Back to Journals » International Journal of Chronic Obstructive Pulmonary Disease » Volume 13
Immune cell response to strenuous resistive breathing: comparison with whole body exercise and the effects of antioxidants
Authors Asimakos A, Toumpanakis D, Karatza MH, Vasileiou S, Katsaounou P, Mastora Z, Vassilakopoulos T
Received 19 October 2017
Accepted for publication 16 December 2017
Published 7 February 2018 Volume 2018:13 Pages 529—545
DOI https://doi.org/10.2147/COPD.S154533
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Richard Russell
Andreas Asimakos,1,2,* Dimitrios Toumpanakis,1,2,* Maria-Helena Karatza,3 Spyridoula Vasileiou,3 Paraskevi Katsaounou,1,2 Zafeiria Mastora,1,2 Theodoros Vassilakopoulos1,2,4
1GP Livanos and M Simou Laboratories, Thorax Foundation, 2Critical Care Department and Pulmonary Unit, Evangelismos Hospital, Medical School, National and Kapodistrian University of Athens, 3Flow Cytometry Unit, Hematology Clinic Evangelismos Hospital, 43rd Department of Critical Care Medicine, Evgenideion Hospital, Medical School, National and Kapodistrian University of Athens, Athens, Greece
*These authors contributed equally to this work
Background/hypothesis: Whole body exercise (WBE) changes lymphocyte subset percentages in peripheral blood. Resistive breathing, a hallmark of diseases of airway obstruction, is a form of exercise for the inspiratory muscles. Strenuous muscle contractions induce oxidative stress that may mediate immune alterations following exercise. We hypothesized that inspiratory resistive breathing (IRB) alters peripheral blood lymphocyte subsets and that oxidative stress mediates lymphocyte subpopulation alterations following both WBE and IRB.
Patients and methods: Six healthy nonathletes performed two WBE and two IRB sessions for 45 minutes at 70% of VO2 maximum and 70% of maximum inspiratory pressure (Pimax), respectively, before and after the administration of antioxidants (vitamins E, A, and C for 75 days, allopurinol for 30 days, and N-acetylcysteine for 3 days). Blood was drawn at baseline, at the end of each session, and 2 hours into recovery. Lymphocyte subsets were determined by flow cytometry.
Results: Before antioxidant supplementation at both WBE end and IRB end, the natural killer cell percentage increased, the T helper cell (CD3+ CD4+) percentage was reduced, and the CD4/CD8 ratio was depressed, a response which was abolished by antioxidants only after IRB. Furthermore, at IRB end, antioxidants promoted CD8+ CD38+ and blunted cytotoxic T-cell percentage increase. CD8+ CD45RA+ cell percentage changes were blunted after antioxidant supplementation in both WBE and IRB.
Conclusion: We conclude that IRB produces (as WBE) changes in peripheral blood lymphocyte subsets and that oxidative stress is a major stimulus predominantly for IRB-induced lymphocyte subset alterations.
Keywords: resistive breathing, exercise, antioxidants, lymphocyte
Introduction
Peripheral blood leukocyte subpopulations respond rather stereotypically to whole body exercise (WBE). During WBE, neutrophil, lymphocyte, and monocyte counts increase, followed by a reduction in lymphocyte count after exercise due to redistribution and apoptosis.1 Regarding specific lymphocyte subpopulations, exercise is mainly associated with an increase in CD8+ T-lymphocyte and CD56+ CD16+ natural killer (NK) cell percentage and a subsequent decrease in CD4+ T-lymphocyte percentage.1 Alterations in lymphocyte count have been associated with suppressed immune function following intense exercise.2
Reactive oxygen species (ROS) generated during intense WBE are among the potential modulators of this response3 though their exact role has not been established. Highly intense exercise induces lymphocyte apoptosis via an ROS-dependent pathway.4 Oxidative stress may induce DNA damage of immunocompetent cells after prolonged and strenuous exercise (marathon run).5 Yet antioxidant supplementation (N-acetylcysteine [NAC]) showed no effect in exercise-induced proliferation and activity of lymphocyte subsets in trained athletes.6 As in WBE, free radicals in the form of ROS and reactive nitrogen species are generated during increased contractile activity of the inspiratory muscles, mainly the diaphragm.7,8
Resistive breathing is encountered in obstructive airway diseases, such as asthma and chronic obstructive pulmonary disease (COPD), especially during exacerbations.9 Inspiratory resistive breathing (IRB) is a form of exercise of the inspiratory muscles and is associated with intense respiratory muscle contractions. When strenuous enough, IRB produces diaphragmatic fatigue and diaphragmatic structural injury and enacts as an immune challenge initiating cytokine upregulation in the diaphragm10,11 and the plasma.12,13 IRB has also been shown to induce pulmonary inflammation and lung injury in experimental animal models.14,15
The immune system’s role and the influence of the excessive production of oxidative derivatives in obstructive pulmonary diseases are active research fields.16–18 In continuation of our previous experiments10–15 in humans and animals where resistive breathing, as a model of airway obstruction, produced oxidative stress-dependent systematic and pulmonary inflammation, we decided to investigate its effects on the cells that orchestrate various immune responses, the lymphocytes. We thus hypothesized that IRB, as WBE, induces lymphocyte subpopulation changes in the peripheral blood and that oxidative stress modulates this response. We also hypothesized that oxidative stress modulates the peripheral blood lymphocyte subpopulation responses to WBE. To test our hypotheses, we conducted WBE and IRB sessions of equal duration before and after in vivo supplementation of antioxidants in healthy nonathlete volunteers.
Subjects and methods
Subjects
Six healthy male volunteers, who were not involved in strenuous manual labor due to their profession, free of any history of asthma and other relevant respiratory conditions, with a mean age of 33 years (28–37 years) were studied. They did not participate in regular exercise training or sports activities and had not had febrile illness in the 3 months before or throughout the duration of the experiment. The subjects were instructed to refrain from intense physical activity or regular exercise training during the study period, to adopt their usual dietary pattern, and were also allowed to have normal daily activities. Once per week, each subject visited the laboratory, was supplied with doses of antioxidants for 7 days, and returned any unused ones. On the same day of the visit, each subject was asked for changes in everyday habits and for symptoms of illness. Each participant was receiving reminder calls every 2 days to ensure compliance with antioxidant supplementation. The Ethics Committee of our institution, Evangelismos Hospital, approved the study protocol, and all the subjects provided written informed consent.
Overall study design
The six subjects performed two sessions of IRB and WBE before and after the administration of antioxidants. Each participant performed an IRB session followed by a WBE session after 15 days, in order to eliminate the possible influences of one session on the other (ie, any muscle injury incurring during one session persisting and thus affecting the immune response of the next session),1 and then, antioxidant supplementation began. The same IRB session was performed after 60 days of antioxidant supplementation followed by the same WBE after 15 days totaling 75 days of antioxidant supplementation and 90 days of overall experimental duration. Therefore, each participant served as a self-control eliminating between subject variability. Experiments were performed at summer time minimizing the chance of (viral) respiratory tract infections, which in fact did not happen to any of our subjects.
After the 1st WBE session, participants started receiving a combination of antioxidants, including 200 mg vitamin E, 50,000 IU vitamin A, and 1,000 mg vitamin C per day, for a total of 75 days. Allopurinol, 600 mg/day, was administered for a total of 30 days starting at day 61 (14 days before the 2nd IRB session) and NAC, 1,800 mg/day, for 3 days before the 2nd IRB and the 2nd WBE sessions. Antioxidants were supplemented in one dose in the morning with breakfast except NAC which was administered in three doses (600 mg each). In the morning of testing, a full dose of antioxidants and 600 mg of NAC were supplemented 2 hours before the session. This design was chosen instead of a randomized crossover design because of the nature of the antioxidants: vitamins with long-lasting effects (especially vitamin E) after the completion of the supplementation.19 The duration and combination of antioxidants were based on our previous experiments with WBE19 and IRB.13
The flowchart of the study design is as follows:
- Day 1: 1st IRB session
- Day 15: 1st WBE session
- Day 16: Initiation of vitamin A (50,000 IU/day), vitamin E (200 mg/day), and vitamin C (1,000 mg/day) in one dose in the morning
- Day 61: Initiation of allopurinol (600 mg/day) one dose in the morning
- Day 72: Initiation of NAC (1,800 mg/day in three doses of 600 mg)
- Day 75: 2nd IRB session; supplementation in the morning 2 hours before session of vitamin A (50,000 IU), vitamin E (200 mg), vitamin C (1,000 mg), allopurinol (600 mg), and NAC (600 mg morning dose); termination of NAC supplementation
- Day 87: Initiation of NAC (1,800 mg/day in three doses of 600 mg)
- Day 90: 2nd WBE session; supplementation in the morning 2 hours before session of vitamin A (50,000 IU), vitamin E (200 mg), vitamin C (1,000 mg), allopurinol (600 mg), and NAC (600 mg morning dose); termination of all antioxidant supplementation. Figure 1 illustrates the flow chart of the study design.
 | Figure 1 Antioxidant supplementation time line. |
Preliminary testing
Preliminary testing was conducted 2 weeks before the initiation of the experimental sessions (1st IRB session).
WBE
The maximum WBE capacity of each subject was assessed at the initial visit to the laboratory. Participants got accustomed to an electrically braked cycle ergometer (Medical Graphics CPX/D; Medical Graphics Corporation, Saint Paul, MN, USA) and were instructed to maintain a constant pedal speed of 60 rpm. Increments of 10 watts every minute were applied until each subject reached a maximum work load (volitional exhaustion) in order to determine the maximum oxygen consumption rate (VO2 max) in accordance with our previous experiments. Heart rate and blood pressure were continuously monitored.19
IRB
The IRB sessions were also performed in accordance with previous experiments.9,13,20 Subjects were placed in the sitting position and were accustomed to the procedure. A mouthpiece was adjusted to a T-piece adapter, and two one-way valves (Hans Rudolph, Inc., Shawnee, KS, USA) were attached to each side in order to determine an inspiration and expiration port. A tube with an adjustable orifice was attached to the inspiratory port in order to increase inspiratory resistance, while the expiratory port was left without load. A rigid tube connected the mouthpiece with a pressure transducer (Validyne, Los Angeles, CA, USA), and the measurements were recorded on a 16-channel electrostatic recorder (Gould ES 1000; Gould Instruments, Cleveland, OH, USA) and displayed on an oscilloscope (Tektronix 2213; Beaverton, OR, USA). To prevent glottis closure during the run, a small hole, 2 mm in diameter, was opened in the mouthpiece. The maximum inspiratory pressure (Pimax) was measured while the subject performed maximal inspiratory effort from functional residual capacity against an occluded inspiratory port. The Pimax was considered the most negative mouth pressure that could be sustained for at least 1 second. Multiple efforts (up to 5 after introductory training on how to perform the maneuver) were performed, and the most negative pressure was considered each subject’s Pimax. Pimax maneuvers were separated by 2 minutes each to avoid potentiation. Preliminary resistive breathing sessions were performed for each subject in order to select the amount of resistance that was required as to achieve inspiratory pressures at 70% of the maximum inspiratory pressure. Excessive dyspnea and hypoxia were criteria for the premature termination of the session. Throughout the IRB run, arterial blood oxygen saturation, arterial blood pressure, and heart rate were monitored.
Main experimental trials
Testing was always performed at 10 am after a night rest of 8 hours in order to avoid possible influences of circadian rhythm variations, and a breakfast rich in carbohydrates was administrated 2 hours before testing (with a full dose of antioxidants and 600 mg of NAC). Each session’s duration was 45 minutes, and blood was drawn at rest (baseline), end (of session), and 120 minutes into recovery, through an intravenous catheter inserted in a forearm vein.
WBE
WBE was conducted at 70% of VO2 max for 45 minutes after a 10-minute warm-up period of increasing intensity (10 W/min). Heart rate and blood pressure were continuously monitored.19
IRB
Subjects begun the IRB run through the precustomized resistive load and were constantly encouraged to achieve pressures of 70% of their Pimax. Subjects were breathing in their own pattern, and the IRB run was terminated after 45 minutes. Throughout the IRB run, arterial blood oxygen saturation, arterial blood pressure, and heart rate were monitored.
Blood samples
Blood samples were collected into sterile syringes and were transferred to precooled sterile ethylenediaminetetraacetic acid tubes or sodium heparin tubes for flow cytometric evaluation (used within 2 hours from venipuncture).
Flow cytometric acquisition and analysis
Fluorescently labeled monoclonal antibodies against cell surface markers characteristic of lymphocyte subpopulations were used for staining. Counting was carried out with flow cytometry (Epics® Elite Esp.; Coulter Electronics, Beckman Coulter, Inc., Brea, CA, USA). We measured CD3, CD19, CD16, CD56, CD8, CD4, CD45RO, CD45RA, HLA-DR, and CD38. The percentages of total T cells (CD3+), B cells (CD19+), NK cells (CD3− CD16+ CD56+), T cytotoxic cells (CD3+ CD8+), T helper cells (CD3+ CD4+), memory T cells (CD45RO+, CD4+, or CD8+), naïve T cells (CD45RA+, CD4+, or CD8+), and markers of activation of CD8+ cells (HLA-DR+ and CD38+) were also measured.
Malondialdehyde (MDA) by high-performance liquid chromatography (MDA-HPLC) assay
The MDA levels as an index of oxidative stress and the efficacy of antioxidant administration were determined at the end of the WBE or IRB sessions, respectively. The MDA-HPLC method was mainly based on the procedure by Nielsen et al.21 The reagents potassium phosphate monobasic, potassium hydroxide, phosphoric acid (H3PO4), sodium hydroxide (NaOH), HPLC-grade methanol, and thiobarbituric acid (TBA) were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). Malondialdehyde tetrabutylammonium salt (96% purity) was also purchased from Sigma-Aldrich Co. and used for standards’ preparation.
For sample preparation, 100 μL of plasma was added to 700 μL of 1% H3PO4 and 200 μL of 42 mmol/L TBA. The mixture was heated for 30 minutes at 100°C and then ice-cooled for 2 minutes. A 250 μL of the aliquot was separated and added to 250 μL of 1 M NaOH in methanol (1:6). After centrifugation (5 minutes at 10,000 g) and filtration, 20 μL of the supernatant was injected in the HPLC apparatus at a flow rate of 1 mL/min, column temperature and pressure at 26°C and 2,200 psi, respectively. The absorbance was read at 532 nm.
For the MDA standard curve preparation, a stock solution of 100 μM of MDA was prepared in 0.01 mmol/L HCl. Dilutions from stock solution of 10–0.5 μmol/L were then performed. The same treatment described for plasma was used for the standard curve reaction.
Statistical analysis
Changes in the percentage of lymphocytes subpopulations over time (rest [baseline], end of WBE or IRB session, and 120 minutes into recovery) were compared by the nonparametric Friedman’s analysis of variance (ANOVA). The Wilcoxon matched pair test was used for post hoc analysis. The percentage of each cell subpopulation at iso-time points and the differences in cell percentages over time, before and after antioxidant supplementation, were compared with the Wilcoxon matched pair test. A p-value of 0.05 was initially considered as statistically significant and was accordingly adjusted using the sharper Bonferroni-type procedure for multiple comparisons described by Hochberg and Benjamini.22 Data are presented as mean values ± standard errors of the mean.
Results
MDA levels at end-exercise or IRB sessions
MDA levels at end exercise were 2.67±0.54 μmol/L before antioxidant supplementation and decreased to 1.86±0.42 μmol/L after antioxidant supplementation (p=0.023). MDA levels at end IRB were 2.14±0.41 μmol/L before antioxidant supplementation and decreased to 1.17±0.28 μmol/L after antioxidant supplementation (p=0.034).
Heart rate and arterial pressure measurements
Heart rate and mean arterial pressure were significantly higher in WBE compared with IRB (Wilcoxon matched pair test: p=0.0001 and p=0.005, respectively).
CD3+ cells (T-lymphocytes)
WBE
T-lymphocyte percentages did not change significantly in the WBE session regardless of antioxidant administration (Table 1).
 | Table 1 Lymphocyte subpopulation percentage changes during WBE |
IRB
T-lymphocyte percentages did not change significantly in the IRB session regardless of antioxidant administration (Table 2).
 | Table 2 Lymphocyte subpopulation percentage changes during IRB |
CD3+ CD4+ cells (T helper cells)
WBE
Both before and after antioxidant supplementation, T helper cell percentage was significantly reduced at exercise end compared with rest (p=0.04 for each) and was restored to rest values at recovery (before antioxidant supplementation, Friedman’s ANOVA x2=8.4, p=0.015; after antioxidant supplementation x2=7.6, p=0.02; Figure 2A). The degree of T helper cell percentage decrease at end-WBE compared with rest was not affected by antioxidants (Figure 2C).
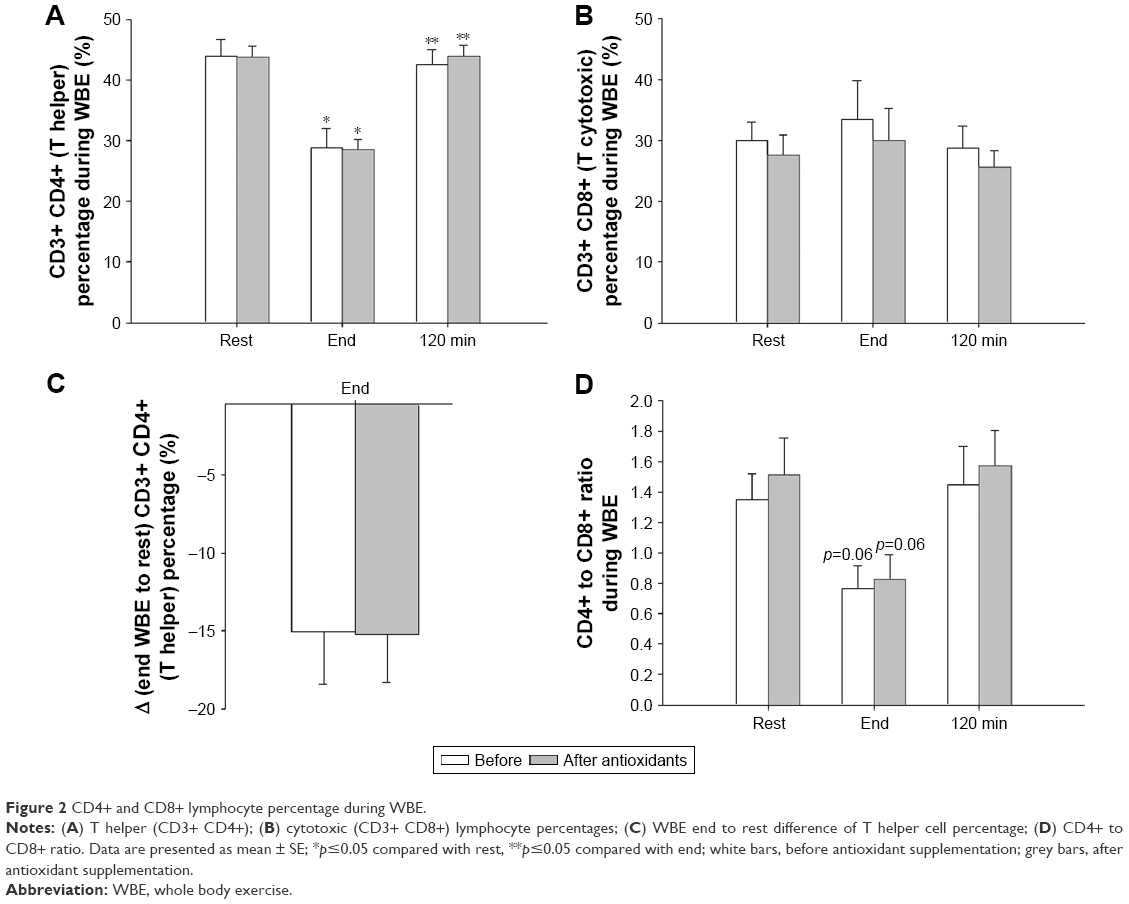 | Figure 2 CD4+ and CD8+ lymphocyte percentage during WBE. |
IRB
Before antioxidant supplementation, the percentage of T helper cells was significantly reduced at the end of resistive breathing compared with rest and were restored to pre-IRB levels at 120 minutes into recovery (Friedman’s ANOVA x2=6.3, p=0.042; Wilcoxon matched pair test, rest-end p=0.027, end-120 minutes p=0.043). Antioxidants attenuated T helper cell percentage decrease at IRB session’s end. (Friedman’s ANOVA x2=8.3, p=0.016; Wilcoxon matched pair test rest-end: p=0.07, end-120 minutes: p=0.027; Figure 3A). The magnitude of T helper cell percentage decrease at end-IRB compared with rest was significantly reduced after antioxidant supplementation (Wilcoxon matched pair test p=0.046; Figure 3B).
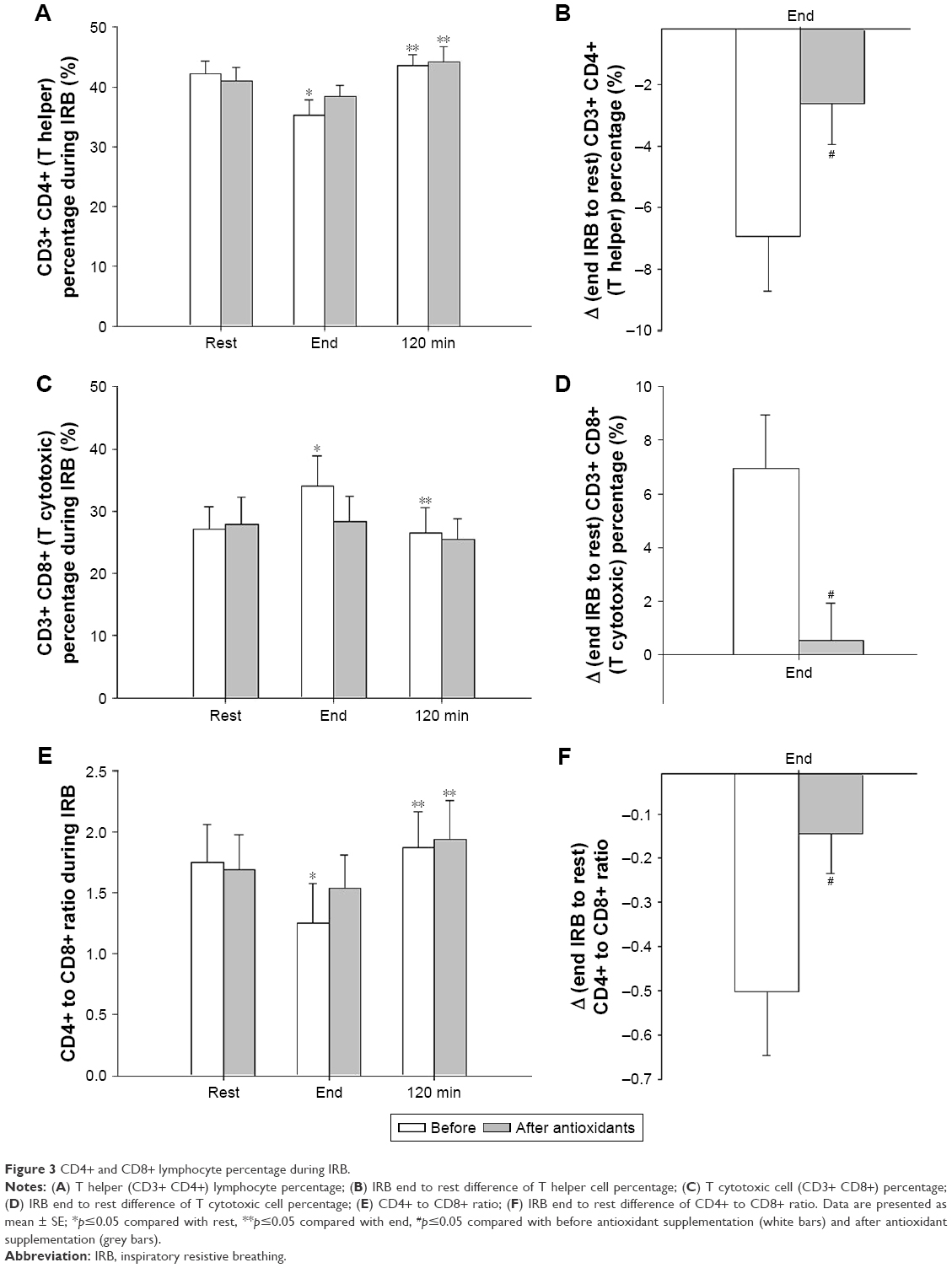 | Figure 3 CD4+ and CD8+ lymphocyte percentage during IRB. |
CD3+ CD8+ cells (cytotoxic T cells)
WBE
The percentage of cytotoxic T cells did not significantly change at either time point, before and after antioxidant supplementation (Figure 2B).
IRB
The percentage of cytotoxic T cells increased significantly after IRB and returned to pre-IRB levels at 120 minutes into recovery (Friedman’s ANOVA x2=7.0, p=0.03; Wilcoxon matched pair test rest-end p=0.046, end-120 minutes p=0.03). Antioxidant supplementation blunted the cytotoxic T cell percentage changes (Friedman’s ANOVA x2=3.0, p=0.22; Figure 3C). The IRB end-to-rest difference of cytotoxic T cell percentage was significantly greater before antioxidant supplementation compared with after antioxidant supplementation (p=0.05; Figure 3D).
CD4/CD8 ratio
WBE
The CD4/CD8 ratio changed significantly over time before (Friedman’s ANOVA x2=6.5, p=0.038) and after antioxidant supplementation (Friedman’s ANOVA x2=6.0, p=0.049). Both before and after antioxidant supplementation, the ratio tended to be reduced at exercise end compared with baseline (Wilcoxon matched pair test, p=0.06; Figure 2D).
IRB
The CD4/CD8 ratio before antioxidant supplementation was significantly decreased at IRB session end and was restored to baseline values at 120 minutes into recovery (Friedman’s ANOVA x2=9.0, p=0.011; Wilcoxon matched pair test rest-end p=0.028, end-120 minutes p=0.028). After antioxidant supplementation, no significant decrease at IRB end was observed compared with rest, although the ratio was increased at 120 minutes into recovery compared with IRB end (Friedman’s ANOVA x2=8.33, p=0.015; Wilcoxon matched pair test, rest-end p=0.17, end-120 minutes p=0.028). The end-to-rest difference of CD4/CD8 ratio was attenuated after antioxidant supplementation compared with before antioxidant supplementation (p=0.05; Figure 3E and F).
CD4+ CD45RA+ (Th cells expressing CD45RA)
WBE
CD45RA+ Th cell percentage was not affected by WBE either before or after antioxidants (Table 1).
IRB
The percentages of CD45RA+ Th cells did not significantly change either before or after antioxidant supplementation (Table 2).
CD4+ CD45RO+ (Th cells expressing CD45RO)
WBE
The percentages of Th cells expressing CD45RO were reduced postexercise (end) and returned to preexercise levels (rest) at 120 minutes into recovery, both before and after antioxidant supplementation (before: Friedman’s ANOVA x2=7.60, p=0.022; Wilcoxon matched pair test rest-end p=0.043; after: Friedman’s ANOVA x2=7.00, p=0.030; Wilcoxon matched pair test rest-end p=0.046; Figure 4A). The WBE end-to-rest difference of CD4+ CD45RO+ cells was not affected by antioxidants (Figure 4B).
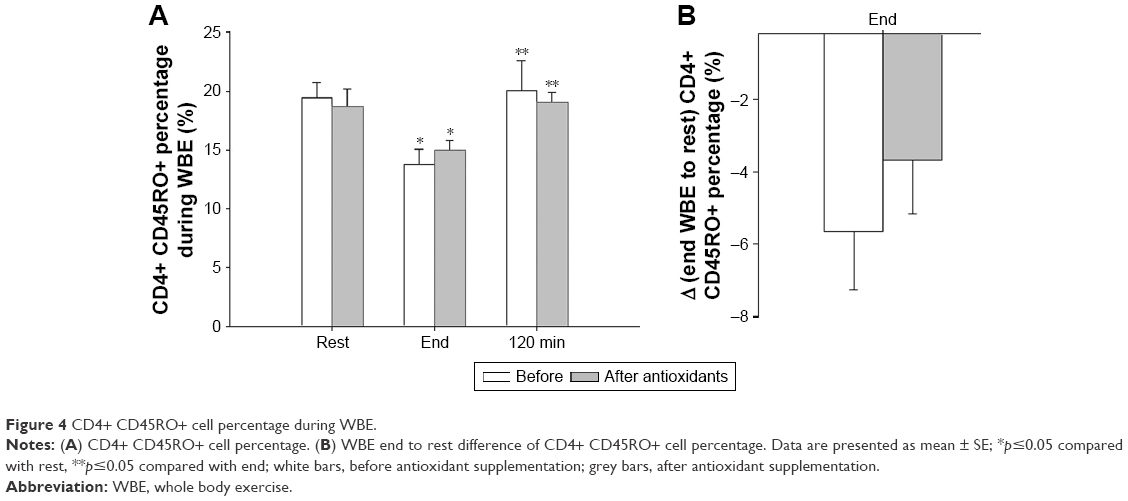 | Figure 4 CD4+ CD45RO+ cell percentage during WBE. |
IRB
The percentages of Th cell-expressing CD45RO did not change during IRB, neither before nor after antioxidant supplementation (Table 2).
CD8+ CD45RA+ (cytotoxic T cells expressing CD45RA)
WBE
The percentage of CD45RA+ cytotoxic T cells increased postexercise (end) and returned to preexercise levels (rest) at 120 minutes into recovery before antioxidant supplementation (Friedman’s ANOVA x2=7.60, p=0.022; Wilcoxon matched pair test; rest-end p=0.043, end-120 minutes p=0.043). In contrast, no significant change was observed after antioxidant supplementation (Friedman’s ANOVA x2=4.00, p=0.14). The iso-time comparison revealed a significant increase at rest percentages after antioxidant supplementation (Wilcoxon matched pair test p=0.043; Figure 5A). The WBE-end to rest increase was not affected by antioxidant supplementation (Figure 5C).
 | Figure 5 CD8+ CD45RA+ cell percentage during WBE and IRB. |
IRB
IRB increased the percentage of CD8+ CD45RA+ T cells after IRB session end that returned to rest values at 120 minutes into recovery (Friedman’s ANOVA x2=9.3, p=0.009; Wilcoxon matched pair test rest-end p=0.03, end-120 minutes p=0.03). After antioxidant supplementation, CD8+ CD45RA+ T cell percentage changes were blunted (Friedman’s ANOVA x2=5.20, p=0.07; Figure 5B). The IRB end to rest increase tended to be reduced after antioxidant supplementation (Wilcoxon matched pair test p=0.07; Figure 5D). The antioxidant-induced upregulation of CD45RA+ percentage at rest of WBE (Figure 5A) was not observed at rest before IRB (Figure 5B).
CD8+ CD45RO+ (cytotoxic T cells expressing CD45RO)
WBE
The percentage of CD45RO+ cytotoxic T cells did not change after WBE irrespective of antioxidant supplementation (Table 1).
IRB
The CD8+ CD45RO+ remained unchanged during IRB both before and after antioxidant supplementation (Table 2).
CD8+ CD38+ cells
WBE
CD8+ CD38+ percentage changes before and after antioxidant supplementation were not significant (Friedman’s ANOVA x2=2.21, p=0.33; Friedman’s ANOVA x2=1.0, p=0.60, respectively). The iso-time comparison revealed that antioxidants upregulated CD8+ CD38+ percentages significantly at 120 minutes into recovery (Wilcoxon matched pair test p=0.043; Table 1).
IRB
Before antioxidant supplementation, the percentage of CD8+ CD38+ cells was not affected from IRB (Friedman’s ANOVA x2=3.5, p=0.17). After antioxidant supplementation, an increase in CD8+ CD38+ cell percentage at IRB end was observed compared with rest. CD8+ CD38+ cell percentage returned to pre-IRB levels at 120 minutes into recovery (Friedman’s ANOVA x2=8.4, p=0.014; Wilcoxon matched pair test rest-to-end p=0.043, end-120 minutes p=0.027; Figure 6A). Iso-time comparisons revealed that the end IRB percentage was higher after antioxidant supplementation (Wilcoxon matched pair test p=0.043; Figure 6A). Indeed, the increase in CD8+ CD38+ cell percentage after antioxidant supplementation was significantly greater than that before antioxidant supplementation (Wilcoxon matched pair test p=0.04; Figure 6B).
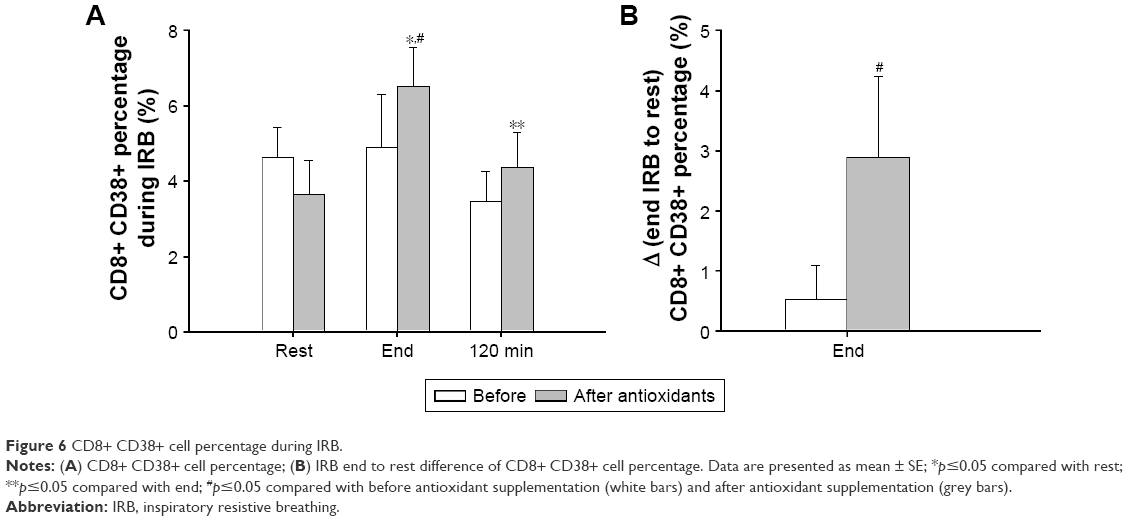 | Figure 6 CD8+ CD38+ cell percentage during IRB. |
CD8+ HLA-DR+ cells
WBE
CD8+ HLA-DR+ cell percentage did not change significantly over time neither before nor after antioxidant supplementation (Table 1).
IRB
CD8+ HLA-DR+ percentage changes were not significant before and after antioxidant supplementation (Table 2).
CD3− CD16+ CD56+ (NK cells)
WBE
NK cell percentages significantly changed over time, regardless of antioxidant supplementation (before antioxidant supplementation: Friedman’s ANOVA x2=8.4, p=0.015; after antioxidant supplementation: Friedman’s ANOVA x2=9.3, p=0.009). Both before and after antioxidant supplementation, NK cell percentage increased postexercise (end) compared with rest (p=0.04 and p=0.02, respectively) and went back to preexercise levels at 120 minutes into recovery (Figure 7A). The difference in NK cell percentage postexercise (end) relative to rest did not differ before and after antioxidant supplementation (Figure 7C).
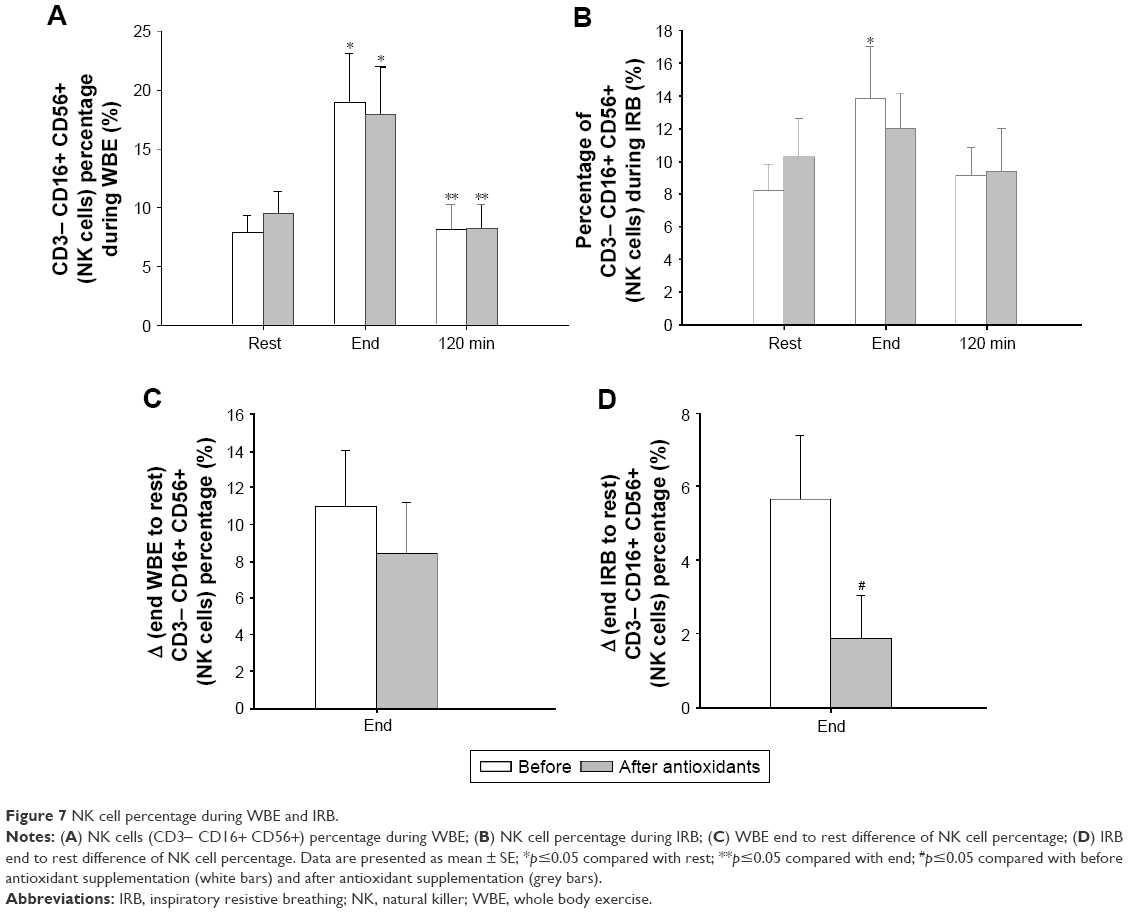 | Figure 7 NK cell percentage during WBE and IRB. |
IRB
Before antioxidant supplementation, NK cell percentage increased at IRB end (Friedman’s ANOVA x2=6.3, p=0.042; end IRB to rest, p=0.028). Antioxidant supplementation blunted the IRB-induced increase in NK cell percentage (Friedman’s ANOVA x2=4.3, p=0.11; Figure 7B). The end-to-rest difference of NK cell percentages was significantly greater before antioxidant supplementation compared with after antioxidant supplementation (p=0.027; Figure 7D).
CD19+ (B cells)
WBE
Both before and after antioxidant supplementation, B-cell percentage decreased postexercise (end) and was restored to rest levels at 120 minutes into recovery (Friedman’s ANOVA, before antioxidants: x2=7.6, p=0.02; after antioxidants, x2=9.3, p=0.009; Figure 8A). The reduction of B-cell percentage at end WBE relative to rest was attenuated after antioxidant supplementation, but this effect did not reach statistical significance, compared with before antioxidant supplementation (p=0.06; Figure 8B).
 | Figure 8 B-lymphocyte percentage during WBE. |
IRB
IRB did not produce any changes in the percentage of B cells regardless of antioxidant supplementation (Table 2).
Discussion
The major finding of our study is that IRB induces significant alterations in peripheral blood lymphocyte subpopulations as does WBE, and this effect is blunted by antioxidants mainly in resistive breathing. During IRB, NK and cytotoxic T-cell percentages increase, T helper cell percentage is reduced, and CD4/CD8 ratio is depressed under the influence of oxidative stress. Two hours into recovery in WBE and IRB, the percentage of most lymphocyte subsets returns to rest levels.
Lymphocyte subset response to WBE and IRB
During exercise, all lymphocytes are recruited to the intravascular space, yet each subset responds differently.1
NK cells are the most sensitive subset to exercise showing a 10-fold increase in the last minute of a short bout (20 minutes) of intense (85% of VO2 max) exercise, while T helper and B cells increase only by 50%–100%.23 Women exercising at 60% of VO2 max for 45 minutes demonstrated an increase in lymphocyte count at exercise end lasting for 1.5 hours with NK cells accounting for two thirds of and with T cells the remaining one third of lymphocytosis.24 Exercise intensity positively correlates with the postexercise increase of the NK cell percentage.25,26
Our findings during WBE confirm these observations. We measured NK cells (CD56+ CD16+) that are considered more responsive to exercise.23,27 After a WBE session with an intensity of 70% of VO2 max, NK cell percentage nearly doubled and was restored to preexercise levels at 2 hours into recovery.26,28 Interestingly, antioxidants had no effect on the aforementioned alterations. To the best of our knowledge, our study is the first to describe NK cell response after resistive breathing. IRB led to a significant rise of NK cell percentage (at end) that returned to rest levels after 2 hours as in WBE. Antioxidants blunted this response, which suggests that oxidative stress is a stimulus for the NK cell increase secondary to resistive breathing.
CD8+ lymphocytes (cytotoxic T cells) are also mobilized and recruited into the vascular compartment in proportion to exercise intensity. Campbell et al have shown an almost 5 times increase in the percentage of cytotoxic T cells after a short session of intense exercise.23 The most actively cytotoxic T cells, the reexpressing CD45RA (CD3+ CD8+ CD45RA+), demonstrate the highest exercise responsiveness.23 Accordingly, the CD45RA+ cell percentage markedly increased after WBE in our experiment. We show here for the first time that the CD45RA+ cell percentage increases during IRB. Another novel finding of our study is that antioxidants abolished the CD45RA+ cell response to IRB and WBE, suggesting that oxidative stress is mediating this response.
CD4+ T helper cells exhibit a more “modest” response to exercise increasing by 50%–100%;23 thus, a depressed CD4/CD8 ratio is expected postexercise.1,24,29 We found that T helper cell percentage was depressed at WBE and IRB end probably due to a higher recruitment of cytotoxic and NK cells in the endovascular compartment, resulting in a depressed CD4/CD8 ratio. Antioxidants attenuated T helper cell percentage and CD4/CD8 reduction only at IRB end. We also found a significant decrease in the percentage of CD4+ CD45RO+ cells at WBE end regardless of antioxidant administration.
In agreement with the literature, antioxidant supplementation tended to upregulate CD8+ CD38+ percentage.30 At 120 minutes into recovery of WBE, CD8+ CD38+ percentage was increased after antioxidant supplementation compared with before antioxidant supplementation. During IRB, CD8+ CD38+ percentage changes were observed only after antioxidant supplementation.
B-lymphocytes (CD19+) seem to be less responsive to exercise. Studies have shown that B-lymphocytes are relatively unaffected by exercise,31,32 whereas others showed an increase during repeated bouts of acute exercise and a reduction in the recovery period.33 We found a significant decrease in their percentage at WBE end and restoration to baseline values 2 hours into recovery regardless of antioxidant administration. By contrast, no change in B-lymphocyte percentage was observed during IRB.
Lymphocyte migration control and redistribution
Lymphocytosis in peripheral blood is a stereotypical result of environmental stress such as surgery, burn, trauma, sepsis, and exercise.34 The immediate postexercise lymphocytosis reflects the transition from reservoirs to target organs where lymphocytes may encounter pathogen invasion or antigen exposure. Consequently, the recovery lymphocytopenia or return to preexercise values is a result of the evolution completion of the migration process. Thus, changes in the percentage of lymphocyte subsets in the vascular compartment at different time points of a stress event (eg, exercise) are part of their functional redistribution to various body compartments. Migration is regulated by adhesion molecules and cell migration cytokines (chemokines), yet the exact control mechanism (mechanisms) remains (remain) relatively elusive. Adrenergic stimulation and, to a lesser extent, steroids and apoptosis have been suggested.35,36
Peripheral blood lymphocyte count and catecholamines are directly related, namely the lymphocytosis during/after stress is dose-dependent on the amount of adrenaline and noradrenaline that are secreted.37 In animal models, exogenous administration of epinephrine increased peripheral blood lymphocytes as observed in exercise.36 NK cells present the highest expression of β2-adrenergic receptors followed by cytotoxic T cells; consequently, the aforementioned subgroups are proportionally responsive to exercise intensity.38 IRB at 70% of Pimax represents a smaller adrenergic response to WBE (at 70% of VO2 max). Interestingly, heart rate and mean arterial pressure (which are indirect measures of adrenergic stimulation) were significantly higher in WBE compared with IRB. Indeed, doubling the work of breathing does not increase sympathetic activation in healthy subjects and COPD patients.39 IRB-induced lymphocyte subset alterations may be influenced by factors other than (a weak) sympathetic activation.
Oxidative stress and exercise
The link between exercise and oxidative stress is firmly established.7,8,40–42 Working muscles are considered the main source of oxidative products (electron leakage from mitochondria, oxidases, myostatin, phospholipase A2, etc.), but not the only one.43 Exercise-induced muscle injury may activate through cytokine production (IL-1β and tumor necrosis factor-alpha [TNF-α]) neutrophils and macrophages, cells capable of producing large amounts of ROS through oxidative burst, their indigenous defense mechanism.44 Endothelial cells and catecholamines are also implicated in ROS generation.43 Oxidative stress derivatives are regulators of cell function and signaling molecules integrated in exercise physiology.45 Our group has shown that oxidative stress is a stimulus for the cytokine induction after both IRB13 and WBE19 and that ROS stimulate IL-6 production from skeletal myotubes.46
We have administrated a cocktail of antioxidants in nonathletes that has already proven effective in attenuating the plasma cytokine response after WBE19 and IRB.13 The rationale behind the composition of the mixture is thoroughly analyzed in our previous work.13,19 Vitamin E is an efficient lipid-soluble free radical scavenger protecting biomembranes, while vitamin C is a water-soluble antioxidant that along with vitamin A can directly scavenge singlet oxygen, superoxide, and hydroxyl radicals in both intra- and extracellular fluid. Allopurinol inhibits the ROS-generating xanthine oxidase, and NAC forms L-cysteine, which is further metabolized to glutathione, a key biological antioxidant molecule.13 The mixture was effective in reducing lipid peroxidation as evidenced by the reduced MDA levels detected postexercise (end) or post-IRB (end) after antioxidant administration. Treatment with allopurinol efficiently inhibits blood glutathione oxidation and lipid peroxidation after strenuous exercise in COPD patients.47 We have used a cocktail of antioxidants with the rationale that multiple mechanisms and sources of oxidative stress exist within cells; thus, a single antioxidant would be unlikely to be effective. We have chosen a relatively long period of administration on the premise that some antioxidants like vitamin E require a long time to become incorporated into membranes.48 On the other hand, as we have already mentioned, the mixture of antioxidants that we used prevented us from adopting a randomized crossover design due to their residual effects, ie, the plasma concentration of vitamin E reached peak values after 15 days of supplementation and maintained this plateau for the next 15 days,49 and the serum concentration of carotenoids peaked at 24–48 hours after a single dose and returned to baseline after 7 days.50 Other investigators who used only one antioxidant in trained athletes failed to produce alterations in lymphocyte subsets.6 We used untrained healthy volunteers and not athletes who present an upregulated antioxidant capacity secondary to training.43,51 Our results suggest that an oxidative stress-dependent pathway regulates lymphocyte redistribution during IRB and possibly in obstructive lung diseases.
The mechanisms by which oxidative stress may regulate lymphocyte redistribution during IRB are not known and are not the scope of this study. However, some speculations are worth pursuing. Lymphocyte migration to and out of lymphoid and nonlymphoid organs presupposes a stepwise chemokine-controlled interaction with the endothelium. The transmigration through the endothelium includes the capture and roll at the vessel wall mediated primarily by selectins and integrins (rolling and adhesion). Activation and transendothelial migration are mediated by adhesion molecules that are present in the membranes of lymphocytes and endothelial cells, the intracellular adhesion molecule 1 (ICAM-1) and vascular intracellular adhesion molecule 1 (VCAM-1).35 ICAMs are upregulated by IL-6, IL-1β, and TNF-α, the three cytokines that were abolished after antioxidant supplementation in IRB.13 The ICAM-1 gene is induced by IL-6 and TNF-α mediated via activator of transcription-3 (Stat3) and nuclear factor-κB.52 TNF-α and IL-1β also induced the expression of ICAM-1 and VCAM-1 by endothelial cells.53 It is therefore tempting to speculate that IL-6, TNF-α, and IL-1β precipitate lymphocyte migration independently of the stress neurohormonal stimulation through the upregulation of adhesion chemokines that are essential in lymphocyte transendothelial migration. In our experiment, antioxidant supplementation may have blunted IRB-induced cytokine release and consequently lymphocyte redistribution. In contrast, WBE lymphocyte subset alterations may be dominated by stress-induced neurohormonal mechanisms that are less influenced by antioxidants and cytokines.
Comparison between WBE and IRB
Although we have used IRB as a model to isolate the effects of respiratory muscle activation, we cannot exclude respiratory muscle activation during WBE. It is known that WBE results in increased inflammation and oxidative stress. One potential source of the immune response observed during WBE may relate to the respiratory muscle metaboreflex/fatigue, which accompanies aerobic exercise. It is known that WBE produces diaphragmatic fatigue when intensity exceeds 85% of VO2 max.54 We have chosen a submaximal exercise intensity of 70% of VO2 max, which is not likely to induce respiratory muscle fatigue. We did not aim for diaphragmatic fatigue through our IRB session, and even with high Pi target (of 70% Pimax), we instructed subjects to breathe in their own pattern with a relatively high respiratory rate (20–25) and inspiratory time/total inspiratory and expiratory time ratio <0.6 using all respiratory muscles, not just the diaphragm. Therefore, all our subjects completed a 45-minute session of IRB, and Pimax at the end of the session was ±10% of initial Pimax. St Croix et al noticed an increase in muscle nerve sympathetic activity (peroneal nerve) and small increases in mean arterial blood pressure and heart rate only when they used a protocol of resistive breathing designed to produce diaphragmatic fatigue (Pi =60% Pimax, inspiratory time/total inspiratory and expiratory time ratio =0, 7, Vt =2x resting Vt and RR =15 breaths/min for 7±3 minutes until exhaustion).55 Sheel et al triggered a similar metaboreflex reducing the blood flow in the femoral artery in a similar protocol of resistive breathing that produced diaphragmatic fatigue identified through changes in mouth twitch pressure in response to bilateral phrenic nerve stimulation, while mean arterial pressure remained unchanged.56 In contrast, increased voluntary ventilation and inspiratory effort even with a Pi close to Pimax (95%) did not produce a sympathetic response.55,56 Taken together, although we cannot exclude the presence of respiratory muscle metaboreflex/fatigue with certainty, we believe that either our IRB protocol or our WBE protocol was not likely to induce such a metaboreflex.
Clinical implications
Obstructive pulmonary diseases such as COPD and asthma are characterized by increased airway resistance that is aggravated during exacerbations and may lead to respiratory failure and death. Our group has focused on the consequences of airway obstruction per se and has created an experimental model of breathing through increased airway resistance (resistive breathing) that was applied on both humans and animals.
In each subject, we applied inspiratory resistive load in order to achieve breathing inspiratory pressures close to 70% of their Pimax. The load we used was extrapolated from data of patients with severe COPD. Direct measurements of the actual load that respiratory muscles face during an exacerbation are extremely difficult since physiological experiments are not conducted in life-threating clinical conditions. However, indirect estimates can be made. In patients with COPD who require mechanical ventilation, respiratory muscles face similar load as in severe exacerbation. Our group showed that in COPD patients requiring invasive mechanical ventilation, the peak/maximum inspiratory pressure value was 0.62±0.15,57 and failure to wean from mechanical ventilation was accompanied by Pimean/Pimax of 0.490±0.09.58
We applied only inspiratory resistance, and this is a limitation of our study. One may argue that our human model is similar to extrathoracic obstruction (upper airway obstruction), where the resistive load is primary inspiratory, and that in COPD and asthma the obstruction is intrathoracic and the resistance is higher during expiration. Nevertheless, during asthma and COPD exacerbations, excess bronchoconstriction induces air-trapping and large negative intrathoracic pressures during inspiration to counteract the intrinsic positive end-expiratory pressure and the increased airway resistance.18 Therefore, our model of IRB, where respiratory muscles contract strenuously and lungs are exposed to large negative intrathoracic pressure gradients, mimics key components of the respiratory pathophysiology of obstructive lung disease, especially during exacerbations. This IRB may induce cytokine upregulation13 and, as shown in the current study, immune alterations.
Increased numbers of T-lymphocytes are observed in lung parenchyma, distal and proximal airways of COPD patients with a predominance of CD8+ over CD4+ cells.16 T-cell numbers are correlated to the extent of alveolar destruction and the severity of airflow obstruction, especially CD8+ cells that induce apoptosis of alveolar cells and cytolysis through the release of perforins, granzyme B, and TNF-α.59,60 COPD patients also show altered peripheral blood lymphocyte subsets that are also correlated to the degree of airway obstruction. The percentage of peripheral blood CD8+ T-lymphocytes was higher in COPD patients compared with nonsmoking healthy controls. Furthermore, patients with low FEV1 had significantly higher CD8+ T-lymphocyte percentage in peripheral blood.61 Smokers with COPD and predominantly small airway disease had higher CD8+ T-lymphocyte percentage in peripheral blood than “healthy” smokers.62
We have shown here for the first time that a brief simulation of airway obstruction (IRB) in healthy humans increases CD8+ T-lymphocytes percentages in peripheral blood in an oxidative stress-dependent pathway. It is tempting to speculate that bouts of increased airway resistance, such as those observed during IRB or COPD exacerbation, increase CD8+ T-lymphocytes in the peripheral circulation and thence to the lung, thus contributing to the increased CD8+ T-lymphocyte levels observed. This would lead to bouts of CD8+ T-cell-mediated lung destruction and would partly explain why COPD patients with frequent exacerbations have more rapid decline in lung function.63,64
NK cell percentages in the peripheral blood and the lung do not differ between stable COPD patients and healthy subjects.59,61,65 However, peripheral blood NK cell cytotoxic activity is impaired in COPD patients.65 In our experiment, IRB resulted in an increase in the NK cells, which was reversed after the end of IRB. This response to the “mechanical stressor” of increased airway resistance (ie, IRB) would be adaptive in the case of COPD exacerbation (which is also associated with increased airway resistance). This is because COPD exacerbations are usually infective in origin, and the increased NK cells would lead to increased immune surveillance against the infective pathogens. However, in the case of COPD exacerbations, the relief of increased airway resistance with bronchodilators would decrease the NK cell percentage in the blood and would lead to “relative immunosuppression” in the face of the offending infective agent causing exacerbation.
Conclusion
We have shown for the first time that IRB induces lymphocyte subset alterations through an oxidative stress-dependent pathway. Lymphocytes are key immune cells that regulate both innate and adaptive immunity. Increased airway resistance may not only induce inflammation and lung injury, but also precipitate, through oxidative stress, peripheral immune alterations.
Acknowledgments
The authors would like to acknowledge the contribution of Dimitri Stathopoulos for proofreading the final draft of this paper. This study was funded by the Thorax Foundation, Athens, Greece.
Disclosure
The authors report no conflicts of interest in this work.
References
Pedersen BK, Hoffman-Goetz L. Exercise and the immune system: regulation, integration, and adaptation. Physiol Rev. 2000;80(3):1055–1081. | ||
Nieman DC. Immune response to heavy exertion. J Appl Physiol (1985). 1997;82(5):1385–1394. | ||
Kruger K, Mooren FC. Exercise-induced leukocyte apoptosis. Exerc Immunol Rev. 2014;20:117–134. | ||
Kruger K, Frost S, Most E, Volker K, Pallauf J, Mooren FC. Exercise affects tissue lymphocyte apoptosis via redox-sensitive and Fas-dependent signaling pathways. Am J Physiol Regul Integr Comp Physiol. 2009;296(5):R1518–R1527. | ||
Tsai K, Hsu TG, Hsu KM, et al. Oxidative DNA damage in human peripheral leukocytes induced by massive aerobic exercise. Free Radic Biol Med. 2001;31(11):1465–1472. | ||
Nielsen HB, Secher NH, Kappel M, Pedersen BK. N-acetylcysteine does not affect the lymphocyte proliferation and natural killer cell activity responses to exercise. Am J Physiol. 1998;275(4 Pt 2):R1227–R1231. | ||
Vassilakopoulos T, Hussain SN. Ventilatory muscle activation and inflammation: cytokines, reactive oxygen species, and nitric oxide. J Appl Physiol (1985). 2007;102(4):1687–1695. | ||
Borzone G, Zhao B, Merola AJ, Berliner L, Clanton TL. Detection of free radicals by electron spin resonance in rat diaphragm after resistive loading. J Appl Physiol (1985). 1994;77(2):812–818. | ||
Vassilakopoulos T, Roussos C, Zakynthinos S. The immune response to resistive breathing. Eur Respir J. 2004;24(6):1033–1043. | ||
Vassilakopoulos T, Divangahi M, Rallis G, et al. Differential cytokine gene expression in the diaphragm in response to strenuous resistive breathing. Am J Respir Crit Care Med. 2004;170(2):154–161. | ||
Sigala I, Zacharatos P, Boulia S, et al. Nitric oxide regulates cytokine induction in the diaphragm in response to inspiratory resistive breathing. J Appl Physiol (1985). 2012;113(10):1594–1603. | ||
Vassilakopoulos T, Zakynthinos S, Roussos C. Strenuous resistive breathing induces proinflammatory cytokines and stimulates the HPA axis in humans. Am J Physiol. 1999;277(4 Pt 2):R1013–R1019. | ||
Vassilakopoulos T, Katsaounou P, Karatza MH, Kollintza A, Zakynthinos S, Roussos C. Strenuous resistive breathing induces plasma cytokines: role of antioxidants and monocytes. Am J Respir Crit Care Med. 2002;166(12 Pt 1):1572–1578. | ||
Toumpanakis D, Kastis GA, Zacharatos P, et al. Inspiratory resistive breathing induces acute lung injury. Am J Respir Crit Care Med. 2010;182(9):1129–1136. | ||
Loverdos K, Toumpanakis D, Litsiou E, et al. The differential effects of inspiratory, expiratory, and combined resistive breathing on healthy lung. Int J Chron Obstruct Pulmon Dis. 2016;11:1623–1638. | ||
Barnes PJ. Inflammatory mechanisms in patients with chronic obstructive pulmonary disease. J Allergy Clin Immunol. 2016;138(1):16–27. | ||
Leidinger P, Keller A, Heisel S, et al. Novel autoantigens immunogenic in COPD patients. Respir Res. 2009;10:20. | ||
Global Initiative for Chronic Obstructive Lung Disease (GOLD). Global Strategy for Diagnosis, Management, and Prevention of COPD – 2016. Available from: http://goldcopd.org/global-strategy-diagnosis-management-prevention-copd-2016/. Accessed November 3, 2016. | ||
Vassilakopoulos T, Karatza MH, Katsaounou P, Kollintza A, Zakynthinos S, Roussos C. Antioxidants attenuate the plasma cytokine response to exercise in humans. J Appl Physiol (1985). 2003;94(3):1025–1032. | ||
Keast D, Cameron K, Morton AR. Exercise and the immune response. Sports Med. 1988;5(4):248–267. | ||
Nielsen F, Mikkelsen BB, Nielsen JB, Andersen HR, Grandjean P. Plasma malondialdehyde as biomarker for oxidative stress: reference interval and effects of life-style factors. Clin Chem. 1997;43(7):1209–1214. | ||
Hochberg Y, Benjamini Y. More powerful procedures for multiple significance testing. Stat Med. 1990;9(7):811–818. | ||
Campbell JP, Riddell NE, Burns VE, et al. Acute exercise mobilises CD8+ T lymphocytes exhibiting an effector-memory phenotype. Brain Behav Immun. 2009;23(6):767–775. | ||
Nieman DC, Nehlsen-Cannarella SL, Donohue KM, et al. The effects of acute moderate exercise on leukocyte and lymphocyte subpopulations. Med Sci Sports Exerc. 1991;23(5):578–585. | ||
Nieman DC, Miller AR, Henson DA, et al. Effects of high- vs moderate-intensity exercise on natural killer cell activity. Med Sci Sports Exerc. 1993;25(10):1126–1134. | ||
Tvede N, Kappel M, Halkjaer-Kristensen J, Galbo H, Pedersen BK. The effect of light, moderate and severe bicycle exercise on lymphocyte subsets, natural and lymphokine activated killer cells, lymphocyte proliferative response and interleukin 2 production. Int J Sports Med. 1993;14(5):275–282. | ||
Timmons BW, Cieslak T. Human natural killer cell subsets and acute exercise: a brief review. Exerc Immunol Rev. 2008;14:8–23. | ||
Yu DT, Clements PJ, Pearson CM. Effect of corticosteroids on exercise-induced lymphocytosis. Clin Exp Immunol. 1977;28(2):326–331. | ||
Fry RW, Morton AR, Crawford GP, Keast D. Cell numbers and in vitro responses of leucocytes and lymphocyte subpopulations following maximal exercise and interval training sessions of different intensities. Eur J Appl Physiol Occup Physiol. 1992;64(3):218–227. | ||
Sandoval-Montes C, Santos-Argumedo L. CD38 is expressed selectively during the activation of a subset of mature T cells with reduced proliferation but improved potential to produce cytokines. J Leukoc Biol. 2005;77(4):513–521. | ||
Shek PN, Sabiston BH, Buguet A, Radomski MW. Strenuous exercise and immunological changes: a multiple-time-point analysis of leukocyte subsets, CD4/CD8 ratio, immunoglobulin production and NK cell response. Int J Sports Med. 1995;16(7):466–474. | ||
Natale VM, Brenner IK, Moldoveanu AI, Vasiliou P, Shek P, Shephard RJ. Effects of three different types of exercise on blood leukocyte count during and following exercise. Sao Paulo Med J. 2003;121(1):9–14. | ||
Nielsen HB, Secher NH, Christensen NJ, Pedersen BK. Lymphocytes and NK cell activity during repeated bouts of maximal exercise. Am J Physiol. 1996;271(1 Pt 2):R222–R227. | ||
Hoffman-Goetz L, Pedersen BK. Exercise and the immune system: a model of the stress response? Immunol Today. 1994;15(8):382–387. | ||
Kruger K, Mooren FC. T cell homing and exercise. Exerc Immunol Rev. 2007;13:37–54. | ||
Kruger K, Lechtermann A, Fobker M, Volker K, Mooren FC. Exercise-induced redistribution of T lymphocytes is regulated by adrenergic mechanisms. Brain Behav Immun. 2008;22(3):324–338. | ||
Bosch JA, Berntson GG, Cacioppo JT, Dhabhar FS, Marucha PT. Acute stress evokes selective mobilization of T cells that differ in chemokine receptor expression: a potential pathway linking immunologic reactivity to cardiovascular disease. Brain Behav Immun. 2003;17(4):251–259. | ||
Turner JE, Bosch JA, Aldred S. Measurement of exercise-induced oxidative stress in lymphocytes. Biochem Soc Trans. 2011;39(5):1299–1304. | ||
Raupach T, Bahr F, Herrmann P, Lüthje L, Hasenfuss G, Andreas S. Inspiratory resistive loading does not increase sympathetic tone in COPD. Respir Med. 2010;104(1):107–113. | ||
Reid MB. Invited review: redox modulation of skeletal muscle contraction: what we know and what we don’t. J Appl Physiol (1985). 2001;90(2):724–731. | ||
Jackson MJ, Pye D, Palomero J. The production of reactive oxygen and nitrogen species by skeletal muscle. J Appl Physiol (1985). 2007;102(4):1664–1670. | ||
Ferreira LF, Reid MB. Muscle-derived ROS and thiol regulation in muscle fatigue. J Appl Physiol (1985). 2008;104(3):853–860. | ||
Steinbacher P, Eckl P. Impact of oxidative stress on exercising skeletal muscle. Biomolecules. 2015;5(2):356–377. | ||
Peake J, Suzuki K. Neutrophil activation, antioxidant supplements and exercise-induced oxidative stress. Exerc Immunol Rev. 2004;10:129–141. | ||
Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol. 2000;279(6):L1005–L1028. | ||
Kosmidou I, Vassilakopoulos T, Xagorari A, Zakynthinos S, Papapetropoulos A, Roussos C. Production of interleukin-6 by skeletal myotubes: role of reactive oxygen species. Am J Respir Cell Mol Biol. 2002;26(5):587–593. | ||
Heunks LM, Viña J, van Herwaarden CL, Folgering HT, Gimeno A, Dekhuijzen PN. Xanthine oxidase is involved in exercise-induced oxidative stress in chronic obstructive pulmonary disease. Am J Physiol. 1999;277(6 Pt 2):R1697–R1704. | ||
Cannon JG, Meydani SN, Fielding RA, et al. Acute phase response in exercise. II. Associations between vitamin E, cytokines, and muscle proteolysis. Am J Physiol. 1991;260(6 Pt 2):R1235–R1240. | ||
Meydani M, Fielding RA, Cannon JG, Blumberg JB, Evans WJ. Muscle uptake of vitamin E and its association with muscle fiber type. J Nutr Biochem. 1997;8(2):74–78. | ||
Johnson EJ, Russell RM. Distribution of orally administered beta-carotene among lipoproteins in healthy men. Am J Clin Nutr. 1992;56(1):128–135. | ||
Sachdev S, Davies KJ. Production, detection, and adaptive responses to free radicals in exercise. Free Radic Biol Med. 2008;44(2):215–223. | ||
Wung BS, Ni CW, Wang DL. ICAM-1 induction by TNFalpha and IL-6 is mediated by distinct pathways via Rac in endothelial cells. J Biomed Sci. 2005;12(1):91–101. | ||
O’Carroll SJ, Kho DT, Wiltshire R, et al. Pro-inflammatory TNFalpha and IL-1beta differentially regulate the inflammatory phenotype of brain microvascular endothelial cells. J Neuroinflammation. 2015;12:131. | ||
Johnson BD, Babcock MA, Suman OE, Dempsey JA. Exercise-induced diaphragmatic fatigue in healthy humans. J Physiol. 1993;460:385–405. | ||
St Croix CM, Morgan BJ, Wetter TJ, Dempsey JA. Fatiguing inspiratory muscle work causes reflex sympathetic activation in humans. J Physiol. 2000;529(Pt 2):493–504. | ||
Sheel AW, Derchak PA, Pegelow DF, Dempsey JA. Threshold effects of respiratory muscle work on limb vascular resistance. Am J Physiol Heart Circ Physiol. 2002;282(5):H1732–H1738. | ||
Zakynthinos SG, Vassilakopoulos T, Roussos C. The load of inspiratory muscles in patients needing mechanical ventilation. Am J Respir Crit Care Med. 1995;152(4 Pt 1):1248–1255. | ||
Vassilakopoulos T, Zakynthinos S, Roussos C. The tension-time index and the frequency/tidal volume ratio are the major pathophysiologic determinants of weaning failure and success. Am J Respir Crit Care Med. 1998;158(2):378–385. | ||
Majo J, Ghezzo H, Cosio MG. Lymphocyte population and apoptosis in the lungs of smokers and their relation to emphysema. Eur Respir J. 2001;17(5):946–953. | ||
Saetta M, Di Stefano A, Turato G, et al. CD8+ T-lymphocytes in peripheral airways of smokers with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 1998;157(3 Pt 1):822–826. | ||
de Jong JW, van der Belt-Gritter B, Koeter GH, Postma DS. Peripheral blood lymphocyte cell subsets in subjects with chronic obstructive pulmonary disease: association with smoking, IgE and lung function. Respir Med. 1997;91(2):67–76. | ||
Kim WD, Kim WS, Koh Y, et al. Abnormal peripheral blood T-lymphocyte subsets in a subgroup of patients with COPD. Chest. 2002;122(2):437–444. | ||
Donaldson GC, Seemungal TA, Bhowmik A, Wedzicha JA. Relationship between exacerbation frequency and lung function decline in chronic obstructive pulmonary disease. Thorax. 2002;57(10):847–852. | ||
Makris D, Moschandreas J, Damianaki A, et al. Exacerbations and lung function decline in COPD: new insights in current and ex-smokers. Respir Med. 2007;101(6):1305–1312. | ||
Prieto A, Reyes E, Bernstein ED, et al. Defective natural killer and phagocytic activities in chronic obstructive pulmonary disease are restored by glycophosphopeptical (inmunoferon). Am J Respir Crit Care Med. 2001;163(7):1578–1583. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.