")
Back to Archived Journals » Vaccine: Development and Therapy » Volume 4
Immunization and chemical conjugation of Bm95 obtained from Pichia pastoris enhances the immune response against vaccinal protein and Neisseria meningitidis capsular polysaccharide
Authors Rodriguez-Valle M , Canan-Hadden L, Niebla O
Received 4 November 2013
Accepted for publication 16 January 2014
Published 7 March 2014 Volume 2014:4 Pages 15—22
DOI https://doi.org/10.2147/VDT.S57043
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Manuel Rodriguez-Valle,1 Leonardo Canan-Hadden,2 Olivia Niebla2
1Animal Biotechnology Division, 2Analytical Division, Centre for Genetic Engineering and Biotechnology, Havana, Cuba
Abstract: The ectoparasite Rhipicephalus (Boophilus) microplus causes severe economic losses to the cattle industry in tropical and subtropical regions, and transmits endoparasites, such as Babesia bovis. The glycoprotein Bm95 is homologous to Bm86, a surface membrane protein of gut epithelial cells in R. microplus, and has been shown to efficiently control this ectoparasite in regions of the Americas. The immunostimulant properties of Bm86 have already been demonstrated after its coinjection with hepatitis B surface antigen (HBsAg) and the infectious bovine rhinotracheitis virus. This study evaluated the carrier and immunostimulant properties of Bm95 using low immunogenic Neisseria meningitidis capsular C polysaccharide (Men CpS) and HBsAg. We produced two polysaccharide-Bm95 conjugates by carbodiimide (MenCpSBm-c) and reductive amination (MenCpSBm-ra) methods. These conjugates were characterized and evaluated in mice. Antibody titers against Men CpS were significantly higher in mice immunized with MenCpSBm-ra (2,350±250, P<0.01) than in those immunized with MenCpSBm-c (250±75) or Men CpS (570±104). The study data indicate effective immunological memory after booster inoculation in mice immunized with MenCpSBm-ra. Additionally, significant humoral immunity against HBsAg was documented in mice coimmunized via the intranasal route with recombinant Bm95 (11,400±345) and HBsAg (128,000±250) compared with mice immunized only with HBsAg (400±40) or Bm95 (5,461±150, P<0.01). In conclusion, the immunostimulatory properties of recombinant Bm95 make it a useful element for developing safer conjugated vaccines against bacterial pathogens and for evaluation against ticks and tick-borne diseases in the context of a polyvalent veterinary vaccine.
Keywords: glycoconjugate, Bm86, capsular polysaccharides, carrier protein
Introduction
The Bm86 glycoprotein is localized on the membrane of gut epithelial cells in Rhipicephalus (Boophilus) microplus, and efficient control of this ectoparasite is possible in cattle vaccinated with this antigen.1 The blood of actively immunized bovine contained Bm86 antibodies that inhibited the biological function of the Bm86 on the surface of the gut epithelial cells.2,3 Additionally, immunological elements such as complement proteins work together with anti-Bm86 antibodies to lyse the intestinal epithelial cells of the tick.4,5 Due to these biological alterations, a reduction in tick number, weight, and reproductive capacity has been documented in R. microplus.6–8 The Bm86 antigen is expressed in the methylotrophic yeast Pichia pastoris, and the recombinant protein is obtained as a highly glycosylated and particulate antigen.3,9 These characteristics of recombinant Bm86 confer strong immunogenic and protective properties in many populations.10,11 Experimental reports confirm the adjuvant and immune-stimulating properties of Bm86.11,12 Bm95 is the homologous gene of Bm86, with only 2.1% of divergence between them, corresponding to approximately 3.4% of divergence between their respective amino acid sequences.13,14 The coding sequence of Bm95 was cloned and expressed in P. pastoris yeast; the recombinant protein was glycosylated, and the particulate antigen was similar to Bm86 and had similar immunogenic properties.14 In this report, the features of Bm95 were harnessed to explore and design conjugated vaccines.
Numerous bacteria responsible for invasive diseases have a polysaccharide capsule enabling these organisms to survive through the transmission and colonization processes. This polysaccharide capsule enables persistence of bacteria in the blood during the pathogenesis of an invasive infectious disease.15,16 The principal defense mechanism of the body against entry of these microorganisms is generation of an immune response against capsular polysaccharides. These antigens are thymus-independent and require the late development of a subset of B-cells.16 The immune response generated is characterized by a brief period of latency followed by a rapid rise of antibodies in serum, predominantly the immunoglobulin (Ig)M subclass. This immune response is associated with the immunological unresponsiveness observed in infants and some animals, and with failure of stimulation of the immunological response at any age.
Conjugation of a bacterial polysaccharide with an immunogenic carrier protein could promote a protective anti-polysaccharide IgG and induction of immunological memory.17–19 The effect of the carrier protein during the primary immunization consists of producing a change in IgG class and a population of B-cells for memory that can be stimulated by T-cell-independent type 2 or TD5 antigens.16,20–23 The polysaccharide component of a conjugated vaccine can be obtained from important veterinary and human pathogens, including Pasteurella hemolytica, Pasteurella multocida, Neisseria meningitidis, Clostridium spp., and Haemophilus spp., that usually are not strong immunogens. Additionally, the most used carrier proteins in conjugated vaccines are the tetanus toxoid and diphtheria antigens. Consequently, there are a limited number of carrier proteins for humans and animals, which constitutes a risk for immunogenicity of the individual conjugates administered in multivalent preparations.24,25 Previous studies have shown a reduced anti-polysaccharide antibody response attributable to the excess of carrier protein or carrier-mediated epitope suppression.26–31 Clinical studies of coadministration of conjugate vaccines using the same carrier protein have yielded inconsistent data related to immune interference and vaccine efficacy.32 The present study aims to take advantage of the immunogenic and particulate properties of Bm95 to develop glycoconjugated vaccines. To achieve this, recombinant Bm95 expressed in the yeast P. pastoris was conjugated by two chemical methods with the N. meningitidis capsular C polysaccharide. This bacterial polysaccharide and the hepatitis B surface antigen (HBsAg) were used to evaluate the carrier and immunostimulant properties of recombinant Bm95 in mice.
Mucosal surfaces such as in the gastrointestinal and respiratory tracts are the principal sites of contact and entry of a vast number of human and animal pathogens, hence stimulation of protective immunity at the mucosal level is very important for developing new vaccines. Also, mucosal vaccines have the ability to stimulate humoral-mediated and cell-mediated immune responses at the mucosal and systemic levels.33 These elements, along with advances in needleless, noninvasive immunization are important and attractive features of mucosal vaccination. In the present study, we also investigated the immunostimulatory properties of recombinant Bm95 administered by the nasal route.
Materials and methods
Native meningococcal C polysaccharide was obtained from the serogroup C meningococcal strain (C11 American Type Culture Collection, Manassas, VA, USA). Recombinant Bm95 and HBsAg were both expressed in the yeast P. pastoris and homogeneously purified as described elsewhere.14,34 The VA-MENGOC-BC® vaccine against N. meningitidis was produced at the Finlay Institute in Havana, Cuba.35 The outer membrane protein of N. meningitidis was obtained as described by Perez et al.36
Conjugation of polysaccharide
Carbodiimide method
The N. meningitidis C polysaccharide (Men CpS) was modified according to the method described by Beuvery et al in 1983.37,38 The polysaccharide (5 mg/mL in 0.2 M NaCl, pH 5.2) was reacted with 0.5 M adipic acid dihydrazide in the presence of 10 mM 1-cyclohexyl-3-(2-morpholinoethyl)-carbodiimide metho-p-toluenesulfonate of 1-cyclohexyl-3-(2-morpholinoethyl)-carbodiimide (EDC). The reaction was maintained with 0.1 M HCl in the pH range of 5.2–5.5 for 4 hours at room temperature. The mixture was dialyzed extensively against a solution of 0.2 M NaCl at pH 7.2 and concentrated using an Amicon-10 filter (Merck Millipore, Billerica, MA, USA). The residual adipic acid was separated by Sepharose-4BCL (Sigma-Aldrich, St Louis, MO, USA) chromatography from the modified Men CpS, which was concentrated using an Amicon-10 filter.
The conjugation was performed by slow addition of 0.5 mL EDC (100 mg/mL) into 20 mL of 0.2 M NaCl solution containing 50 mg of modified Men CpS and 50 mg of Bm95 to a final concentration of 2.5 mg/mL. The pH was maintained at 5.5 by addition of 0.1 M HCl. The reaction was carried out for 1 hour at room temperature. The conjugate was centrifuged for 30 minutes at 50,000× g, and the unreacted protein and dihydrazide polysaccharide were removed by Sepharose-4BCL chromatography previously equilibrated with 0.2 M phosphate buffer (pH 7.2). The chromatography flow speed was 0.5 mL per minute, and the conjugate was sterilized by filtration for its conservation after separation. The conjugate was designated as MenCpSBm-c.
Reductive amination method
The Men CpS was oxidized by a modified method of reductive amination reported by Jennings and Lugowski in 1981.39 A total of 200 mg of oxidized Men CpS was fragmented in 5% sodium acetate (10 mL, pH 8.1), and 200 mg of sodium metaperiodate was added at room temperature in the dark for 10 minutes. Next, 2 mL of ethylene glycol was added to expend the excess sodium metaperiodate, and the reaction mixture was left at room temperature for a further 60 minutes. The oxidized Men CpS with an average molecular weight of 40,000 Da was purified using a 1.5×90 cm Bio-Gel A 0.5 column (Bio-Rad, Hercules, CA, USA) by direct application of the reaction mixture treated with ethylene glycol. The column was previously calibrated with dextran (40,000 Da) and equilibrated with 250 mM sodium chloride and 25 mM disodium phosphate (pH 7). The oxidized Men CpS was desalted using a Sephadex G-25 column (Amersham Biosciences, Buckinghamshire, UK) and lyophilized.
The conjugation was performed by dissolving 5 mg of oxidized Men CpS, 5 mg of Bm95, and 5 mg of sodium cyanoborohydride in 1 mL of 0.1 M sodium hydrogen carbonate and 0.1 M NaCl solution (pH 8.3). The reaction mixture was maintained for 24 hours at 37°C with slow agitation. This mixture was applied to a Bio-Gel A 0.5 column (1.6× 100 cm) previously equilibrated with 25 mM of sodium phosphate and 250 mM of sodium chloride (pH 7.0) at a flow speed of 0.5 mL per minute. The conjugate eluted in the void volume of the column. The final product was designated as MenCpSBm-ra.
Determination of amino groups
The amino content was determined using the 2,4,6-trinitrobenzene sulfonic acid method40 (Thermo Fisher Scientific, Waltham, MA, USA). The concentration of amino groups is expressed in μmol/mg of Men CpS. Alcohol dehydrogenase was used as the standard for the standard curve.
Conjugate characterization
The N-acetylneuraminic acid content was determined by the resorcinol method.41 The polysaccharide content was expressed in mg/mL. Neuraminic acid was used as the standard in the standard curves, and the bicinchoninic acid assay was used to determine the protein concentration (Thermo Scientific Pierce Inc., Rockford, IL, USA).
Animals and immunization
Experiment 1
Female Balb/c mice aged 6–8 weeks of similar weight were used in five experimental groups. The mice were selected at random to form five groups with a total of five animals per group, and were maintained under controlled lighting conditions at 22°C, with access to water and laboratory chow ad libitum, as previously described.42 The Bm95 conjugates and polysaccharides were mixed with incomplete Freund’s adjuvant (Sigma-Aldrich, St Louis, MO, USA). The mice in group A were immunized with conjugated MenCpSBm-ra. Group B mice were immunized with MenCpSBm-c, group C mice were immunized with 2 μg of Men CpS, group D mice were immunized with 2 μg of unconjugated Bm95, and group E mice were vaccinated with VA-MENGOC-BC® as a positive control. Each dose of glycoconjugate was normalized to contain only 2 μg of Bm95 in a total volume of 100 μL administered subcutaneously. The second dose was administered 21 days after the initial dose. Blood was taken from the mice via the retroorbital vein, and the serum was collected and stored at −20°C until testing for antibodies against Bm95 and Men CpS by enzyme-linked immunosorbent assay (ELISA). Blood was taken before the first vaccination (preimmune serum) and two additional blood extractions were performed 15 days after the first and last doses.
Experiment 2
To study the immunostimulant properties of Bm95, five groups of 10-week-old female Balb/c mice were immunized intranasally with the following: 10 μg of Bm95 (group 1), 5 μg of proteoliposome (group 2) or 2 μg of HBsAg (group 3), while group 4 was inoculated with 10 μg of Bm95 combined with 5 μg of the outer membrane protein of N. meningitidis and group 5 was inoculated with 2 μg of HBsAg combined with 10 μg of Bm95. All immunogens were dissolved in 50 μL of phosphate-buffered saline, and a final volume of 100 μL was administered to each animal. Mice immunized in groups 4 and 5 received their respective immunogens mixed into the same preparation. The mice were immunized on days 0, 14, and 28. Samples of blood were obtained 12 days after the final immunization on day 40. Blood was collected from the retroorbital vein, and the serum was isolated and stored at −20°C until testing for antibodies.
Immunoassays
The antibody response against Men CpS was measured by enzyme-linked immunosorbent assay using a procedure described elsewhere.42 Microtiter plates (Nunc Immunoplate I, Gibco Ltd., Paisley, UK) were activated using poly-L-lysine at a concentration of 2 μg/mL in carbonate/bicarbonate (0–0.5 M) coating buffer (pH 9.5). Three washes were done with Milli Q water (18.2 mΩ · cm; EMD Millipore, Bedford, MA, USA). The plates were coated with 1 μg/mL of Men CpS in phosphate-buffered saline at 4°C overnight. The plates were washed three times with T-PBS (0.05% [v/v] Tween-20 in phosphate-buffered saline) and blocked with 1% skim milk in T-PBS for 1 hour at room temperature. The specific antibodies against Men CpS were developed with goat anti-mouse IgG-horseradish peroxidase conjugate for 1 hour at room temperature. The plates were washed, and the chromogenic reaction was developed in substrate buffer (3% of H2O2, o-phenylenediamine [0.5 mg/mL] in 0.1 M citrate buffer, pH 5). The reaction was stopped after 15 minutes at 25°C with 50 μL of 2 M orthophosphoric acid per well. The optical density was read in a plate reader at 492 nm (Sensidentscan; Merck, Darmstadt, Germany). End-point titers were determined as two-fold optical density values of preimmune sera. An enzyme-linked immunosorbent assay for detecting the antibody response against Bm95 and HBsAg was conducted as described elsewhere.12,13,43
Statistical analysis
Log-transformed titer data were used. The Student’s t-test (P<0.01) was used to test for statistically significant differences in antibody levels between the experimental groups.
Results
Effect of conjugation method on immunogenicity of MenCpSBm
Synthesis of the glycoconjugates was done using the carbodiimide and reductive amination methods. The composition of both conjugates was determined by chemical assay. The results of these assays indicate that the composition of the conjugates obtained by amination (MenCpSBm-ra) and oxidation (MenCpSBm-c) methods have a molar ratio of Men CpS to protein of 2.019±0.137 and 0.878±0.044, respectively. Gel filtration chromatography (Bio-Gel A 0.5 column) was used to separate the protein conjugates from the polysaccharide and chemical residual. The crude conjugate (Men CpS-Bm95-EDC) contained a 280 nm absorbing material in the void volume fraction, indicating that the protein had been linked to the Men CpS (data not shown). The antibody titers against meningococcal C polysaccharide induced after the first dose of MenCpSBm-ra (group A), MenCpSBm-c (group B), Men CpS (group C) and VA-MENGOC-BC® were very similar between the experimental groups (550±175, 550±225, 700±203, and 250±100, respectively). However, after the second dose of the immunization scheme (day 36), groups B and C showed a plateau in the immune response, with titers of 250±75 and 570±104, respectively (Figure 1). The response in group A showed significantly higher IgG titers (P<0.01) of 2,350±250 compared with the control group E (1,100±100). The placebo group (group D) did not have IgG titers against Men CpS detectable by enzyme-linked immunosorbent assay. Consequently, we can conclude that MenCpSBm-ra induced a potent antibody response against Men CpS under our experimental conditions.
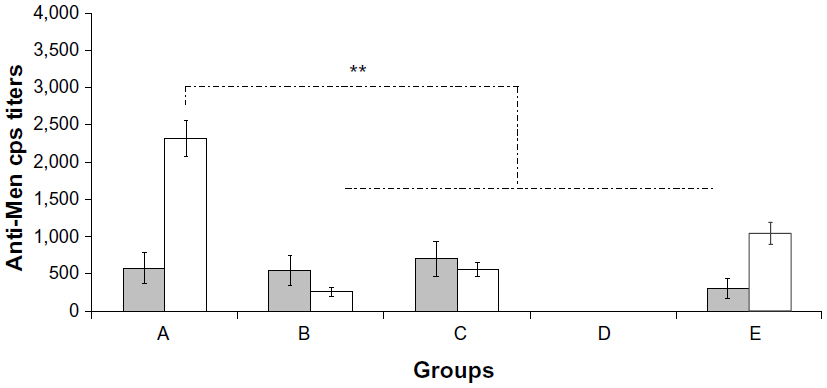 | Figure 1 Immunogenicity of MenCpSBm95 conjugates obtained against Men CpS. The gray bars represent immunoglobulin G titers 15 days after the first dose. The white bars represent immunoglobulin G titers 15 days after the second dose. The experimental groups were immunized with the MenCpSBm-ra (A) and MenCpSBm-c (B) conjugates, N. meningitidis C polysaccharide (C), Bm95 (D), and VA-MENGOC-BC® (E). **P<0.01, statistically significant difference by Student’s t-test. |
An enzyme-linked immunosorbent assay was used to measure the anti-carrier (Bm95) immune response, and showed only IgG titers against Bm95 in sera obtained from group A (MenCpSBm-ra), group B (MenCpSBm-c), and group D (Bm95) mice (Figure 2). The data obtained did not show interference by the anti-carrier IgG with the immune response against the meningococcal C polysaccharide fraction of the glycoconjugate.
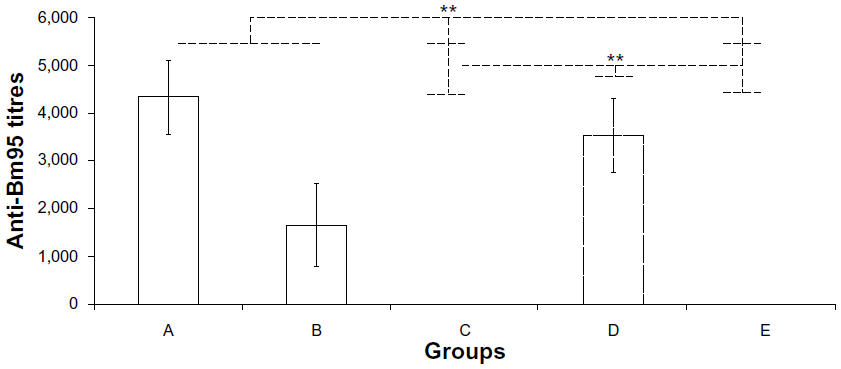 | Figure 2 Immunogenicity of the MenCpSBm95 conjugates against Bm95. An enzyme-linked immunosorbent assay was used to determine immunoglobulin G titers against Bm95 as described in the methods. The experimental groups were immunized with MenCpSBm-ra (A), MenCpSBm-c (B) conjugates, Men CpS (C), Bm95 (D), and VA-MENGOC-BC (E). **P< 0.01, statistically significant difference by Student’s t-test. |
Immunostimulant analysis by intranasal immunization
The immunostimulant capability of Bm95 was studied further by intranasal immunization using recombinant HBsAg. We observed that all animals immunized with Bm95 and HBsAg (group 5) seroconverted and reached mean titers of 11,400±345 and 128,000±250 against Bm95 and HBsAg, respectively. These titers were significantly (P<0.01) higher than the titers obtained in mice immunized only with Bm95 (5,461±150, group 1) or HBsAg (400±40, group 3). However, the anti-Bm95 titers (11,400±345) in group 5 were similar to the mean titers of group 4 immunized with Bm95 and the outer membrane protein of N. meningitidis (12,800±180, Figure 3).
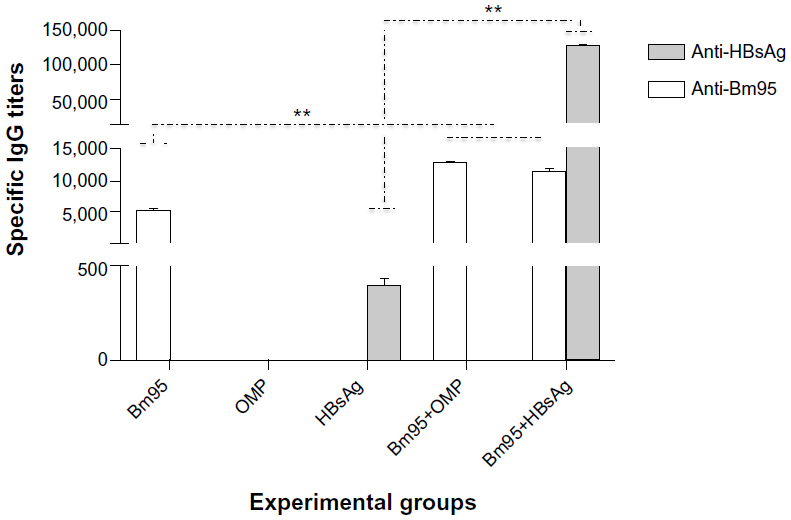 | Figure 3 Immunostimulant properties of Bm95 in mice immunized by the intranasal route. The data are shown as the mean ± standard deviation. **P<0.01, statistically significant difference by Student’s t-test. |
Discussion
The present study investigated the immunostimulatory properties of Bm95 used alone and as a carrier protein in a conjugated vaccinal preparation. Therefore, a rational approach for using Bm95 as a novel carrier molecule for preventing infections caused by polysaccharide encapsulated bacteria was conceived.42 The results obtained in this study showed high immunogenicity of the Bm95-based conjugate (MenCpSBm-ra) compared with the meningococcal C polysaccharide. Additionally, an enhanced immune response typical of the T-cell-dependent immune response against Men CpS was observed. The Bm95 conjugates induced a more rapid and stronger immune response after the first dose against Men CpS, as high titers against Bm95 were maintained. However, MenCpSBm-ra induced a significantly higher antibody titer against Men CpS than MenCpSBm-c after the second dose, and similar results have been reported previously using the same conjugation reaction.42 The effect of conjugation chemistry on the immunogenicity of polysaccharide-protein conjugates and the advantage of the reductive amination method to develop an effective immune response against polysaccharide antigens have already been reported.42,44–48 The ratio of Bm95 to Men CpS had an important influence on the results. The conjugate obtained by the reductive amination method had a high ratio of Men CpS to Bm95 compared with the carbodiimide conjugate. Consequently, this ratio of carrier protein was observed to provide enough T-cell epitopes for the development of the most efficient T-cell-dependent response and improved the presentation of carbohydrate epitopes. This explains the robust immune response observed after administration of the second doses with MenCpSBm-ra.
Intranasal immunization is a distinct approach that offers advantages in terms of protection against infection at both local and distant sites.49 The immune response after vaccination using the intranasal route is usually characterized by mucosal IgA, as well as systemic IgG. This response is mediated by type 1 and type 2 helper T cells lymphocytes and complements the protection against a number of pathogens affecting the upper and lower respiratory tracts of humans and animals.50–54 Further, immunization via the mucosal surface is an effective approach for avoiding maternal antibody interference53,55 and is a less stressful immunization procedure.
Consequently, the immunogenicity of Bm95 was further tested in this study by intranasal co-immunization with HBsAg. Enhancement of the humoral immune response against HBsAg following intranasal administration of both antigens was observed. However, the specific IgG titer obtained against Bm95 after coadministration of both antigens is similar to that of Bm95 mixed with outer membrane protein, which is an important immune potentiator.56–60 Previously, similar adjuvant properties were reported for Bm86 antigen obtained from P. pastoris, which is homologous with Bm95. A small number of proteins have adjuvant characteristics and act principally as B-cell mitogens capable of inducing high antibody levels. The particulate antigen obtained from P. pastoris could trigger important immunological signals to gather immunological factors that improve the immune response. Whereas the mechanism of interaction between Bm95 or Bm86 and the immune system is imprecise, there are important experimental data showing a better immune response to vaccinal proteins coadministered with these antigens.12,56,61 However, this is the first study that similarly shows the adjuvant and immunostimulant properties of Bm95 administered intranasally in mice.
Conclusion
The present study demonstrates for the first time, to the authors’ knowledge, that the conjugation procedure using recombinant Bm95 as a carrier protein improves antibody titers against bacterial polysaccharide antigens, which usually have poor immunogenicity. Hence, Bm95 seems to be an attractive candidate for use as a carrier protein or in combination with other vaccine antigens because of its availability in high purity, its yeast origin and its immunogenicity. This protein could be used with polysaccharide components obtained from different bacterial entities to develop multivalent vaccines conjugated against veterinary pathogens.
Acknowledgment
The authors are grateful to the staff from the Animal Biotechnology Division at Centre for Genetic Engineering and Biotechnology (Havana, Cuba) for their technical assistance.
Disclosure
The authors report no conflicts of interest in this work.
References
Willadsen P, Riding GA, McKenna RV, et al. Immunological control of a parasitic arthropod: identification of a protective antigen from Boophilus microplus. J Immunol. 1989;143:1346–1351. | |
Rand KN, Moore T, Sriskantha A, et al. Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proc Nat Acad Sci U S A. 1989;86:9657–9661. | |
Rodriguez Valle M, Rubiera R, Penichet M, et al. High-level expression of the B. microplus Bm86 antigen in the yeast Pichia pastoris forming highly immunogenic particles for cattle. J Biotechnol. 1994;33:135–146. | |
Willadsen P. Immunological control of ectoparasites: past achievements and future research priorities. Genet Anal. 1999;15:131–137. | |
Willadsen P. Tick control: thoughts on a research agenda. Vet Parasitol. 2006;138:161–168. | |
Rodriguez Valle M, Penichet mL, Mouris AE, et al. Control of Boophilus microplus populations in grazing cattle vaccinated with a recombinant Bm86 antigen preparation. Vet Parasitol. 1995;57:339–349. | |
de la Fuente J, Rodriguez Valle M, Garcia-Garcia JC. Immunological control of ticks through vaccination with Boophilus microplus gut antigens. Trop Vet Dis. 2000;916:617–621. | |
Rodriguez Valle M, Mendez L, Valdez M, et al. Integrated control of Boophilus microplus ticks in Cuba based on vaccination with the anti-tick vaccine Gavac(™). Exp Appl Acarol. 2004;34:375–382. | |
Canales M, Enriquez A, Ramos E, et al. Large-scale production in Pichia pastoris of the recombinant vaccine Gavac(™) against cattle tick. Vaccine. 1997;15:414–422. | |
Garcia-Garcia JC, Montero C, Rodriguez Valle M, et al. Effect of particulation on the immunogenic and protective properties of the recombinant Bm86 antigen expressed in Pichia pastoris. Vaccine. 1998;16:374–380. | |
Rodriguez Valle M, Montero C, Machado H, Joglar M. The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnol. 2001;1:2. | |
Garcia-Garcia JC, Soto A, Nigro F, et al. Adjuvant and immunostimulating properties of the recombinant Bm86 protein expressed in Pichia pastoris. Vaccine. 1998;16:1053–1055. | |
Garcia-Garcia JC, Gonzalez IL, Gonzalez DM, et al. Sequence variations in the Boophilus microplus Bm86 locus and implications for immunoprotection in cattle vaccinated with this antigen. Exp Appl Acarol. 1999;23:883–895. | |
Garcia-Garcia JC, Montero C, Redondo M, et al. Control of ticks resistant to immunization with Bm86 in cattle vaccinated with the recombinant antigen Bm95 isolated from the cattle tick, Boophilus microplus. Vaccine. 2000;18:2275–2287. | |
Deangelis PL, White CL. Identification of a distinct, cryptic heparosan synthase from Pasteurella multocida types A, D, and F. J Bacteriol. 2004;186:8529–8532. | |
Kelly DF, Moxon ER, Pollard AJ. Haemophilus influenzae type b conjugate vaccines. Immunology. 2004;113:163–174. | |
Robbins JB, Schneerson R. Polysaccharide-protein conjugates: a new generation of vaccines. J Infect Dis. 1990;161:821–832. | |
Granoff DM, Rathore MH, Holmes SJ, Granoff PD, Lucas AH. Effect of immunity to the carrier protein on antibody responses to Haemophilus influenzae type b conjugate vaccines. Vaccine. 1993;11 Suppl 1:S46–S51. | |
Granoff DM, Moe GR, Giuliani MM, et al. A novel mimetic antigen eliciting protective antibody to Neisseria meningitidis. J Immunol. 2001;167:6487–6496. | |
Donnelly JJ, Deck RR, Liu MA. Immunogenicity of a Haemophilus influenzae polysaccharide-Neisseria meningitidis outer membrane protein complex conjugate vaccine. J Immunol. 1990;145:3071–3079. | |
Brogden KA, DeBey B, Audibert F, Lehmkuhl H, Chedid L. Protection of ruminants by Pasteurella haemolytica A1 capsular polysaccharide vaccines containing muramyl dipeptide analogs. Vaccine. 1995;13:1677–1684. | |
Rubinstein LJ, Garcia-Ojeda PA, Michon F, Jennings HJ, Stein KE. Murine immune responses to Neisseria meningitidis group C capsular polysaccharide and a thymus-dependent toxoid conjugate vaccine. Infect Immun. 1998;66:5450–5456. | |
Vos Q, Lees A, Wu ZQ, Snapper CM, Mond JJ. B-cell activation by T-cell-independent type 2 antigens as an integral part of the humoral immune response to pathogenic microorganisms. Immunol Rev. 2000;176:154–170. | |
Insel RA. Potential alterations in immunogenicity by combining or simultaneously administering vaccine components. Ann N Y Acad Sci. 1995;754:35–47. | |
Bogaert D, Hermans PW, Adrian PV, Rumke HC, de Groot R. Pneumococcal vaccines: an update on current strategies. Vaccine. 2004;22:2209–2220. | |
Di John D, Wasserman SS, Torres JR, et al. Effect of priming with carrier on response to conjugate vaccine. Lancet. 1989;2:1415–1418. | |
Peeters CC, Tenbergen-Meekes AM, Poolman JT, Beurret M, Zegers BJ, Rijkers GT. Effect of carrier priming on immunogenicity of saccharide-protein conjugate vaccines. Infect Immun. 1991;59:3504–3510. | |
Bergquist C, Lagergard T, Holmgren J. Anticarrier immunity suppresses the antibody response to polysaccharide antigens after intranasal immunization with the polysaccharide-protein conjugate. Infect Immun. 1997;65:1579–1583. | |
Dagan R, Eskola J, Leclerc C, Leroy O. Reduced response to multiple vaccines sharing common protein epitopes that are administered simultaneously to infants. Infect Immun. 1998;66:2093–2098. | |
Renjifo X, Wolf S, Pastoret PP, et al. Carrier-induced, hapten-specific suppression: a problem of antigen presentation? J Immunol. 1998;161:702–706. | |
Fattom A, Cho YH, Chu C, Fuller S, Fries L, Naso R. Epitopic overload at the site of injection may result in suppression of the immune response to combined capsular polysaccharide conjugate vaccines. Vaccine. 1999;17:126–133. | |
Knuf M, Kowalzik F, Kieninger D. Comparative effects of carrier proteins on vaccine-induced immune response. Vaccine. 2011;29:4881–4890. | |
Neutra MR, Kozlowski PA. Mucosal vaccines: the promise and the challenge. Nat Rev Immunol. 2006;6:148–158. | |
Herrera L, Del la Fuente J, Quinones Y, et al. Biologicals production by recombinant DNA technology in Cuba. Dev Biol Stand. 1989;70:257–269. | |
Castellanos L, Betancourt L, Sardinas G, et al. [Characterization of N. meningitidis proteoliposome proteins. Consistency and reproducibility among batches of VA-MENGOC-BC, assessed by proteomic techniques]. Biotecnología Aplicada. 2007;24:168–171. Portuguese. | |
Perez O, Lastre M, Lapinet J, et al. Immune response induction and new effector mechanisms possibly involved in protection conferred by the Cuban anti-meningococcal BC vaccine. Infect Immun. 2001;69:4502–4508. | |
Beuvery EC, Miedema F, van Delft R, Haverkamp J. Preparation and immunochemical characterization of meningococcal group C polysaccharide-tetanus toxoid conjugates as a new generation of vaccines. Infect Immun. 1983;40:39–45. | |
Beuvery EC, van Delft RW, Miedema F, Kanhai V, Nagel J. Immunological evaluation of meningococcal group C polysaccharide-tetanus toxoid conjugate in mice. Infect Immun. 1983;41:609–617. | |
Jennings HJ, Lugowski C. Immunochemistry of groups A, B, and C meningococcal polysaccharide-tetanus toxoid conjugates. J Immunol. 1981;127:1011–1018. | |
Goodwin JF, Choi SY. Quantification of protein solutions with trinitrobenzenesulfonic acid. Clin Chem. 1970;16:24–31. | |
Svennerholm L. Quantitative estimation of sialic acids. II. A colorimetric resorcinol-hydrochloric acid method. Biochim Biophys Acta. 1957;24:604–611. | |
Carmenate T, Canaan L, Alvarez A, et al. Effect of conjugation methodology on the immunogenicity and protective efficacy of meningococcal group C polysaccharide-P64k protein conjugates. FEMS Immunol Med Microbiol. 2004;40:193–199. | |
Triguero A, Blanco R, Machado H, Rodriguez Valle M, de la Fuente J. Development of enzyme linked immunosorbent assays to measure Bm86 antigen of Boophilus microplus (cattle tick) and to detect anti-Bm86 antibodies in serum samples. Biotechnol Tech. 1999;13:119–125. | |
Fattom A, Li X, Cho YH, et al. Effect of conjugation methodology, carrier protein, and adjuvants on the immune response to Staphylococcus aureus capsular polysaccharides. Vaccine. 1995;13:1288–1293. | |
Gupta RK, Egan W, Bryla DA, Robbins JB, Szu SC. Comparative immunogenicity of conjugates composed of Escherichia coli O111 O-specific polysaccharide, prepared by treatment with acetic acid or hydrazine, bound to tetanus toxoid by two synthetic schemes. Infect Immun. 1995;63:2805–2810. | |
Schneerson R, Robbins JB, Chu C, et al. Serum antibody responses of juvenile and infant rhesus monkeys injected with Haemophilus influenzae type b and pneumococcus type 6A capsular polysaccharide-protein conjugates. Infect Immun. 1984;45:582–591. | |
Seppala I, Makela O. Antigenicity of dextran-protein conjugates in mice. Effect of molecular weight of the carbohydrate and comparison of two modes of coupling. J Immunol. 1989;143:1259–1264. | |
Shen X, Lagergard T, Yang Y, Lindblad M, Fredriksson M, Holmgren J. Group B Streptococcus capsular polysaccharide-cholera toxin B subunit conjugate vaccines prepared by different methods for intranasal immunization. Infect Immun. 2001;69:297–306. | |
Bergquist C, Johansson EL, Lagergard T, Holmgren J, Rudin A. Intranasal vaccination of humans with recombinant cholera toxin B subunit induces systemic and local antibody responses in the upper respiratory tract and the vagina. Infect Immun. 1997;65:2676–2684. | |
Kurono Y, Yamamoto M, Fujihashi K, et al. Nasal immunization induces Haemophilus influenzae-specific Th1 and Th2 responses with mucosal IgA and systemic IgG antibodies for protective immunity. J Infect Dis. 1999;180:122–132. | |
Mielcarek N, Riveau G, Remoue F, Antoine R, Capron A, Locht C. Homologous and heterologous protection after single intranasal administration of live attenuated recombinant Bordetella pertussis. Nat Biotechnol. 1998;16:454–457. | |
Jakobsen H, Saeland E, Gizurarson S, Schulz D, Jonsdottir I. Intranasal immunization with pneumococcal polysaccharide conjugate vaccines protects mice against invasive pneumococcal infections. Infect Immun. 1999;67:4128–4133. | |
Shewen PE, Carrasco-Medina L, McBey BA, Hodgins DC. Challenges in mucosal vaccination of cattle. Vet Immunol Immunopathol. 2009;128:192–198. | |
Todd JD. Intranasal vaccination of cattle against IBR and PI3: field and laboratory observations in dairy, beef, and neonatal calf populations. Dev Biol Stand. 1976;33:391–395. | |
Prado ME, Prado TM, Payton M, Confer AW. Maternally and naturally acquired antibodies to Mannheimia haemolytica and Pasteurella multocida in beef calves. Vet Immunol Immunopathol. 2006;111:301–307. | |
Wetzler LM. Immunopotentiating ability of neisserial major outer membrane proteins. Use as an adjuvant for poorly immunogenic substances and potential use in vaccines. Ann N Y Acad Sci. 1994;730:367–370. | |
Verheul AF, Snippe H, Poolman JT. Meningococcal lipopolysaccharides: virulence factor and potential vaccine component. Microbiol Rev. 1993;57:34–49. | |
Verheul AF, Van Gaans JA, Wiertz EJ, Snippe H, Verhoef J, Poolman JT. Meningococcal lipopolysaccharide (LPS)-derived oligosaccharide-protein conjugates evoke outer membrane protein- but not LPS-specific bactericidal antibodies in mice: influence of adjuvants. Infect Immun. 1993;61:187–196. | |
Mesa C, De Leon J, Rigley K, Fernandez LE. Very small size proteoliposomes derived from Neisseria meningitidis: an effective adjuvant for Th1 induction and dendritic cell activation. Vaccine. 2004;22:3045–3052. | |
Tamargo B, Marquez Y, Ramirez W, Cedre B, Fresno M, Sierra G. New proteoliposome vaccine formulation from N. meningitidis serogroup B, without aluminum hydroxide, retains its antimeningococcal protectogenic potential as well as Th-1 adjuvant capacity. BMC Immunol. 2013; 14 Suppl 1:S12. | |
Lycke N, Holmgren J. Strong adjuvant properties of cholera toxin on gut mucosal immune responses to orally presented antigens. Immunology. 1986;59:301–308. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.