Back to Journals » Nature and Science of Sleep » Volume 17

Impact of Obstructive Sleep Apnea on Endometrial Function in Female Rats: Mechanism Exploration
Authors Zhang D ![]() , Bian W
, Bian W ![]() , Gao Z
, Gao Z
Received 2 June 2025
Accepted for publication 25 August 2025
Published 4 October 2025 Volume 2025:17 Pages 2485—2499
DOI https://doi.org/10.2147/NSS.S540493
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah L Appleton
Dong Zhang,* Wenli Bian,* Zhihua Gao
Department of Gynecology, Haihe Hospital, Tianjin University, Tianjin, 300350, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dong Zhang, Department of Gynecology, Haihe Hospital, Tianjin University, Tianjin, 300350, People’s Republic of China, Tel +86 13920148868, Fax +86 02258830336, Email [email protected]
Background: Elevated systemic oxidative stress contributes to endometrial damage. Individuals with obstructive sleep apnea (OSA) exhibit significantly elevated oxidative stress; however, the potential role of oxidative stress in OSA-induced endometrial injury remains unclear.
Objective: To investigate the effects of OSA on systemic oxidative stress and endometrial morphological alterations in a female rat model.
Methods: We randomly assigned 15 female Sprague-Dawley (SD) rats to three groups: (1) Control group: Normal feeding for 8 weeks; (2) Short-term OSA group: 4 weeks of normal feeding followed by 4 weeks of Sleep Apnea (SA) modeling; (3) Long-term OSA group: 8 weeks of SA modeling.Assessments included: Body weight; uterine index; Oxidative stress markers: superoxide dismutase (SOD), reactive oxygen species (ROS) and malondialdehyde (MDA);Endometrial histomorphology: thickness, microvessel density and gland count via Hematoxylin and Eosin (H&E) staining; immunohistochemical (IHC) analysis of Kiel 67 (Ki-67) antigen and vascular endothelial growth factor (VEGF); Apoptosis detection by terminal deoxynucleotidyl transferase dUTP Nick-End Labeling (TUNEL) assay.
Results: Long-term OSA exposure significantly increased body weight vs control (P< 0.05). Both OSA groups showed reduced uterine indices and elevated oxidative stress (P< 0.05). Progressive structural impairment was observed with OSA duration: endometrial thickness and microvessel density decreased sequentially (control > short-term > long-term; P< 0.05), and gland number was reduced in the long-term group vs control (P< 0.05). IHC showed duration-dependent suppression of Ki-67 (proliferation) and VEGF (angiogenesis) expression (P< 0.05), while apoptosis increased with OSA exposure (P< 0.05).
Conclusion: In a preclinical model, OSA-like exposure promoted weight gain, uterine atrophy, and progressive endometrial damage. Mechanistic analyses revealed that this impairment resulted from oxidative stress-mediated inhibition of cellular proliferation (reflected by reduced Ki-67 expression) and suppression of angiogenesis (indicated by decreased VEGF levels), concurrent with enhanced apoptotic activity. Given the observed duration-dependent pathological progression, our findings establish that sleep apnea contributes to female reproductive dysfunction, warranting early clinical intervention in women with sleep-disordered breathing.
Keywords: sleep apnea, oxidative stress, endometrium, Ki-67
Introduction
Obstructive sleep apnea (OSA), now recognized as the fourth most prevalent chronic disease worldwide, has emerged as a critical public health challenge. This condition is pathologically characterized by recurrent partial or complete upper airway collapse during sleep, leading to intermittent hypoxia (IH), hypercapnia, and sleep fragmentation.1 Epidemiological data reveal a striking increase in OSA prevalence in developed countries, rising from 11% in 1993 to 34% in 2017, with rates reaching 50% in certain obese populations.2 Notably, while OSA has traditionally been considered a male-dominant disorder (male-to-female ratio ~2-3:1), recent studies indicate that 6–15% of adult women are affected, with prevalence soaring to 30% among obese women.3 Particularly alarming is the 3.1-fold higher risk in perimenopausal women compared to those of reproductive age.4 Despite these figures, 80% of female OSA cases remain undiagnosed due to atypical symptomatology, suggesting that the disease burden in women has been substantially underestimated.3
The multisystem damage induced by OSA has been conceptualized as a “hypoxia-reoxygenation-inflammation” pathological axis. Key mechanisms including IH, hypercapnia, sleep fragmentation, and exaggerated intrathoracic pressure swings drive systemic complications, with well-documented impacts on metabolic/endocrine, cardiovascular, reproductive, neurocognitive, oncological, urinary, and hematopoietic systems through both experimental and clinical evidence.5
Within the reproductive system, OSA-induced oxidative stress appears to play a pivotal role. Studies demonstrate that cyclic hypoxia-reoxygenation in OSA patients sustains a state of elevated oxidative stress, which may trigger endometrial damage and subsequent uterine dysfunction, ultimately compromising female reproductive capacity.6,7 Despite these findings, direct evidence linking OSA to endometrial injury remains scarce.
To address this gap, we developed a novel female rat model replicating OSA-like intermittent hypoxia. Kiel 67 (Ki-67) antigen expression is a well-established indicator of proliferative activity in the endometrium, while vascular endothelial growth factor (VEGF) expression serves as a critical marker for assessing physiological status and pathological alterations. Through comprehensive evaluation of: (1) serum oxidative stress markers (superoxide dismutase [SOD], reactive oxygen species [ROS] and malondialdehyde [MDA]); (2) endometrial histomorphometry (thickness, gland density, vascularity); (3) proliferation marker Ki-67; and (4) angiogenic factor VEGF, we provide the first systematic investigation into OSA-specific endometrial pathology, establishing a mechanistic foundation for its reproductive consequences.
This study, utilizing an OSA animal model, for the first time reveals the pathological mechanism whereby intermittent hypoxia disrupts the endometrial regenerative niche via the oxidative stress axis. These findings deliver two pivotal clinical implications: (1) It indicates that OSA may induce reproductive disorders through endometrial impairment, advocating for OSA screening and early intervention in women with infertility or recurrent pregnancy loss. (2) It identifies the prognostic potential of serum oxidative markers (MDA/SOD) in predicting OSA-related endometrial damage outcomes.
Materials and Methods
Materials
Experimental Animals
The Animal Research Reporting Guidelines state that a minimum of 5 biologically independent animals per group for histopathology studies.8 Fifteen female Sprague-Dawley (SD) rats (specific pathogen-free [SPF] grade, aged 6–8 weeks, body weight 170–190 g) were purchased from Sibeifu (Beijing) Biotechnology Co., Ltd. All animal husbandry and management procedures complied with the Laboratory Animal Management Regulations of Tianjin University Haihe Hospital. This study was approved by the Ethics Committee of Tianjin University Haihe Hospital (Approval No. 2025HHWZ(A)-003).
Main Reagents and Instruments
Intermittent hypoxia chamber: Purchased from Shanghai Tawang Intelligent Technology Co., Ltd. (China); Primary antibodies: Ki-67 and VEGF antibodies were purchased from Abcam (USA);Secondary antibodies and chemicals: Biotin-labeled goat anti-rabbit secondary antibody, phosphate-buffered saline (PBS), and 3,3’-diaminobenzidine (DAB) were obtained from Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd. (China); Biochemical kits: SOD, ROS, and MDA assay kits (colorimetric/fluorometric methods) were purchased from Nanjing Jiancheng Bioengineering Institute (China); Paraffin microtome: Leica RM2235 (Leica Microsystems, Germany);Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) assay kit: One-step TUNEL In Situ Apoptosis Detection Kit (Elabscience, China); Optical microscope: Olympus CX23 (Olympus Corporation, Japan); Inverted fluorescence microscope: SOPTOP-XD Series (SOPTOP, China).
Methods
Model Preparation
The OSA model was established using a hypoxia chamber as previously described.9 For OSA modeling: Gas delivery: Compressed air and a gas mixture (90% N₂ + 10% O₂) were alternately delivered into the hypoxia chamber at a flow rate of 2 L/min via solenoid valve control. Oxygen cycling: Oxygen concentration cycled between 10% and 21% at 30-second intervals (1 cycle/min) for 8 h/day (8:00 AM–4:00 PM). Post-hypoxia protocol: Outside hypoxia exposure, rats were returned to standard cages with normal air, room temperature (22–25°C), and ad libitum feeding. Specimens were collected on the day following the experimental endpoint. Control group: Rats were housed in identical polymethyl methacrylate (PMMA) chambers with normoxic air (21% O₂) and no hypoxia intervention. Oxygen concentrations during intermittent hypoxia experiments were monitored in real-time using an oxygen analyzer (Shanghai Tawang Intelligent Technology Co., Ltd). The experimental designer was aware of group allocation only during the design and assignment phases, whereas operators, outcome assessors, and the statistical analyst remained blinded throughout all subsequent stages (including experiment conduction, data collection, and analysis).
Animal Grouping
Fifteen rats were randomly assigned to three groups (n=5 per group) using a random number table method: Long-term group (Exposed to the OSA model for 8 weeks), Short-term group (Normal feeding for 4 weeks followed by OSA model exposure for 4 weeks), and Control group (Normal feeding for 8 weeks under identical environmental conditions). All animals were housed in a SPF animal facility. At the experimental endpoint, animals were euthanized by intraperitoneal injection of an overdose of pentobarbital sodium, in strict accordance with the American Veterinary Medical Association Guidelines (2020). All procedures were approved by the Ethics Committee of Tianjin Haihe Hospital (Approval No. 2025HHWZ(A)-003, which waived the requirement for predefined exclusion criteria due to the exploratory nature. The reporting of this study adhered to the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines 2.0. This protocol did not preset any exclusion thresholds; consequently, all collected datapoints were incorporated into the final analytical dataset. The primary outcome measures in this study were histopathological alterations in the rat endometrium (including endometrial thickness, gland count, and vascular density) along with immunohistochemical expression levels of Ki-67 and VEGF.
Body Weight and Uterine Index
Body weight was measured for all animals before the experiment and on the day after the experimental endpoint. At the endpoint, rats were euthanized, and uteri were dissected and weighed. The uterine index was calculated as follows:
Uterine Index (%) = Uterine Weight (g)/Body Weight (g)×100.
Histopathological Observation
After uterine tissue collection, specimens were fixed in 4% paraformaldehyde for 48 h, paraffin-embedded, and sectioned into 5 μm slices. Three sections per rat were subjected to routine Hematoxylin and Eosin (H&E) staining. Endometrial histomorphological changes were observed under an optical microscope (Olympus CX23, Japan). Endometrial thickness, defined as the vertical distance from the endometrial-myometrial junction to the uterine lumen, was measured using ImageJ software (National Institutes of Health, USA). Additionally, endometrial vascular density and gland number were quantified by counting vessels and glands in five randomly selected high-power fields (HPF, 40× magnification) per section. The average values from three sections were calculated as the final vascular density and gland number for each sample.
IHC Detection of Ki-67 and VEGF Expression in the Endometrium
Staining Protocol
Section Preparation: Paraffin sections were baked overnight at 37°C, rehydrated through a graded ethanol series, and rinsed with phosphate-buffered saline (PBS) three times (3 min each).
Antigen retrieval was performed via microwave heating (3 min × 2 cycles).
Endogenous Peroxidase Blocking: Sections were incubated with 3% H₂O₂ at room temperature (RT) for 15 min to block endogenous peroxidase activity, followed by PBS rinses (3 min × 3).
Blocking and Antibody Incubation: Sections were incubated with blocking serum (ready-to-use) for 15 min. Excess serum was gently removed without washing.
Primary antibodies (Ki-67 and VEGF, diluted as per manufacturer’s instructions) were applied and incubated overnight at 4°C. Sections were rinsed with PBS (3 min × 3).
Biotin-labeled secondary antibodies (goat anti-rabbit IgG) were added for 15 min at RT, followed by PBS rinses (3 min × 3).
Streptavidin-horseradish peroxidase (HRP) working solution was applied for 15 min, and sections were rinsed with PBS (3 min × 3).
Chromogenic Reaction and Counterstaining
3,3′-Diaminobenzidine (DAB) was used for chromogenic development. Sections were rinsed with tap water for 3 min. Hematoxylin counterstaining was performed for 15 min, followed by bluing in PBS for 4 min. Sections were dehydrated through a graded ethanol series and mounted with neutral resin.
Observation and Quantification
Microscopic Evaluation: Ki-67-positive cells exhibited distinct brown-yellow granular staining in the nucleus. VEGF-positive cells displayed cytoplasmic brown-yellow staining.
Image Analysis: Stained sections were imaged under an Olympus CX23 microscope.
ImageJ software (National Institutes of Health, USA) was used to quantify the positive expression rates of Ki-67 and VEGF. Positive cells were counted in five randomly selected high-power fields (40×) per section, and the average value was calculated to reflect endometrial cell proliferation.
Measurement of Serum SOD, ROS, and MDA Levels Using Biochemical Kits
Venous blood was collected from rats and centrifuged at 3,000 rpm for 10 min to isolate serum. Serum levels of SOD, ROS, and MDA were quantified using commercial biochemical kits (Nanjing Jiancheng Bioengineering Institute, China) following the manufacturer’s instructions. SOD activity: Measured via the xanthine oxidase method (colorimetric assay, absorbance at 550 nm).ROS levels: Detected using a 2’,7’-dichlorodihydrofluorescein diacetate (DCFH-DA) probe (fluorometric assay, excitation/emission: 488/525 nm).MDA content: Determined by the thiobarbituric acid (TBA) method (colorimetric assay, absorbance at 532 nm). All measurements were performed in triplicate, and results were normalized to total protein concentration (Bradford assay).
Detection of Endometrial Cell Apoptosis
Paraffin sections of rat uterine tissues were dewaxed, rehydrated, and incubated with TUNEL equilibration buffer (One-Step TUNEL In Situ Apoptosis Detection Kit, Elabscience, China). After rinsing, sections were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and mounted with anti-fade medium. Apoptotic cells were visualized and imaged under a fluorescence microscope (SOPTOP-XD, China) using green fluorescence (excitation/emission: 488/530 nm). Fluorescence intensity was quantified using ImageJ software (National Institutes of Health, USA). Apoptosis levels were expressed as the ratio of TUNEL-positive cells to total DAPI-stained nuclei in five randomly selected fields per section.
Statistical Analysis
All statistical analyses were performed using SPSS 21.0 software (IBM Corp). Normality assumptions for the linear regression model were verified using Shapiro–Wilk tests and residual Q-Q plots, while homogeneity of variances was assessed via Levene’s tests. When the normality assumption was violated, logarithmic transformation was applied to the data; when variance homogeneity was not established, Welch’s analysis of variance (ANOVA) was employed as the alternative approach. Continuous variables were expressed as mean ± standard deviation (mean ± SD). For comparisons among three or more groups, one-way ANOVA was applied. If the ANOVA revealed statistically significant differences (threshold: P < 0.05), Fisher’s least significant difference (LSD) post hoc test was subsequently employed for pairwise comparisons. A two-tailed P-value < 0.05 was considered statistically significant for all tests.
Results
Body Weight and Uterine Index Across Experimental Groups
Table 1 and Figure 1 demonstrate that at baseline, no significant differences in body weight were observed among the three groups (P > 0.05). At the experimental endpoint: Body weight: A statistically significant difference was detected between the control group and long-term group (P < 0.05), whereas no differences existed between control vs short-term group or short-term vs long-term groups (P > 0.05 for all).
 |
Table 1 Comparison of Body Weight and Uterine Index Across Experimental Groups |
 |
Figure 1 Initial body weight, final body weight, and uterine index in three experimental groups. Notes: control group, normal feeding for 8 weeks under identical environmental conditions; short-term group, normal feeding for 4 weeks followed by obstructive sleep apnea (OSA) model exposure for 4 weeks; long-term group,exposed to the OSA model for 8 weeks; *P <0.05; ns, P≥0.05. |
Uterine index: Significant intergroup differences were identified at the endpoint, with the control group differing significantly from both short-term (P < 0.05) and long-term groups (P < 0.05). However, no difference was observed between short-term and long-term groups (P > 0.05).
These findings suggest that sleep apnea may contribute to increased body weight in rats.
Serum Levels of MDA, SOD, and ROS Across Experimental Groups
Table 2 and Figure 2 demonstrate that Serum MDA concentrations progressively increased across control, short-term, and long-term groups, with statistically significant differences observed in all pairwise comparisons (P < 0.05 for each).Serum ROS levels followed a similar ascending trend, showing significant intergroup differences (control vs short-term: P < 0.05; control vs long-term: P < 0.05; short-term vs long-term: P < 0.05).Serum SOD activity exhibited a dose-dependent decline: Significant reductions were detected in control vs short-term (P < 0.05) and control vs long-term groups (P < 0.05).
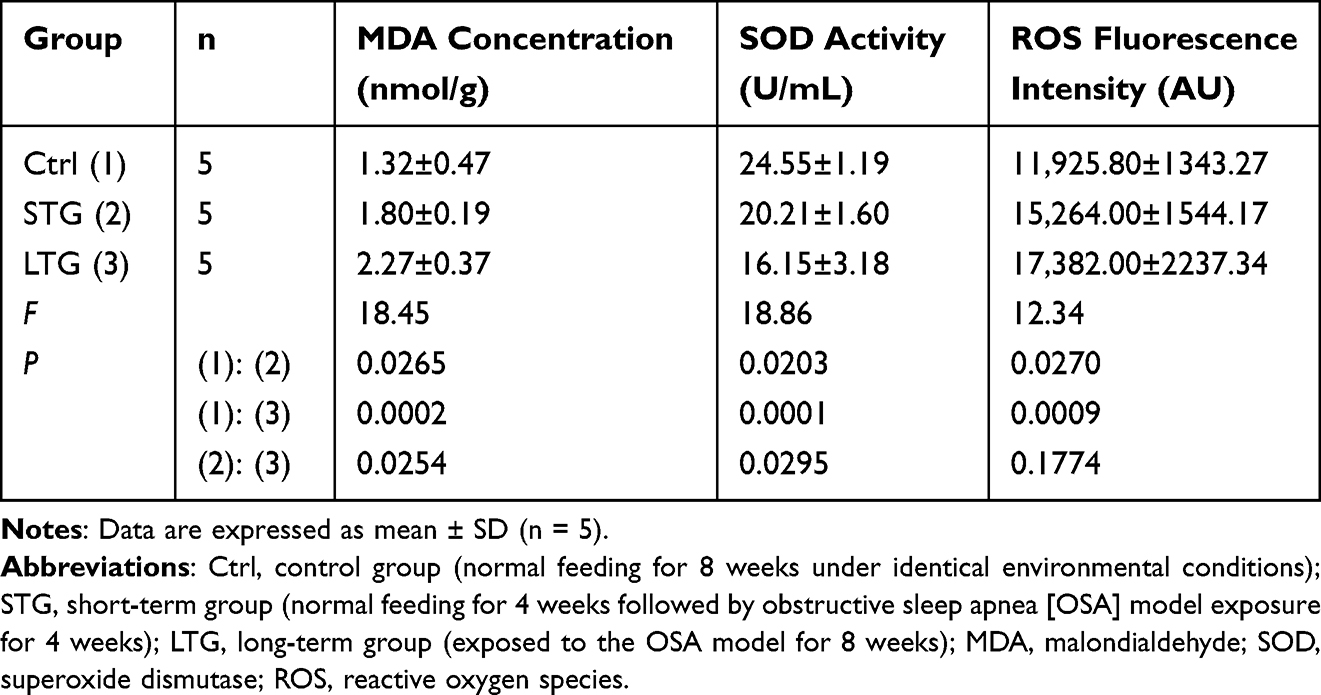 |
Table 2 Serum Levels of MDA, Superoxide Dismutase SOD, and Reactive Oxygen Species ROS in Experimental Groups |
 |
Figure 2 Serum levels of MDA, SOD activity, and ROS in three experimental groups. Abbreviations: MDA, malondialdehyde; SOD, superoxide dismutase; ROS, reactive oxygen species. Notes: control group, normal feeding for 8 weeks under identical environmental conditions; short-term group, normal feeding for 4 weeks followed by obstructive sleep apnea (OSA) model exposure for 4 weeks; long-term group,exposed to the OSA model for 8 weeks; *, P< 0.05; ns, P≥0.05. |
No significant difference was observed between short-term and long-term groups (P > 0.05).
These findings collectively indicate elevated systemic oxidative stress in rats with sleep apnea, characterized by exacerbated lipid peroxidation (MDA ↑), accumulated reactive oxygen species (ROS ↑), and impaired antioxidant capacity (SOD ↓).
Histopathological Alterations in Endometrial Tissues Across Experimental Groups: Endometrial Thickness, Vascular Density, and Glandular Number
As shown in Table 3, Figures 3, and 4, the endometrial thickness of rats in the control group, short-term group, and long-term group decreased sequentially, with intergroup comparisons showing significant differences (all P < 0.05). Similarly, the number of endometrial blood vessels in these groups also declined progressively, and statistically significant differences were observed between all groups (P < 0.05). For the number of endometrial glands, a significant reduction was noted between the control group and the long-term group (P < 0.05), whereas no significant differences were detected between the control group and the short-term group or between the short-term group and the long-term group (P > 0.05). These results indicate that sleep apnea leads to a decline in endometrial thickness, vascular density, and glandular number in rats.
 |
Table 3 Comparative Analysis of Endometrial Structural Parameters Across Experimental Groups |
 |
Figure 3 Representative hematoxylin and eosin (H&E)-stained endometrial sections. Notes: (a) control group (normal feeding for 8 weeks under identical environmental conditions); (b) short-term group (normal feeding for 4 weeks followed by obstructive sleep apnea [OSA] model exposure for 4 weeks); (c) long-term group (exposed to the OSA model for 8 weeks). Endometrial vessels and glands are indicated by red and white arrows, respectively. Scale bar: 100 μm. |
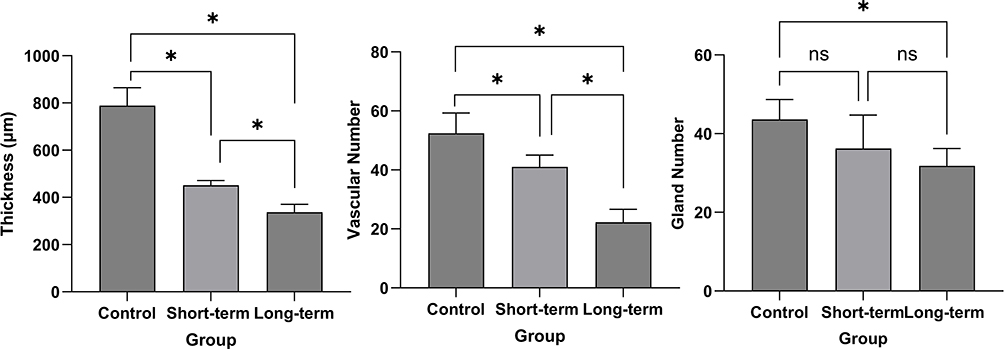 |
Figure 4 Structural parameters of the endometrium in three groups of rats. Notes: control group, normal feeding for 8 weeks under identical environmental conditions; short-term group, normal feeding for 4 weeks followed by obstructive sleep apnea (OSA) model exposure for 4 weeks; long-term group,exposed to the OSA model for 8 weeks; *, P < 0.05; ns, P≥0.05. |
Expression Levels of Ki-67 and VEGF in the Endometrium of Rats Across Experimental Groups
As illustrated in Figures 5–7, and Table 4, the expression rates of Ki-67 and VEGF in the endometrium of rats decreased sequentially across the control group, short-term group, and long-term group. For Ki-67, significant differences were observed between the control group and the short-term group, as well as the control group and the long-term group, with intergroup comparisons showing significant differences (both P < 0.05).
 |
Table 4 Expression Rates of Ki-67 and VEGF in the Endometrium of Rats Across Experimental Groups |
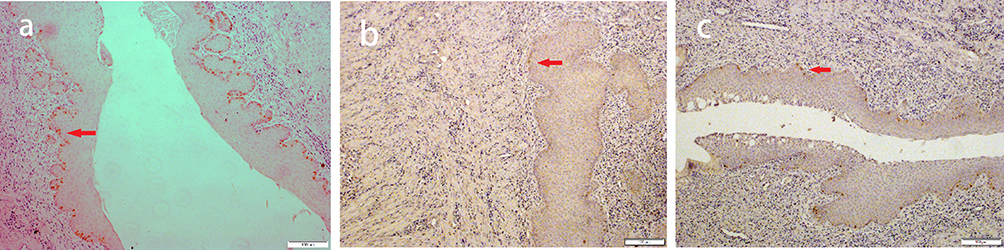 |
Figure 5 Immunohistochemical staining of Ki-67 in the endometrium. Abbreviation: Ki-67, Kiel 67. Notes: (a) control group (normal feeding for 8 weeks under identical environmental conditions); (b) short-term group (normal feeding for 4 weeks followed by obstructive sleep apnea [OSA] model exposure for 4 weeks); (c) long-term group (exposed to the OSA model for 8 weeks). Ki-67+ (Kiel 67 antigen positive) nuclei (brown) indicated by red arrows. Scale bar: 100 μm. |
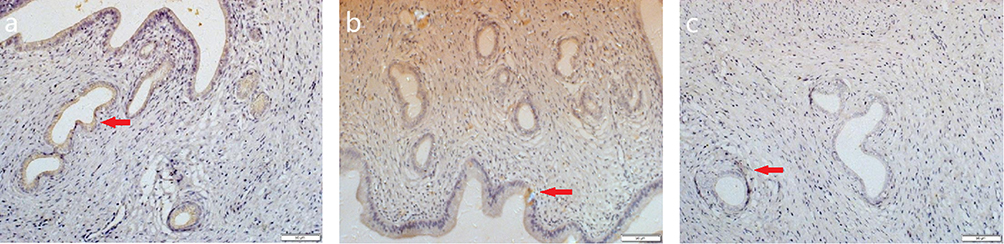 |
Figure 6 Immunohistochemical staining of VEGF in the endometrium. Abbreviation: VEGF,vascular endothelial growth factor. Notes: (a) control group (normal feeding for 8 weeks under identical environmental conditions); (b) short-term group (normal feeding for 4 weeks followed by OSA model exposure for 4 weeks); (c) long-term group (exposed to the OSA model for 8 weeks). VEGF+ (vascular endothelial growth factor positive) cells (brown cytoplasmic granules) are indicated by red arrows. Scale bar: 50 μm. |
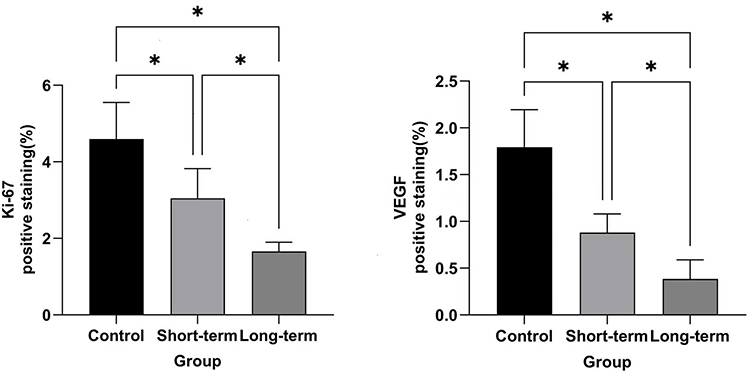 |
Figure 7 Expression rates of Ki-67 and VEGF in the endometrium of rats across experimental groups. Abbreviations: Ki-67, Kiel 67; VEGF, vascular endothelial growth factor. Notes: control group, normal feeding for 8 weeks under identical environmental conditions; short-term group, normal feeding for 4 weeks followed by obstructive sleep apnea (OSA) model exposure for 4 weeks; long-term group,exposed to the OSA model for 8 weeks; *, P< 0.05.r. |
Apoptosis Levels in the Endometrial Tissue of Rats Across Experimental Groups
As illustrated in Table 5 and Figures 8 and 9, the apoptosis levels in the endometrium of rats progressively increased across the control group, short-term group, and long-term group. Significant differences were observed between the control group and the short-term group, as well as the control group and the long-term group, with intergroup comparisons showing significant differences (both P < 0.05). However, no significant difference was detected between the short-term group and the long-term group (P > 0.05). Notably, sleep apnea resulted in elevated endometrial apoptosis levels in rats.
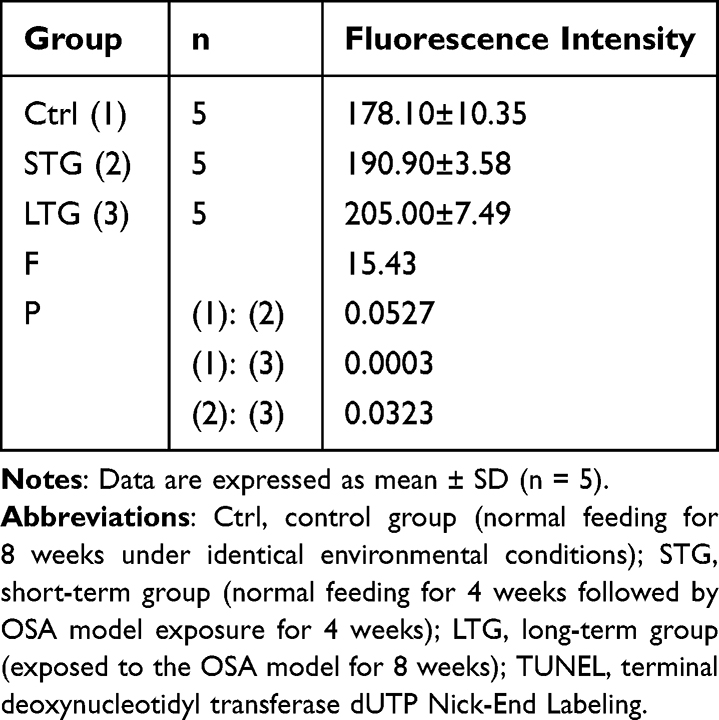 |
Table 5 TUNEL Fluorescence Intensity in Uterine Tissue Across Experimental Groups |
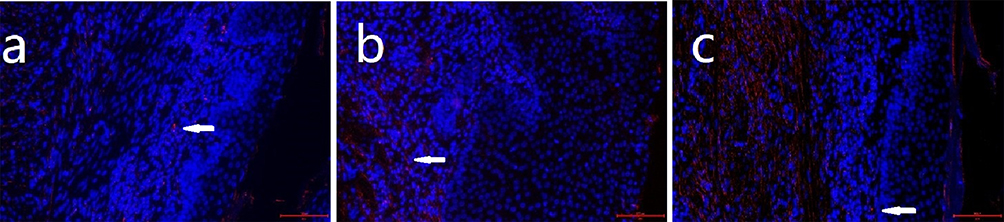 |
Figure 8 Apoptosis level of endometrial cells in three groups of rats. Notes: (a) control group (normal feeding for 8 weeks under identical environmental conditions); (b) short-term group (normal feeding for 4 weeks followed by obstructive sleep apnea [OSA] model exposure for 4 weeks); (c) long-term group (exposed to the OSA model for 8 weeks). TUNEL+ (Terminal deoxynucleotidyl transferase dUTP nick-end labeling positive) nuclei (red, indicated by white arrowheads) indicate apoptotic cells. Nuclei were counterstained with DAPI (4′,6-diamidino-2-phenylindole) (blue). Scale bar: 200 μm. |
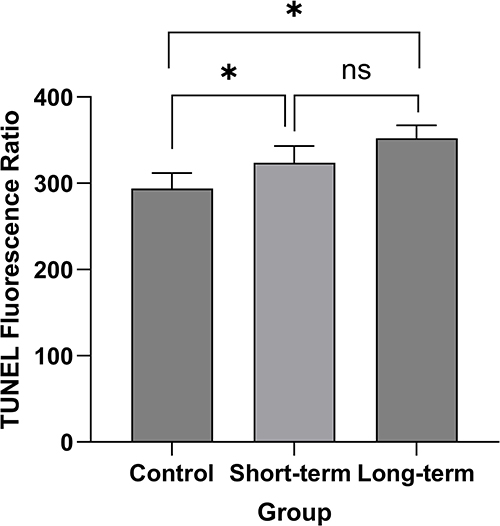 |
Figure 9 Apoptosis level in the endometrium of rats across three experimental groups. Notes: control group, normal feeding for 8 weeks under identical environmental conditions; short-term group, normal feeding for 4 weeks followed by obstructive sleep apnea (OSA) model exposure for 4 weeks; long-term group, exposed to the OSA model for 8 weeks; *, P< 0.05; ns, P≥ 0.05. |
Discussion
Obstructive Sleep Apnea Hypopnea Syndrome (OSAHS) is a disorder characterized by nocturnal snoring, recurrent awakenings, apnea episodes, daytime fatigue, and excessive somnolence. It induces multiple pathophysiological alterations, including recurrent hypoxia, hypercapnia, disruption of sleep architecture, and increased sympathetic nerve activity, leading to chronic hypoxia, sleep fragmentation, and systemic oxidative stress. According to a US survey, the estimated prevalence of at least mild OSA among women aged 30–70 years is 5%, while moderate to severe OSA affects approximately 6% of this population.10 Under OSAHS conditions, the imbalance between oxidative and antioxidant systems results in elevated levels of ROS, which damage lipids, proteins, and deoxyribonucleic acid (DNA), thereby impairing cellular function.
In this study, we established a chronic intermittent hypoxia (CIH) model to simulate OSA. This non-invasive, anesthesia-free model allows precise control of gas concentrations within the chamber, ensuring safe, reproducible, and standardized experimental conditions. Currently, it is the most widely utilized animal model for investigating OSA-related mechanisms.
Effects of OSA on Body Weight and Uterine Index in Female Rats
OSA and Body Weight in Female Rats: Relationship and Underlying Mechanisms
We demonstrate that OSA induces increased body weight and decreased uterine index in rats. While obesity is a recognized risk factor for OSA, our findings highlight a critical bidirectional relationship where OSA actively promotes weight gain through metabolic dysregulation.11,12 Specifically, OSA-induced intermittent hypoxia triggers neuroendocrine disturbances: sleep fragmentation impairs insulin sensitivity and activates central reward pathways, leading to hyperphagia disproportionate to energy expenditure. This aligns with clinical evidence indicating that OSA independently contributes to obesity progression, as demonstrated by elevated weight gain in patients compared to BMI-matched controls.
This metabolic disruption is further compounded by hypoxia-driven pathways, including: Hypoxia-Inducible Factor 1 Alpha (HIF-1α) activation, which enhances hepatic gluconeogenesis and ectopic lipid accumulation; Mitochondrial dysfunction, reducing lipid oxidative capacity; Systemic inflammation (eg, Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells [NF-κB]-mediated tumor necrosis factor alpha [TNF-α]/lnterleukin-6[IL-6] release) exacerbating insulin resistance.13–18
Although obesity contributes to OSA pathogenesis via pharyngeal adipose deposition and reduced lung volume (as summarized below), our experimental focus remains on OSA-initiated metabolic sequelae.19 The observed weight gain in rats substantiates that OSA is not merely a consequence of but a driver for adiposity, establishing a self-perpetuating cycle.20 This framework explains our key finding: OSA directly contributes to impaired reproductive function via metabolic dysregulation and weight gain.
OSA and Uterine Index: Correlation and Underlying Mechanisms
This study revealed that OSA significantly reduces uterine index (p<0.0001), suggesting compromised uterine morphogenesis and indicating potential fertility impairment in female rats. Oxidative stress critically modulates uterine developmental processes and endometrial homeostasis through coordinated regulation of cellular proliferation/apoptosis balance, angiogenic remodeling, and inflammatory cascade activation. Emerging evidence has established close associations between oxidative stress and the pathogenesis of endometriosis, recurrent pregnancy loss, and uterine developmental anomalies. Uterine development relies on stromal-epithelial interactions and hormonal signaling (eg, estrogen and progesterone). Animal studies demonstrate that aberrant ROS levels during embryogenesis disrupt uterine gland formation and myometrial differentiation. For instance, nuclear factor erythroid 2-related factor 2 (Nrf2)-knockout mice exhibit diminished uterine gland density and stromal fibrosis due to attenuated antioxidant defenses, demonstrating that redox imbalance directly compromises uterine morphogenesis.21 Wang et al demonstrated that microwave radiation exposure induces elevated oxidative stress in female mice, resulting in decreased uterine index and suppressed uterine growth under oxidative conditions.22 These findings align with the pathological outcomes observed in our study.
Impact of Obstructive Sleep Apnea on Systemic Redox Status in Female Rats
ROS are chemically defined as oxygen-containing, chemically reactive entities generated endogenously or exogenously, which collectively represent the oxidative stress burden of an organism through their redox activity.²³ SOD constitutes a ubiquitous family of metalloenzymes that have been evolutionarily conserved. By catalyzing the dismutation of the superoxide anion radical (O₂•−) into molecular oxygen and hydrogen peroxide, SOD serves as a primary defense mechanism against oxidative damage. This enzymatic activity maintains metabolic homeostasis, neutralizes free radical-mediated cellular injury, and facilitates cellular repair processes.23 Our experimental data demonstrate that OSA exposure significantly disrupts redox homeostasis in rats, manifested by elevated ROS levels, suppressed SOD activity, and increased MDA production (Table 2 and Figure 2). This triad of biomarkers mechanistically accounts for the observed endometrial pathology, where ROS overproduction (45.75% increase versus controls; P=0.0009) directly induces glandular atrophy via oxidative DNA damage in epithelial cells. SOD reduction (34.22% decrease versus controls; P=0.0001) compromises the primary antioxidant defense, exacerbating membrane lipid peroxidation as quantitatively measured by MDA accumulation (71.97% increase versus controls; P=0.0002).23,24
These findings align with the established pathophysiology of OSA in humans: recurrent apneic events generate intermittent hypoxia-reoxygenation cycles that uncouple mitochondrial respiration, triggering superoxide bursts during reoxygenation phases.25,26 This process recapitulates ischemia-reperfusion injury, explaining the observed ROS elevation in our model.6 Crucially, our study provides the first experimental evidence linking this oxidative cascade to uterine impairment. The clinical implications are twofold: the ROS-SOD-MDA axis may serve as a quantifiable biomarker panel for OSA-associated reproductive damage; and therapeutic strategies targeting mitochondrial oxidative stress (eg, SOD mimetics) may protect endometrial integrity. We posit that chronic oxidative injury likely underlies OSA-associated reproductive dysfunction.
Obstructive Sleep Apnea-Induced Endometrial Pathological Injury in Female Rats
This study conclusively demonstrates significant endometrial pathological injury in OSA-model rats, characterized by reduced endometrial thickness, decreased vascular density, and diminished glandular count compared to controls. Histopathological analysis further revealed a substantial increase in apoptotic cells within the OSA group, directly supporting CIH as a key driver of endometrial injury.
Notably, the observed endometrial thinning and microstructural degeneration align with the effects of estrogen depletion. Yang et al previously reported that oxidative stress-mediated pathways induce excessive apoptosis in ovarian granulosa cells and oocytes, precipitating premature ovarian insufficiency (POF) and consequent estrogen reduction.27 Given the critical role of estrogen in endometrial proliferation, our findings are consistent with established estrogen-dependent endometrial maintenance mechanisms.
Endometrial receptivity, fundamentally determined by histological architecture and thickness, constitutes an independent predictor of pregnancy outcomes. Our study identifies oxidative stress as central to OSA-associated endometrial injury: While physiological ROS levels activate pro-proliferative pathways like Wnt/β-catenin, sustained oxidative stress and ROS overload in OSA suppress this pathway, inducing epithelial morphological abnormalities. More critically, ROS activate pro-inflammatory NF-κB and p38 Mitogen-Activated Protein Kinase (MAPK) pathways, triggering cytokine release that inhibits proliferation and promotes apoptosis. This oxidative microenvironment impairs early pregnancy through dual mechanisms: (1) ROS-mediated oxidation of pinopodial surface proteins compromises embryo adhesion capacity; (2) Mitochondrial ROS accumulation in trophoblasts increases their apoptotic susceptibility. These findings resonate with clinical observations: Endometrial thickness <6 mm correlates with significantly reduced implantation rates in assisted reproduction (29.43% vs 41.67% in controls, p<0.01). Although OSA-induced endometrial hypoplasia and elevated apoptosis strongly suggest fertility impairment, definitive causal relationships require validation through future longitudinal studies.
A key finding of this work is the significant suppression of endometrial proliferative activity in OSA-model rats, evidenced by markedly reduced Ki-67 positivity (Figure 2) accompanied by decreased endometrial thickness (P < 0.05). Ki-67 serves as the gold-standard biomarker for cellular proliferation, and its downregulation unequivocally indicates disrupted cell cycle progression in the endometrium.28,29 We postulate two underlying mechanisms: CIH: CIH-induced oxidative stress triggers mitochondrial dysfunction and DNA damage, culminating in cell cycle arrest (eg, G1/S checkpoint activation).30 Hormonal Dysregulation: OSA disrupts the estrogen/progesterone ratio, impairing estrogen-driven proliferative signaling essential for cyclical endometrial regeneration.31 Furthermore, the concurrent decline in proliferative activity (Ki-67↓) and increase in apoptosis within the OSA group collectively indicate severely compromised endometrial growth dynamics. This pathophysiological cascade likely contributes to fertility impairment in female rats by diminishing endometrial receptivity.
Another significant finding is the substantial reduction in endometrial VEGF expression and vascular density in OSA-model rats. VEGF plays a pivotal role in endometrial angiogenesis and vascular permeability maintenance.32,33 Its insufficient expression inevitably suppresses neovascularization, reduces microvascular density, and impairs blood perfusion, thereby collectively impairing endometrial receptivity and contributing to embryo implantation failure. Combined with observed proliferative suppression, these results strongly suggest OSA impairs endometrial function through dual pathways: directly inhibiting cellular proliferation (manifested as reduced Ki-67 expression) and downregulating the pro-angiogenic factor VEGF.
Synthesizing these findings, OSA exacerbates endometrial proliferative suppression by inhibiting VEGF signaling. This combined effect likely impedes embryo implantation by compromising endometrial receptivity, ultimately leading to reduced fertility. These insights not only deepen the understanding of mechanisms underlying OSA-associated reproductive injury but also suggest that ROS levels, Ki-67 expression, and VEGF signaling may serve as a potential biomarker panel for assessing endometrial damage.
Limitations and Future Perspectives
This study employed a single model approach involving nitrogen gas infusion to reduce oxygen concentration in the experimental chamber, thereby establishing an intermittent hypoxia model. However, potential confounding factors may exist due to undefined biological effects of nitrogen gas exposure. Future studies should validate these findings using alternative OSA animal models (eg, chronic intermittent hypoxia chambers or tracheal occlusion techniques). Additionally, while we quantified systemic oxidative stress markers in serum, subsequent investigations should measure organ-specific oxidative stress parameters (eg, ovarian and endometrial tissue levels of ROS, SOD activity, and MDA concentration) to strengthen the evidentiary chain.
Conclusion
Our study demonstrates that OSA-like intermittent hypoxia induces systemic oxidative stress, endometrial hypoplasia, and suppressed cellular proliferation in female rats. These findings underscore the potential role of OSA in female reproductive dysfunction and highlight the need for clinical investigations into endometrial health in OSA-affected women. Future research should prioritize translational approaches, including antioxidant interventions (eg, N-acetylcysteine) and hormonal therapies, to mitigate OSA-related endometrial damage.
Data Sharing Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Statement of Ethics Approval
This study was approved by the Ethics Committee of Tianjin Haihe Hospital (Approval No. 2025HHWZ(A)-003). Our study adheres to the principles of the Declaration of Helsinki.
Author Contributions
Dong Zhang: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing-review and editing. Wenli Bian: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing-Original Draft, Writing-review and editing. Zhihua Gao: Data curation, Formal analysis, Investigation, Project administration, Resources, Software, Supervision, Validation, Writing-Original Draft, Writing-review and editing. All authors have reviewed and agreed on all versions of the article before submission, during revision, the final version accepted for publication, and any significant changes introduced at the proofing stage. They have agreed on the journal to which the article will be submitted and agree to take responsibility and be accountable for the contents of the article.
Funding
This study was funded by Science and Technology Planning Project of Tianjin Municipal Education Commission (No.2022YGYB15).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lavie L. Oxidative stress in obstructive sleep apnea and intermittent hypoxia--revisited--the bad ugly and good: implications to the heart and brain. sleep medicine. Reviews. 2015;20:27–45.doi:10.1016/j.smrv.2014.07.003.
2. Peppard PE, Young T, Barnet JH, et al. Increased prevalence of sleep-disordered breathing in adults. American. Journal of Epidemiology.2013. 177(9):1006–1014. doi:10.1093/aje/kws342
3. Franklin KA, Lindberg E. Obstructive sleep apnea is a common disorder in the population-a review on the epidemiology of sleep apnea. Journal of Thoracic. Disease. 2015;7(8):1311–1322. doi:10.3978/j.issn.2072-1439.2015.06.11
4. Beverly Hery CM, Hale L, Naughton MJ. Contributions of the Women’s Health Initiative to understanding associations between sleep duration, insomnia symptoms, and sleep-disordered breathing across a range of health outcomes in postmenopausal women. Sleep Health. 2020;6(1):48–59. doi:10.1016/j.sleh.2019.09.005
5. Ma LS, Liu CY. Research status and progress of obstructive sleep apnea. Journal of Hebei Medical University. 2021;42(5):609–612. doi:10.3969/j.issn.1007-3205.2021.05.023
6. Maniaci A, Iannella G, Cocuzza S, et al. Oxidative stress and inflammation biomarker expression in obstructive sleep apnea patients. J clin Med. 2021;10(2):282. doi:10.3390/jcm10020277
7. Gao W, Feng F, Ma X, et al. Progress of oxidative stress in endometrium decidualization. J Obstetrics Gynaecol. 2022;42(8):3429–3434. doi:10.1080/01443615
8. Vollert J, Schenker E, Macleod M, et al. EQIPD WP3 study group members. Systematic review of guidelines for internal validity in the design, conduct and analysis of preclinical biomedical experiments involving laboratory animals. BMJ Open Sci. 2020;4(1):e100046. doi:10.1136/bmjos-2019-100046
9. Shen YB, Ou XW, Liu S. Research progress on animal models of obstructive sleep apnea. Journal of Shanghai Jiaotong University. 2024;44(4):501–508. doi:10.3969/j.issn.1674-8115.2024.04.011
10. Mangione CM, Barry MJ, Nicholson WK, et al. US Preventive Services Task Force, Mangione, C M. Screening for obstructive sleep apnea in adults: us preventive services task force recommendation statement. JAMA. 2022;328(19):1945–1950. doi:10.1001/jama.2022.20304.
11. Young T, Palta M, Dempsey J, et al. The occurrence of sleep-disordered breathing among middle-aged adults. New England J Med. 1993;328(17):1230–1235. doi:10.1056/NEJM199304293281704
12. Schwartz AR, Patil SP, Laffan AM, et al. Obesity and obstructive sleep apnea: pathogenic mechanisms and therapeutic approaches. Proceedings Am Thoracic Soc. 2008;5(2):185–192. doi:10.1513/pats.200708-137MG
13. Vgontzas AN, Papanicolaou DA, Bixler E. O. et al.Sleep apnea and daytime sleepiness and fatigue: relation to visceral obesity, insulin resistance, and hypercytokinemia. J Clin Endocrinol Metabolism. 2000;85(3):1151–1158. doi:10.1210/jcem.85.3.6484
14. Arnardottir ES, Bjornsdottir E, Olafsdottir K, Benediktsdottir B, Gislason T. A. et al.Obstructive sleep apnoea in the general population: highly prevalent but minimal symptoms. Eur Respir J. 2016;47(1):194–202. doi:10.1183/13993003.01148-2015
15. Lf D, Li J, Reinke C, et al. Intermittent hypoxia exacerbates metabolic effects of diet-induced obesity. Obesity. 2011;19(11):2167–2174. doi:10.1038/oby.2011.240
16. Zhang H, Gao P, Fukuda R, et al. HIF-1 inhibits mitochondrial biogenesis and cellular respiration in VHL-deficient renal cell carcinoma by repression of C-MYC activity. Cancer Cell. 2007;11(5):407–420. doi:10.1016/j.ccr.2007.04.001
17. Koh JH, Hancock CR, Terada S, et al. PPARβ is essential for maintaining normal levels of PGC-1α and mitochondria and for the increase in muscle mitochondria induced by exercise. Cell Metabolism. 2017;25(5):1176–1185. doi:10.1016/j.cmet.2017.04.029
18. Lee SJ, Zhang J, Choi AM, et al. Mitochondrial dysfunction induces formation of lipid droplets as a generalized response to stress. Oxidative Med Cellular Longevity. 327167;2013. doi:10.1155/2013/327167
19. Xu PC, Zhang H, Cheng Z. M. et al.Pathophysiological connections and therapeutic advances in obstructive sleep apnea syndrome, obesity, and metabolic syndrome. Laboratory Medicine and Clinic. 2024;21(8):1176–1181. doi:10.3969/j.issn.1672-9455.2024.08.032
20. Hawkins MS, Levine MD, Buysse DJ, et al. Sleep health characteristics among adults who attempted weight loss in the past year: NHANES 2017-2018. International. Journal of Environmental Research and PublicHealth. 2021;18(19):10170. doi:10.3390/ijerph181910170
21. Jakaria M, Azam S, Haque ME, et al. Taurine and its analogs in neurological disorders: focus on therapeutic potential and molecular mechanisms. Redox Biology. 2019;24:101223. doi:10.1016/j.redox.2019.101223
22. Wang JJ, Chen SQ, Dong J, et al. Short-term effects of microwave radiation on ovarian function in female mice. Journal of Chinese PLA MedicalCollege. 2021;42(11):1167–1171. doi:10.3969/j.issn.1672-9455.2024.08.032
23. Chen Y. Recent research on ROS, SOD, and GSH in diabetic vascular complications. J Jiangxi Unive Traditional Chin Med. 2013;25(3):93–97. doi:10.3969/j.issn.1005-9431.2013.03.031
24. Deng HP, Yu SY, Chen YQ, et al. Effects of X-ray radiation on body weight, fur growth, and SOD, CAT activities and MDA content in liver and kidney tissues of neonatal mice. Acta Zoologica Sinica. 2008;54(6):1029–1037.
25. Semenza GL. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Ann Rev Pathol. 2014;9:47–71. doi:10.1146/annurev-pathol-012513-104720
26. Hirte H, Yao X, Ferguson SE, et al. An Ontario Health (Cancer Care Ontario) clinical practice guideline: consolidation or maintenance systemic therapy for newly diagnosed stage II, III, or IV epithelial ovary, fallopian tube, or primary peritoneal carcinoma. Curr Oncol. 2021;28(2):1114–1124. doi:10.3390/curroncol28020107
27. Yang KF, Li L, Zhou H, et al. Research progress on regulation of oxidative stress, autophagy, and apoptosis in premature ovarian failure. J Hunan Univ Chin Med. 2021;41(5):809–814. doi:10.3969/j.issn.1674-070X.2021.05.031
28. Gerdes J, Lemke H, Baisch H, et al. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. The. J Immunol. 1984;133(4):1710–1715. doi:10.4049/jimmunol.133.4.1710
29. Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. J Cell Physiol. 2000;182(3):311–322.
30. Endl E, Gerdes J. Posttranslational modifications of the KI-67 protein coincide with two major checkpoints during mitosis. J Cell Physiol. 2000;182(3):371–380. doi:10.1002/(SICI)1097-4652(200003)182:3<371::AID-JCP8>3.0.CO;2-J
31. Linz D, Linz B, Hohl M, et al. Atrial arrhythmogenesis in obstructive sleep apnea: therapeutic implications. Sleep Med Rev. 2016;26:87–94. doi:10.1016/j.smrv.2015.03.003
32. Dey SK, Lim H, Das SK, et al. Molecular cues to implantation. Endocrine Rev. 2004;25(3):341–373. doi:10.1210/er.2003-0020
33. AbdelHafez F, Bedaiwy M, El-Nashar SA, et al. Techniques for cryopreservation of individual or small numbers of human spermatozoa: a systematic review. Human Reproduction Update. 2009;15(2):153–164. doi:10.1093/humupd/dmn061
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.