Back to Journals » Journal of Inflammation Research » Volume 18
Impact of Echinococcus granulosus Cyst Fluid on the Phenotypic Dynamics and Degranulation of RBL-2H3 Mast Cells
Authors Yue Z, Zhou JR, Wang CS, Pu XL, Parehati G, Cao LW, Bayina B, Ye JR
Received 13 February 2025
Accepted for publication 14 July 2025
Published 24 July 2025 Volume 2025:18 Pages 9837—9848
DOI https://doi.org/10.2147/JIR.S522640
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Anish R. Maskey
Zhen Yue,1,2,* Jing-Ru Zhou,3,* Chun-Sheng Wang,1 Xue-Li Pu,1 Guligeiya Parehati,1 Li-Wei Cao,1 Batesurong Bayina,1 Jian-Rong Ye1
1Department of Anesthesiology, The First Affiliated Hospital of Xinjiang Medical University, Urumqi, 830054, People’s Republic of China; 2Department of Anesthesiology, Changji People’s Hospital, Changji, 831100, People’s Republic of China; 3Department of Anesthesiology, The Sixth Affiliated Hospital of Xinjiang Medical University, Urumqi, 830063, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jian-Rong Ye, Department of Anesthesiology, The First Affiliated Hospital of Xinjiang Medical University, No. 137, Liyushan South Road, Urumqi, 830054, People’s Republic of China, Tel +86 18999938777, Email [email protected]
Objective: This study aimed to evaluate the effects of Echinococcus granulosus cyst fluid (EgCF) on the phenotypic characteristics and degranulation activity of mast cells during echinococcosis-associated hypersensitivity reactions, using the RBL-2H3 rat basophilic leukemia cell line as an in vitro model.
Methods: The cytotoxic effects of EgCF on RBL-2H3 cells were assessed using the Cell Counting Kit-8 assay. Degranulation activity was quantified by measuring β-hexosaminidase release. Morphological alterations and cytoskeletal changes were evaluated via toluidine blue staining and phalloidin-labeled confocal laser scanning microscopy. Apoptotic cell populations were quantified using flow cytometry.
Results: EgCF at mass concentrations ranging from 0.5 to 5 mg/mL significantly enhanced cell proliferation. Higher EgCF concentrations were associated with increased morphological deformation, as evidenced by toluidine blue staining, and with significantly elevated β-hexosaminidase release compared to the control group (p < 0.01). Flow cytometry revealed a statistically significant increase in apoptosis rates in the treated groups (p < 0.01). Within 30 minutes of EgCF exposure, phalloidin staining showed notable cytoskeletal reorganization, including cellular wrinkling and pseudopodia extension. These morphological changes were partially reversed within 45 to 60 minutes.
Conclusion: EgCF induces morphological transformation and promotes degranulation in mast cells, with the intensity of these responses correlating positively with EgCF concentration. These findings suggest a dose-dependent role of EgCF in modulating mast cell activity during parasitic hypersensitivity responses.
Keywords: allergic hypersensitivity, Echinococcus granulosus cyst fluid, mast cell degranulation, parasitic infection, RBL-2H3 cells
Introduction
Cystic echinococcosis (CE) is a zoonotic parasitic disease with high endemicity in pastoral regions of northwest China. The condition is primarily caused by infection with E. granulosus, resulting in the development of hydatid cysts within intermediate hosts.1–3 Physical trauma, such as blunt force injury or surgical manipulation, may cause cyst rupture and subsequent leakage of cystic fluid, which can precipitate acute hypersensitivity reactions that are potentially life-threatening if not addressed promptly.4,5
The pathophysiology of echinococcosis-associated hypersensitivity involves complex immune mechanisms mediated by various effector cells. Among these, mast cells, located abundantly in perivascular tissues, connective tissues, and mucosal surfaces, play a central role in the host immune response during parasitic infections.6,7 These cells contribute to immune regulation primarily through degranulation, releasing proinflammatory mediators that rapidly exert effects on local tissues and facilitate the recruitment of additional immune cells.8,9
Mast cell activation often occurs via antibody-dependent mechanisms. Upon pathogen exposure, B cells are stimulated to produce immunoglobulin E (IgE), which subsequently binds to high-affinity IgE receptors (FcεRI) on mast cell surfaces. This interaction upregulates receptor expression and results in sensitization of the mast cells. Upon re-exposure to the same antigen, the FcεRI-bound IgE recognizes the antigen, triggering mast cell activation and subsequent degranulation.10–12 This degranulation process, characterized by morphological changes and the release of chemokines, cytokines, and lipid mediators, is a hallmark of allergic and inflammatory responses.13
Following E. granulosus infection, antigen-specific IgE binds to FcεRI on mast cells, rendering them sensitized. Rupture of hydatid cysts, whether spontaneous or surgically induced, can activate these sensitized mast cells, leading to a massive release of immunomodulatory substances. This cascade may result in systemic allergic responses, including anaphylactic shock, which can be fatal. Consequently, investigating the mechanisms underlying mast cell-mediated allergic responses in echinococcosis is of critical clinical importance.
To investigate the effects of E. granulosus cyst fluid (EgCF) on mast cell morphology and activation, this study utilized RBL-2H3 cells, a rat basophilic leukemia cell line widely employed as an in vitro model for mast cell degranulation due to its functional similarity to native mast cells.14 In this study, IgE-sensitized RBL-2H3 cells were exposed to varying concentrations of EgCF. The objective was to assess the impact of EgCF on cell morphology, cytoskeletal reorganization, and degranulation dynamics. Findings from this investigation may inform therapeutic strategies and provide mechanistic insights into the prevention of hypersensitivity reactions in individuals with cystic echinococcosis.
Materials and Methods
Experimental Material
Cell Line
The rat basophilic leukemia cell line (RBL-2H3) was obtained from Procell Life Science & Technology Co., Ltd. (China).
Main Reagents
Dinitrophenyl-immunoglobulin E (DNP-IgE; Batch No. D8406) and Compound 48/80 (Batch No. C2313) were procured from Sigma-Aldrich (USA). Toluidine blue (Batch No. 1002341393) and 4-Nitrophenyl N-acetyl-D-glucosaminide (Batch No. S10146) were utilized as staining and enzymatic substrates, respectively. TRITC-phalloidin (Batch No. CA1610) was obtained from Beijing Solarbio Science & Technology Co., Ltd. (China). The flow cytometry apoptosis detection kit (Batch No. BB-4101) was purchased from Bestbio (China). The bicinchoninic acid (BCA) protein assay kit was supplied by Thermo Fisher Scientific (USA). Fetal bovine serum (FBS; Batch No. A5669801) was acquired from Gibco (USA).
Equipment
Cell culture was conducted in an incubator from Thermo Fisher Scientific (USA). Microscopic observations were made using an inverted microscope from Leica (Germany). Absorbance measurements were performed using a multifunctional microplate reader from Thermo Fisher Scientific (USA). Cytoskeletal imaging was conducted with a laser confocal scanning microscope (Model 1570000) from Nikon (Japan). Apoptosis analyses were carried out using a flow cytometer (Model LSR-II) from Bestbio (China).
Cell Treatment Methods
Cell Culture
Prior to cell culture, pipettes and related laboratory equipment were placed on a clean bench that had been disinfected with alcohol and exposed to ultraviolet (UV) light for 30 minutes. RBL-2H3 cells previously stored at –80 °C were rapidly thawed in a 37 °C water bath with gentle agitation. The thawed cells were transferred into Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 15% FBS, inoculated into T25 culture flasks, and incubated at 37 °C in a humidified atmosphere containing 5% CO2. Cells were passaged every 2 to 3 days at a 1:3 ratio. Cells in the logarithmic growth phase were selected for subsequent experimental procedures.
Preparation and Storage of EgCF
Fresh sheep liver infected with E. granulosus was obtained from a local slaughterhouse. The surface of the liver was disinfected by wiping with alcohol for 3 minutes. Cyst fluid was aspirated aseptically from the hydatid cysts using a sterile syringe and transferred into sterile centrifuge tubes. The fluid was centrifuged at 4 °C at 2500 rpm for 10 minutes. The supernatant was collected, and the sediment was discarded.
Approximately 10 mL of the supernatant was transferred into a sterile Petri dish placed within a clean bench previously disinfected with UV light for 30 minutes. The dish was sealed with plastic wrap and subjected to lyophilization using a freeze dryer for 24 hours. The lyophilized cyst fluid was then reconstituted in sterile phosphate-buffered saline (PBS). Protein concentration was determined using a BCA protein assay kit, and the solution was diluted to the desired concentration with PBS. Remaining samples were aliquoted and stored at –80 °C until further use.
Assessment of Cell Viability Following EgCF Exposure Using the CCK8 Assay
RBL-2H3 cells in the logarithmic growth phase were harvested using 0.25% trypsin and processed under aseptic conditions in a UV-sterilized clean bench. The resulting cell suspension was centrifuged at 4 °C at 1000 rpm for 5 minutes. The supernatant was discarded, and the cell pellet was resuspended in complete culture medium and counted. Cells were seeded into 96-well plates at a density of 5 × 10³ cells per well and incubated at 37 °C in a humidified 5% CO2 incubator for 12 hours.
The experimental design included four groups: blank control group, control group, positive control groups, and experimental groups, each with three replicates. The blank group contained culture medium and CCK-8 reagent without cells to serve as a background reference. The control group consisted of cells cultured in complete medium without any treatment, representing baseline viability. The positive control groups were treated with Compound 48/80 (C48/80) at final concentrations of 25 µg/mL and 50 µg/mL. The experimental groups received EgCF at final concentrations of 0.5, 1, 2, 3, 4, and 5 mg/mL.
After 12 hours of incubation, 10 µL of Cell Counting Kit-8 (CCK-8) solution was added to each well, followed by a 30-minute incubation. Optical density (OD) values were measured at 450 nm using a microplate reader.
The assay was performed in triplicate at different time points. Cell viability was calculated using the following formula:
Cell viability = (Experimental group’s OD value - Blank group’s OD value) / (Control group’s OD value -Blank group’s OD value) × 100%.
Assessment of Cell Morphology Using Toluidine Blue Staining
RBL-2H3 cells in the logarithmic growth phase were harvested, treated with 0.25% trypsin, centrifuged, and resuspended in complete medium. Cells were seeded into 6-well plates at a density of 5 × 105 cells per well. EgCF was added to achieve final concentrations of 0.5, 1, 2, 3, 4, and 5 mg/mL. Following a 60-minute incubation at 37 °C, 20 µL of 0.5% toluidine blue solution was added to each well and incubated for 3 minutes. Cells were then rinsed with PBS and examined under an inverted microscope to assess morphological alterations.
Assessment of β-Hexosaminidase (β-HEX) Release Following EgCF Exposure
RBL-2H3 cells in the logarithmic growth phase were harvested, trypsinized with 0.25% trypsin, centrifuged, and resuspended under aseptic conditions in a UV-sterilized clean bench. Cells were seeded into 24-well plates at a density of 2 × 105; cells per well and incubated for 6 hours to allow adherence. Subsequently, cells were washed with PBS and sensitized overnight with DNP-IgE (0.125 µg/mL) prepared in DMEM containing 1% FBS to maintain cell stability.
The following day, cells were washed three times with PBS and exposed to EgCF at final concentrations of 0.5, 1, 2, 3, 4, and 5 mg/mL for 1 hour. To terminate the reaction, cells were placed on ice for 10 minutes. Supernatants were collected, and in the control group, cells were lysed with 0.5% Triton X-100 on ice for 30 minutes to determine total enzyme release.
Both the cell lysates and supernatants were transferred into sterile, enzyme-free 1.5 mL Eppendorf tubes and centrifuged at 4 °C at 800 × g for 15 minutes. Subsequently, 50 µL of supernatant was transferred to a 96-well plate, followed by the addition of 50 µL of chromogenic substrate. Plates were sealed and incubated for 2 hours. After incubation, 200 µL of stop solution was added to each well, and optical density (OD) was measured at 405 nm using a microplate reader.
The β-HEX release rate was calculated using the following formula: β-HEX release rate (%) = (Experimental group’s OD - Control group’s OD) / (Total enzyme well’s OD - Control group’s OD) × 100%.
Evaluation of Cytoskeletal Alterations Following Exposure to EgCF
RBL-2H3 cells in the logarithmic growth phase were seeded into laser confocal culture dishes at a density of 1 × 104 cells per well under aseptic conditions in a UV-sterilized clean bench. After cell adherence, cells were sensitized overnight with DNP-IgE at a concentration of 0.125 µg/mL in complete medium.
The following day, cells were rinsed with PBS and treated with EgCF at a final concentration of 3 mg/mL. At time points of 0, 15, 30, 45, and 60 minutes, the reaction was terminated by placing the corresponding dishes on ice. A positive control group was treated with Compound 48/80 (C48/80) at 50 µg/mL for 60 minutes.
Following treatment, cells were fixed with 4% paraformaldehyde at room temperature, then washed with PBS 2–3 times (10 minutes per wash). Permeabilization was carried out using 0.5% Triton X-100 for 5 minutes, followed by an additional 2–3 PBS washes (10 minutes each). Subsequently, 200 µL of TRITC-phalloidin working solution was added to each well to stain the actin cytoskeleton. Cells were incubated at room temperature in the dark for 30 minutes, followed by three PBS washes (5 minutes each).
To visualize nuclei, cells were stained with 4′,6-diamidino-2-phenylindole at a concentration of 0.1 mM for 30 seconds, then washed with PBS at room temperature. Immediately following staining, cytoskeletal morphology was observed and imaged using a laser confocal scanning microscope.
Evaluation of Apoptosis Following Exposure to EgCF
RBL-2H3 cells in the logarithmic growth phase were seeded into 6-well plates at a density of 1 × 106 cells per well under aseptic conditions in a UV-sterilized clean bench. After cell adherence, cells were sensitized overnight with DNP-IgE at a final concentration of 0.125 µg/mL in complete medium.
The following day, cells were rinsed with PBS and then treated with EgCF at final concentrations of 0.5, 1, 2, 3, 4, and 5 mg/mL. Following a 1-hour incubation at 37 °C, the supernatant was discarded. Cells were subsequently harvested using 0.25% trypsin without ethylenediaminetetraacetic acid and stained according to the manufacturer’s protocol provided with the apoptosis detection kit. Apoptotic cell populations were immediately analyzed using flow cytometry.
Statistical Analysis
Data were analyzed using SPSS software version 29.0, with graphical visualization performed using GraphPad Prism version 8.3. Flow cytometry data were processed and visualized using FlowJo software.
Statistical comparisons were performed using one-way analysis of variance (ANOVA) or the Kruskal–Wallis test, depending on the distribution of the data. Post-hoc analyses for multiple comparisons were conducted using the least significant difference (LSD) test. Normally distributed data are presented as mean ± standard deviation (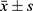 ), while categorical (enumeration) data are expressed as percentages. A p-value of less than 0.05 was considered statistically significant.
), while categorical (enumeration) data are expressed as percentages. A p-value of less than 0.05 was considered statistically significant.
Results
Effects of EgCF on the Viability of RBL-2H3 Cells
Results from the CCK-8 assay (Figure 1) indicated that treatment with EgCF at various concentrations promoted cell proliferation relative to the control group; however, these differences were not statistically significant. In contrast, cells treated with the positive control Compound 48/80 exhibited significantly reduced proliferation compared to the control group (p < 0.001).
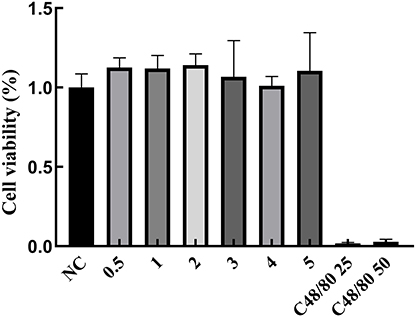 |
Figure 1 The impact of EgCF on the viability of RBL-2H3 cells. Abbreviations: EgCF, Echinococcus granulosus cyst fluid; RBL-2H3, rat basophilic leukemia cells. Note: EgCF concentration (mg/mL); the intervention time of C48 / 80 and EgCF on cells was 12h. |
Toluidine Blue Staining
Morphological assessment via toluidine blue staining revealed that the control group exhibited typical spindle-shaped morphology with intact cell membranes. In contrast, cells exposed to increasing concentrations of EgCF for 60 minutes demonstrated progressive morphological alterations, including a shift toward rounded or polygonal shapes, accompanied by observable degranulation (Figure 2).
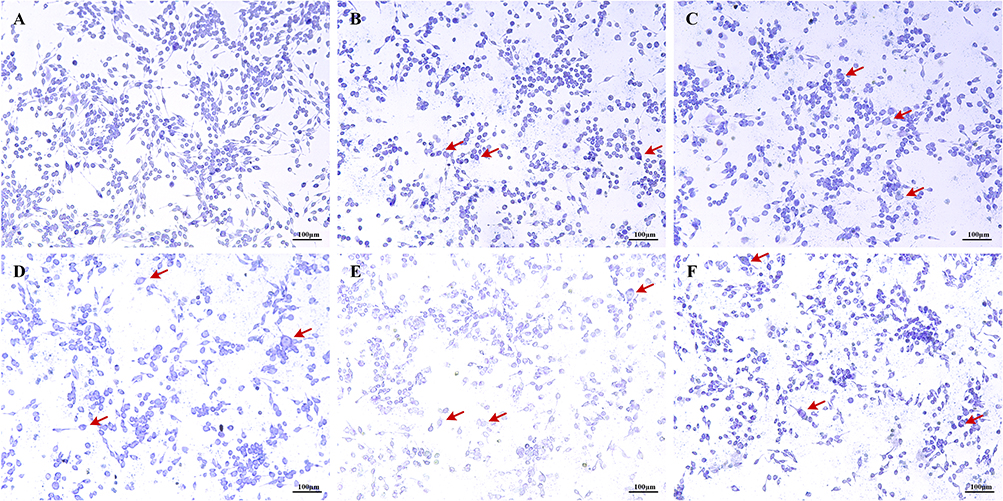 |
Figure 2 Effects of varying EgCF concentrations on cell morphology (200X). Abbreviation: EgCF, Echinococcus granulosus cyst fluid. Note: (A) control group, (B) EgCF0.5mg/mL, (C) group EgCF1mg/mL, (D) group EgCF2mg/mL, (E) group EgCF4mg/mL, (F) group EgCF5mg/mL; C48 / 80 and EgCF intervened cells for 1h; The red arrow points to the deformed cell. |
β-HEX Release Rate
Treatment with the positive control Compound 48/80 led to a marked increase in β-HEX release compared to the control group (p < 0.001) (Figure 3). Similarly, EgCF exposure induced a concentration-dependent elevation in β-HEX release, which was statistically significant at all tested concentrations compared to the control group (p < 0.001).
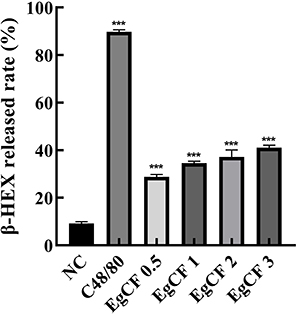 |
Figure 3 Effects of EgCF concentrations on β-HEX release rate. Abbreviations: EgCF, Echinococcus granulosus cyst fluid; β-HEX, β-hexosaminidase. Note: EgCF concentration (mg/mL); *** Delegates compared with the control group, P < 0.001; C48/80 50ug/mL and EgCF 3mg/mL; C48 / 80 and EgCF intervened cells for 1h. |
Effects of EgCF on Cytoskeletal Structure
After 60 minutes of treatment with Compound 48/80, cells in the positive control group exhibited a rounded morphology with signs of membrane rupture and cytoskeletal discontinuity (Figure 4). In the EgCF-treated groups, cells became enlarged and developed distinct morphological changes, including surface wrinkling and the extension of lamellipodia-like pseudopodia. These alterations were most pronounced at the 30-minute time point. With continued incubation, cells gradually regained their original morphology.
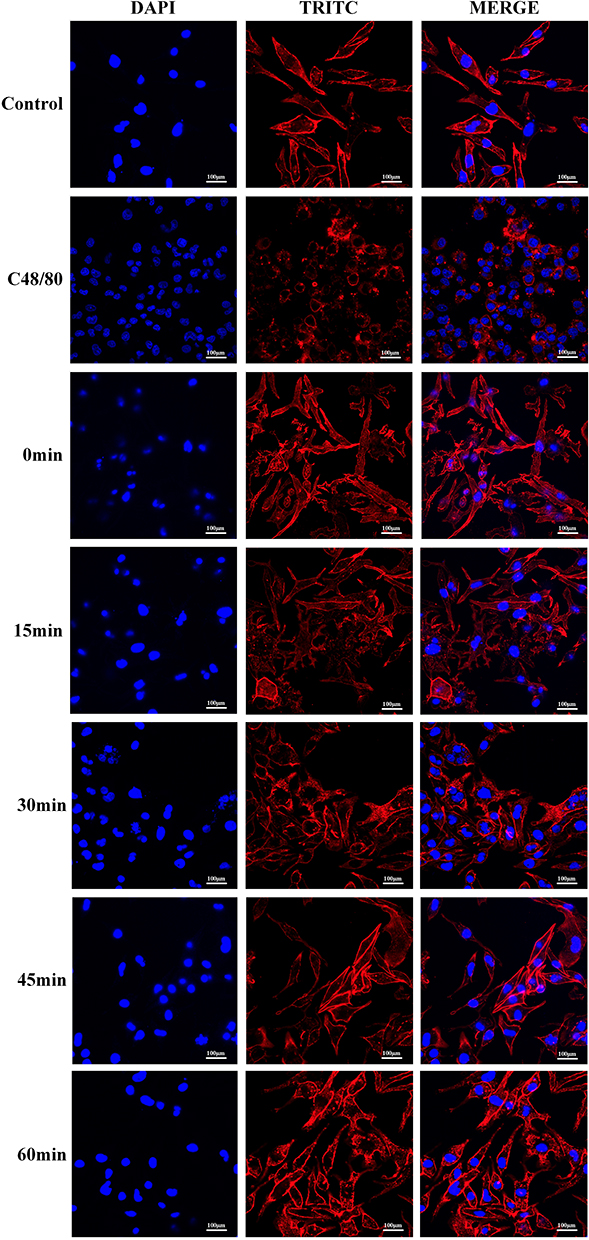 |
Figure 4 Effects of EgCF on the cytoskeleton. Abbreviations: EgCF, Echinococcus granulosus cyst fluid; TRITC, Tetramethylrhodamine isothiocyanate (The effect is similar to actin). Note: C48/80 50ug/mL and EgCF 3mg/mL; intervened cells for 0,15,30,45 and 60 min, respectively. |
Effects of EgCF on Apoptosis
Apoptosis rates increased significantly in a dose-dependent manner with rising concentrations of EgCF. All experimental groups demonstrated significantly elevated apoptosis compared to the control group (p < 0.001) (Figure 5). Similarly, both positive control groups showed a substantial increase in apoptotic cells relative to the control.
 |
Figure 5 Effects of EgCF on Cell Apoptosis. Abbreviation: EgCF, Echinococcus granulosus cyst fluid. Note: EgCF concentration (mg/mL); *** Delegates compared with the control group, P < 0.001. Group (A) is the control group, Group (B) EgCF group 0.5mg/mL, Group (C) EgCF group 1mg/mL, Group (D) EgCF group 2mg/mL, Group (E) EgCF 3mg/mL, Group (F) EgCF 4mg/mL, Group (G) EgCF group 5mg/mL; The total apoptosis rate was Q2 + Q4; C48 / 80 and EgCF intervened cells for 1h. |
Discussion
Leakage of EgCF following cyst rupture is known to provoke severe hypersensitivity reactions in the host. In vivo, rupture results in the release of EgCF into the pericystic space—a region already enriched with parasitic antigens due to selective molecular filtration by the laminated layer.15,16 The EgCF concentrations used in this study (0.5–5 mg/mL) were selected based on biochemical analyses of pericystic fluid in infected sheep livers,17 supporting the physiological relevance of the in vitro exposure model (Figures 3–5). These parasite-induced hypersensitivity responses are distinctive, and if improperly managed, may progress to life-threatening anaphylaxis.18
Previous reports have attributed echinococcosis-associated anaphylactic shock primarily to type I hypersensitivity reactions, potentially compounded by endotoxin-mediated shock, with the former considered the predominant mechanism.19 This complex immunopathological process involves multiple immune cell types,20 with mast cells playing a central effector role. Originating from hematopoietic stem cells in the bone marrow, mast cells undergo terminal maturation in peripheral tissues, where they act as key mediators of allergic inflammation.21,22
In the context of hydatid disease, an in vitro model that mimics mast cell activation in response to EgCF exposure is essential for mechanistic insights. The RBL-2H3 cell line, derived from rat basophilic leukemia cells, is widely used due to its phenotypic and functional similarity to native mast cells, including its ability to express FcεRI and undergo IgE-mediated degranulation.23,24
The present study employed this cell model to simulate allergic activation upon EgCF stimulation. Morphological observations revealed significant EgCF-induced cellular remodeling, including pseudopodia formation and membrane wrinkling (Figure 4), indicative of cytoskeletal reorganization, a hallmark of mast cell activation.25 Notably, this EgCF-induced cytoskeletal alteration in mast cells has not been previously reported in hydatid disease models. Phalloidin and toluidine blue staining provided clear visualization of dynamic cytoskeletal responses and degranulation events, underscoring the model’s utility in visualizing allergen-induced mast cell activation.
Additionally, the release of β-HEX, a granule-associated lysosomal enzyme functionally analogous to histamine and tryptase,26,27 further confirmed EgCF-induced degranulation. The observed dose-dependent increase in β-HEX release supports the hypothesis that EgCF plays a direct role in promoting mast cell activation and mediator release. Collectively, these findings offer a cellular and molecular basis for the severe allergic manifestations seen in individuals with cystic echinococcosis.
Unlike DNP-BSA, which selectively activates mast cells via the FcεRI receptor, EgCF comprises pathogen-associated molecular patterns, including glycans and parasitic DNA, which may additionally engage Toll-like receptors.28 This potential dual activation mechanism could underlie the enhanced degranulation observed and the rapid cytoskeletal remodeling noted in EgCF-treated cells. Supporting this hypothesis, prior studies have demonstrated that nematode-derived galectins can modulate crosstalk between FcεRI and TLR pathways,29 although EgCF-specific signaling mechanisms remain poorly characterized.30
In echinococcosis, EgCF functions as a key trigger of allergic responses following cyst rupture. To mimic this sensitization process in vitro, RBL-2H3 cells were sensitized using a monoclonal IgE antibody, and Compound 48/80 was employed as a positive control for inducing degranulation.31 The release of β-HEX, a recognized marker of mast cell activation and degranulation, was used to optimize the experimental parameters. Based on preliminary experiments, literature guidance, and laboratory feasibility, an IgE concentration of 0.125 µg/mL and a 12-hour sensitization period were selected.
To establish non-cytotoxic yet effective concentrations of EgCF, the CCK-8 assay was used to screen for viability-preserving doses. Final concentrations of 0.5, 1, 2, 3, 4, and 5 mg/mL were identified as suitable for degranulation assays. Following one hour of EgCF exposure, β-HEX release was significantly elevated across all concentrations, confirming their functional potency.
Toluidine blue staining further substantiated the induction of degranulation. In the control group, cells exhibited typical spindle-shaped morphology with intact and continuous membranes. EgCF-treated cells appeared more rounded or polygonal, and under high magnification, light blue granules indicative of degranulation were observed in the pericellular space. The degree of morphological deformation increased proportionally with EgCF concentration, and high concentrations ultimately resulted in partial membrane rupture. Based on β-HEX release profiles, 3 mg/mL was selected as the optimal working concentration for subsequent experiments. The positive control concentration of Compound 48/80 (50 µg/mL) was adopted from established protocols.32
Upon confirming successful model establishment, cytoskeletal dynamics during degranulation were evaluated using phalloidin staining. Mast cell activation is known to be accompanied by significant morphological changes, including cytoskeletal remodeling and mediator release. In this study, following IgE sensitization and EgCF stimulation, immunofluorescence imaging revealed increased cell volume and surface irregularity. Within 15 minutes of EgCF exposure, cells exhibited prominent wrinkling and lamellipodia extension. These structural changes peaked at 30 minutes and subsequently diminished, with most cells returning to baseline morphology by 60 minutes. This transient cytoskeletal rearrangement is consistent with classical IgE-mediated mast cell activation, but contrasts with prolonged deformation seen in endotoxin-induced or sepsis-related models.33
Notably, the reversible nature of EgCF-induced morphological changes (Figure 4) distinguishes it from irreversible membrane damage typically observed in complement-mediated cytotoxicity.34 This implies a distinct pathophysiological mechanism specific to parasitic anaphylaxis. Furthermore, findings by Zhou et al indicated that formononetin suppresses cytoskeletal alterations via FcεRI degradation,27 while the present study suggests that EgCF may actively leverage FcεRI pathways to promote cytoskeletal deformation and degranulation.
Collectively, these data confirm that EgCF induces robust degranulation in IgE-sensitized RBL-2H3 cells and support the development of a reliable in vitro model for studying E. granulosus-mediated allergic responses.
Further analysis integrating toluidine blue and phalloidin staining demonstrated that increasing concentrations of EgCF induced progressively more pronounced cellular deformation, with membrane rupture correlating with elevated β-HEX release. To determine whether this degranulation was associated with apoptosis, flow cytometric analysis was performed. The results revealed a concentration-dependent increase in apoptosis, particularly at EgCF concentrations of 4–5 mg/mL. These findings suggest that high EgCF concentrations not only induce mast cell degranulation but also promote apoptotic cell death, potentially intensifying the allergic response. This dual effect—simultaneous degranulation and apoptosis—distinguishes EgCF from other mast cell stimuli such as plant-derived allergens35 or synthetic drug candidates,36 which typically induce degranulation in the absence of cytotoxicity.
Notably, while Wang et al reported IL-10-mediated immunosuppression during chronic E. granulosus infection,37 the present findings uncover an acute pro-apoptotic effect of EgCF, indicative of a biphasic immune strategy: long-term immunological tolerance during chronic infection versus acute cytotoxicity upon cyst rupture. This model is consistent with clinical observations of anaphylactic shock in patients with cystic echinococcosis,38 and this study provides the first in vitro evidence linking mast cell apoptosis to the severity of hypersensitivity responses.
The observed coupling of degranulation and apoptosis may reflect a parasitic immune evasion strategy, wherein the rapid elimination of mast cells following degranulation impairs sustained host defense, contributing to the severity and lethality of anaphylactic reactions.
Despite the insights offered by this study, several mechanistic limitations should be acknowledged. First, although β-HEX is a well-established surrogate marker of degranulation,39 direct quantification of histamine—the principal mediator of acute anaphylaxis—was not performed. Future studies should assess histamine release to establish a direct link between enzymatic activity and the physiological mediators of hypotension and shock commonly observed during cyst rupture.40 Second, intracellular calcium flux, a critical signaling event in FcεRI-mediated mast cell activation,41 was not evaluated. Third, this study did not examine phosphorylation events in proximal signaling cascades, such as the Lyn–Syk–Gab2 axis. Given previous reports that nematode galectins can modulate Syk phosphorylation in basophils,29 it is plausible that components within EgCF similarly influence this pathway, contributing to the observed cytoskeletal dynamics (Figure 4).
Clarifying these molecular mechanisms will be critical for developing targeted interventions aimed at preventing or mitigating echinococcosis-related anaphylaxis.
This study demonstrates that EgCF induces dose-dependent mast cell degranulation and apoptosis, with higher concentrations (≥4 mg/mL) eliciting a cytotoxic response consistent with an immune evasion mechanism (Figures 3–5). These findings underscore three mechanistically distinct effects of EgCF: (1) degranulation surpassing that of synthetic agonists, (2) transient but pronounced cytoskeletal remodeling, and (3) apoptosis-mediated cytotoxicity. Collectively, these features delineate parasitic anaphylaxis as immunologically distinct from conventional allergic models.42
While previous research by Maruszewska-Cheruiyot et al reported that nematode-derived galectin inhibits mast cell apoptosis,29 the current findings reveal that E. granulosus components promote apoptotic cell death, suggesting a phylum-specific divergence in immune modulation strategies. This contrast highlights the necessity for further investigation into the molecular pathways, such as MAPK and NF-κB signaling, which may mediate these effects.
In conclusion, this in vitro model provides novel insights into the pathophysiological mechanisms underlying echinococcal allergic responses and may serve as a valuable platform for future studies aimed at developing targeted interventions for parasitic anaphylaxis.
Abbreviation
EgCF, Echinococcus granulosus cyst fluid; CCK-8, Cell Counting Kit-8; CE, Cystic echinococcosis; FBS, fetal bovine serum; IgE, Immunoglobulin E; C48/80, Compound 48/80; BCA, Bicinchoninic Acid Assay; DMEM, Dulbecco’s Modified Eagle Medium; PBS, Phosphate Buffered Saline; OD, Optical Density; β-HEX, β-Hexosaminidase; FBS, Fetal Bovine Serum; DNP-IgE, Anti-Dinitrophenol-Immunoglobulin E; DAPI, 4′,6-diamidino-2-phenylindole; TRITC, Tetramethylrhodamine isothiocyanate; Fc ε RI, high affinity IgE receptor; DNP-BSA, Anti-Dinitrophenol Bovine Serum Albumin.
Data Sharing Statement
All data generated or analysed during this study are included in this article. Further enquiries can be directed to the corresponding author.
Ethical Approval
All experiments were evaluated and approved by the ethics Committee of The First Affiliated Hospital of Xinjiang Medical University (IACUC-20200318-04) and complied with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Acknowledgments
We would like to acknowledge the hard and dedicated work of all the staff that implemented the intervention and evaluation components of the study.
Funding
National Natural Science Foundation of China (No.82060581).
Disclosure
The authors declare that they have no competing interests.
References
1. Woolsey ID, Miller AL. Echinococcus granulosus sensu lato and Echinococcus multilocularis: a review. Res Vet Sci. 2021;135:517–522. doi:10.1016/j.rvsc.2020.11.010
2. Zhang W, Li J, McManus DP. Concepts in immunology and diagnosis of hydatid disease. Clin Microbiol Rev. 2003;16(1):18–36. doi:10.1128/CMR.16.1.18-36.2003
3. Liu GH, Yang JY, Tang MJ. Research progress on the pathogenesis of human hepatic echinococcosis. Chin J Zoonoses. 2021;37(2):165–170. [in Chinese].
4. Feng SP, Guo YM, Han XM. The status and progress of minimally invasive surgical treatment on hepatic alveolar Echinococcosis. Parasitoses Infect Dis. 2021;19(01):51–56. [in Chinese].
5. Ye J, Zhang Q, Xuan Y, et al. Factors associated with Echinococcosis-induced perioperative anaphylactic shock. Korean J Parasitol. 2016;54(6):769–775. doi:10.3347/kjp.2016.54.6.769
6. Mendoza RP, Fudge DH, Brown JM. Cellular energetics of mast cell development and activation. Cells. 2021;10(3):524. doi:10.3390/cells10030524
7. da Silva EZ, Jamur MC, Oliver C. Mast cell function: a new vision of an old cell. J Histochem Cytochem. 2014;62(10):698–738. doi:10.1369/0022155414545334
8. Zeng H, Zhang H, Liu Z, et al. Dynactin subunit 1 facilitates mast cell degranulation to drive food allergy pathogenesis. Immunol Lett. 2025;20:107035.
9. Ghrairi N, Elhechmi YZ. Physiopathology of allergic asthma: a comprehensive review. Scand J Immunol. 2025;101(5):e70032. doi:10.1111/sji.70032
10. Huang R, Zhang W, Hu Y, et al. Houttuynia cordata polysaccharides ameliorate atopic dermatitis in mice through modulation of skin immune barrier and lipid metabolism. Int J Biol Macromol. 2025;314:144264. doi:10.1016/j.ijbiomac.2025.144264
11. Xia Y, Bin P, Zhou Y, et al. Glycerophospholipid metabolism licenses IgE-mediated mast cell degranulation. Cell Rep. 2025;44(6):115742. doi:10.1016/j.celrep.2025.115742
12. Guo X, Lei Y, Xu Y, et al. PRL2 negatively regulates FcεRI mediated activation of mast cells. Cell Death Dis. 2025;16(1):322. doi:10.1038/s41419-025-07649-2
13. Aitella E, Romano C, Ginaldi L, Cozzolino D. Mast cells at the crossroads of hypersensitivity reactions and neurogenic inflammation. Int J Mol Sci. 2025;26(3):927. doi:10.3390/ijms26030927
14. Liu ZC, Liu SF, Li F, et al. Study on degranulation model of RBL-2H3 cells. J Hebei Univ. 2018;38(02):159–167.
15. Yu X, Yasen Y, Wang C, et al. Construction of ceRNA regulatory network in mice with Echinococcosis-induced allergic reactions. Acta Trop. 2021;224:106120. doi:10.1016/j.actatropica.2021.106120
16. Ye J, Zhang Q, Ma L, Zheng H. Immunological characteristics of recurrent echinococcosis-induced anaphylactic shock. Am J Trop Med Hyg. 2016;94(2):371–377. doi:10.4269/ajtmh.15-0386
17. Mohammed AA, Allen JT, Rogan MT. Echinococcus granulosus cyst fluid enhances epithelial-mesenchymal transition. Parasite Immunol. 2018;40(6):e12533. doi:10.1111/pim.12533
18. Wang H, Li J, Pu H, et al. Echinococcus granulosus infection reduces airway inflammation of mice likely through enhancing IL-10 and down-regulation of IL-5 and IL-17A. Parasit Vectors. 2014;7:522. doi:10.1186/s13071-014-0522-6
19. Li Y, Zheng H, Cao X, et al. Demographic and clinical characteristics of patients with anaphylactic shock after surgery for cystic echinococcosis. American JTrop Med Hyg. 2011;85(3):452–455. doi:10.4269/ajtmh.2011.10-0448
20. Mendoza RP, Anderson CC, Fudge DH, et al. Metabolic consequences of IgE- and non-IgE-mediated mast cell degranulation. J Immunol. 2021;207(11):2637–2648. doi:10.4049/jimmunol.2001278
21. Arock M, Wedeh G, Hoermann G, et al. Preclinical human models and emerging therapeutics for advanced systemic mastocytosis. Haematologica. 2018;103(11):1760–1771. doi:10.3324/haematol.2018.195867
22. Smeding C, Gamadia LE, Heidt J. Allergie en anafylaxie [Allergy and anaphylaxis]. Ned Tijdschr Geneeskd. 2024;168:D8119. Dutch.
23. Ma D, Qin SX, Hao HC, et al. Screening and identification of in vitro anti-allergic active probiotics based on the degranulation inhibition of RBL-2H3 cells. Food Sci. 2024;45(06):55–63.
24. Barido FH, Lee SK. Effect of detoxified Rhus verniciflua extract on oxidative stability and quality improvement of raw chicken breast during cold storage. J Anim Sci Technol. 2022;64(2):380–395. doi:10.5187/jast.2022.e20
25. Bacci S. The evolution of mast cells across all vertebrate classes: the mystery continues. Histol Histopathol. 2025;18926. doi:10.14670/HH-18-926
26. Yang C, Zhang JW, Qi J, Yu BY. A novel method for evaluating pseudoallergy based on β-hexosaminidase and its application for traditional Chinese medicine injections. J Sep Sci. 2024;47(1):e2300545. doi:10.1002/jssc.202300545
27. Zhou ZW, Ji K, Zhu XY, et al. Natural isoflavone formononetin inhibits IgE-mediated mast cell activation and allergic inflammation by increasing IgE receptor degradation. Food Funct. 2023;14(6):2857–2869. doi:10.1039/D2FO03997D
28. Zhang Y, Yue Y, Cheng Y, Jiao H, Yan M. Antigen B from Echinococcus granulosus regulates macrophage phagocytosis by controlling TLR4 endocytosis in immune thrombocytopenia. Chem Biol Interact. 2025;406:111350. doi:10.1016/j.cbi.2024.111350
29. Maruszewska-Cheruiyot M, Szewczak L, Krawczak-Wójcik K, Stear MJ, Donskow-łysoniewska K. Nematode galectin inhibits Basophilic leukaemia RBL-2H3 cells apoptosis in IgE-mediated activation. Int J Mol Sci. 2024;25(13):7419. doi:10.3390/ijms25137419
30. Wang Z, Pu N, Zhao W, et al. RNA sequencing reveals dynamic expression of genes related to innate immune responses in canine small intestinal epithelial cells induced by Echinococcus granulosus protoscoleces. Front Vet Sci. 2024;11:1503995. doi:10.3389/fvets.2024.1503995
31. Wang J, Liu J, Yang Y, et al. Inhibitory effect of phellodendrine on C48/80-induced allergic reaction in vitro and in vivo. Int Immunopharmacol. 2024;134:112256. doi:10.1016/j.intimp.2024.112256
32. Kaushik A, Singh DP, Sahu M, et al. Protective effect of Achyranthes aspera against compound 48/80, histamine and ovalbumin-induced allergic disorders in murine model. Mol Biol Rep. 2024;51(1):202. doi:10.1007/s11033-023-09137-2
33. Huang J, Dai M, Huang X, et al. Shikonin inhibits NLRP3 inflammasome activation and controls inflammatory disease. Sci Rep. 2025;15(1):19037. doi:10.1038/s41598-025-03512-8
34. Zhang Z, Yin M, Geng X, et al. Ficolin-3 activates complement and triggers necroptosis in cholangiocarcinoma cells via the RIPK1/RIPK3/MLKL Signaling Pathway. FASEB J. 2025;39(10):e70673. doi:10.1096/fj.202403313R
35. Smith SA, Khan YW, Shrem RA, et al. Antigenic determinants underlying IgE-mediated anaphylaxis to peanut. J Allergy Clin Immunol. 2025;155(5):1595–1606.e10. doi:10.1016/j.jaci.2024.12.1094
36. Chen YC, Lee YR, Chang YC, et al. Scutellaria barbata ameliorates acute respiratory distress syndrome by inhibiting neutrophil-mediated inflammatory responses. J Ethnopharmacol. 2025;346:119653. doi:10.1016/j.jep.2025.119653
37. Wang X, Wang X, Wang D, et al. Interleukin-10 overexpression in 4T1 cells: a gateway to suppressing mammary carcinoma growth. Int Immunopharmacol. 2024;142(Pt A):113089. doi:10.1016/j.intimp.2024.113089
38. Zakariaei Z, Fakhar M, Sharifpour A, Banimostafavi ES, Soleymani M, Zakariaei A. Anaphylactic shock due to ruptured pulmonary hydatid cyst in a young patient from Iran. Ann Med Surg Lond. 2021;68:102675. doi:10.1016/j.amsu.2021.102675
39. Zhou YM, Jiao YX, Fan JK, et al. Ubiquitin-specific protease 13 regulates FcεRI-mediated mast cell activation and allergic inflammation via SYK protein modulation. Int J Biol Macromol. 2025;308(Pt 2):142302. doi:10.1016/j.ijbiomac.2025.142302
40. Shrestha S, Ghimire SK, Shrestha M, Maharjan S, Bishwakarma K. “Intraperitoneal ruptured hydatid cyst of liver with cystobiliary communication: a case report”. Int J Surg Case Rep. 2025;127:111012. doi:10.1016/j.ijscr.2025.111012
41. Kammala AK, Syed M, Yang C, Occhiuto CJ, Subramanian H. A critical role for Na+ /H+ exchanger regulatory factor 1 in modulating FcεRI-mediated mast cell activation. J Immunol. 2021;206(3):471–480. doi:10.4049/jimmunol.2000671
42. Tang RY, Yin L, Yao L, Zhang QF, Chen XP. A co-expressed natural antisense RNA FCER1A-AS controls IgE-dependent immunity by promoting expression of FcεRIα. Microbiol Spectr. 2023;11(3):e0073323. doi:10.1128/spectrum.00733-23
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.