")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
IL-33 Contributes to the Pathological Changes of Hair Follicles in Psoriasis: A Potential Target for Psoriatic Alopecia
Authors Dai C , Chen H, Jiao M , Zhang N, Tang X , Fan A , Liu S , Qian Z , Wang C , Xu Y , Tan Z, Zeng F , Zheng F
Received 29 December 2022
Accepted for publication 7 March 2023
Published 12 March 2023 Volume 2023:16 Pages 639—650
DOI https://doi.org/10.2147/CCID.S403075
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Jeffrey Weinberg
Chan Dai,1 Huoying Chen,2 Mengya Jiao,1 Na Zhang,1 Xuhuan Tang,1 Anqi Fan,3 Shiwang Liu,1 Zhigang Qian,1 Chenchen Wang,1 Yong Xu,1 Zheng Tan,1,4 Fanfan Zeng,5 Fang Zheng1,4
1Department of Immunology, School of Basic Medicine, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China; 2Department of Laboratory Medicine, The Second Affiliated Hospital of Guilin Medical University, Guilin, Guizhou, People’s Republic of China; 3College of Life Science, Yangtze University, Jingzhou, Hubei, People’s Republic of China; 4Key Laboratory of Organ Transplantation, Ministry of Education, NHC Key Laboratory of Organ Transplantation, Key Laboratory of Organ Transplantation, Chinese Academy of Medical Sciences, Wuhan, Hubei, People’s Republic of China; 5Department of Clinical Laboratory, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China
Correspondence: Fang Zheng, Department of Immunology, School of Basic Medicine, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430000, Hubei, People’s Republic of China, Email [email protected] Fanfan Zeng, Department of Clinical Laboratory, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430000, Hubei, People’s Republic of China, Email [email protected]
Purpose: IL-33 is constitutively expressed in skin tissues. Alopecia, a T cells-driven disorder of the hair follicles (HFs), is a common complication in the development of psoriasis. However, the role of IL-33 in psoriatic alopecia remains uncovered. Here, we investigated the roles of IL-33 in inducing pathological changes of hair follicles in psoriasis.
Patients and Methods: Clinical samples and imiquimod (IMQ)-induced psoriatic mice samples were used to investigate the pathological changes and T-cell infiltration of HFs. By using immunohistochemistry staining, the distribution and expression alteration of IL-33 in HFs were determined. Next, by using IL-33 and ST2 knockout mice, we investigated the role of IL-33/ST2 axis in the pathological changes of HFs in psoriasis. Meanwhile, recombinant IL-33 protein was subcutaneous injected to confirm its effect. Finally, RNA sequencing was used to clarify the genes and signaling pathways that involved in this process. Differentially expressed genes were further verified by RT-PCR in cultured HFs in vitro.
Results: We found that the pathological changes of HFs and T cells infiltration in imiquimod-induced psoriatic mice were similar to that in psoriasis patients. The IL-33 positive keratinocytes in the outer root sheath of HFs were increased in both psoriasis patients and psoriatic model mice compared with the controls. By using gene knockout mice, we found that the pathological changes and T cell infiltration were attenuated in IL-33−/− and ST2−/− psoriatic model mice. In addition, subcutaneous injection of recombinant IL-33 exacerbated the pathological changes of HFs and T cell infiltration. RNA sequencing and RT-RCR revealed that IL-33 upregulated the transcription of genes related to keratinocytes proliferation and T lymphocytes chemotaxis.
Conclusion: Our study identifies that IL-33 promotes the pathological changes of HFs in psoriasis, which contributes to psoriatic alopecia. Inhibition of IL-33 may be a potential therapeutic approach for psoriatic alopecia.
Keywords: IL-33, psoriatic alopecia, hair follicle, keratinocytes
Introduction
Psoriatic alopecia is one of the most common complications in the development of psoriasis, especially in severe cases. More than 80% of psoriasis patients suffered from alopecia throughout the course of the disease. Psoriasis causes different types of alopecia, including patchy or diffuse alopecia in the psoriatic lesion area, telogen alopecia and scar baldness, among which patchy alopecia accounts for about 75% of patients with psoriasis alopecia.1 Psoriatic alopecia affects not only the scalp but also other parts of the body. It seriously affects patients’ daily life and work, and brings heavy burdens to patients’ psychological and social life.2,3
Several anti-psoriatic drugs show an alleviating effect on psoriatic skin, such as glucocorticoids, cyclosporine A,4 TNF-α inhibitors, an anti-IL-23 antibody, an IL-17 receptor blocker, and IL-17 inhibitors.5–7 However, for patients with psoriatic alopecia, those drugs are not satisfactory and cause hair growth disorders.8,9 Therefore, seeking a treatment that can effectively treat psoriasis while also restoring hair follicles to health, which warrants further exploration.
In psoriatic alopecia, there are some specific pathological changes in HFs.10 The pathological changes include increasing proportion of catagen or telogen hair follicles, atrophy of sebaceous gland, hyperplasia of keratinocytes in the outer root sheath of HFs and T cell infiltration.11–14 The inner root sheaths and outer root sheaths of HF are composed by keratinocytes, which is originated from epidermal keratinocytes that grow vertical downward.15–17 Under inflammatory stimulation, keratinocytes release various cytokines and chemokines that mediate T cell activation and migration. Cytokines released by activated T cells, such as IL-17, TNF-α, IL-23, IL-22 and IFN-γ, act on keratinocytes in HFs and cause HFs pathological changes, ultimately leading to alopecia.18 However, the regulatory factors driving T cell accumulation and HFs pathological changes in the psoriatic inflammatory microenvironment have not been well reported.
IL-33 is a nuclear cytokine and constitutively expressed in epithelial cells, endothelial cells, fibroblasts and astrocytes.19 IL-33 is a dual-function protein that acts as a cytokine or as an intracellular nuclear factor. As an intracellular nuclear factor, IL-33 has transcriptional regulatory functions associated with the homeodomain-like helix-turn-helix motif.20 As a cytokine, IL-33 is released upon tissue damage, inflammatory stimulation and mechanical stress.21,22 The released IL-33 acts on the target cells expressing ST2 receptor to exert its biological function.23 Previous studies have reported the important role of IL-33 in allergic, fibrotic, infectious, inflammatory and autoimmune diseases.24,25 A recent study reported that people with psoriatic alopecia had increased level of IL-33.26 However, the potential role of IL-33 in psoriatic alopecia has not been clarified.
In this study, the distribution and expression of IL-33 was analyzed in psoriatic HFs. We further found the effect of IL-33/ST2 axis on the pathological changes in HFs of IMQ-induced psoriatic mice. Finally, we determined the direct effect of IL-33 on keratinocytes. We identified a potential therapeutic target associated with the pathological changes of HFs in psoriasis.
Materials and Methods
Human Skin Samples
Normal skin samples were obtained from surgical waste. The psoriatic skin samples were obtained from the biopsy of psoriasis patients in the Second Affiliated Hospital of Guilin Medical University (Guilin, China). All human studies were approved by Ethics Committee of the Second Affiliated Hospital of Guilin Medical University (Ethical approval number: NO.GZR2018001; Approval date: August 21, 2018), and were conformed to the Declaration of Helsinki. Oral and written explanations about this study were obtained for all the participants.
Mice
The WT, IL-33−/− and ST2−/− were C57BL/6 background. All mice in this study were maintained in specific pathogen-free conditions. All mice experiments performed in this study were in strict accordance with the National Institutes of Health guide for the care and use of Laboratory animals and the Institutional Animal Care and Use Committee, Tongji Medical College, Huazhong University of Science and Technology.
Induction of Psoriatic HFs
7–8 week-old male mice were used to induce psoriatic HFs. First, mice were anesthetized. Next, their hair in the back was shaved. Then, 62.5 mg of imiquimod cream (MedShine, Chengdu, China) was smeared on the back skin for 5 consecutive days.
HE Staining
HE staining performed in this study followed standard operating procedures. After dewaxing, slices were incubated in hematoxylin for 2 mins followed by eosin staining. Then the slices were dehydrated with ethanol and sealed with neutral gum.
Immunohistochemistry and Immunofluorescence Staining
The skin samples were fixed at 4% paraformaldehyde and embedded in paraffin. Repair antigen was conducted using the thermal repair method. For immunohistochemistry and immunofluorescence, paraffin sections were stained as reported previously.27 The slices were incubated with the primary antibodies anti-IL-33 (R&D, cat: af3626, USA), anti-CD3 (Boster, cat: PB9093, China) and anti-ST2 (Abcam, cat: ab25877, USA) overnight at 4℃. For immunohistochemistry, the DAB substrate kit and the secondary antibody (Zhongshan Golden Bridge, Beijing, China) were used according to the manufacturer’s manual. For immunofluorescence, the slices were incubated in the dark with the secondary antibody at 37℃ for 30 mins. Nuclei were counterstained by incubation with DAPI at room temperature for 10 mins.
Primary Keratinocyte Culture
The primary keratinocytes were isolated and cultured as previously described.28 The primary keratinocytes were separated from neonatal mice and incubated in 154CF medium (Gibco, cat: M-154CF-500, USA) containing 2mg/mL dispase II at 4℃ for 10 hours. The epidermis was peeled off and washed with cold Hank’s Balanced Salt Solution (Gibco, cat:14175–095, USA). Then, the epidermal keratinocytes were separated by digestion with 0.05% trypsin for 10 mins. Primary keratinocytes were seeded in 48-well plate and cultured in a 5% carbon dioxide incubator.
Cell Immunofluorescence
Keratinocytes were fixed with 4% paraformaldehyde solution for 15 mins and then rinsed with PBS. Cells were incubated with blocking buffer (0.3% Triton X-100 and 10% donkey serum in PBS) for 45 mins at 37℃. Then, cells were incubated with the primary antibody Keratin 14 (Abcam, cat: ab181595, USA) overnight at 4℃. After being washed with PBST, cells were incubated in the dark with the secondary antibody solution at 37℃ for 45 mins. Nuclei were counterstained with DAPI for 10 mins at room temperature. Afterwards, cells were washed three times to remove unbound DAPI. The fluorescence images were acquired using a confocal laser-scanning microscope (LSM 780, Carl Zeiss AG) and analyzed with ImageJ software.
RNA Sequencing Analysis
After two days of culture, primary keratinocytes were treated with 200 ng/mL IL-33 (Biolegend, cat: 580504, USA). After incubation for 12 h, keratinocytes were harvested. Total RNA was extracted with TRIzol (Invitrogen, USA) and used for RNA sequencing analysis. RNA sequencing was performed by Genminix Informatics co., Ltd. (Shanghai, China). Datasets related to this article can be found at the Gene Expression Omnibus datasets using accession numbers at: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE145963
Human Anagen HFs
Anagen HFs were obtained from a human post-mortem occipital scalp region through the body donor program of the Huazhong University of Science and Technology, China, in accordance with legal requirements. Before their deaths, the donors voluntarily declared that their remains would be donated as materials for scientific research. The dissected occipital scalp was cut into three pieces (2×4 cm). As previously described,29 HFs were plucked from one piece and cultured in the Williams E medium (Gibco, USA) with 10 ng/mL hydrocortisone, 10ng/mL insulin, 2 mM L-glutamine, and 100 U/mL penicillin streptomycin (Gibco, USA) at 37℃ in a 5% carbon dioxide incubator. 200 ng/mL IL-33 (R&D, cat: 3625-IL-010, USA) was added to the basal Williams E medium. After being cultured for 12 hours, HFs were harvested and used for the next experiments.
Mouse Anagen HFs
Hair growth was induced by depilating the back hairs with a hair plucking sticker in 7 or 8-week-old male C57BL/6 mice. At day 9 after depilation, HFs were in anagen VI.30 Mouse anagen HFs were dissected from the back depilated region and divided into single follicular units under the dissecting microscope. HFs were cultured at 37℃ in a 5% carbon dioxide incubator. The composition of culture media was conducted as above. 200 ng/mL IL-33 (Biolegend, cat: 580504, USA) was added to the basal Williams E medium. After being cultured for 12 hours, HFs were harvested and used for the next experiments.
Real-Time PCR
Human and mouse HFs were homogenized in TRIzol (Invitrogen, USA). The total RNA of HFs was reversely transcribed into cDNA using the Reverse Transcription Kit (Thermo Fisher Scientific, USA). All primers were prepared by Sangon Biotech (Shanghai, China). The primers used in this study were listed in Supplementary Table S1.
Statistical Analysis
All data were presented as the mean ± SEM. The difference between the two groups was evaluated using by a two-tailed Student’s t-test. P value < 0.05 were considered to be statistically significant. *P <0.05, **P<0.01, ***P< 0.001.
Results
The Expression of IL-33 in the HFs of Psoriatic Patients and Psoriatic Model Mice is Increased
To observe the effects of psoriasis on HFs, skin samples from patients with and without psoriasis were collected and performed HE staining. The results showed that, in contrast to normal skin, most HFs in psoriatic skin were showed hyperkeratosis of infundibulum, thickening of outer root sheaths and atrophy of sebaceous glands (Figure 1a). Immunohistochemical staining showed that for normal skin, IL-33 was stained predominantly in epidermal keratinocytes and rarely in the keratinocytes of the outer root sheath, whereas for psoriatic skin, IL-33 was stained obviously in the keratinocytes of both epidermis and outer root sheath. More importantly, compared with normal skin, IL-33-positive keratinocytes were significantly increased in the outer root sheath of HFs in the psoriatic skin (Figure 1b and d). In addition, immunohistochemical staining of CD3 was performed to determine the infiltration and distribution of T cells. It showed that CD3-positive cells were seldomly observed in the HFs of normal skin, while easily observed in the HFs of psoriatic skin. Statistically, CD3-positive cells were significantly increased in and around the HFs of psoriatic patients relative to normal skin (Figure 1c and d). Taken together, above data indicated that IL-33-positive keratinocytes and T cells were increased in the outer root sheath of HFs in psoriasis. Next, IMQ was applied to induce psoriatic mice model. Compared with normal skin, IMQ-induced psoriatic skin showed apparently hyperplasia in the outer root sheath of HFs. As a result, the diameter and length of HFs in IMQ-induced psoriatic skin were obviously increased relative to normal skin. In addition, the number of sebaceous glands in the IMQ-induced psoriatic skin was significantly reduced relative to normal skin (Figure 1e and h). Similar to psoriasis patients, immunohistochemical staining showed that the number of IL-33-positive keratinocytes in HFs was markedly increased in IMQ-induced psoriatic mice skin (Figure 1f and h). Besides, immunohistochemical staining of CD3 indicated that the number of T cells infiltrated in and around the HFs was also increased in IMQ-induced psoriatic skin (Figure 1g and h). In conclusion, above data indicated that the pathological changes and T cell infiltration of HFs in psoriatic model mice were similar to those in psoriatic patients.
 |
Figure 1 The pathological changes of HFs in IMQ-induced psoriatic mice were similar to those seen in psoriatic patients. (a) H & E staining of the normal skin and psoriatic skin. Scale bar = 100μm. (i) infundibulum, (ii) outer root sheath, (iii) sebaceous glands. (b) Immunohistochemical staining of IL-33 in normal skin and psoriatic skin. Scale bar=100μm. (c) Immunohistochemical staining of CD3 in normal skin and psoriatic skin. Scale bar = 100μm. (d) Statistical data of HFs diameter, IL-33-positive cells and CD3-positive cells. (e) H & E staining of the normal skin and IMQ-induced psoriatic skin. Scale bar=100μm. (i) infundibulum, (ii) outer root sheath, (iii) sebaceous glands. (f) Immunohistochemical staining of IL-33 in normal skin and IMQ-induced psoriatic skin. Scale bar=100μm. (g) Immunohistochemical staining of CD3 in normal skin and IMQ-induced psoriatic skin. Scale bar=100μm. Red arrows indicate CD3-positive T cells. (h) Statistical data of HFs number, HFs length, HFs diameter, IL-33-positive cells and CD3-positive cells. Data are representative of three independent experiments. Significance was determined by a two-tailed Student’s t-test. * P <0.05, ** P<0.01, *** P< 0.001. |
The Pathological Changes in HFs of IMQ-Induced Psoriatic Mice are Alleviated in IL-33 or ST2 Knockout Mice
Firstly, we confirmed that IL-33 or ST2 knockout had no effect on the morphology of HFs under physiological conditions (Table S2). Next, to clarify the potential role of IL-33 in the pathological changes of HFs in psoriasis, IL-33−/− mice were used to induce psoriatic mice model. The lesion skin samples were collected for HE staining and immunohistochemical staining. The data showed that, compared with WT mice, the pathological changes, including infundibular hyperkeratosis, outer root sheath thickening and sebaceous glands atrophy, were obviously attenuated in the HFs of IL-33−/− mice. The length and diameter of HFs were significantly decreased in IL-33−/− psoriatic model mice compared to WT psoriatic model mice (Figure 2a and c). Immunohistochemical staining showed that the number of CD3+ cells infiltrated in and around the HFs was significantly reduced in IL-33−/− psoriatic model mice relative to WT psoriatic model mice (Figure 2b and d). To furtherly validate the role of IL-33 in HFs pathological changes, ST2−/− mice was applied to psoriatic model induction and pathological analysis. Consistent with the alteration observed above, ST2−/− psoriatic model mice also showed similar attenuation of pathological changes and decreased number of CD3+ cells in and around the HFs compared with the WT psoriatic model mice (Figure 2e–h). Collectively, these data suggested that IL-33/ST2 signaling promoted pathological changes of HFs in psoriasis.
 |
Figure 2 The pathological changes of HFs and T cells infiltration were alleviated in IL-33−/− and ST2−/− mice in IMQ-induced psoriatic model. (a) H & E staining of lesion skin in WT psoriatic mice and IL-33−/− psoriatic mice (n = 6 per group). Scale bar=100μm. (i) infundibulum, (ii) outer root sheath, (iii) sebaceous glands. (b) Immunohistochemical staining of CD3 in WT psoriatic mice and IL-33−/− psoriatic mice (n = 6 per group). Scale bar = 100μm. Red arrows indicate CD3-positive T cells. (c and d) Statistical data of HFs number, HFs length, HFs diameter, sebaceous gland number and CD3-positive cells. (e) H & E staining of lesion skin in WT psoriatic mice and ST2−/− psoriatic mice (n = 6 per group). Scale bar=100μm. (i) infundibulum, (ii) outer root sheath, (iii) sebaceous glands. (f) Immunohistochemical staining of CD3 in WT psoriatic mice and ST2−/− psoriatic mice (n = 6 per group). Scale bar = 100μm. Red arrows indicate CD3-positive T cells. (g and h) Statistical data of HFs number, HFs length, HFs diameter, sebaceous gland number and CD3-positive cells. Data are representative of three independent experiments. Significance was determined by a two-tailed Student’s t-test. * P<0.05, ** P<0.01, *** P<0.001. |
Recombinant IL-33 Injection Aggravates the Pathological Changes in HFs of IMQ-Induced Psoriatic Mice
Although above data showed that IL-33/ST2 signaling deficiency suppressed pathological changes of HFs and T cells infiltration in IMQ-induced psoriatic mice, whether IL-33 had a positive function still needs further confirmation. Next, a subcutaneous injection assay was performed in WT mice to investigate the role of IL-33. The schematic of the treatment performed seen in Figure 3a. The lesion skin in the treated area was collected for HE staining and immunohistochemical staining. The data showed that, compared with IMQ treatment alone, recombinant-IL-33 aggravated HFs pathological changes including infundibular hyperkeratosis, outer root sheaths thickening and sebaceous glands atrophy (Figure 3b and d). Besides, the number of CD3+ cells infiltrated in and around the HFs was significantly increased in recombinant-IL-33 injected mice (Figure 3c and e). Together, these data indicated that IL-33 promoted pathological changes in HFs as well as T cells infiltration, implying the role of IL-33 in HFs probably by affecting T cells infiltration.
 |
Figure 3 Subcutaneous injection of recombinant IL-33 aggravated the pathological changes of HFs and T cells infiltration in IMQ-induced psoriatic mice. (a) Schematic of the treatment performed. (b) H & E staining (n = 4 per group), PBS vs IL-33. Scale bar=100μm. (i) infundibulum, (ii) outer root sheath. (c) Immunohistochemical staining for CD3 (n=4 per group), PBS vs IL-33. Red arrows indicate CD3-positive T cells. Scale bar=100μm. (d and e) Statistical data of HFs number, HFs length, HFs diameter, sebaceous gland number and CD3-positive cells. Data are representative of three independent experiments. Significance was determined by a two-tailed Student’s t-test. * P<0.05, ** P<0.01, *** P<0.001. |
IL-33 Treatment Upregulates Some Genes Related to Keratinocytes Proliferation and T Cell Chemotaxis
In order to investigate the potential target cell of IL-33, immunofluorescent staining of ST2 was performed with mice normal skin and IMQ-induced psoriatic skin. The image showed that ST2 was mainly expressed by outer root sheath keratinocytes as well as epidermal keratinocytes (Figure S1). To determine the effects of IL-33 signaling on the transcription of downstream genes, primary keratinocytes were isolated from neonatal mice and cultured in vitro. The purity was identified by immunofluorescent staining of keratin 14 (Krt14) and it showed to be more than 95% (Figure 4a). Next, primary keratinocytes were treated with recombinant IL-33 for 12 hours and collected for RNA sequencing. GO enrichment analysis showed that IL-33 upregulated genes were closely related to epithelial cell proliferation, keratinocyte differentiation and lymphocyte chemotaxis (Figure 4b). Among these upregulated genes, CCL2 and CCL20 played pivotal role in T cells chemotaxis. The heat map indicated that CCL2 and CCL20 were obviously upregulated by recombinant IL-33 (Figure 4c). Next, RT-PCR was performed to further validate the role of IL-33. The data showed that IL-33 upregulated the transcription level of CCL2 and CCL20 in both mice and human HFs in vitro (Figure 4d and e). Collectively, these data indicated that IL-33 acted on keratinocytes and promoted T cell infiltration by upregulating CCL2 and CCL20.
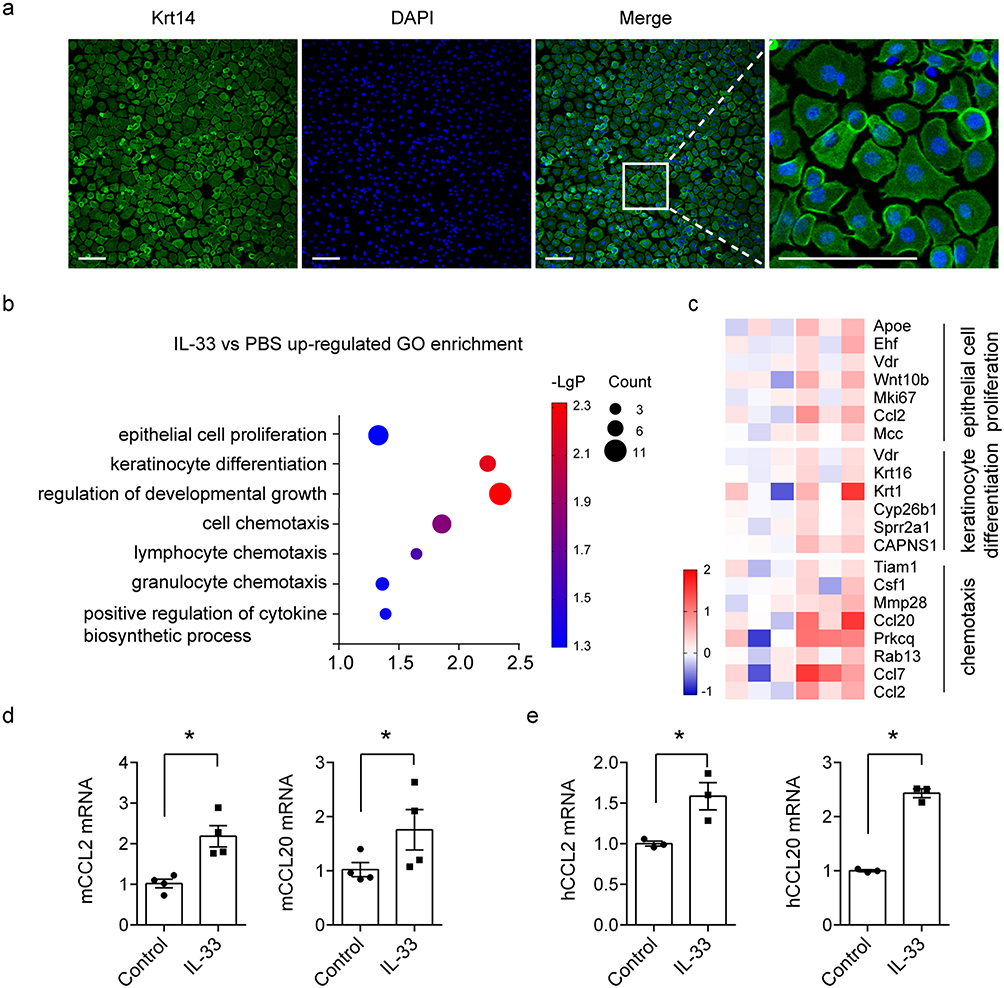 |
Figure 4 Recombinant IL-33 upregulated downstream genes associated with proliferation and chemotaxis in keratinocytes in vitro. (a) Immunofluorescence staining of Krt14 (green) in primary keratinocytes, Scale bar = 100μm. (b) GO enrichment analysis of up-regulated genes upon IL-33 stimulation in mouse primary keratinocytes. 3 samples per group. Count means the number of genes. (c) Heatmap of up-regulated gene. (d and e). Real-time PCR analysis of representative genes, CCL2 and CCL20, in mice and human HFs. Recombinant IL-33 (200 ng/mL) treated for 12 hours. Data are representative of three independent experiments. Significance was determined by a two-tailed Student’s t-test. * P<0.05. |
Discussion
Currently, there is no completely effective treatment to attenuate pathological damage of HFs and psoriatic alopecia. Although present drugs used in clinic have achieved promising results in the treatment of psoriasis, most of these drugs have limited effect on psoriatic alopecia. Some of the anti-psoriasis drugs induce hypertrichosis, and others cause psoriatic alopecia. Immunosuppressive drugs, cyclosporine A, which has been used in psoriasis treatment for quite a few years, caused hypertrichosis via regulating canonical Wnt/β-catenin activity.31,32 Target drugs, TNF-inhibitors were associated with drug-induced psoriasiform alopecia.33,34 Monoclonal antibody, adalimumab, was also reported to induce severe alopecia in scalp psoriasis patients.35 Thus, it is of great necessity to investigate potential and effective targets to improve the treatment of psoriatic alopecia. Here, by means of human samples, psoriatic mouse models, in vivo and in vitro experiments, we confirmed the involvement of IL-33/ST2 axis in the pathological changes of HFs in psoriasis, and proposed that IL-33 may be a potential therapeutic target for the treatment of psoriatic alopecia.
In this study, we believed that IL-33 upregulated the expression of chemokines to promote the infiltration of T cells, thereby contributing to the pathological changes of HFs in psoriasis. In our study, some typical pathological changes were observed in the HFs in psoriasis patients, and IL-33 positive keratinocytes as well as T cells were increased in the outer root sheath of HFs in psoriasis patients. The pathological changes and T cell alteration were then confirmed in IMQ-induced psoriatic model mice in vivo. Next, In vitro experiments proved that IL-33 upregulated the transcription of CCL2 and CCL20. These two chemokines have potent chemotactic roles on T cells. CCL2 was a ligand of CCR4 which was expressed on Th17 cells. CCL20 was bound exclusively to CCR6 which was highly expressed on Th17 and Th1 cells. The recruited T cells released cytokines, such as IL-17, IL-22, IL-23 and TNF-α, to alter the balance of proliferation and differentiation in keratinocytes. Previous studies had reported that T cells played a pivotal role in pathological changes of HFs and caused psoriatic alopecia.36–38 Therefore, based on our findings and previous researches, we concluded that IL-33 contributed to pathological changes of HFs in psoriasis probably by regulating T cell infiltration.
The role of IL-33 in promoting pathological changes of HFs was by means of a cytokine, not a nuclear factor. Because of its nuclear location and its release upon cell damage or stress, IL-33 was proposed to be a dual-function alarmin.19,39–41 Nuclear IL-33 was able to repress gene expression by facilitating chromatin compaction. In addition, IL-33 nuclear location was shown to act as a strong regulator of IL-33 extracellular function. Once released, IL-33 was cleaved by neutrophil elastase and cathepsin G. The resulting C-terminal cleavage products, containing the IL-1-like domain, was considered to be a mature form and had a stronger proinflammatory activity.42–44 The released IL-33 bound to receptor ST2 and activated downstream signaling pathways of the target cells.45 In this study, our data indicated that receptor ST2 was mainly expressed by keratinocytes of the skin. In vivo results demonstrated that receptor ST2 deficiency ameliorated pathological changes of HFs as well as T cell infiltration. In addition, cytokine (recombinant) IL-33 was showed to aggravate HFs pathological changes and T cell infiltration. Furthermore, in vitro results proved that cytokine (recombinant) IL-33 regulated the transcription of some genes associated with T cells chemotaxis in both mice and human HFs. Taken together, we concluded that IL-33 acted as a cytokine to promote the pathological changes of HFs via ST2.
Previous reports showed that the different psoriasis-related cytokines caused various pathological alterations in psoriatic HF. For example, VEGF prolonged the duration of HFs anagen via promoting substantial angiogenesis.46–49 IGF-1 maintained HFs anagen by regulating Wnt/β-catenin activity.50,51 TNF-α and IFN-γ promoted a transformation from anagen to catagen or telogen, and aggravated psoriatic alopecia.52,53 Our study clarified the role of IL-33 in psoriatic alopecia and enriched our understanding of the networks involved in psoriasis-related cytokines.
In summary, the present study reported an unrevealed role of IL-33 in pathological changes of HFs in psoriasis. Our data identified that IL-33 promoted the expression of T-cell chemokines to regulate T-cell infiltration and ultimately contribute to pathological changes of HFs. Our study indicated that inhibition IL-33/ST2 signaling pathway probably provides a potential therapeutic approach for the treatment of psoriatic alopecia.
Conclusion
In psoriatic alopecia, IL-33 was upregulated in the outer root sheath keratinocytes of hair follicles. Upon psoriatic inflammatory stimuli, IL-33 actively released and upregulated the expression of chemokines in keratinocytes to promote the infiltration of T cells, thereby contributing to the pathological changes of HFs and alopecia.
Ethics Statement
Our manuscript, entitled “IL-33 contributes to the pathological changes of hair follicles in psoriasis: a potential target for psoriatic alopecia”, is being submitted. We collected skin samples from patients with and without psoriasis, which were used for HE staining and immunohistochemical staining. The studies were approved by no informed consent is necessary in the scope of this study, since the specimen were analyzed retrospectively and already belonged to the Second Affiliated Hospital of Guilin Medical University at the time of the analyses.
We obtained anagen hair follicles from a human post-mortem occipital scalp region through the body donor program of the Tongji Medical College, the Wuhan Red Cross, China. The HFs were harvested and used for culture in vitro. Before their deaths, the donors voluntarily declared that their remains would be donated as materials for scientific research.
All human studies were approved by Ethics Committee of the Second Affiliated Hospital of Guilin Medical University (Ethical approval number: NO.GZR2018001; Approval date: August 21, 2018).
Acknowledgments
This work was supported by the grant awarded by the National Natural Science Foundation of China [Grant No. 82171761] to Fang Zheng, and National Natural Science Foundation of China [Grant No. 82101900] to Fanfan Zeng.
Disclosure
The authors declare that there are no conflicts of interest.
References
1. Chan CS, Van Voorhees AS, Lebwohl MG, et al. Treatment of severe scalp psoriasis: from the medical board of the national psoriasis foundation. J Am Acad Dermatol. 2009;60(6):962–971. doi:10.1016/j.jaad.2008.11.890
2. Marahatta S, Agrawal S, Adhikari BR. Psychological impact of alopecia areata. Dermatol Res Pract. 2020;2020:8879343.
3. Aghaei S, Saki N, Daneshmand E, Kardeh B. Prevalence of psychological disorders in patients with alopecia areata in comparison with normal subjects. ISRN Dermatol. 2014;2014:304370.
4. Wang T-S, Tsai T-F. Managing scalp psoriasis: an evidence-based review. Am J Clin Dermatol. 2017;18(1):17–43.
5. Megna M, Potestio L, Ruggiero A, Camela E, Fabbrocini G. Guselkumab is efficacious and safe in psoriasis patients who failed anti-IL17: a 52-week real-life study. J Dermatolog Treat. 2022;33(5):2560–2564.
6. Megna M, Potestio L, Fabbrocini G, Ruggiero A. Long-term efficacy and safety of guselkumab for moderate to severe psoriasis: a 3-year real-life retrospective study. Psoriasis Targets Ther. 2022;12:205–212.
7. Megna M, Tommasino N, Potestio L, et al. Real-world practice indirect comparison between guselkumab, risankizumab, and tildrakizumab: results from an Italian 28-week retrospective study. J Dermatol Treat. 2022;33(6):2813–2820.
8. Wong BL, Rybalsky I, Shellenbarger KC, et al. Long-term outcome of interdisciplinary management of patients with Duchenne muscular dystrophy receiving daily glucocorticoid treatment. J Pediatr. 2017;182:296–303.
9. Ngwanya MR, Gray NA, Gumedze F, Ndyenga A, Khumalo NP. Higher concentrations of dithranol appear to induce hair growth even in severe alopecia areata. Dermatol Ther. 2017;30:4.
10. Bruno de Castro S, Bandeira LG, Cunha T, Valente NYS. Follicular psoriasis: an underdiagnosed entity? An Bras Dermatol. 2019;94(1):116–118. doi:10.1590/abd1806-4841.20197987
11. Rittié L, Tejasvi T, Harms PW, et al. Sebaceous gland atrophy in psoriasis: an explanation for psoriatic alopecia? J Invest Dermatol. 2016;136(9):1792–1800. doi:10.1016/j.jid.2016.05.113
12. Sawan S, Descamps V. Scalp psoriasis: a paradigm of “switch-on” mechanism to anagen hair growth? Arch Dermatol. 2008;144(8):1064–1066. doi:10.1001/archdermatol.2008.2
13. Bruni F, Alessandrini A, Starace M, Orlando G, Piraccini BM. Clinical and trichoscopic features in various forms of scalp psoriasis. J Eur Acad Dermatol Venereol. 2021;35(9):1830–1837. doi:10.1111/jdv.17354
14. George SM, Taylor MR, Farrant PB. Psoriatic alopecia. Clin Exp Dermatol. 2015;40(7):717–721. doi:10.1111/ced.12715
15. Lee J, Tumbar T. Hairy tale of signaling in hair follicle development and cycling. Semin Cell Dev Biol. 2012;23(8):906–916.
16. Liu S, Zhang H, Duan E. Epidermal development in mammals: key regulators, signals from beneath, and stem cells. Int J Mol Sci. 2013;14(6):10869–10895.
17. Magerl M, Tobin DJ, Muller-Rover S, et al. Patterns of proliferation and apoptosis during murine hair follicle morphogenesis. J Invest Dermatol. 2001;116(6):947–955.
18. Chiricozzi A, Romanelli P, Volpe E, Borsellino G, Romanelli M. Scanning the Immunopathogenesis of psoriasis. Int J Mol Sci. 2018;19:1.
19. Cayrol C, Girard JP. Interleukin-33 (IL-33): a nuclear cytokine from the IL-1 family. Immunol Rev. 2018;281(1):154–168.
20. Roussel L, Erard M, Cayrol C, Girard JP. Molecular mimicry between IL-33 and KSHV for attachment to chromatin through the H2A-H2B acidic pocket. EMBO Rep. 2008;9(10):1006–1012.
21. Cayrol C. IL-33, an alarmin of the IL-1 family involved in allergic and non allergic inflammation: focus on the mechanisms of regulation of its activity. Cells. 2021;11:1.
22. Martin NT, Martin MU. Interleukin 33 is a guardian of barriers and a local alarmin. Nat Immunol. 2016;17(2):122–131.
23. Bae S, Kang T, Hong J, et al. Contradictory functions (activation/termination) of neutrophil proteinase 3 enzyme (PR3) in interleukin-33 biological activity. J Biol Chem. 2012;287(11):8205–8213.
24. Dwyer GK, D’Cruz LM, Turnquist HR. Emerging functions of IL-33 in homeostasis and immunity. Annu Rev Immunol. 2022;40:15–43.
25. Liew FY, Girard JP, Turnquist HR. Interleukin-33 in health and disease. Nat Rev Immunol. 2016;16(11):676–689.
26. Bain KA, McDonald E, Moffat F, et al. Alopecia areata is characterized by dysregulation in systemic type 17 and type 2 cytokines, which may contribute to disease-associated psychological morbidity. Br J Dermatol. 2020;182(1):130–137.
27. Chen H, Sun Y, Lai L, et al. Interleukin-33 is released in spinal cord and suppresses experimental autoimmune encephalomyelitis in mice. Neuroscience. 2015;308:157–168.
28. Zeng F, Chen H, Chen L, et al. An autocrine circuit of IL-33 in keratinocytes is involved in the progression of psoriasis. J Invest Dermatol. 2020;2020:1.
29. Langan EA, Philpott MP, Kloepper JE, Paus R. Human hair follicle organ culture: theory, application and perspectives. Exp Dermatol. 2015;24(12):903–911.
30. Hendrix S, Handjiski B, Peters EM, Paus R. A guide to assessing damage response pathways of the hair follicle: lessons from cyclophosphamide-induced alopecia in mice. J Invest Dermatol. 2005;125(1):42–51.
31. Hawkshaw NJ, Haslam IS, Ansell DM, Shamalak A, Paus R. Re-evaluating cyclosporine a as a hair growth-promoting agent in human scalp hair follicles. J Invest Dermatol. 2015;135(8):2129–2132.
32. Hawkshaw NJ, Hardman JA, Haslam IS, et al. Identifying novel strategies for treating human hair loss disorders: cyclosporine A suppresses the Wnt inhibitor, SFRP1, in the dermal papilla of human scalp hair follicles. PLoS Biol. 2018;16(5):e2003705.
33. Afanasiev OK, Zhang CZ, Ruhoy SM. TNF-inhibitor associated psoriatic alopecia: diagnostic utility of sebaceous lobule atrophy. J Cutan Pathol. 2017;44(6):563–569.
34. Lauro W, Picone V, Abategiovanni L, et al. A case of psoriatic alopecia secondary to certolizumab pegol: clinical and trichoscopic evaluation. Int J Dermatol. 2022;2022:1.
35. Melé-Ninot G, Expósito-Serrano V, Quintana Codina M, et al. Adalimumab-related alopecia in a patient affected by psoriasis. Dermatol Online J. 2017;23:7.
36. Hong JW, Lee CY, Ha SM, et al. The contributory roles of Th17 lymphocyte and cytotoxic T lymphocyte at the hair bulge region as well as the hair bulb area in the chronic alopecia areata patients. Ann Dermatol. 2017;29(2):156–166.
37. Oka A, Takagi A, Komiyama E, et al. Alopecia areata susceptibility variant in MHC region impacts expressions of genes contributing to hair keratinization and is involved in hair loss. EBioMedicine. 2020;57:102810.
38. Milling S, Siebert S. T cells and cytokines in inflamed psoriatic skin. Who’s in charge? Immunology. 2020;160(4):311–312.
39. Travers J, Rochman M, Miracle CE, et al. Chromatin regulates IL-33 release and extracellular cytokine activity. Nat Commun. 2018;9(1):3244.
40. Aizawa H, Koarai A. Oxidative stress enhances the expression of IL-33 in human airway epithelial cells. Respir Res. 2018;19(1):52.
41. Uchida M, Anderson EL, Squillace DL, et al. Oxidative stress serves as a key checkpoint for IL-33 release by airway epithelium. Allergy. 2017;72(10):1521–1531.
42. Calabrese L, Fiocco Z, Satoh TK, Peris K, French LE. Therapeutic potential of targeting interleukin-1 family cytokines in chronic inflammatory skin diseases. Br J Dermatol. 2022;186(6):925–941.
43. Liu X, Hu R, Pei L, et al. Regulatory T cell is critical for interleukin-33-mediated neuroprotection against stroke. Exp Neurol. 2020;328:113233.
44. Chen Z, Hu Y, Gong Y, et al. Interleukin-33 alleviates psoriatic inflammation by suppressing the T helper type 17 immune response. Immunology. 2020;160(4):382–392.
45. Pinto SM, Subbannayya Y, Rex DAB, et al. A network map of IL-33 signaling pathway. J Cell Commun Signal. 2018;12(3):615–624.
46. Suzuki T, Ito T, Gilhar A, Tokura Y, Reich K, Paus R. The hair follicle-psoriasis axis: shared regulatory mechanisms and therapeutic targets. Exp Dermatol. 2021;2021:1.
47. Mecklenburg L, Tobin DJ, Müller-Röver S, et al. Active hair growth (anagen) is associated with angiogenesis. J Invest Dermatol. 2000;114(5):909–916.
48. Yano K, Brown LF, Detmar M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J Clin Invest. 2001;107(4):409–417.
49. Hou C, Miao Y, Wang J, Wang X, Chen CY, Hu ZQ. Collagenase IV plays an important role in regulating hair cycle by inducing VEGF, IGF-1, and TGF-β expression. Drug Des Devel Ther. 2015;9:5373–5383.
50. Oh JY, Park MA, Kim YC. Peppermint oil promotes hair growth without toxic signs. Toxicol Res. 2014;30(4):297–304.
51. Hodak E, Gottlieb AB, Anzilotti M, Krueger JG. The insulin-like growth factor 1 receptor is expressed by epithelial cells with proliferative potential in human epidermis and skin appendages: correlation of increased expression with epidermal hyperplasia. J Invest Dermatol. 1996;106(3):564–570.
52. Philpott MP, Sanders DA, Bowen J, Kealey T. Effects of interleukins, colony-stimulating factor and tumour necrosis factor on human hair follicle growth in vitro: a possible role for interleukin-1 and tumour necrosis factor-alpha in alopecia areata. Br J Dermatol. 1996;135(6):942–948.
53. Ito T, Ito N, Saathoff M, Bettermann A, Takigawa M, Paus R. Interferon-gamma is a potent inducer of catagen-like changes in cultured human anagen hair follicles. Br J Dermatol. 2005;152(4):623–631.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.