Back to Journals » Clinical Ophthalmology » Volume 19
Immune Camouflage in Pythium insidiosum Keratitis: A Hypothesis on Molecular Mimicry and Host Pattern Recognition Receptor Evasion
Received 13 August 2025
Accepted for publication 11 December 2025
Published 19 December 2025 Volume 2025:19 Pages 4855—4873
DOI https://doi.org/10.2147/OPTH.S560311
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Scott Fraser
Bharat Gurnani,1 Kirandeep Kaur2
1Department of Cataract, Cornea and Refractive Surgery, Gomabai Netralaya and Research Centre, Neemuch, MP, 458441, India; 2Department of Cataract, Pediatric Ophthalmology and Strabismus, Gomabai Netralaya and Research Centre, Neemuch, MP, 458441, India
Correspondence: Bharat Gurnani, Department of Cataract, Cornea and Refractive Surgery, Gomabai Netralaya and Research Centre, Neemuch, MP, 458441, India, Tel +919080523059, Email [email protected]
Abstract: Pythium insidiosum keratitis is a vision-threatening corneal infection that often mimics fungal keratitis, yet it responds poorly to standard antifungals. Historically, approximately 80% of cases have required surgical excision of the cornea. Unlike fungi, P. insidiosum lacks ergosterol in its cell membrane and possesses a cellulose-rich wall with minimal β-1,3-glucan exposure, reducing Dectin-1–mediated detection. Surface hydroxyproline-rich glycoproteins structurally resemble host collagen, potentially engaging inhibitory lectin receptors and dampening early innate immune responses. Delayed Toll-like receptor (TLR4) activation, limited TLR2 signaling, and altered complement activation contribute to subdued inflammatory recruitment, allowing the pathogen to establish deep stromal infection before overt clinical signs emerge. We hypothesize that the organism’s aggressive behaviour stems from immune camouflage. P. insidiosum evades early host immune detection by masking its pathogen-associated molecular patterns (PAMPs) and mimicking host molecules. This allows the oomycete to establish infection with minimal initial inflammation. Supporting evidence includes the atypical, cellulose-rich cell wall (with minimal β-glucan and no ergosterol) of P. insidiosum blunts early cytokine responses. Moreover, the corneal cells infected with Pythium initially produce very low levels of IL-1β, underscoring the potential need for adjunctive immunotherapies to effectively clear the infection. If validated, this immuno-evasion hypothesis has major implications: diagnostic assays could incorporate host immune response patterns, and novel treatments might combine cell wall–degrading enzymes or Pattern Recognition Receptor (PRR) agonists with immunotherapy to “unmask” the pathogen for immune elimination. Ultimately, viewing P. insidiosum as an immuno-camouflaged pathogen offers a new paradigm to explain its clinical course and to improve outcomes in this often-devastating keratitis.
Keywords: Pythium insidiosum, keratitis, molecular mimicry, pattern recognition receptor evasion, immuno-camouflage
Background
Pythium insidiosum is an aquatic oomycete (fungus-like organism) that causes Pythiosis – an infection previously known mainly in horses and dogs, but now increasingly recognized in humans.1 In the eye, P. insidiosum causes suppurative keratitis (corneal ulceration) that closely mimics fungal keratitis in appearance, earning it the nickname “parafungus”.2 Patients often have a history of corneal trauma in water or soil environments and present with features resembling severe fungal ulcer – such as reticular infiltrates and peripheral “tentacle” extensions in the cornea.2 Critically, however, standard antifungal medications are ineffective against Pythium due to its unique cellular structure. Unlike true fungi, P. insidiosum lacks ergosterol in its cell membrane and instead incorporates cellulose in its cell wall.3 Its cell wall is composed primarily of cellulose with some β-glucans and even hydroxyproline-containing proteins,4 whereas fungal pathogens have chitin and abundant β-glucan. This fundamental difference explains why conventional antifungals (targeting ergosterol or fungal cell wall synthesis) often fail and why extreme measures, such as therapeutic keratoplasty (corneal transplantation), have been required in a high proportion of cases.5 Beyond the lack of therapeutic targets, P. insidiosum keratitis poses a diagnostic and immunological challenge. Clinically, it can be mistaken for fungal or amoebic keratitis, delaying appropriate therapy.6 Microbiologically, Pythium grows as broad, aseptate hyphae in tissue that can resemble zygomycete fungi.7 However, an astute clue is that corneal scrapings show ribbon-like hyphae that fail to stain with chitin-specific fungal stains but bind calcofluor white (which highlights cellulose). This indicates that the pathogen’s cell wall composition differs markedly from that of fungi, potentially affecting how the host’s immune system recognizes it.8
The host immune response to P. insidiosum in the cornea is not yet fully elucidated, but emerging studies show distinctive patterns. In fungal keratitis, resident macrophages and dendritic cells rapidly detect fungal cell wall components via Toll-like receptors (TLRs) and C-type lectin receptors (CLRs), triggering an intense neutrophil influx.9 For example, in Aspergillus keratitis, TLR4 and the CLR Dectin-1 (β-glucan receptor) are critical for fungal killing and neutrophil recruitment, respectively.10 By contrast, the immune response to Pythium appears initially muted. An in vivo study in a rabbit model of P.insidiosum keratitis found that key innate inflammatory cytokines (IL-1β, IL-6, IL-8) in the cornea did not surge until days 3–7 post-infection,11 suggesting a delay in robust immune activation. In in vitro experiments, human corneal epithelial cells stimulated with P. insidiosum failed to upregulate IL-1β, in stark contrast to their response to typical pathogens.12 Pythium induced IL-6 and IL-8 in these cells, indicating partial host recognition, but the absence of early IL-1β suggests that certain innate pathways (e.g., inflammasome activation) may be bypassed.13 Monocyte-derived macrophages likewise produce a distinct cytokine profile in response to Pythium: IL-1β and IL-8 are produced, partly via TLR2-dependent signalling, but neutralizing TLR2 only modestly reduced these cytokines.14 This implies that Pythium triggers multiple receptors sub-optimally. Intriguingly, patients with predisposing immunological conditions – for example, individuals with β-thalassemia – are disproportionately susceptible to systemic Pythiosis.15 Monocytes from such patients show deficient TNF-α and IFN-γ responses to Pythium zoospores, which may explain their vulnerability. All these observations point toward an organism that, unlike typical corneal pathogens, initially “flies under the radar” of the host immune system.16 Despite increasing recognition of Pythium insidiosum as a major cause of infectious keratitis, its immune evasion mechanisms remain poorly understood compared to fungal and protozoal keratitis. While fungi primarily exploit β-glucan shielding and Acanthamoeba employs cyst-mediated resistance, Pythium exhibits a unique cellulose-dominant cell wall and potential molecular mimicry, suggesting a distinct form of immune camouflage. Understanding these differences is crucial for developing targeted immunomodulatory therapies and improving diagnostic precision.
Given this background, we explore the hypothesis that P. insidiosum keratitis is severe not only because of diagnostic delays or drug resistance, but because the pathogen actively evades and subverts the host’s innate immune recognition. Furthermore, we propose an immuno-camouflage hypothesis: P. insidiosum avoids triggering full-blown immune responses by molecular mimicry of host tissues and by concealing its PAMPs from pattern recognition receptors (PRRs). We discuss how this hypothesis diverges from conventional thinking and examine supportive and contrary evidence. If Pythium has evolved to evade the immune system, unravelling this could open new avenues for diagnosis (e.g., immune-based assays) and treatment (e.g., therapies to expose or counteract the camouflage).
The Hypothesis
Pythium insidiosum keratitis represents an infection in which the pathogen’s success is due to immuno-evasion. We hypothesize that Pythium acts as an “immuno-camouflaging” oomycete. It employs molecular mimicry of host structures and strategic evasion of PRRs to dampen early innate immune detection in the cornea (Figure 1). In essence, Pythium disguises itself as “self” or at least as a non-threatening presence, buying time to invade tissue unchecked. This concept diverges from the traditional view that corneal infections cause pathology primarily by overwhelming the host response; here we propose the opposite – that Pythium causes such extensive damage because it initially underwhelms or misdirects the host response, leading to delayed containment and subsequent uncontrolled spread (Figure 2).17 The pathogenicity of Pythium insidiosum appears to result from a complex interplay between microbial persistence and host-mediated stromal damage. Unlike fungi that produce potent proteases or keratolytic toxins, Pythium exhibits a silent invasion pattern with limited enzymatic degradation yet extensive stromal infiltration. Recent evidence indicates that host-derived matrix metalloproteinases (MMP-2 and MMP-9) become upregulated in response to delayed immune recognition, leading to progressive stromal melt and perforation. This supports our hypothesis of immune camouflage, in which early evasion of pattern recognition receptors (PRRs) leads to a dysregulated secondary inflammatory cascade once the pathogen is detected. Thus, corneal damage in Pythium keratitis results not only from direct invasion but also from an exaggerated host response driven by sustained MMP activity and delayed immune activation.
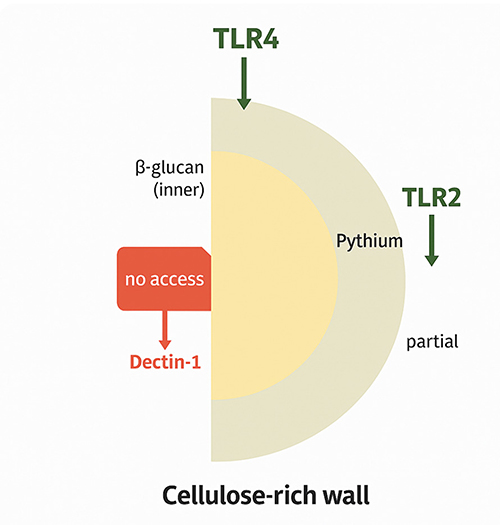 |
Figure 1 Pythium insidiosum PRR Evasion Model—The cellulose-rich outer wall limits immune recognition by masking inner β-glucan layers, preventing access to Dectin-1 and allowing only partial engagement of Toll-Like Receptor 2 (TLR2) and Toll-Like Receptor 2 (TLR4). This structural adaptation facilitates evasion of host pattern recognition receptors (PRRs) and contributes to immune evasion. |
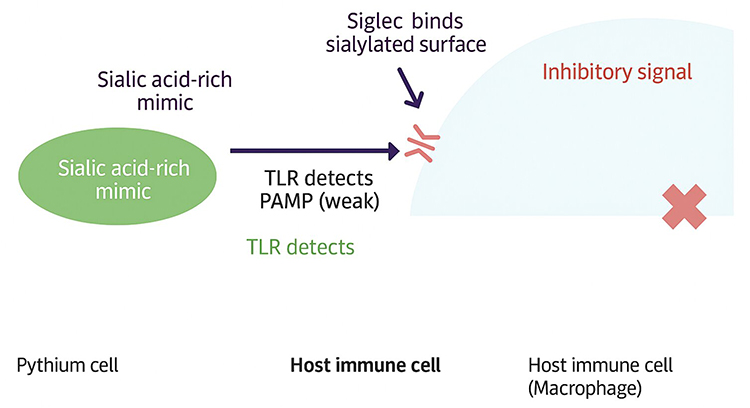 |
Figure 2 The diagram depicts molecular mimicry by Pythium through a sialic acid-rich surface that engages host Siglec receptors, delivering inhibitory signals to macrophages. This interaction weakens Toll-like receptor (TLR) recognition of pathogen-associated molecular patterns (PAMPs), allowing immune evasion. |
Key Immunological Steps in the Proposed Mechanism
Failure of Early PAMP Recognition
Upon entering corneal tissue (eg via a small wound), P. insidiosum zoospores encyst and germinate into hyphae. Unlike fungal conidia or hyphae, which carry exposed β-glucans and mannans, Pythium presents an outer cell wall rich in cellulose and possibly mucilaginous polymers. Cellulose is not a common component of human pathogens and correspondingly, humans lack dedicated PRRs for cellulose18 Important fungal PAMPs like β-1,3-glucan may be present in Pythium’s wall but are buried beneath the cellulose-rich outer layer.19 We propose that this architecture conceals β-glucan of Pythium from Dectin-1 on host phagocytes, at least in the early phase. Similarly, Pythium contains little or no chitin, so chitin-sensing pathways are not triggered. The net effect is that crucial receptors such as Dectin-1, TLR4, and NOD-like receptors that would normally signal an alarm in response to a fungal invader are either not engaged or only weakly engaged initially.20 Corneal epithelial cells and resident macrophages thus mount an attenuated response – for example, releasing some IL-8 for neutrophil recruitment, but minimal IL-1β or TNF-α (key drivers of inflammation). This mechanistic step explains the empirical finding that IL-1β is low in early Pythium infection despite the presence of a large organism in the cornea. Essentially, Pythium is like an “invisible intruder,” sneaking past the cornea’s pattern recognition system by wearing an atypical cell wall cloak.21
Molecular Mimicry of Host Molecules
In addition to concealing its microbial signatures, we hypothesize that P. insidiosum actively mimics host tissue components to mislead the immune system. Oomycete cell walls and secreted proteins may contain epitopes that resemble those found in the human cornea or extracellular matrix.22 For instance, the presence of hydroxyproline-rich proteins in Pythium’s cell wall is notable – hydroxyproline is abundant in human collagens. Pythium could produce a collagen-like glycoprotein that the host immune system perceives as “self.” Another potential mimicry target is in glycosylation patterns: Pythium might decorate its surface with sugars commonly found on human cells. One candidate is sialic acid (N-acetylneuraminic acid), a terminal sugar on many mammalian cell surface glycoproteins. Some pathogens (notably certain bacteria) coat themselves with sialic acid to engage host inhibitory receptors such as Siglecs on immune cells, thereby inhibiting immune activation.23 We propose Pythium may employ a similar strategy – by displaying host-like carbohydrates or other antigens, it engages negative regulators on macrophages and dendritic cells, further dampening their response. This molecular mimicry could induce immune tolerance or confusion: pattern recognition receptors are not triggered effectively, and any immune cells that do bind might interpret the signal as “self” and activate inhibitory pathways. The result is an inappropriate lack of acute inflammation at a time when the pathogen is actively spreading through the corneal stroma.24
Delayed and Misdirected Inflammation
By evading early detection, Pythium gains a foothold. The organism’s hyphae infiltrate extensively, often reaching the sclera or causing ring infiltrates in the cornea. Eventually, as the infection load increases and tissue damage accumulates, the immune system is inevitably alerted – necrotic cells may release damage signals or Pythium PAMPs become exposed (for instance, as hyphae age or are partially damaged, β-glucan could become accessible to Dectin-1).25 When the immune system finally recognizes the invader, it may do so explosively: neutrophils flood in (correlating with high corneal IL-8 levels by day 7) and pro-inflammatory cytokines like IL-1β and IL-17 surge in later stages.21 By this point, however, the infection is well established, and the inflammatory response, though finally robust, can cause collateral tissue damage (corneal melts, scarring). In essence, the host response is “fashionably late” – arriving so delayed that it must react with overwhelming force, which unfortunately harms the delicate cornea. This is a departure from typical fungal keratitis, in which early immune detection often reveals infection confined to a focal ulcer (with some tissue damage from inflammation), with the pathogen at least constrained early. In Pythium keratitis, immune evasion leads to a scenario where, by the time the alarm is fully raised, the infection is widespread, and the ensuing inflammation contributes more to pathology (rapid perforation, for example) than to adequate clearance 26
Immune Exhaustion or Deviation
We further speculate that Pythium’s mimicry might not only delay immunity but also skew it. If Pythium antigens mimic host proteins, the adaptive immune system may avoid targeting those antigens strongly (to prevent autoimmunity). There is precedent in infectious diseases for such mimicry, which can lead to inadequate immune responses or even tolerance. While this aspect remains speculative for Pythium, it raises the question of whether repeated exposure to Pythium (in environmental settings) could induce some immune deviation in susceptible individuals. Clinically, some patients with Pythium keratitis fail to mount high antibody titers until given immunotherapeutic vaccines, again hinting that the pathogen is not immunologically conspicuous (Figure 3).27
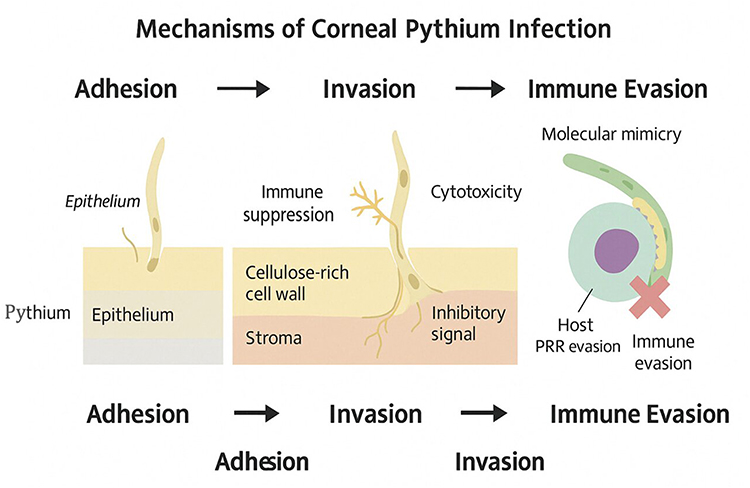 |
Figure 3 Schematic representation of the mechanisms of corneal Pythium infection, progressing through adhesion, invasion, and immune evasion. The organism adheres to the corneal epithelium, invades the stroma via a cellulose-rich cell wall, and induces immune suppression and cytotoxicity. Immune evasion is facilitated by molecular mimicry and host pattern recognition receptor (PRR) evasion, enabling persistence within the corneal tissue. |
In summary, the hypothesis posits a stepwise subversion of host defences by P. insidiosum. Initially, through structural camouflage (an atypical cell wall) and active molecular disguise, Pythium establishes infection with only a mild immune response. A delayed hyperinflammatory phase follows this silent invasion phase once the immune system finally recognizes the threat. This two-phase immunopathology – stealth then storm – could explain why Pythium keratitis often progresses rapidly to perforation and why treatments that target the organism pharmacologically have limited success if the immune system is not appropriately engaged. It diverges from existing thinking by suggesting that the host’s failure to mount an early defense, rather than the organism’s inherent virulence or the lack of drugs, is a key driver of disease severity.18
Evaluation of the Hypothesis
This is a hypothesis-driven conceptual framework intended to propose potential mechanisms and future experimental directions. The assays and models described herein, including PRR activation studies and cell wall unmasking experiments, are proposed for future validation. These experiments will be undertaken in subsequent studies. To evaluate this immuno-camouflage hypothesis, we consider evidence from the literature that supports or challenges each component of the proposed mechanism, and we suggest experimental approaches to test these ideas.
Supporting Evidence for Immune Evasion
Unique Cell Wall and PRR Evasion
There is strong biochemical and experimental evidence that P. insidiosum is not recognized by the host in the same way as fungal pathogens. The cell wall composition (summarized in Table 1) provides the first clue. Unlike Candida or Aspergillus, Pythium incorporates cellulose and only a small amount of β-glucan, and it lacks chitin.28 This means key fungal PAMPs are either absent or shielded. Indeed, studies have shown that TLR4 and Dectin-1, which are crucial for immune detection of fungi (TLR4 drives fungal killing, Dectin-1 triggers neutrophil recruitment), are not robustly activated at the onset of Pythium infection.29 Ahirwar et al found that in vitro exposure of human corneal cells to P. insidiosum led to upregulation of Dectin-1 and TLR4 transcripts only after 12 hours, implying that initial contact (0–6 hours) did not strongly engage these receptors.11 In contrast, fungal components often trigger Dectin-1 within minutes, leading to rapid IL-1β release. The delayed engagement of Dectin-1/TLR4 in Pythium aligns with our hypothesis that early during infection these PRRs “see” little to react to. Additionally, Pythium does stimulate TLR2 to some extent (likely via lipoproteins or glycoproteins on its surface),11 but TLR2 activation tends to induce a more regulatory or IL-10–skewed response in many contexts, and in corneal macrophages TLR2 alone was insufficient to drive IL-6 production.11 Thus, the pattern of PRR usage by Pythium (more TLR2, delayed Dectin-1/TLR4) is consistent with a pathogen that avoids triggering the host’s most potent antifungal sensors initially. This supportive evidence comes from both the organism’s biochemistry and host gene expression data.30
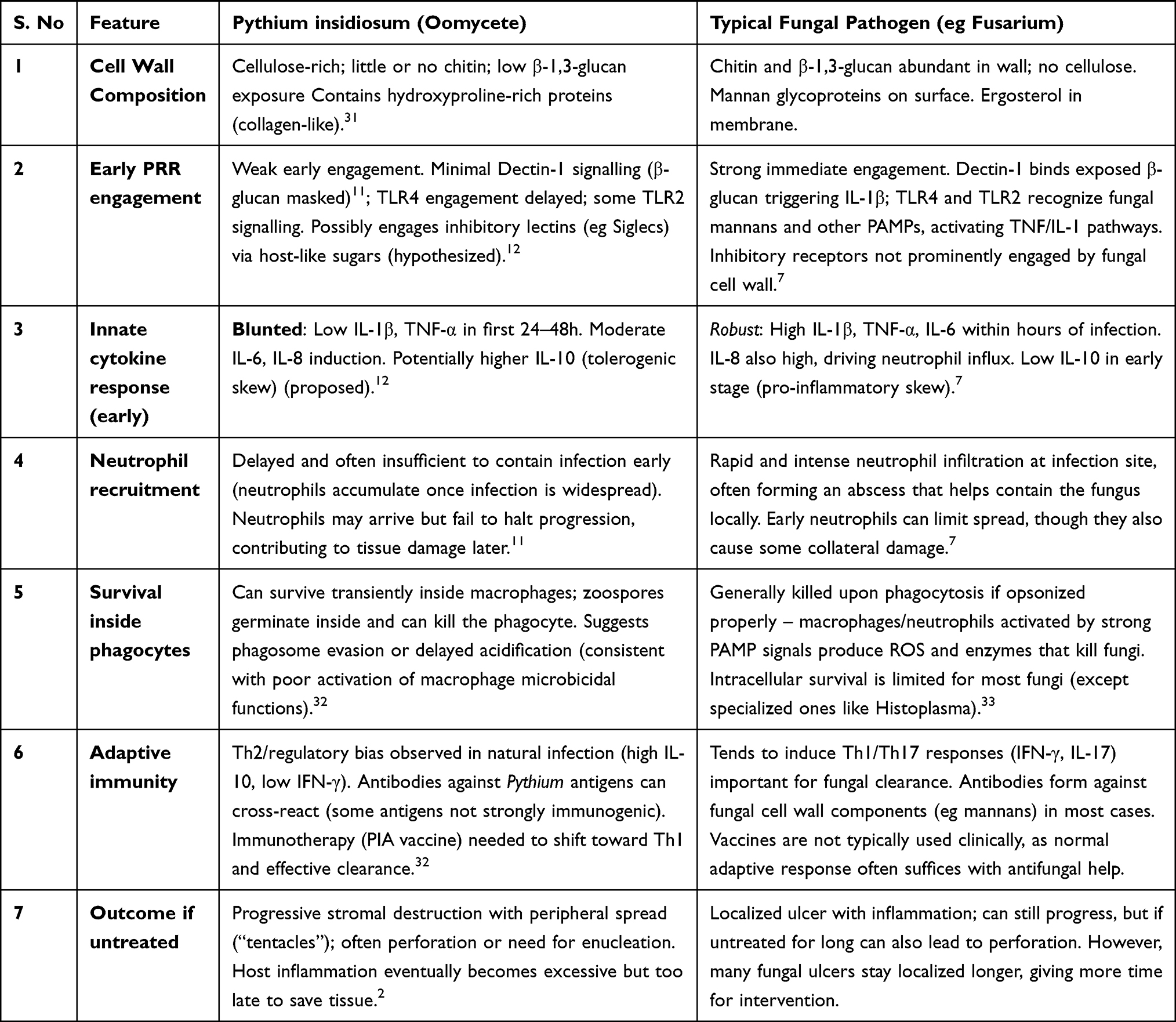 |
Table 1 Comparative Analysis of Immune Evasion Strategies of Pythium insidiosum versus Typical Fungal Keratitis Pathogens, Highlighting Differences in Cell Wall Composition, Pattern Recognition Receptor Engagement, Cytokine Responses, Neutrophil Recruitment, Intracellular Survival, Adaptive Immunity Bias, and Disease Outcomes |
Damped Early Cytokine Response
Clinical reports and animal studies corroborate that Pythium fails to elicit a fulminant early immune response. In a rabbit model, the expression of pro-inflammatory cytokines was modest at 3 days post-infection and peaked at 7 days despite significant infection being present. Specifically, IL-1β – a master pro-inflammatory cytokine usually elevated early in infections – was not significantly up on day 3 compared to control, whereas by day 7 it rose substantially.34 This timing suggests that the initial immune activation was delayed. For comparison, in fungal keratitis IL-1β can rise within hours as Dectin-1 and TLRs trigger inflammasome pathways.35 The absence of early IL-1β in Pythium infection (despite the organism proliferating in the cornea) strongly supports the idea of early immune evasion. The pattern of chemokines also fits: IL-8 was elevated in Pythium infection, but mostly at later time points, mirroring the massive neutrophil influx observed clinically only once the infection is well advanced (often giving the appearance of a “dense infiltrate” days into the disease). Thus, both the in vitro epithelial data and in vivo cytokine profiles indicate a muted early innate response, as predicted by the hypothesis.36
Molecular Mimicry and Immune Modulation
Direct evidence of molecular mimicry is more difficult to obtain, but there are tantalizing clues. One clue comes from comparative immunology: Pythium and a phylogenetically distant fungus, Basidiobolus, share cross-reactive antigens that can confuse diagnostic tests37 If Pythium shares antigens with Basidiobolus, it is conceivable it also shares epitopes with mammalian proteins (given that convergent mimicry is possible). More directly, the composition of Pythium’s extracellular matrix includes hydroxyproline, an amino acid prevalent in collagen and plant cell wall glycoproteins but rare in most pathogens.38 This unusual feature suggests cell wall of Pythium proteins might resemble host structural proteins (like collagen or mucins), potentially reducing their immunogenicity. Additionally, studies of Pythium immunotherapy give hints about baseline immune skewing. Untreated patients with vascular Pythiosis often show a Th2-biased immune profile (high IL-5, IL-10, low IFN-γ), whereas after receiving the Pythium antigen vaccine, they shift toward a Th1 profile (high IFN-γ, IL-12) and improved macrophage activity.39 This implies that Pythium infection in its natural course might promote a non-protective Th2/regulatory environment – possibly via IL-10 induction or lack of strong Th1 cues – which would be consistent with immune evasion. For example, if Pythium’s surface mimicry engages macrophage Siglec receptors, it could drive an anti-inflammatory, IL-10^high phenotype in those cells, echoing what is seen in certain chronic parasitic infections that exploit similar mechanisms.40 While specific Siglec-Pythium interactions have not been documented, the concept is supported by analogies in microbial pathogenesis (several pathogens coat themselves in host-like sialic acid to evade complement and neutrophils). Our hypothesis is that Pythium similarly decorates itself in such a way to appear as “self” to the innate immune system, and this is indirectly supported by the need to artificially boost Th1 immunity via a vaccine to clear the organism in disseminated cases (Figure 4).41
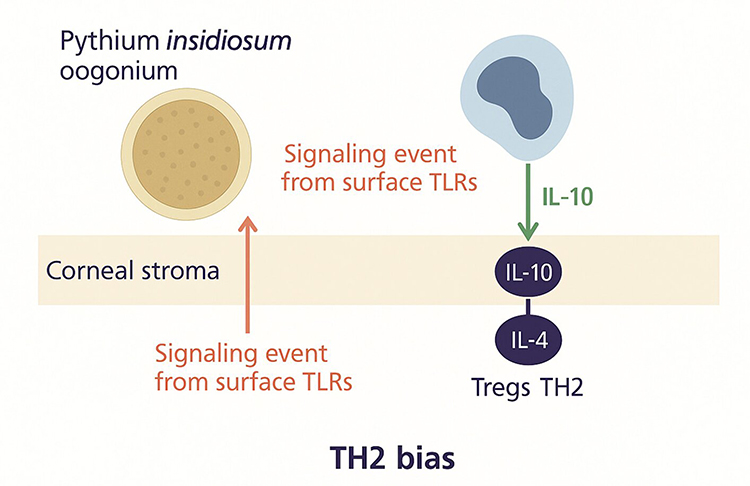 |
Figure 4 Illustration of stromal immune deviation in Pythium insidiosum infection, where signaling events from surface Toll-like receptors (TLRs) in the corneal stroma promote a T-Helper cells-2 (TH2)-biased immune response. The process involves Interleukin-10 production, leading to regulatory T cell (Treg) and TH2 cell activation with increased Interleukin-4, facilitating immune modulation and persistence of infection. |
Survival Inside Phagocytes
A hallmark of many immune-evasive pathogens is the ability to survive phagocytosis (eg Mycobacterium tuberculosis in macrophages). There is evidence that Pythium zoospores can survive after being engulfed by phagocytes.42 Medhasi et al (2024) observed that even when patient-derived macrophages phagocytosed Pythium zoospores, the zoospores were able to germinate inside the macrophages and ultimately kill the host cells.32 This implies that not only does Pythium initially avoid activation of the macrophage’s microbicidal arsenal, but it can also resist or delay destruction within the phagosome. One reason could be that the usual phagolysosomal responses (eg production of reactive oxygen species or acidification) are not effectively triggered without strong PRR signals. If the macrophage does not “realize” it has engulfed a dangerous pathogen (due to mimicry), it might not activate the full spectrum of killing mechanisms, allowing Pythium to persist intracellularly temporarily.43 While Pythium is primarily extracellular in tissues, this intracellular phase could help it disseminate or at least survive initial contact with immune cells. The ability to germinate inside a macrophage and kill it is direct evidence of a failure of innate immunity – something one would not expect if the macrophage were properly activated. This observation strongly supports the notion that Pythium can disarm or evade cellular immunity at least for a window of time (Figure 5).44
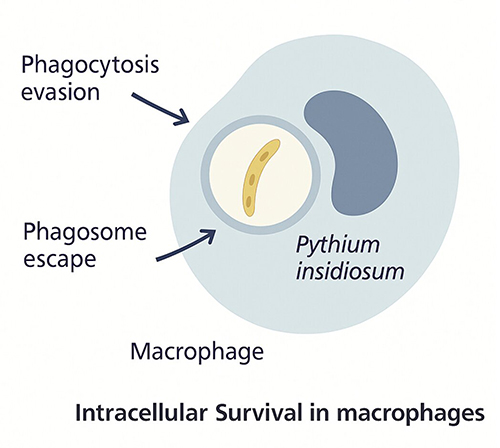 |
Figure 5 Diagram illustrating intracellular survival of Pythium insidiosum within macrophages. The organism evades phagocytosis and escapes from the phagosome, enabling persistence inside host immune cells and contributing to immune evasion. |
Clinical Correlations
Certain clinical features of Pythium keratitis make more sense under the immuno-camouflage hypothesis. One example is the frequent necessity of enucleation or evisceration in advanced cases despite intense inflammatory response in the eye. By the time the immune system fully responds, the infection has often involved most of the cornea and even beyond, making immune-mediated clearance without destroying the eye nearly impossible.45 Another correlation is the risk factors: patients with iron overload or thalassemia have impaired neutrophil and macrophage function and are at higher risk of fulminant Pythiosis. If Pythium relies on slipping past a suboptimal immune system, then those with any baseline immune defect (even subtle, like iron-overloaded phagocytes) would be at a marked disadvantage, which matches clinical observations.46 In fungal keratitis, by contrast, otherwise healthy individuals mount a vigorous response and usually wall off the infection unless pathogen load is very high or virulence is extreme. The fact that Pythium often requires such drastic intervention suggests that, in essence, the immune system “drops the ball” early on – aligning with our hypothesis.16
Contradictory Evidence or Alternate Explanations
While the above points support the hypothesis, it is important to acknowledge evidence that could challenge it.
Intensity of Inflammation
One might argue that Pythium keratitis is actually characterized by too much inflammation rather than too little – corneas can undergo rapid melt with extensive neutrophil infiltration (sometimes described as a “sterile” immune ring). Could this not indicate that the immune system is responding strongly to Pythium, rather than being fooled by it? Indeed, by mid to late infection, the host response is extreme.47 However, our hypothesis does not claim Pythium lacks immunogenicity entirely; rather, it suggests an initial delay or blunting. The later excessive inflammation is not contradictory – it’s expected once the immune system catches up. Another interpretation of the excessive inflammation is that it results from the immune system suddenly recognizing the pathogen after a period of unopposed growth, leading to an over-reactive response (analogous to an immune reconstitution phenomenon). Thus, the presence of inflammation in Pythium keratitis is not evidence against immune evasion; it may actually be a consequence of it. Nevertheless, it will be important to quantify the timing of immune events carefully in experimental models to confirm this sequence.
No Known Pythium Immune Effector Molecules
In plant-pathogenic oomycetes (like Phytophthora), numerous secreted effectors help the pathogen evade plant immunity. For P. insidiosum, such virulence factors are less studied. We currently lack identified Pythium proteins that actively disable human immune signaling. The hypothesis of molecular mimicry is somewhat speculative in this regard – we infer its presence from outcomes, not from having isolated a “stealth protein.” This is a gap that future research must address.22 It is possible that Pythium, being an evolutionary generalist (infecting plants, animals, humans), has not evolved highly specific immune suppressors for humans, and that its immune evasion is more due to passive lack of PAMPs than active immune suppression. In evaluating the hypothesis, one should consider the simpler explanation: Pythium might evade detection simply because our immune system is not adapted to recognize oomycete features (an evolutionary blind spot), rather than Pythium specifically mimicking us. This is a valid alternative interpretation.1 The hypothesis would still hold in spirit (immune evasion), but the emphasis would shift from active mimicry to passive “stealth” by unusual biochemistry. Testing this will require deeper analysis of Pythium’s genome and secretome for known immune-modulating motifs (eg does it have proteins with host motif domains, or known inhibitor domains?).
Adaptive Immunity Eventually Clears Infection
Another point to consider is that many patients do eventually seroconvert and can clear Pythium with appropriate treatment (including immunotherapy). If Pythium were completely camouflaged, one might expect it to cause chronic infection indefinitely (like some parasites that establish latency). Instead, in many cases of keratitis that undergo therapeutic keratoplasty combined with aggressive medical therapy, the patient recovers without systemic spread, indicating the immune system can recognize and eliminate residual Pythium.48 This suggests that any immune evasion by Pythium is temporary or partial. Our hypothesis accounts for this by the idea that once enough damage occurs or enough antigens are present, the jig is up – Pythium gets recognized (albeit late). So this observation does not refute the hypothesis, but it emphasizes that Pythium’s camouflage is not perfect or permanent. It buys time, but not infinite time. Experimentally, this could be seen in vitro: if one exposes immune cells to increasing doses of Pythium antigen or longer exposure, eventually typical inflammatory responses might appear. This dose-response could be explicitly tested.
Experimental Approaches to Test the Hypothesis
Although this is a hypothesis-driven conceptual study, the proposed experimental framework is designed to be practically implementable Feasibility testing could involve in vitro macrophage stimulation assays with Pythium antigens to assess PRR activation, alongside comparative β-glucan unmasking assays. Validation in ex vivo or animal models could subsequently evaluate cytokine responses (IL-1β, TNF-α, IL-6) and confirm immune activation patterns. These approaches would provide empirical support for the proposed immune camouflage model of P. insidiosum. To move from circumstantial evidence to direct proof of immuno-camouflage, the following studies are proposed:
Comparative PRR Activation Assay
Take P. insidiosum hyphae and a control fungus (eg Aspergillus or Fusarium hyphae) and expose them to human or mouse dendritic cells or macrophages in vitro. Measure early signaling events (NF-κB activation, inflammasome activation) and cytokine release (IL-1β, TNF-α, IL-6, IL-10) at very short time points (eg 4, 8, 12 hours) (Figure 6). We predict Pythium will induce significantly lower IL-1β and TNF-α than the fungus, but possibly similar IL-8 (since neutrophils do eventually come) and perhaps higher IL-10 (Table 2).24 Furthermore, using blocking antibodies or knockout cells for specific PRRs can pinpoint differences: for instance, blocking Dectin-1 should have little effect on Pythium-stimulated cytokines (since Pythium is not effectively using Dectin-1 pathways initially), whereas it would drop cytokines in Fusarium-stimulated cells (where β-glucan–Dectin-1 is key). Early results of this kind of experiment would directly confirm if Pythium fails to trigger the usual PRR signals (Figure 7).49
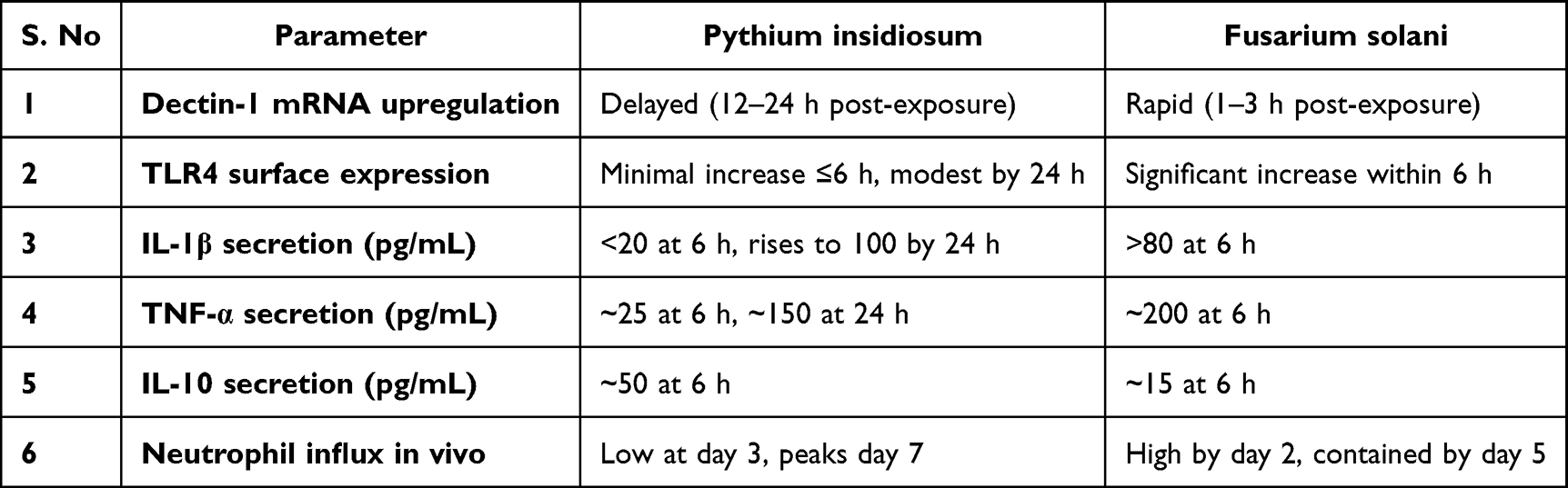 |
Table 2 Proposed Omparative Kinetics of PRR Activation and Cytokine Responses in Pythium insidiosum versus Fusarium solani Keratitis, Highlighting Delayed Innate Immune Engagement and Prolonged Inflammatory Profile in Pythium Infections |
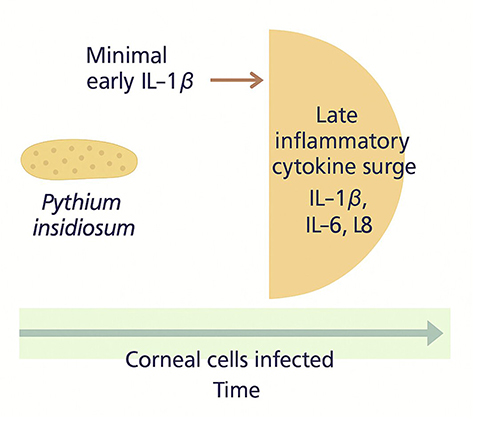 |
Figure 6 Delayed hyperinflammation model of Pythium insidiosum infection, showing minimal production followed by a late surge of inflammatory cytokines, including Interleukin −1β, Interleukin −6, and Interleukin −8, during ongoing corneal cell infection. This delayed response contributes to tissue damage and disease progression. |
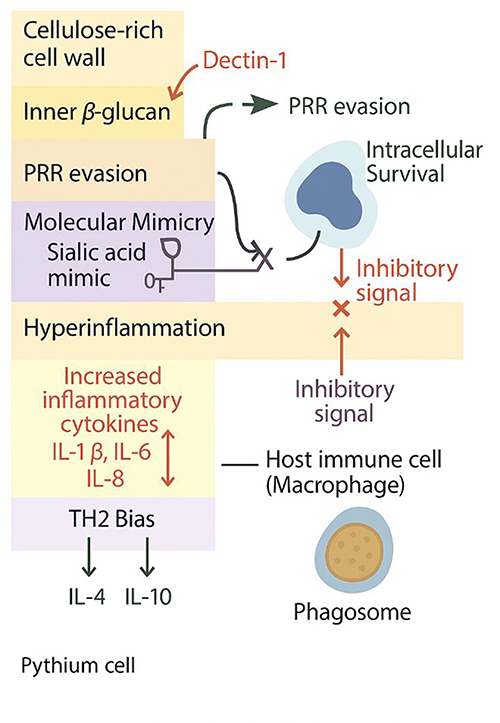 |
Figure 7 Schematic illustration of the mechanisms of Pythium insidiosum immune evasion. Strategies include pathogen recognition receptor (PRR) evasion via a cellulose-rich cell wall and β-glucan masking, molecular mimicry through sialic acid-rich surfaces, and intracellular survival within macrophages with inhibitory signaling. Additional mechanisms involve inducing hyperinflammation with elevated IL-1β, IL-6, and IL-8, and skewing the immune response toward a TH2 bias characterized by increased IL-4 and IL-10 production. |
Cell Wall “Unmasking” Experiment
To test if cellulose is hiding immunostimulatory β-glucan in Pythium, we can enzymatically treat P. insidiosum hyphae with cellulase (which digests cellulose) or with a β-glucanase control. Treated and untreated hyphae can then be added to immune cells or even injected into a mouse cornea. We hypothesize that cellulase-treated Pythium (with cellulose removed, exposing inner β-glucan) will elicit a much stronger immune response – eg immediate neutrophil infiltration, high IL-1β – compared to intact Pythium.50 Preliminary support for this idea comes from a recent proposal to use cellulose synthesis inhibitors in treating Pythium keratitis; while that was aimed at directly weakening the organism, an added benefit might be enhancing immune recognition. If our hypothesis is correct, cellulase exposure will “uncloak” Pythium, making it behave more like a fungus immunologically. This can be quantified by an increase in Dectin-1–dependent cytokines after cellulase treatment. A converse experiment is to coat a true fungus (like Candida) with exogenous cellulose or a synthetic polymer and see if its recognition by immune cells is reduced – this would mimic Pythium’s strategy and strengthen the concept of physical PAMP masking.2
Mimicry Detection Assays
To directly detect molecular mimicry, we could perform a proteomic and glycomic analysis of Pythium cell surfaces. High-performance liquid chromatography or mass spectrometry can identify if cell wall of Pythium contains sialic acids, glycosaminoglycans, or other host-like carbohydrates. We can also test if Pythium binds to host immune inhibitory receptors: for instance, a fluorescence binding assay with recombinant human Siglec proteins could show whether Pythium extracts have affinity for these receptors. If Pythium indeed carries sialylated epitopes, one would see binding to Siglec-7 or Siglec-9 (common on monocytes and neutrophils).51 We can also treat Pythium with neuraminidase (which removes sialic acids) and see if immune recognition improves (similar logic to the cellulase experiment). On the protein side, immunoblotting could be done to see if antibodies against human corneal proteins cross-react with Pythium proteins. For example, one might test whether anti-collagen antibodies bind any Pythium antigens on a blot – unusual binding could indicate a collagen-like protein in Pythium. Additionally, any Pythium gene encoding a protein with significant sequence homology to a human protein (detected via BLAST searches) would be a candidate mimic; knocking it out (if feasible) and seeing if immune detection of the mutant improves would be a definitive test. These experiments are more challenging but would directly demonstrate mimicry.52
In vivo Immune Challenge Models
An elegant in vivo experiment would be a comparative infection in PRR-deficient vs normal mice. For example, use P. insidiosum to infect the corneas of wild-type mice and Dectin-1 knockout mice (or MyD88 knockout mice, etc). If our hypothesis is correct that Pythium largely evades Dectin-1, then Pythium keratitis outcomes might be similar in Dectin-1 KO and wild-type mice (ie knocking out that sensor does not make things drastically worse, because it was not contributing much to defense anyway). In contrast, a fungal keratitis in Dectin-1 KO mice would be much more severe than in wild-type.49 Such a result – Pythium causing severe disease irrespective of certain PRRs – would bolster the idea that it does not trip the normal alarms. Conversely, if knocking out an inhibitory pathway (for instance, mice lacking Siglec-E, the murine equivalent of human Siglec-9) leads to Pythium being better controlled (because the inhibitory signal is gone), that would strongly support the mimicry aspect. Measuring bacterial vs Pythium load and host damage in these models can pinpoint which immune pathways are bypassed by Pythium. Additionally, adoptive transfer experiments (eg transferring macrophages educated in the presence of Pythium antigens into naive hosts) could show whether Pythium induces a regulatory phenotype that dampens clearance.53
Testing Molecular Mimicry
A direct way to evaluate mimicry is to see if disrupting the suspected mimic confers a host advantage. If we suspect, for instance, that Pythium’s sialylation of its surface is critical, we could create conditions to grow Pythium in presence of sialic acid analog inhibitors (to reduce any sialylation) and then test its virulence or immune recognition. Alternatively, use a Siglec-Fc fusion protein as a decoy in an infection model – if adding a soluble Siglec that binds Pythium’s surface leads to greater immune activation (by blocking the pathogen’s engagement of inhibitory receptors on actual immune cells), that would strongly validate the mimicry concept.54 These are sophisticated experiments but feasible with modern immunology techniques. In evaluating contradictory data, we must consider that severe inflammation in Pythium keratitis is not evidence against immune evasion but a likely consequence of initial evasion. However, to be thorough, the hypothesis should also be tested in scenarios where immune evasion might not be at play. For example, if we introduce Pythium antigens alone (without live organism) into a cornea, do they provoke less response than equivalent fungal antigens? If yes, the antigens themselves are inherently less immunostimulatory (supporting evasion). If no, and the live infection is needed to see the dampening (meaning only live Pythium causes low response), it could mean Pythium actively secretes an immunosuppressive factor during infection. This distinction – passive evasion via “stealth” PAMPs vs active immunosuppression – is subtle but important, and experiments can be designed to differentiate (eg co-culture Pythium with another pathogen to see if it suppresses the response to the second pathogen).55
Consequences and Discussion
If P. insidiosum is indeed an immuno-camouflaging pathogen, the implications for clinical management and our understanding of host-pathogen interactions are profound. While the Th2-skewed immune response and absence of ergosterol in Pythium insidiosum have been recognized as major contributors to its persistence and antifungal resistance, these mechanisms do not fully explain its aggressive corneal behaviour. The present hypothesis introduces an ocular-specific model of immune camouflage, integrating both pathogen and host microenvironmental factors. The corneal surface represents a unique immunological niche characterized by relative immune privilege, avascularity, and localized innate defense through pattern-recognition receptors (PRRs). We propose that P. insidiosum adapts to this setting by masking β-glucan with a cellulose-rich outer wall, thereby limiting Dectin-1–mediated recognition, and by expressing sialic acid–rich surface components that engage Siglec receptors on host immune cells to deliver inhibitory “self-like” signals. This dual strategy of PRR evasion and molecular mimicry blunts early corneal immune activation, explaining the delayed inflammatory onset and rapid stromal infiltration observed clinically. By reframing P. insidiosum infection as an active immune-camouflage process rather than passive immune evasion, this model provides a new perspective on ocular pathogenesis and suggests potential therapeutic avenues targeting PRR activation or Siglec blockade to restore effective immune recognition in Pythium keratitis. This section discusses the potential consequences, spanning diagnostics, treatment, and broader immunological insight:
Rethinking Diagnostics
Current diagnostic methods for Pythium keratitis rely on microbiological culture, PCR, or cytology (eg seeing aseptate hyphae on smear). These can be slow or inconclusive, leading to delays in proper therapy. If the immuno-camouflage hypothesis holds true, it suggests a novel diagnostic approach: detecting the host’s lack of immune response. For instance, in a patient with a severe corneal ulcer, an unusually low level of IL-1β or TNF-α in the tear fluid or aqueous humor (relative to the size of the ulcer) could raise suspicion for Pythium.56 In contrast, fungal ulcers typically induce a marked inflammatory cytokine milieu. Thus, measuring a panel of inflammatory mediators from tear samples might serve as an “immune fingerprint” – Pythium could show an IL-10^high^/IL-1^low^ signature early on, tipping off clinicians that this is not a typical fungus. Additionally, the presence of host-like components in the lesion (eg detecting cellulose in corneal scrapes with fluorescent brighteners, already used, or even finding human collagen fragments bound to Pythium – hypothetically if Pythium cloaks itself in host debris) could be exploited as indirect markers.57 A fascinating possibility is the development of a skin test or blood test akin to a delayed-type hypersensitivity test: inoculate a small amount of killed Pythium antigen intradermally and observe if the patient mounts a normal Th1 response. A patient with Pythium infection might exhibit anergy or a skewed response due to immune deviation, whereas they would respond normally to common antigens. Such immunological tests could complement molecular diagnostics.58
Therapeutic Innovations
Recognizing immune evasion opens up entirely new therapeutic strategies beyond simply trying to kill the microbe. One strategy is immunomodulation – essentially, boosting the host’s ability to recognize and fight Pythium. This could be as straightforward as adding an immunostimulatory adjuvant to the treatment regimen.59 For example, topical TLR agonists (like a TLR7/8 agonist imiquimod, or a CpG ODN for TLR9) might be applied to the cornea to activate innate immunity broadly, counteracting Pythium’s suppression. There is precedence for using imiquimod in difficult fungal or protist infections of the skin; a careful use in the cornea could potentially ramp up local immunity (though with risk of inflammation).60 Another angle is checkpoint inhibition in the immune sense: if Pythium engages inhibitory receptors like Siglecs or PD-1/PD-L1 pathways on macrophages (the latter is speculative), one could use blocking antibodies to those (akin to cancer immunotherapy but locally) to prevent the pathogen from exploiting those “brakes”. Of course, safety in the eye is a concern, but short-term local application might be feasible.61 Perhaps, the most immediately practical idea is to combine Pythium-targeted therapy with agents that strip its camouflage. The recently proposed use of cellulose biosynthesis inhibitors (CBIs) falls in this category. Indaziflam or similar agents could weaken the oomycete’s cell wall and simultaneously expose its underbelly to the immune system. Our hypothesis provides a second rationale for CBIs: beyond fungistatic effects, they might turn Pythium from an invisible foe to a visible one. Likewise, enzymatic adjunctive therapy could be tried – for instance, hourly topical cellulase or chitinase (though Pythium has little chitin, cellulase is relevant) might digest the pathogen’s protective layer in vivo. This concept parallels how biofilm-degrading enzymes are used to break down bacterial biofilms to improve antibiotic penetration and immune access.2
Another therapy inspired by mimicry would be to target the mimicry itself. If we confirm Pythium uses host-like sialic acids, we could use neuraminidase eye drops to remove those sugars. Neuraminidase (sialidase) eye drops are not standard, but interestingly, certain bacteria produce sialidases that could potentially be harnessed or formulated if needed. Alternatively, a competitive inhibitor that saturates Siglec receptors (preventing Pythium from engaging them) might be applied – though this is quite experimental.62 Immunotherapy (as already practiced in some settings for Pythium, where patients receive injections of Pythium antigen to stimulate immunity) would also benefit from understanding of evasion. For instance, knowing that a Th1 response is protective, clinicians could monitor cytokine profiles after immunotherapy doses and ensure patients are mounting the right type of response. In the future, a more refined Pythium vaccine could be developed that not only exposes the immune system to Pythium antigens but also includes an adjuvant that blocks the organism’s inhibitory tricks (perhaps a conjugate that ties up inhibitory receptors) (Table 3).63
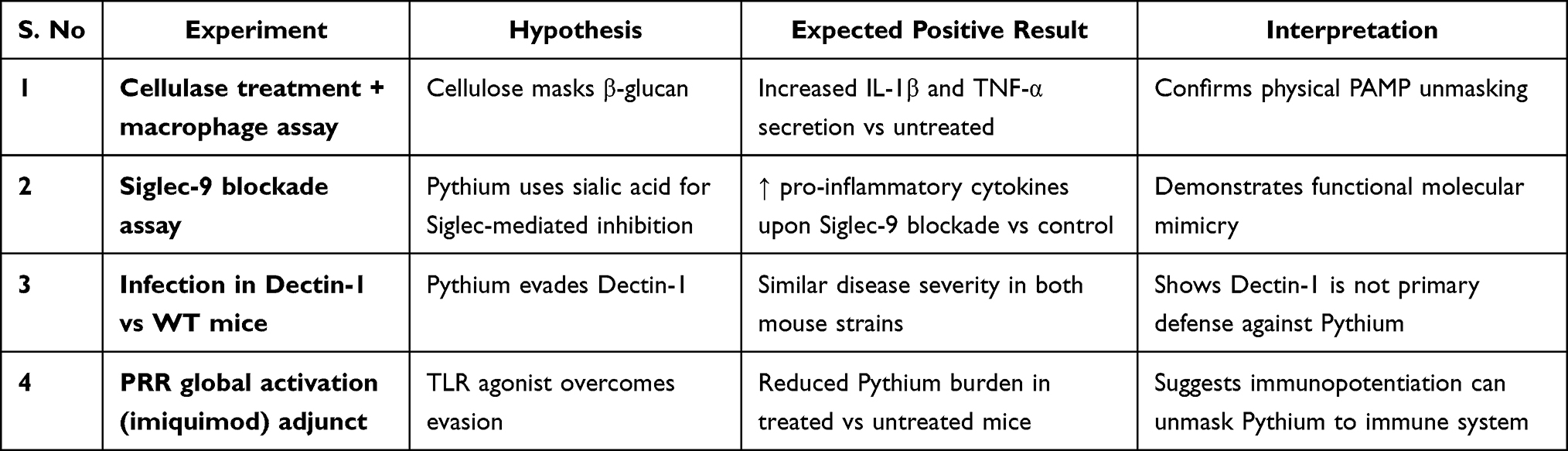 |
Table 3 Proposed Experimental Approaches to Investigate Pythium insidiosum Immune Evasion via Cellulose Masking, Siglec-Mediated Inhibition, and PRR Circumvention, with Anticipated Outcomes to Validate These Mechanisms |
Rationale for Combination Therapy
If immune evasion is key, then successful treatment likely requires both killing the organism and engaging the immune system. This might explain why combinations of antibiotics (like azithromycin or linezolid) with surgery have had partial success, some antibiotics like azithromycin have immunomodulatory properties (azithromycin is known to dampen neutrophil responses in other contexts, but perhaps its inhibition of protein synthesis in Pythium also stresses it and exposes PAMPs).64 The hypothesis encourages combination therapy where one component targets the pathogen and another “enables” the immune system. For example, a combination of linezolid (to inhibit Pythium’s protein synthesis) + interferon-gamma eye drops (to activate macrophages) could be envisioned. While speculative, if Pythium often infects individuals with suboptimal immune function (like those with thalassemia or even local immune deviation in the eye), providing immune stimulants externally could tip the balance.65
Rethinking Pathogenesis in Other Contexts
Validating immune camouflage in Pythium might prompt a re-examination of other “atypical” infections. For instance, Acanthamoeba keratitis is another difficult corneal infection, caused by a protozoan that also has a cyst form with cellulose in the wall – does it similarly evade immune detection? Some evidence in animal models shows Acanthamoeba cysts induce less inflammation, and like Pythium, it often requires vigorous immune activation (eg via miltefosine or PHMB that might also act on host immune cells). The parallels suggest that immune evasion might be a broader theme among organisms that are not classical bacteria or fungi. Studying Pythium could thus shed light on a spectrum of “stealth” pathogens.
Rethinking Host Damage
The hypothesis also reframes the cause of tissue damage in Pythium keratitis. Rather than simply attributing it to the organism’s invasiveness or toxins, it suggests a two-phase damage model: early damage is due to the organism digesting tissue unchecked (since immune cells are not stopping it), and late damage is due to an overzealous immune response (once the immune cells finally engage, they release a storm of enzymes and radicals that harm tissue).4 This aligns with the observation that Pythium can cause a cornea to melt rapidly – likely a combination of proteolytic enzymes from the pathogen and matrix metalloproteinases from neutrophils once they arrive. Therefore, therapies might need to protect the cornea in two ways: by bringing in immune help earlier, and by controlling excessive inflammation later. For the latter, adding collagen cross-linking or anti-proteinase therapy could be beneficial. Indeed, one could speculate that early immune evasion followed by late inflammation means an ideal therapy might oddly include both an immune stimulant (early) and an anti-inflammatory (later) in a timed sequence.66 This is complicated to implement, but worth investigating (eg in animal models, give an immune booster at inoculation and a steroid 4 days later once treatment has started, to see if outcome improves). Traditionally steroids are contraindicated in infectious keratitis, but in Pythium, if the immune response is tardy and then destructive, a carefully timed anti-inflammatory might preserve vision once adequate antimicrobial action is in place. This is conjectural but follows logically from the hypothesis.
Rationale for Early Surgical Intervention
Currently, early therapeutic keratoplasty is often done in Pythium keratitis because medical therapy is unreliable. Our hypothesis lends immunological justification to this practice: by removing the bulk of the camouflaged pathogen, surgery essentially removes the source of immune evasion. The wound from surgery will trigger a classic wound-healing immune response, which can then clean up any remaining Pythium (especially if combined with immunotherapy).67 Surgeons have noted that even after therapeutic keratoplasty, recurrence at the graft margin can happen, presumably from residual organism – one might interpret that as Pythium trying to reinvade, but now in the presence of a lot of post-surgical inflammation it sometimes fails to take hold. Ensuring the immune system is activated (perhaps by debriding edges, etc.) is likely one reason that large excisions with a good rim of clear tissue have better outcomes. In this context, surgery is an immune-restoring measure (in addition to pathogen debulking). If immune-based therapies improve, perhaps surgery could be less extensive or avoided. Still, until then, it remains a mainstay partly because it sidesteps the need for the immune system to do the initial heavy lifting.31
Rethinking Epidemiology and Prevention
Emerging evidence indicates that P.insidiosum keratitis is not limited to a single geographic region but has a truly global footprint. Although initially described in Thailand, subsequent case reports and case series have detailed infections in India, Australia, China, Israel, and the USA.3 A recent systematic review noted that Pythiosis—including its ocular form—has been documented in more than 23 tropical, subtropical and temperate countries, with mean incidence increasing over the past decade.4 In South India, for example, a prevalence of approximately 5.9% of microbial keratitis cases was attributed to P. insidiosum in one series.2 These data emphasise that P. insidiosum should be considered a globally relevant ocular pathogen, particularly in water-related and agrarian exposures, and highlight the need for heightened awareness, region-adapted diagnostic pathways, and surveillance systems in both endemic and non-endemic regions. If Pythium mainly causes disease in those whose immune system does not react properly, identifying those at risk (like individuals with subtle immunosuppression or genetic predispositions in PRRs) could be important. Perhaps in the future, genetic screening or immune profiling could identify people who might need prophylactic measures when in endemic areas (for example, avid swimmers with diabetes or iron overload could avoid stagnant water exposure). Also, understanding that Pythium is an environmental organism that our immune system is not “trained” to recognize might encourage development of prophylactic measures for high-risk groups, maybe even an experimental vaccine for veterinarians or others who frequently encounter it. In the broader sense, confirming immune camouflage in Pythium would underscore the importance of looking at host-pathogen interactions in infections that are atypical. It would remind clinicians that a pathogen’s “virulence” is often a two-way street – sometimes the most dangerous pathogens are those that do not immediately send our immune system into battle mode. This has analogies in other fields (for instance, certain cancers escape immune detection in similar ways; chronic infections like hepatitis B have phases of immune tolerance). Pythium keratitis could become a model for studying immune tolerance in acute infection (Table 4).
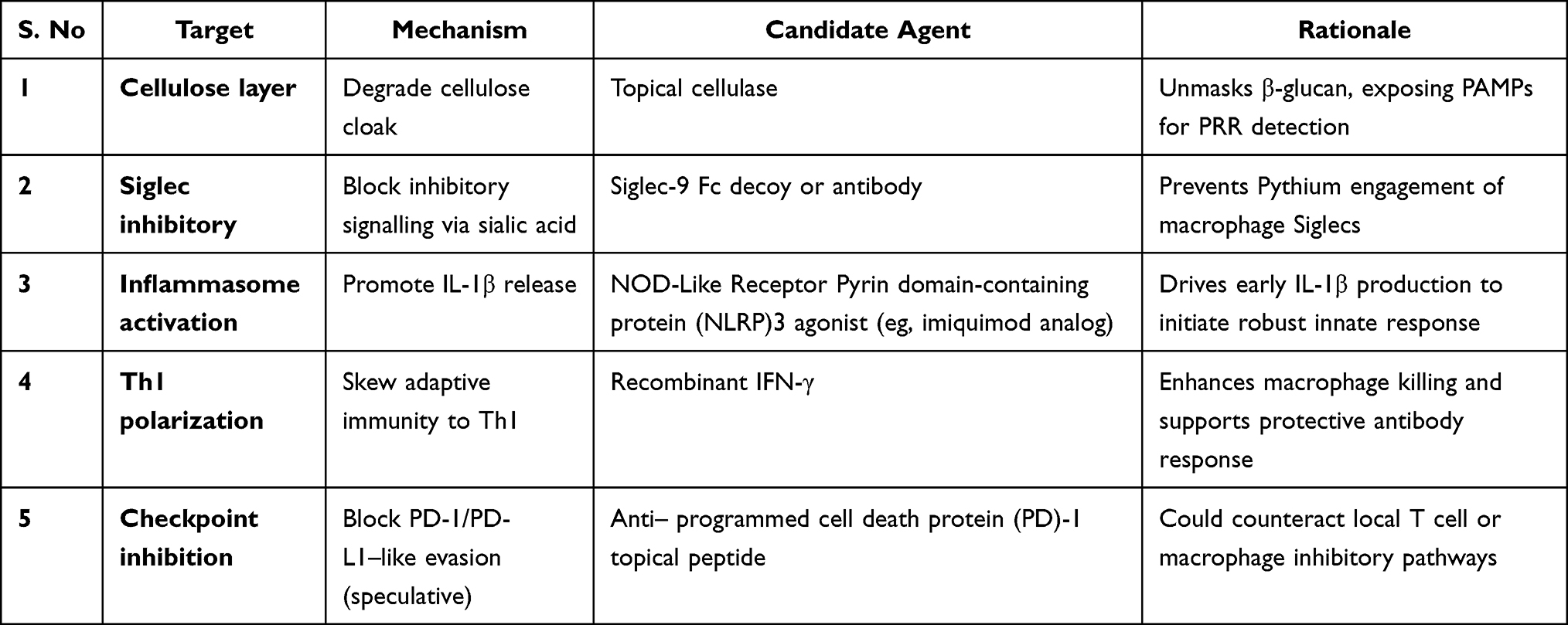 |
Table 4 Potential Immunotherapeutic Targets and Candidate Agents for Pythium insidiosum, Highlighting Mechanisms to Unmask Pathogen Antigens, Boost Innate and Adaptive Immunity, and Counteract Immune Evasion |
While the hypothesis integrates existing evidence on immune evasion mechanisms, several uncertainties remain regarding the specific molecular interactions between P. insidiosum and host immune receptors. Future research should focus on validating these proposed pathways through in vitro PRR-binding studies, cytokine response assays, and in vivo infection models. Addressing these questions will be essential to confirm whether Pythium employs immune camouflage as a dominant survival strategy and to identify potential therapeutic targets for intervention.
The current hypothesis lays a conceptual foundation for understanding immune evasion by Pythium insidiosum in the cornea, and further empirical validation is essential to substantiate the proposed mechanisms. To strengthen the translational potential of this hypothesis, it is acknowledged that the proposed experimental design remains conceptual and requires validation in controlled corneal infection models. Future studies will focus on minimizing confounding factors that may influence immune escape, including host immune variability, corneal epithelial integrity, microbial load, and environmental factors. Standardized ex vivo human corneal models and in vivo animal models will be employed to simulate physiological immune responses within the ocular microenvironment. Appropriate experimental controls, reproducible inoculum preparation, and temporal cytokine profiling will be integrated to distinguish pathogen-driven immune modulation from secondary host effects. These approaches will enable precise delineation of the structural and molecular components of P. insidiosum—including its cellulose-rich wall and hydroxyproline-rich glycoproteins—that contribute to altered recognition by pattern recognition receptors and dampened inflammatory signalling. Such refined validation strategies will help substantiate the immune-camouflage hypothesis and deepen understanding of corneal immune evasion mechanisms.
Challenges and Future Directions
While the present work is conceptual, translating the immune-camouflage hypothesis into experimental validation poses particular challenges. Pythium insidiosum is a fastidious organism, and standard fungal immunoassays may not adequately capture its cellulose-rich, β-glucan–deficient wall interactions. Future studies could focus on Pattern Recognition Receptor (PRR) activation assays, particularly those targeting the TLR2, TLR4, and Dectin-1 pathways, to elucidate innate immune signalling in response to Pythium antigens. In vitro cytokine profiling of corneal epithelial and stromal cells, along with cell wall unmasking experiments using β-glucan- or cellulose-targeted enzymes, may provide insights into how the pathogen modulates host recognition. Establishing reproducible infection models in corneal tissue or organoids could further strengthen mechanistic understanding. Acknowledging these limitations and defining potential experimental frameworks not only reinforces the hypothesis's plausibility but also provides a roadmap for future translational studies exploring immunomodulatory or enzyme-assisted therapies for Pythium keratitis.
Conclusion
The hypothesis that Pythium insidiosum keratitis is driven by immuno-camouflage provides a coherent explanation for many perplexing aspects of this disease – from the subtle clinical onset and diagnostic difficulty, to the failure of typical antifungals and the dramatic tissue destruction. While some molecular details remain to be established, the pieces of the puzzle (cellulose shielding, delayed cytokine responses, survival in macrophages, etc.) fit a narrative in which the host is initially “fooled” and then retaliates too late. Validating this theory would not only help tailor more effective therapies (e.g., by combining unmasking agents with targeted drugs and immunotherapy) but also shift the paradigm for managing recalcitrant infections: sometimes, helping the immune system recognize the pathogen is as essential as directly attacking the microbe. Ultimately, the interplay of Pythium and the host immune system is a battle of disguise and discovery. By uncovering the strategies Pythium employs, we can tip the balance in the host’s favour, potentially transforming an almost always blinding infection into one that can be detected early, treated effectively, and even prevented in those at risk. The story of Pythium insidiosum may well exemplify that in the contest between pathogens and the immune system, it’s not always brute force that wins the day – sometimes, it’s stealth. And knowing that, we can respond in kind: by finding ways to illuminate the infiltrator and call the immune system to arms at the right place and time.
In conclusion, this hypothesis proposes that Pythium insidiosum employs immune camouflage to evade early host recognition, representing a novel conceptual advance in understanding its corneal pathogenicity. Future validation of this hypothesis will require targeted experimental approaches, including PRR activation assays, cytokine profiling, and cell wall–unmasking studies to confirm the pathogen’s immune-modulatory behaviour. These experiments, coupled with molecular and immunohistochemical analyses, could provide definitive insights into the mechanisms of immune evasion and guide the development of effective immunotherapeutic strategies.
Funding
No external support, either public or private, was received for the conduct of this study.
Disclosure
The authors declare no conflict of interest.
References
1. Vanittanakom N, Supabandhu J, Khamwan C, et al. Identification of emerging human-pathogenic pythium insidiosum by serological and molecular assay-based methods. J Clin Microbiol. 2004;42(9):3970–3974. PMID: 15364977; PMCID: PMC516349. doi:10.1128/JCM.42.9.3970-3974.2004
2. Gurnani B, Natarajan R, Mohan M, Kaur K. Breaking-down barriers: proposal of using cellulose biosynthesis inhibitors and cellulase enzyme as a novel treatment modality for vision threatening pythium insidiosum keratitis. Clin Ophthalmol.;18:765–776. PMID: 38495678; PMCID: PMC10941664. doi:10.2147/OPTH.S450665
3. Gurnani B, Kaur K, Agarwal S, et al. Pythium insidiosum Keratitis: past, Present, and Future. Ophthalmol Ther. 2022;11(5):1629–1653. PMID: 35788551; PMCID: PMC9255487. doi:10.1007/s40123-022-00542-7
4. Gurnani B, Kaur K, Chaudhary S, et al. Decoding the rare diagnostic enigma of Pythium insidiosum keratitis – a narrative review. TNOA J Ophthalmic Sci Res. 2023;61(4):408–426. doi:10.4103/tjosr.tjosr_97_23
5. Gurnani B, Kaur K. Anti-infective therapies for Pythium insidiosum keratitis. Expert Rev Anti Infect Ther. 2024;22(10):805–817. PMID: 39268901. doi:10.1080/14787210.2024.2403146
6. Cao B, Gonugunta VT, Radhakrishnan N, et al. Outcomes of Pythium keratitis: a meta-analysis of individual patient data. Curr Ophthalmol Rep. 2022;10(4):198–208. PMID: 37250102; PMCID: PMC10211475. doi:10.1007/s40135-022-00302-7
7. Montgomery ML, Fuller KK. Experimental models for fungal keratitis: an overview of principles and protocols. Cells.;9(7):1713. PMID: 32708830; PMCID: PMC7408389. doi:10.3390/cells9071713
8. Chander J, Chakrabarti A, Sharma A, Saini JS, Panigarhi D. Evaluation of calcofluor staining in the diagnosis of fungal corneal ulcer. Mycoses. 1993;36(7–8):243–245. PMID: 7509453. doi:10.1111/j.1439-0507.1993.tb00758.x
9. Mpakosi A, Kaliouli-Antonopoulou C. Immune mechanisms of filamentous fungal keratitis. Cureus.;16(6):e61954. PMID: 38855487; PMCID: PMC11162199. doi:10.7759/cureus.61954
10. Leal SM, Cowden S, Hsia YC, Ghannoum MA, Momany M, Pearlman E. Distinct roles for Dectin-1 and TLR4 in the pathogenesis of aspergillus fumigatus keratitis. PLoS Pathog.;6(7):e1000976. PMID: 20617171; PMCID: PMC2895653. doi:10.1371/journal.ppat.1000976
11. Ahirwar LK, Sharma S. Elevated levels of interleukins, leukocyte protein and cathelicidin antimicrobial peptide are strongly associated with early to mid-stage of Pythium insidiosum infection in rabbit corneas. Curr Eye Res. 2022;47(5):677–687. PMID: 35179411. doi:10.1080/02713683.2021.2023192
12. Wongprompitak P, Pleewan N, Tantibhedhyangkul W, et al. Involvement of Toll-like receptor 2 on human corneal epithelium during an infection of Pythium insidiosum. Asian Pac J Allergy Immunol. 2020;38(2):129–138. PMID: 30118247. doi:10.12932/AP-110518-0311
13. Broz P, Monack DM. Molecular mechanisms of inflammasome activation during microbial infections. Immunol Rev. 2011;243(1):174–190. PMID: 21884176; PMCID: PMC3170129. doi:10.1111/j.1600-065X.2011.01041.x
14. Schenk M, Fabri M, Krutzik SR, et al. Interleukin-1β triggers the differentiation of macrophages with enhanced capacity to present mycobacterial antigen to T cells. Immunology. 2014;141(2):174–180. PMID: 24032597; PMCID: PMC3904238. doi:10.1111/imm.12167
15. Sathapatayavongs B, Leelachaikul P, Prachaktam R, et al. Human pythiosis associated with thalassemia hemoglobinopathy syndrome. J Infect Dis. 1989;159(2):274–280. PMID: 2644370. doi:10.1093/infdis/159.2.274
16. Gurnani B, Kaur K. Pythium Keratitis. In: StatPearls. Treasure Island (FL): StatPearls Publishing; 2025. PMID: 34424645.
17. Gurnani B, Christy J, Narayana S, Rajkumar P, Kaur K, Gubert J. Retrospective multifactorial analysis of Pythium keratitis and review of literature. Indian J Ophthalmol. 2021;69(5):1095–1101. PMID: 33913840; PMCID: PMC8186601. doi:10.4103/ijo.IJO_1808_20
18. Hu L, Huang X, Yee NH, et al. Pythium insidiosum: an emerging pathogen that is easily misdiagnosed and given treatment as a fungus. Front Cell Infect Microbiol.;14:1430032. PMID: 39268488; PMCID: PMC11390559. doi:10.3389/fcimb.2024.1430032
19. Ruiz-Herrera J, Ortiz-Castellanos L. Cell wall glucans of fungi. A review. Cell Surf.;5:100022. PMID: 32743138; PMCID: PMC7389562. doi:10.1016/j.tcsw.2019.100022
20. Batbayar S, Lee DH, Kim HW. Immunomodulation of fungal β-Glucan in host defense signaling by Dectin-1. Biomol Ther. 2012;20(5):433–445. PMID: 24009832; PMCID: PMC3762275. doi:10.4062/biomolther.2012.20.5.433
21. Qazi BS, Tang K, Qazi A. Recent advances in underlying pathologies provide insight into interleukin-8 expression-mediated inflammation and angiogenesis. Int J Inflam. 2011;2011:908468. PMID: 22235381; PMCID: PMC3253461. doi:10.4061/2011/908468
22. Krajaejun T, Rujirawat T, Lohnoo T, et al. Secretome profiling by proteogenomic analysis shows species-specific, temperature-dependent, and putative virulence proteins of Pythium insidiosum. 8(5):527. doi:10.3390/jof8050527 J Fungi (Basel)
23. Ghosh S. Sialic acid and biology of life: an introduction. sialic acids and sialoglycoconjugates in the biology of life. Health and Disease. 2020:1–61. PMCID: PMC7153325. doi:10.1016/B978-0-12-816126-5.00001-9
24. Ledur PC, Tondolo JSM, Jesus FPK, et al. Dendritic cells pulsed with Pythium insidiosum (1,3)(1,6)-β-glucan, Heat-inactivated zoospores and immunotherapy prime naïve T cells to Th1 differentiation in vitro. Immunobiology. 2018;223(3):294–299. PMID: 29074300. doi:10.1016/j.imbio.2017.10.033
25. Puangsricharern V, Chotikkakamthorn P, Tulvatana W, et al. Clinical characteristics, histopathology, and treatment outcomes of Pythium keratitis: a retrospective cohort study. Clin Ophthalmol.;15:1691–1701. PMID: 33935486; PMCID: PMC8080432. doi:10.2147/OPTH.S303721
26. Ryu JS, Kim SY, Kim MK, Oh JY. Inflammation confers healing advantage to corneal epithelium following subsequent injury. Int J Mol Sci.;24(4):3329. PMID: 36834749; PMCID: PMC9962668. doi:10.3390/ijms24043329
27. Cusick MF, Libbey JE, Fujinami RS. Molecular mimicry as a mechanism of autoimmune disease. Clin Rev Allergy Immunol. 2012;42(1):102–111. PMID: 22095454; PMCID: PMC3266166. doi:10.1007/s12016-011-8294-7
28. Gurnani B, Kaur K, Chaudhary S, et al. Decoding the rare diagnostic enigma of Pythium insidiosum keratitis – a narrative review. TNOA J Ophthalmic Sci Res. 2023;61(4):408–426. doi:10.4103/tjosr.tjosr_97_23
29. Goodridge HS, Underhill DM. Fungal recognition by TLR2 and Dectin-1. Handb Exp Pharmacol. 2008; (183):87–109. PMID: 18071656. doi:10.1007/978-3-540-72167-3_5
30. Plato A, Hardison SE, Brown GD. Pattern recognition receptors in antifungal immunity. Semin Immunopathol. 2015;37(2):97–106. PMID: 25420452; PMCID: PMC4326652. doi:10.1007/s00281-014-0462-4
31. Wan J, Lin J, Hu Y, Wei M, Zou Y, Yuan Z. Repeat keratoplasty for failed therapeutic keratoplasty for microbial keratitis: an analysis of characteristics and risk factors. J Ophthalmol. 2020;2020:9041837. PMID: 32104598; PMCID: PMC7036106. doi:10.1155/2020/9041837
32. Medhasi S, Sangphech N, Permpalung N, Torvorapanit P, Plongla R, Worasilchai N. Functional characterization of macrophages and change of Th1/Th2 balance in patients with pythiosis after Pythium insidiosum antigen immunotherapy. Sci Rep.;14(1):27363. PMID: 39521871; PMCID: PMC11550834. doi:10.1038/s41598-024-78756-x
33. Uribe-Querol E, Rosales C. Control of phagocytosis by microbial pathogens. Front Immunol.;8:1368. PMID: 29114249; PMCID: PMC5660709. doi:10.3389/fimmu.2017.01368
34. Zambrano CG, Braga CQ, Dal Ben V, et al. Experimental Pythium aphanidermatum Infection in Rabbits. J Comp Pathol. 2022;190:30–35. PMID: 35152969. doi:10.1016/j.jcpa.2021.12.001
35. Ma H, Chan JFW, Tan YP, et al. NLRP3 inflammasome contributes to host defense against Talaromyces marneffei Infection. Front Immunol.;12:760095. PMID: 34912336; PMCID: PMC8666893. doi:10.3389/fimmu.2021.760095
36. Sanches JM, Ayilam Ramachandran R, Mussi N, Baniasadi HR, Robertson DM. IL-1β-Mediated immunometabolic adaptation in corneal epithelial cells. J Inflamm Res.;18:9537–9555. PMID: 40697419; PMCID: PMC12282541. doi:10.2147/JIR.S495323
37. Oldstone MB. Molecular mimicry and immune-mediated diseases. FASEB J. 1998;12(13):1255–1265. PMID: 9761770; PMCID: PMC7164021. doi:10.1096/fasebj.12.13.1255
38. Rotchanapreeda T, Sae-Chew P, Lohnoo T, et al. Immunological cross-reactivity of proteins extracted from the oomycete Pythium insidiosum and the fungus Basidiobolus ranarum compromises the detection specificity of immunodiagnostic assays for pythiosis. 7(6):474. doi:10.3390/jof7060474 J Fungi
39. Wanachiwanawin W, Mendoza L, Visuthisakchai S, et al. Efficacy of immunotherapy using antigens of Pythium insidiosum in the treatment of vascular pythiosis in humans. 22(27–28):3613–3621. doi:10.1016/j.vaccine.2004.03.031 Vaccine
40. Figueiredo CA, Alcantara-Neves NM, Amorim LD, et al. Evidence for a modulatory effect of IL-10 on both Th1 and Th2 cytokine production: the role of the environment. Clin Immunol. 2011;139(1):57–64. PMID: 21285005; PMCID: PMC3070800. doi:10.1016/j.clim.2010.12.019
41. Khatua B, Roy S, Mandal C. Sialic acids siglec interaction: a unique strategy to circumvent innate immune response by pathogens. Indian J Med Res. 2013;138(5):648–662. PMID: 24434319; PMCID: PMC3928697.
42. Zhai W, Wu F, Zhang Y, Fu Y, Liu Z. The immune escape mechanisms of Mycobacterium Tuberculosis. Int J Mol Sci.;20(2):340. PMID: 30650615; PMCID: PMC6359177. doi:10.3390/ijms20020340
43. Chitasombat MN, Jongkhajornpong P, Lekhanont K, Krajaejun T. Recent update in diagnosis and treatment of human pythiosis. PeerJ.;8:e8555. PMID: 32117626; PMCID: PMC7036273. doi:10.7717/peerj.8555
44. Yolanda H, Krajaejun T. History and perspective of immunotherapy for pythiosis. Vaccines.;9(10):1080. PMID: 34696188; PMCID: PMC8539095. doi:10.3390/vaccines9101080
45. Gurnani B, Kaur K, Venugopal A, et al. Pythium insidiosum keratitis - A review. Indian J Ophthalmol. 2022;70(4):1107–1120. PMID: 35325996; PMCID: PMC9240499. doi:10.4103/ijo.IJO_1534_21
46. Ud-Naen S, Tansit T, Kanistanon D, Chaiprasert A, Wanachiwanawin W, Srinoulprasert Y. Defective cytokine production from monocytes/macrophages of E-beta thalassemia patients in response to Pythium insidiosum infection. Immunobiology. 2019;224(3):427–432. PMID: 30765134. doi:10.1016/j.imbio.2019.02.002
47. Samanta I. Cutaneous, subcutaneous and systemic mycology. Veterinary Mycology. 2015;31:11–153. PMCID: PMC7122059. doi:10.1007/978-81-322-2280-4_4
48. Mendoza L, Newton JC. Immunology and immunotherapy of the infections caused by Pythium insidiosum. Med Mycol. 2005;43(6):477–486. PMID: 16320491. doi:10.1080/13693780500279882
49. Taylor PR, Tsoni SV, Willment JA, et al. Dectin-1 is required for beta-glucan recognition and control of fungal infection. Nat Immunol. 2007;8(1):31–38. PMID: 17159984; PMCID: PMC1888731. doi:10.1038/ni1408
50. Bulmer GS, de Andrade P, Field RA, van Munster JM. Recent advances in enzymatic synthesis of β-glucan and cellulose. Carbohydr Res. 2021;508:108411. PMID: 34392134; PMCID: PMC8425183. doi:10.1016/j.carres.2021.108411
51. Vos GM, Hooijschuur KC, Li Z, et al. Sialic acid O-acetylation patterns and glycosidic linkage type determination by ion mobility-mass spectrometry. Nat Commun.;14(1):6795. PMID: 37880209; PMCID: PMC10600165. doi:10.1038/s41467-023-42575-x
52. Klarlund JK, Callaghan JD, Stella NA, Kowalski RP, McNamara NA, Shanks RMQ. Use of collagen binding domains to deliver molecules to the cornea. J Ocul Pharmacol Ther. 2019;35(9):491–496. PMID: 31593501; PMCID: PMC6839418. doi:10.1089/jop.2019.0065
53. Andes FT, Adam S, Hahn M, et al. The human sialic acid-binding immunoglobulin-like lectin Siglec-9 and its murine homolog Siglec-E control osteoclast activity and bone resorption. Bone. 2021;143:115665. PMID: 33007530. doi:10.1016/j.bone.2020.115665
54. Saini P, Adeniji OS, Abdel-Mohsen M. Inhibitory Siglec-sialic acid interactions in balancing immunological activation and tolerance during viral infections. EBioMedicine. 2022;86:104354. PMID: 36371982; PMCID: PMC9663867. doi:10.1016/j.ebiom.2022.104354
55. Xia D, Toy R, Pradhan P, et al. Enhanced immune responses to vaccine antigens in the corneal stroma. J Control Release. 2023;353:434–446. PMID: 36462639; PMCID: PMC9892265. doi:10.1016/j.jconrel.2022.11.045
56. Gurnani B, Kaur K. Understanding barriers, recommended solutions, and future prospects for the diagnosis and management of Pythium insidiosum keratitis. Indian J Ophthalmol.;71(12):3584–3586. PMID: 37991287; PMCID: PMC10788758. doi:10.4103/IJO.IJO_1041_23
57. Wagener J, Malireddi RK, Lenardon MD, et al. Fungal chitin dampens inflammation through IL-10 induction mediated by NOD2 and TLR9 activation. PLoS Pathog.;10(4):e1004050. PMID: 24722226; PMCID: PMC3983064. doi:10.1371/journal.ppat.1004050
58. Sridapan T, Krajaejun T. Nucleic acid-based detection of Pythium insidiosum: a systematic review. J Fungi.;9(1):27. PMID: 36675848; PMCID: PMC9863793. doi:10.3390/jof9010027
59. Strzelec M, Detka J, Mieszczak P, Sobocińska MK, Majka M. Immunomodulation-a general review of the current state-of-the-art and new therapeutic strategies for targeting the immune system. Front Immunol.;14:1127704. PMID: 36969193; PMCID: PMC10033545. doi:10.3389/fimmu.2023.1127704
60. Wang Y, Abel K, Lantz K, Krieg AM, McChesney MB, Miller CJ. The Toll-like receptor 7 (TLR7) agonist, imiquimod, and the TLR9 agonist, CpG ODN, induce antiviral cytokines and chemokines but do not prevent vaginal transmission of simian immunodeficiency virus when applied intravaginally to rhesus macaques. J Virol. 2005;79(22):14355–14370. Erratum in: J Virol. 2006;80(17):8846. PMID: 16254370; PMCID: PMC1280235. doi:10.1128/JVI.79.22.14355-14370.2005
61. Alsaab HO, Sau S, Alzhrani R, et al. Checkpoint signaling inhibition for cancer immunotherapy: mechanism, combinations, and clinical outcome. Front Pharmacol.;8:561. PMID: 28878676; PMCID: PMC5572324. doi:10.3389/fphar.2017.00561
62. Glanz VY, Myasoedova VA, Grechko AV, Orekhov AN. Inhibition of sialidase activity as a therapeutic approach. Drug Des Devel Ther.;12:3431–3437. PMID: 30349196; PMCID: PMC6186905. doi:10.2147/DDDT.S176220
63. Tondolo JSM, Loreto ES, de Jesus FPK, Ledur PC, Verdi CM, Santurio JM. Immunotherapy based on Pythium insidiosum mycelia drives a Th1/Th17 response in mice. Med Mycol.;58(8):1120–1125. PMID: 32396166. doi:10.1093/mmy/myaa023
64. Saris A, Qin W, van Linge CCA, et al. The azithromycin Pro-drug CSY5669 boosts bacterial killing while attenuating lung inflammation associated with pneumonia caused by methicillin-resistant staphylococcus aureus. Antimicrob Agents Chemother. 66(9):e0229821. doi:10.1128/aac.02298-21
65. Firdaus NHW, Mallick B, Nandakumar KS, Shakya AK. Advantages of using complement components in preventive and therapeutic vaccine strategies for infectious and non-infectious diseases. J R Soc Interface. 2025;22(228):20250138. PMID: 40592462; PMCID: PMC12212990. doi:10.1098/rsif.2025.0138
66. Zhu BB, Zhou J, Zheng J, et al. Corneal graft melting: a systematic review. Int J Ophthalmol. 13(3):493–502. doi:10.18240/ijo.2020.03.19
67. Gurnani B, Kaur K. Therapeutic Keratoplasty. In: StatPearls. Treasure Island (FL): StatPearls Publishing; 2025. PMID: 37276297.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Predicting Prognosis Based on Regional Prevalence, Ulcer Morphology and Treatment Strategy in Vision-Threatening Pythium insidiosum Keratitis
Gurnani B, Kaur K
Clinical Ophthalmology 2023, 17:1307-1314
Published Date: 5 May 2023
Breaking-Down Barriers: Proposal of Using Cellulose Biosynthesis Inhibitors and Cellulase Enzyme as a Novel Treatment Modality for Vision Threatening Pythium Insidiosum Keratitis
Gurnani B, Natarajan R, Mohan M, Kaur K
Clinical Ophthalmology 2024, 18:765-776
Published Date: 11 March 2024