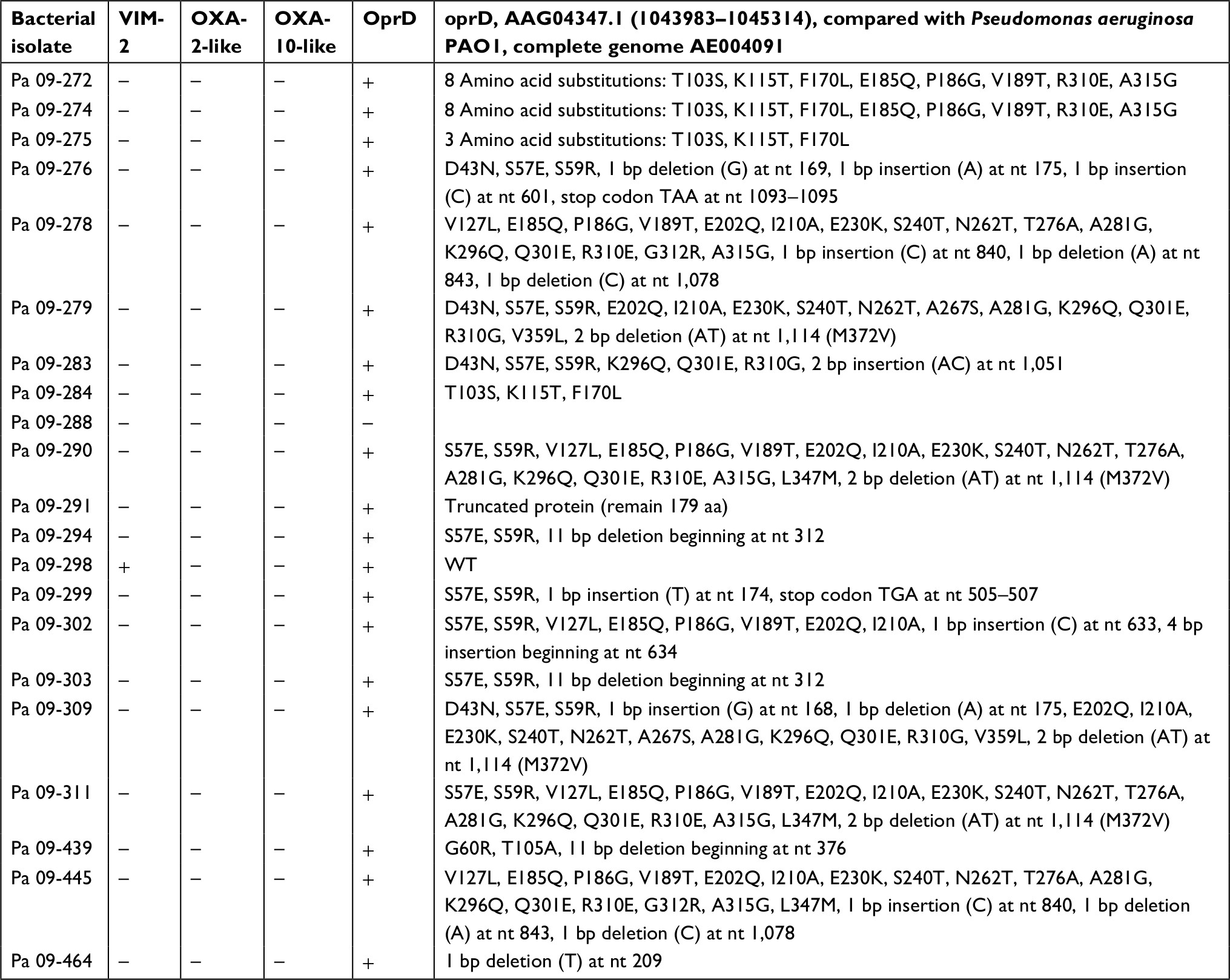 | Table 2 Resistance mechanisms of carbapenem-resistant Pseudomonas aeruginosa (Pa) isolates |
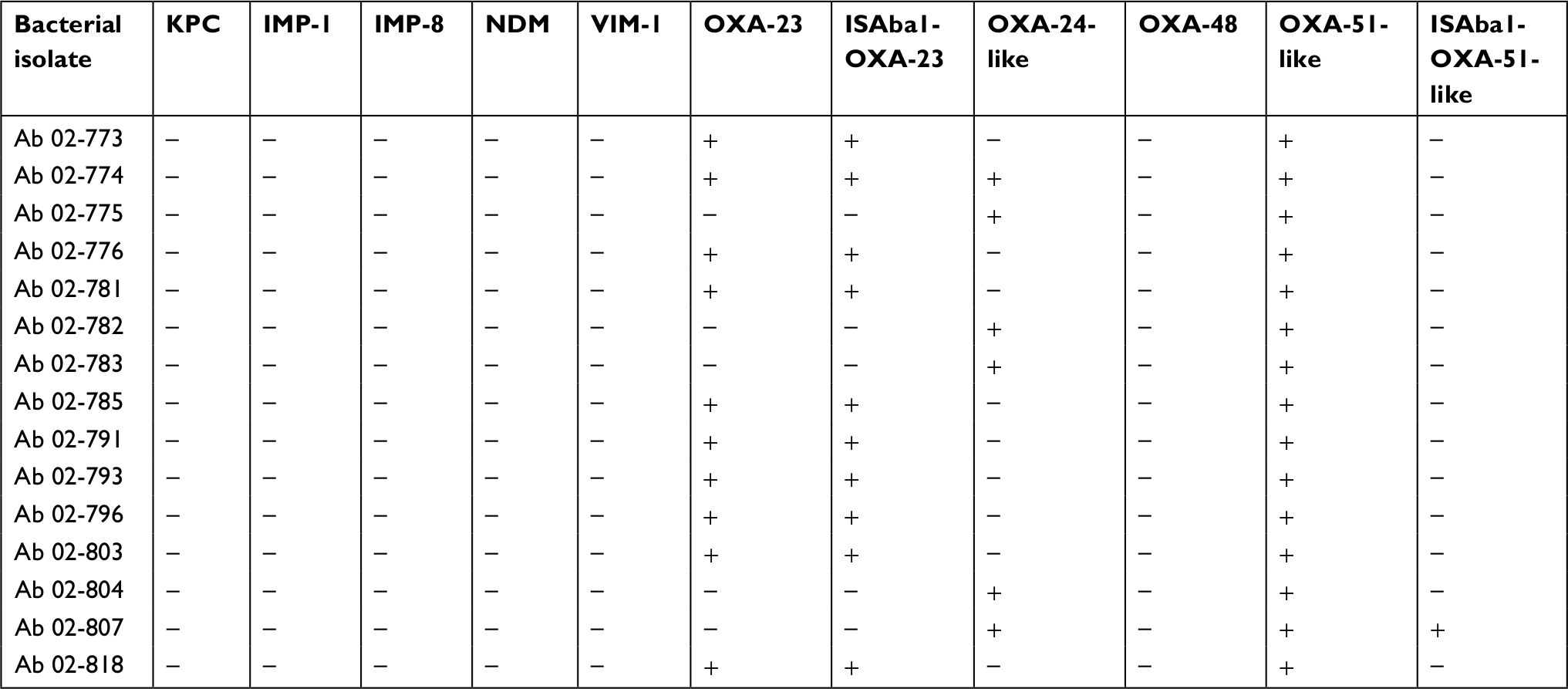 | Table 3 Resistance mechanisms of carbapenem-resistant Acinetobacter baumannii (Ab) isolates |
Discussion
This study investigated the in vitro activities of different compositions of cefoperazone–sulbactam and cefoperazone alone against carbapenem-resistant P. aeruginosa and A. baumannii. We also assessed their antibiotic resistance mechanisms to help us better understand the association between antibiotic activity and resistance mechanisms in these two microorganisms. First, after the addition of sulbactam with cefoperazone, the in vitro antibiotic activity, in terms of the MIC values and antibiotic susceptibility rates, against carbapenem-resistant A. baumannii improved. In contrast, this changed antibiotic composition did not change the antibiotic activity against carbapenem-resistant P. aeruginosa. In summary, after adding sulbactam with cefoperazone, we can enhance the in vitro activity against the carbapenem-resistant A. baumannii complex but not against carbapenem-resistant P. aeruginosa.
In addition, to explain the different activities of cefoperazone or cefoperazone–sulbactam against carbapenem-resistant P. aeruginosa and A. baumannii isolates, this study investigated the resistance mechanism of these two bacteria. The resistance mechanism of carbapenem-resistant P. aeruginosa was mainly attributed to nucleotide substitution or deletion in the oprD gene, and few isolates carried the VIM-2 gene. This finding is consistent with a previous study of 78 non-duplicated imipenem-resistant P. aeruginosa isolates that showed alterations in the oprD protein and the presence of an active efflux pump are the main antibiotic resistance mechanisms of P. aeruginosa.10 Such a resistance mechanism may keep carbapenem-resistant P. aeruginosa isolates resistant to β-lactams even after adding β-lactamase inhibitors, such as sulbactam. Therefore, we did not find a synergistic effect between the β-lactam (cefoperazone) and the β-lactamase inhibitor (sulbactam) against carbapenem-resistant P. aeruginosa in this study.
Unlike serine-dependent β-lactamases (classes A, C and D), class B β-lactamases are metallo-β-lactamases (MBLs) that require zinc or another heavy metal for catalysis. MBLs are not inhibited by mechanism-based inhibitors, such as clavulanate, sulbactam or tazobactam.11 The most common families of identified acquired class B MBLs include the VIM and IMP groups, along with the emerging NDM group, none of which was found in our carbapenem-resistant A. baumannii isolates. In contrast, most of the carbapenem-resistant A. baumannii isolates in this study carried resistance genes, including the OXA-23 gene, ISAba1-OXA-23 gene, OXA-24-like gene and OXA-51-like gene, as previously described.12 The β-lactamases, OXA-23, OXA-24 and OXA-51, in the carbapenem-resistant A. baumannii in this study belonged to class D, and were their major resistance mechanism to sulbactam.13 Sulbactam is chemically a penicillanic acid sulfone and shows particular activity against class A β-lactamases, whereas its activity against class D enzymes is less potent. Similarly, inhibition of OXA-type enzymes by sulbactam is not as strong as the inhibition produced by TEM-1 and other clinically used inhibitors.14 In this study, the concentration of sulbactam was higher than the conventional concentration, and this concentration may increase the affinity of sulbactam for class D β-lactamases and therefore the activity against these carbapenem-resistant A. baumannii isolates. Thus, we found that after adding a relatively high concentration of sulbactam with cefoperazone, the in vitro activity against carbapenem-resistant A. baumannii could be enhanced.15 However, further studies are warranted to elucidate the role of cefoperazone and sulbactam in the treatment of infection and assess β-lactamase gene expression.
This study had one major limitation. Only small numbers of clinical isolates were available in our institution because we wanted to investigate isolates with different PFGE patterns; thus, the clinical utility of the results may be limited. Further large-scale studies are warranted to confirm our findings.
Conclusion
The in vitro activity against carbapenem-resistant A. baumannii can be enhanced by adding sulbactam with cefoperazone, but this treatment does not work for carbapenem-resistant P. aeruginosa. This significant difference can be explained by the difference in resistance mechanisms between carbapenem-resistant A. baumannii and P. aeruginosa.
Data sharing statement
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Disclosure
The authors report no conflicts of interest in this work.
References
Huang YT, Hsueh PR. Antimicrobial drug resistance in Taiwan. Int J Antimicrob Agents. 2008;32(Suppl 3):S174–S178. | ||
Lin KY, Lauderdale TL, Wang JT, Chang SC. Carbapenem-resistant Pseudomonas aeruginosa in Taiwan: prevalence, risk factors, and impact on outcome of infections. J Microbiol Immunol Infect. 2016;49(1):52–59. | ||
Kuo HY, Wang FD, Yen YF, Lin ML, Liu CY. In vitro activities of piperacillin or cefoperazone alone and in combination with beta-lactamase inhibitors against Gram-negative bacilli. New Microbiol. 2009;32(1):49–55. | ||
Lai CC, Chen CC, Lu YC, Lin TP, Chuang YC, Tang HJ. Appropriate composites of cefoperazone-sulbactam against multidrug-resistant organisms. Infect Drug Resist. 2018;11:1441–1445. | ||
Chang PC, Chen CC, Lu YC, et al. The impact of inoculum size on the activity of cefoperazone-sulbactam against multidrug resistant organisms. J Microbiol Immunol Infect. 2018;51(2):207–213. | ||
Lai CC, Chen CC, Huang HL, Chuang YC, Tang HJ. The role of doxycycline in the therapy of multidrug-resistant E. coli – an in vitro study. Sci Rep. 2016;6:31964. | ||
Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Testing of Bacteria that Grow Aerobically; Approved Standard. 8th ed. CLSI document M07-A9. Wayne, PA: CLSI; 2012. | ||
Lin YC, Hsia KC, Chen YC, et al. Genetic basis of multidrug resistance in Acinetobacter clinical isolates in Taiwan. Antimicrob Agents Chemother. 2010;54(5):2078–2084. | ||
Moubareck C, Brémont S, Conroy MC, Courvalin P, Lambert T. GES-11, a novel integron-associated GES variant in Acinetobacter baumannii. Antimicrob Agents Chemother. 2009;53(8):3579–3581. | ||
Kao CY, Chen SS, Hung KH, et al. Overproduction of active efflux pump and variations of OprD dominate in imipenem-resistant Pseudomonas aeruginosa isolated from patients with bloodstream infections in Taiwan. BMC Microbiol. 2016;16(1):107. | ||
Jeon JH, Lee JH, Lee JJ, et al. Structural basis for carbapenem-hydrolyzing mechanisms of carbapenemases conferring antibiotic resistance. Int J Mol Sci. 2015;16(5):9654–9692. | ||
Sen B, Joshi SG. Studies on Acinetobacter baumannii involving multiple mechanisms of carbapenem resistance. J Appl Microbiol. 2016;120(3):619–629. | ||
Kuo SC, Lee YT, Yang Lauderdale TL, et al. Contribution of Acinetobacter-derived cephalosporinase-30 to sulbactam resistance in Acinetobacter baumannii. Front Microbiol. 2015;6:231. | ||
Akova M. Sulbactam-containing beta-lactamase inhibitor combinations. Clin Microbiol Infect. 2008;14(Suppl 1):185–188. | ||
Chaudhary M, Payasi A. Prevalence, genotyping of Escherichia coli and Pseudomonas aeruginosa clinical isolates for oxacillinase resistance and mapping susceptibility behavior. J Microb Biochem Technol. 2014;6:63–67. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.