Back to Journals » Journal of Inflammation Research » Volume 18
Identification of SIN3A as a Promising Epigenetic Target Against Allergic Rhinitis
Authors Li H ![]() , Wang R, Yu B, Zeng J, Xiao Z
, Wang R, Yu B, Zeng J, Xiao Z
Received 6 June 2025
Accepted for publication 10 October 2025
Published 27 October 2025 Volume 2025:18 Pages 14971—14987
DOI https://doi.org/10.2147/JIR.S535690
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Yuhan Xing
Hanqiao Li,1– 3 Renkang Wang,1– 3 Benquan Yu,1– 3 Junjie Zeng,1– 3 Zian Xiao1– 3
1Department of Otorhinolaryngology Head and Neck Surgery, The Second Xiangya Hospital, Central South University, Changsha, Hunan Province, People’s Republic of China; 2Institute of Otology, Central South University, Changsha, Hunan Province, People’s Republic of China; 3Laboratory of Otorhinolaryngology Head and Neck Cancer, The Second Xiangya Hospital, Central South University, Changsha, Hunan Province, People’s Republic of China
Correspondence: Zian Xiao, Department of Otorhinolaryngology Head and Neck Surgery, The Second Xiangya Hospital, Central South University, Changsha, 410011, Hunan Province, People’s Republic of China, Email [email protected]
Purpose: The involvement of histone modifications in the development of other airway allergic diseases has been widely studied and followed, however, the role in allergic rhinitis(AR) has not been explored and discussed.
Methods: We downloaded GSE50223 from Gene Expression Omnibus database, obtained histone modification related genes in FACER database, and identified key functional genes related to histone modification in AR by WGCNA and single cell sequencing analysis. Finally, we collected nasal mucosa tissues from AR patients and healthy individuals to verify the expression of the target gene. At the functional level, a co-culture system of house dust mite (HDM)-sensitized human nasal epithelial cells and naive CD4⁺ T cells was employed. Using gene knockdown and overexpression techniques, the role of SIN3A in the differentiation of Th17 and Treg cells was analyzed.
Results: SIN3A was identified as a key functional gene related to histone modification with differential expression in AR, and single-cell data analysis showed that SIN3A might be involved in the immune infiltration difference by regulating Treg cells and thus. Upregulation of SIN3A was confirmed in nasal mucosal tissues of AR patients. Functional experiments demonstrated that SIN3A promotes the differentiation of naive T cells into Th17 cells and inhibits their differentiation into Treg cells, thereby disrupting the Th17/Treg balance.
Conclusion: Histone modification-related gene SIN3A is differentially expressed in AR and healthy populations. SIN3A may play a significant role in the pathogenesis of AR by regulating the Th17/Treg immune balance. This study first revealed a novel epigenetic mechanism by which SIN3A mediates immune imbalance in allergic rhinitis (AR), providing a theoretical basis for the development of intervention strategies targeting SIN3A.
Keywords: allergic rhinitis, histone modification, bioinformatics
Introduction
Allergic rhinitis (AR) is an allergic condition of the nasal mucosa mediated by particular immunoglobulin E (IgE) and involves several immunoreactive cells and cytokines, among other elements.1 Upon re-exposure of atopic persons to allergens, allergic rhinitis is clinically characterized by nasal congestion, rhinorrhea, sneezing, and pruritus of the nasal passages.2 It frequently coexists with asthma or conjunctivitis. AR constitutes a worldwide health issue, affecting patients from many nations, ethnicities, socioeconomic statuses, and age demographics.3,4 Statistical data reveals that between 20% to 30% of adults and as many as 40% of youngsters worldwide are impacted by this illness.4 The prevalence is escalating, with a growing number of patients diagnosed each year.5,6 This substantially affects patients’ social interactions, sleep quality, academic achievement, and work efficiency. Approximately 90% of these individuals receive insufficient treatment, leading to a considerable economic burden.
Although the disease burden of allergic rhinitis (AR) is severe, its pathological mechanism remains incompletely understood. In recent years, the role of epigenetic regulation in allergic diseases has received extensive attention. Histone modifications (such as methylation, acetylation, etc.) as key epigenetic mechanisms can participate in the fine regulation of immune responses by altering chromatin structure and gene transcriptional activity. Existing studies have shown that abnormal histone modifications are closely related to allergic diseases such as asthma and atopic dermatitis.7–9 Some studies have also found that specific probiotics can alleviate AR symptoms by affecting histone H1.2 and HMGB1.10 However, most epigenetic studies still focus on other allergic diseases, and there is a significant knowledge gap regarding the specific histone modification regulatory network in AR and its specific role in immune imbalance. In particular, which histone modification-related genes play a key role in AR and how they affect the differentiation and function of key immune cell subsets such as Th17/Treg have not been systematically explored.
To fill this gap, this study, based on bioinformatics methods, integrated the transcriptome data from the GEO database with the gene set related to histone modification, and screened the hub genes in AR through differential expression analysis and WGCNA. Further, the correlation with the immune microenvironment was revealed through immune infiltration analysis, and the expression was verified using clinical nasal mucosa samples. Finally, the regulatory effect of key genes on the differentiation direction of Th17/Treg was clarified through in vitro cell experiments, aiming to deepen the understanding of the immune mechanism of AR from the epigenetic level and provide a theoretical basis for finding potential therapeutic targets.
Materials and Methods
Materials
This study selected the experimental group of AR patients who underwent postnasal neurectomy at the Department of Otorhinolaryngology Head and Neck Surgery of the Second Xiangya Hospital of Central South University. The control group consisted of patients with uncomplicated deviated nasal septum, devoid of allergic conditions such as allergic rhinitis and asthma, and was selected between February 2023 and September 2023. The Human Research Ethics Committee of the Second Xiangya Hospital of Central South University accepted the study (Ethics approval number: LYF2022222). All patients executed an informed consent document. The research complied with the principles of the Declaration of Helsinki.
RNA Isolation and Real-Time Quantitative PCR (RT-qPCR)
Total RNA was extracted from nasal mucosal tissue samples with TRIzol reagent (Takara, Japan). cDNA was generated using reverse transcription employing a PrimeScript™ RT kit (Takara, Japan) alongside random 6-polymers and oligo dT primers, by the manufacturer’s protocol (Takara, Japan). The relative expression of the target genes was standardized to GAPDH mRNA. All responses were conducted in triplicate, and the data are presented as standard deviation ± mean. The following are the primer sequences:
SIN3A_forward: GCCTGTGGAGTTTAATCATGCC;
SIN3A_reverse: CCTCTTGCTCAGTCAAAGCTG;
GAPDH_forward: AGGTCGGTGTGAACGGATTTG;
GAPDH_reverse: GGCAGAGATGATGACCCTTTT。
Western Blot Analysis
Samples were lysed in RIPA buffer, and protein concentrations were measured with the BCA Protein Assay Kit (Beyotime, China). Proteins were separated via SDS-PAGE, electrotransferred to a PVDF membrane, treated with a primary antibody at 4°C overnight, washed three times, incubated with a secondary antibody for one hour at room temperature, and detected using chemiluminescent reagents. The Anti-SIN3A antibody was acquired from Proteintech (USA).
Immunofluorescence Staining
Following antigen repair, nasal mucosa tissue sections were incubated with 3% H2O2 for 15 minutes, and subsequently treated with 10% goat serum for 1 hour. Subsequently, sections were treated overnight at 4°C with a primary antibody, followed by a secondary antibody. Cell nuclei were labeled with DAPI (Beyotime, China). Angiogenesis was examined utilizing a confocal microscope (Olympus, Japan).
Cell Culture and Reagents
The human nasal mucosal epithelial cells (HNEpCs) line was procured from Ningbo Mingzhou Biotechnology Co., Ltd. The cell passage number was maintained at no more than 15. The cells were cultured in EpiCM (Procell, China), a complete epithelial cell culture medium. All cell cultures were carried out in a humidified incubator maintained at 37°C and 5% carbon dioxide (CO2) concentration. The nasal mucosal epithelial cells were stimulated with 100 μg/mL of house dust mite extract (HDM, GREER, USA). Twenty-four hours post-stimulation, the expression levels of epithelial cell-derived cytokines, namely IL-25, TSLP, and IL-33, were detected.
Co-Culture of Transfected Cell Lines with Naive T Cells
HENpCs were seeded in 6-well plates and grown to approximately 70–80% confluence before transfection. SIN3A siRNA (AM16708; clone ID: 108731) was purchased from Invitrogen. The SIN3A overexpression plasmid (OHS6085-213584153; clone ID: ccsbBroad304-11788) was constructed for this study. Transfection reagents were diluted in serum-free medium using the RNAimax system (including NC, si-SIN3A and oe-SIN3A). The transfection efficiency of SIN3A was measured by RT-qPCR 48 hours after transfection to confirm successful transfection. Four groups were co-cultured with (1.5×106 cells/well) primary T cells (SAIOS, China) for 48 hours: untransfected HENpC without HDM intervention; empty vector-transfected HENpCs with HDM intervention; si-SIN3A-transfected HENpCs with HDM intervention; oe-SIN3A-transfected HENpCs with HDM stimulation.
Flow Cytometry for the Detection of Th17 and Treg Differentiation
After resuspending the cells, Cell Stimulation Cocktail was added and the cells were stimulated at 37°C for 4 hours, followed by centrifugation and washing. CD3 and CD4 antibodies were used for surface staining of the cells, which were incubated in the dark for 30 minutes and then washed with PBS. Subsequently, the cells were fixed with Intracellular Fixation Buffer for 30 minutes, permeabilized with 1×Permeabilization Buffer, and stained intracellularly with IL-17A antibody for 30 minutes in the dark. After washing, the cells were resuspended in PBS containing 0.5% BSA and analyzed on the flow cytometer to determine the proportion of CD3+CD4+IL-17A+ Th17 cells. The detection process for Treg cells was similar. CD4 and CD25 antibodies were used to label surface antigens of the cells, and Foxp3 antibody was used for intracellular staining. The cells were then analyzed on the flow cytometer to determine the proportion of CD4+CD25+Foxp3+ Treg cells.
Data Sources
Gene expression datasets were queried utilizing allergic rhinitis as a keyword. The subsequent criteria were utilized during the screening: 1. It encompassed individuals with allergic rhinitis necessitating a clinically validated diagnosis; 2. The dataset had an extensive number of samples, incorporating both patients and healthy controls; and 3. The samples analyzed were human tissues. The dataset GSE50223 was acquired from the NCBI GEO public database with the annotation platform GPL6884. The dataset comprised 42 transcriptome samples, consisting of 21 control samples and 21 AR samples. From the FACER database, 510 histone modification-related genes were identified, of which 439 were expressed in the GSE50223 dataset.
Differential Expression Analysis
Differential expression analysis was conducted utilizing the most recent version of the “Limma” package in the R 4.0.3 program. Fold change (FC) was determined using the average gene expression from the AR and control groups. Genes with differential expression were identified using threshold values (|logFC|>1 and P < 0.05).
Functional Enrichment Analysis of Differential Genes
To thoroughly investigate the functional significance of these differentially expressed genes (DEGs), we analyzed the DEGs in the regular and AR groups utilizing the “limma” package from Bioconductor in R. We examined the differentially expressed genes using the “clusterProfiler” package in R and conducted gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses. The “clusterProfiler” package in R was utilized to identify differentially expressed genes in the regular and AR groups, and to assess the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment of these genes, applying a significance criterion of P < 0.05. Only upon meeting these criteria were the differentially expressed genes subjected to analysis. Significance criterion; only paths meeting the aforementioned conditions were deemed considerably enriched.
Immune Infiltration Analysis
To assess the impact of differentially expressed AR genes on immune infiltration, ssGSEA was employed to measure the degree of immune cell infiltration in each sample. A Pearson correlation analysis was conducted for gene expression and immune cell composition. A p-value of less than 0.05 was deemed statistically significant.
WGCNA Network Construction
To identify critical genes connected with pathophysiological pathways in allergic rhinitis patients, we employed the expression profiling data from GSE50223 to form a WGCNA network, thereby investigating the co-expression regulatory networks linked to allergic rhinitis. The modules exhibiting substantial association were identified as essential based on their relationship with sample categories.
ROC Analysis of Hub Genes in Histone Modification, Analysis of Associated Drugs, and Immune Cell Infiltration Assessment
The Receiver Operating Characteristic (ROC) function is graphically shown by plotting the true positive rate on the horizontal axis and the false positive rate on the vertical axis, used to evaluate the model’s accuracy. The Area Under the Curve (AUC) represents the area beneath the ROC function, with values approaching 1 indicating superior performance. The efficacy of the effect was evaluated using the ROC function to compute the AUC value of the Hub genes for the sample categories, determining their suitability as indicators for these categories. Subsequently, the Drug-Gene Interaction databases (DGIdb) were employed to identify and present the medications interacting with the Hub gene. The relationship between histone-modified Hub genes and different immune cells was further analyzed.
Single-Cell Sequencing Analysis
The Seurat package was utilized for quality control and subsequent data analysis. The positional links among the clusters were determined by UMAP algorithm analysis, and the clusters were annotated utilizing MonacoImmuneData files. Ultimately, we refined each subgroup for marker genes utilizing the FindAllMarkers algorithm, with |avg_log2FC| > 1 and a corrected P-value < 0.05 deemed as distinctive marker genes for each cell subtype.
Statistical Analysis
A two-tailed p-value of less than 0.05 was deemed significant. All analyses were conducted using R software (version 4.1.1) and Prism (version 8.1) for Windows 64.0.
Results
Identification and Functional Study of Histone-Associated Differentially Expressed Genes (DEGs)
Acquire the gene chip dataset of GSE50223 from the NCBI GEO public database, then employ the limma package to identify differentially expressed genes between regular and AR groups, with the screening criteria of |logFC|>1 and P<0.05, resulting in the identification of 316 differentially expressed genes (Figure 1a and b). Subsequently, we intersected with 439 histone modification-related genes, resulting in the identification of 7 differentially expressed genes associated with histone modifications: SIN3A, SIRT2, UBE2E1, PRDM10, BANP, YWHAZ, and SET. To enhance our comprehension of the potential roles of histone modification-associated differentially expressed genes, we employed the clusterProfiler package in R to conduct Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) functional enrichment analyses, with a significance threshold of P<0.05. The results indicated that these differential genes were significantly enriched in biological processes such as DNA replication (GO:0006260), cellular component disassembly (GO:0022411), protein-containing complex disassembly (GO:0032984), and mitochondrial gene expression (GO:0140053), among others (Figure 1c). No substantial enrichment pathway for differential genes was identified in KEGG.
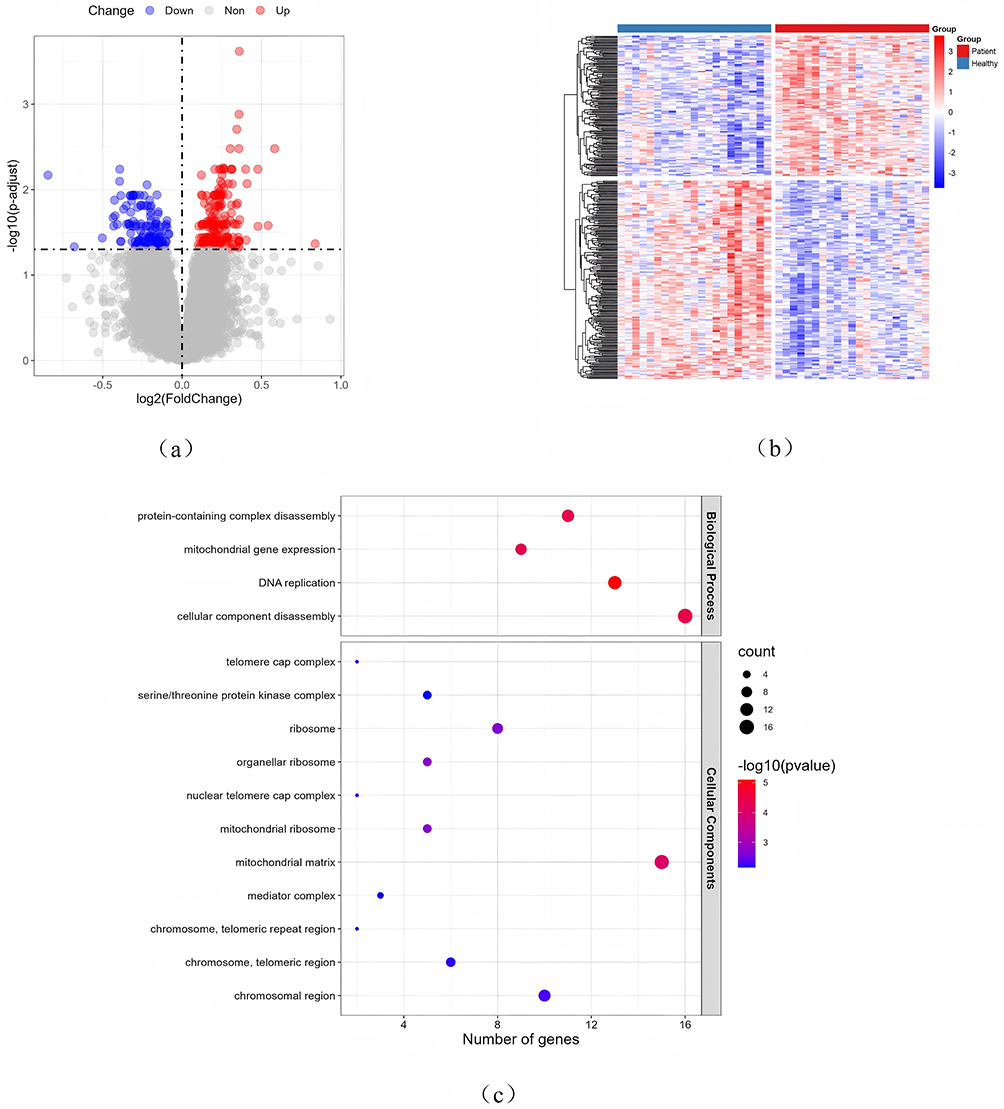 |
Figure 1 Identification and functional analysis of histone-related DEGs. (a) Volcano map of differentially expressed genes (DEGs) in GSE50223; (b) Heat map of DEGs expression; (c) GO enrichment pathway analysis of histone modification-associated DEGs. |
Modified Immunological Microenvironment in Individuals with Allergic Rhinitis
The etiology of allergic rhinitis (AR) encompasses inflammation and immunological responses, with numerous immune cells and proteins participating in its pathophysiological pathways. To investigate the mechanisms of its immune response, we employed ssGSEA to evaluate the disparities in the abundance of various immune cells between AR patients and healthy individuals, subsequently comparing the discrepancies in component samples. Marked disparities were observed between AR patients and the healthy control group in cell types such as Effector memory CD8 T cells, Monocytes, and Natural killer cells (Figure 2). This aligns with earlier data indicating that NK cells may play a role in the pathophysiology of allergic disorders by elevating type 2 cytokines and facilitating eosinophil migration, and that NK cell levels are heightened in individuals with allergic rhinitis.11,12
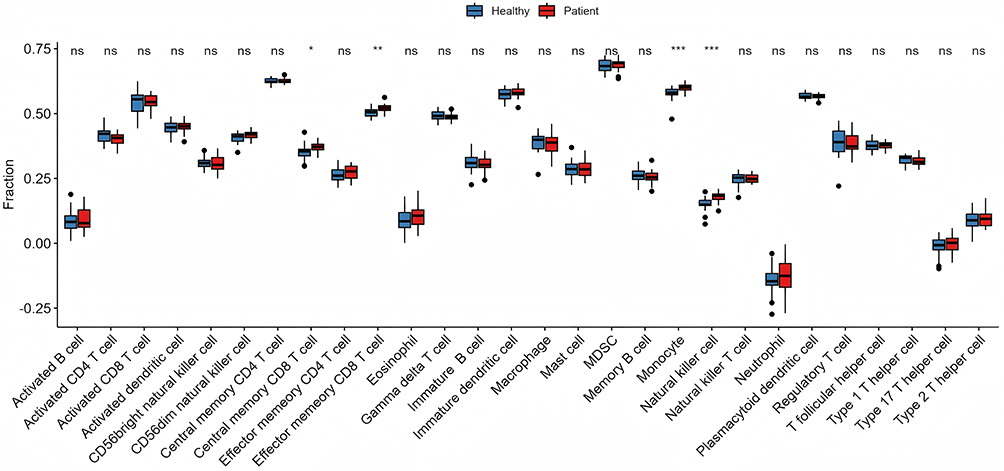 |
Figure 2 Immune microenvironment analysis. Non-allergic rhinitis group in blue and allergic rhinitis group in red. “*” indicates p<0.05, “**” indicates p<0.01, and “***” indicates p<0.001, “ns” indicates ‘not significant’. |
Identification of Pivotal Genes Associated with Histone Modification by WGCNA
To identify essential genes associated with the pathophysiological mechanisms of allergic rhinitis (AR) patients, we employed the expression profile data from GSE50223 to develop a weighted gene co-expression network analysis (WGCNA) for investigating the co-expression regulatory network pertinent to AR. Based on the association between modules and sample types, we identified the modules with considerable strong correlation as essential modules. Upon thorough investigation, we discovered a substantial negative correlation between the log-log(k) of nodes with connectivity k and the log-log(P(k)) of the corresponding node’s occurrence probability. The intensity of this negative correlation is quantified by a correlation coefficient exceeding 0.8, as illustrated in Figure 3a. According to the assessment to ascertain the power=12, the log10(P(k)) and log10(k) were By plotting and squaring the correlation coefficient, which is 0.89—indicative of a value approaching 1—it is evident that the two variables exhibit a strong linear relationship, suggesting the construction of a scale-free network in this instance (Figure 3b).
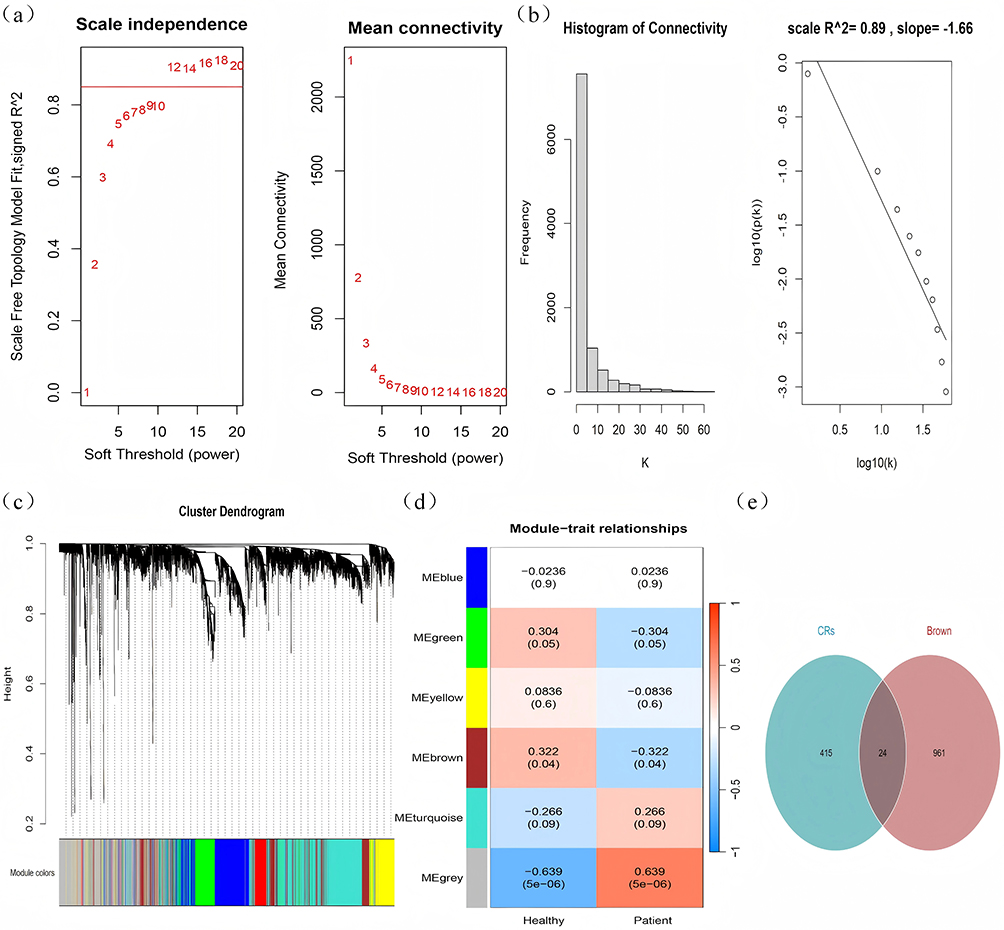 |
Figure 3 Construction of WGCNA modules. (a) Evaluation of optimal soft threshold; (b) study of scale-free fit index and average connectivity; (c) dendrogram for gene clustering; (d) heatmap depicting module-phenotype associations. (e) Venn diagram illustrating the intersection of WGCNA brown module genes and histone modification-related genes, with green circles representing the 439 histone modification-related genes expressed in the GSE50223 dataset and brown circles denoting the WGCNA brown module genes. |
The tom matrix ultimately classified these genes into six modules (Figure 3c), with each row of Figure 3d depicting the eigenvector gene of each module, while the columns display the classification information of the samples. The figure in each cell represents the correlation coefficient of the gene module with the categorization of the respective sample, whilst the figure in parentheses denotes the significance level p-value. The findings revealed that the brown module exhibited the most substantial significant positive connection with AR, leading us to identify the brown module as the principal module associated with AR, comprising 985 genes. We subsequently identified the histone modification genes within the brown module, designating them as histone modification hub genes (Figure 3d), which included ASF1A, BRD2, CDK2, CHD7, SIN3A, and 24 other genes (Figure 3e). SIN3A was identified as a differentially expressed gene, as previously described.
ROC Analysis of Histone Modification Hub Genes, Associated Medication Analysis, and Immune Cell Infiltration Assessment
Initially, the ROC function was employed to compute the AUC values of the Hub genes relative to the sample categories, revealing that the expression levels of the majority of genes effectively indicated the AR sample categories. Notably, 16 genes exhibited AUC values exceeding 0.6, with SIN3A, SUPT16H, and NCOA5 demonstrating AUCs of 0.87, 0.77, and 0.76, respectively (Figure 4a). The findings indicated that SIN3A is a crucial element in allergic rhinitis and is a differentially expressed gene in Result 2.3.1, potentially serving as a target for elucidating the mechanisms underlying AR pathogenesis.
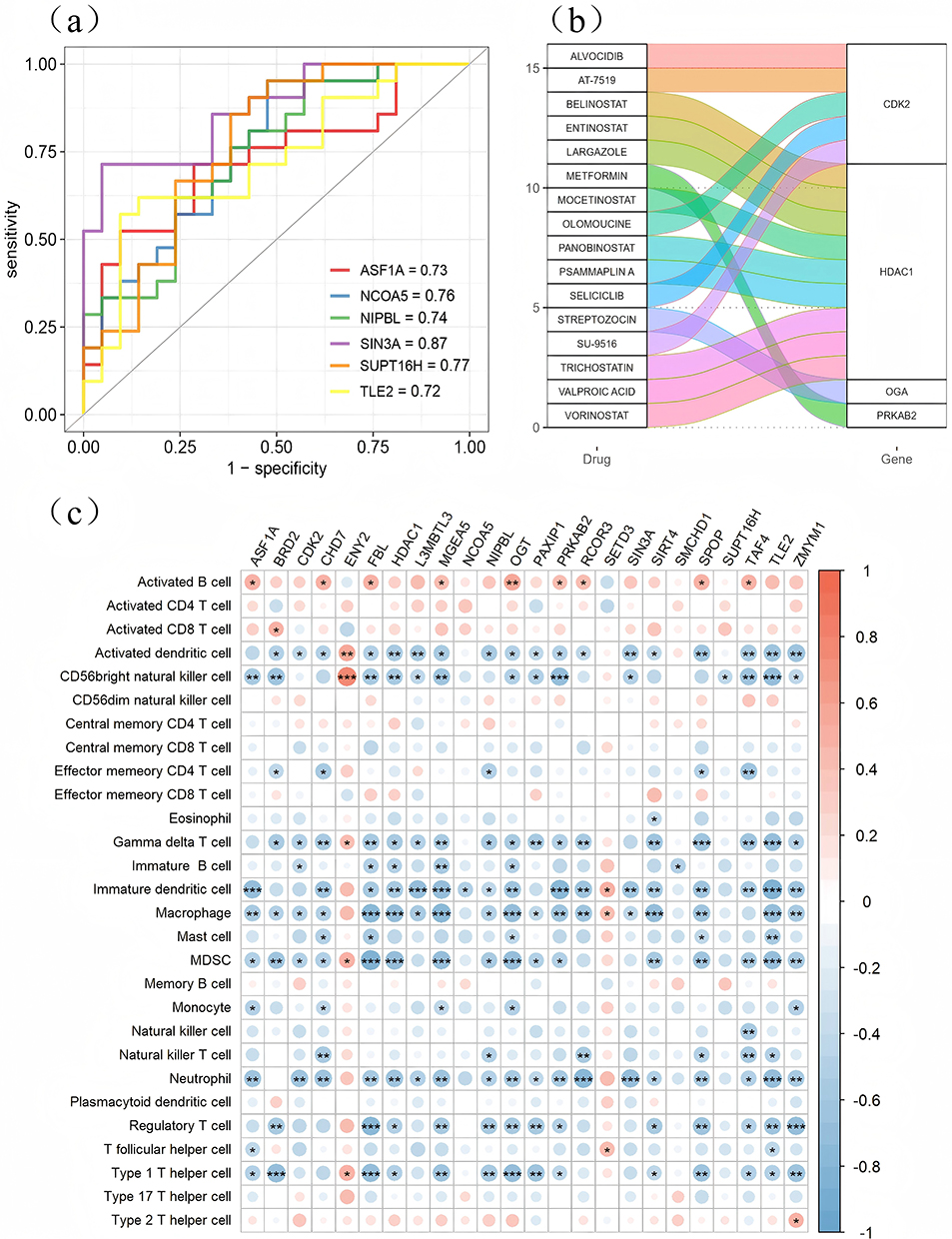 |
Figure 4 Correlation analysis of hub genes associated with histone modifications. (a) The ROC curves for histone modification-associated Hub genes are presented, highlighting the six Hub genes with the highest AUC values in the figure; (b) Interaction between histone modification-associated hub genes and pharmaceuticals; (c) Correlation between histone modification-associated hub genes and immune cells, with the left side of the figure depicting the types of immune cells and the upper side displaying 24 histone modification-associated hub genes. Red dots signify positive correlations, while blue dots denote negative correlations. “*” indicates p<0.05, “**” indicates p<0.01, and “***” indicates p<0.001, ’ns’ indicates ‘not significant’. |
Subsequently, the DGIdb database was employed to identify drugs that interact with the Hub genes, focusing exclusively on gene-drug interactions documented in a minimum of three publications to ensure credibility. This process yielded four histone-modified genes (PRKAB2, OGA, HDAC1, and CDK2) along with their corresponding interacting drugs (Figure 4b), presenting novel candidates for allergic rhinitis treatment. SIN3A failed to obtain pertinent drug matches.
We examined the link between histone-modified Hub genes and immunological infiltration, observing a significant and robust correlation between the majority of these histone-modified Hub genes and immune cells (Figure 4c).
SIN3A is the Only Histone Modification Hub Gene with Significant Differential Expression in the AR
We screened seven histone modification-related genes exhibiting significant differential expression in AR and intersected them with the histone modification Hub key genes identified via the WGCNA network. The final results indicated that among these Hub key genes, only SIN3A demonstrated significant differential expression between the disease and control groups. Figure 4c illustrates that SIN3A exhibits a substantial positive connection with neutrophils, immature dendritic cells, activated dendritic cells, and other immune cells. This suggests that SIN3A, a cluster of essential genes for protein modification, may influence the immunological milieu and immune control in AR patients, hence contributing to the pathophysiological mechanisms of AR.
Single-Cell Analysis of SIN3A and AR Immune Microenvironment
To further examine the function of SIN3A in the AR immune microenvironment and to confirm the expression of SIN3A at the single-cell level, the dataset GSE2001074 (peripheral blood samples from patients with allergic rhinitis before and after SLIT treatment) was retrieved from the GEO database, and its scRNA data were utilized for subsequent analysis. Following the filtration of low-quality cells as per the threshold established in Method 2.2.6, the resultant single-cell data were downscaled and clustered utilizing the UMAP method (Figure 5a). Figure 5b illustrates the clustering of various samples prior to and subsequent to de-batching, executed at a resolution of 0.5, resulting in the division into 14 cell clusters (Figure 5c). Four principal cell types were identified through annotation: CD4+ cytotoxic T-cells (CD4+ CTL, marker gene GZMA), CD4+ regulatory T cells (CD4+ Treg, marker genes FOXP3, IL2RA), CD4+ naive T cells (Naive CD4+ T cells, marker gene SELL), and CD4+ helper T cells (CD4+ Th17, marker genes KLRB1, CCR6) (Figure 5). As illustrated in Figure 5d, the expression of SIN3A is observed in these four types of immune cells.
 |
Figure 5 Single-cell data analysis of SIN3A in AR. (a–d) UMAP clustering results: (a) “pre” on the right signifies the period prior to SLIT treatment, while “post” denotes the period following SLIT treatment; (b) illustrates the clustering of various samples before and after de-batching; (c) on the right, the four principal cell types identified through annotation are: CD4+ cytotoxic T cells, CD4+ regulatory T cells, CD4+ naïve T cells, and CD4+ helper T cells17; (d) depicts SIN3A expression across each immune cell type. (e) Differential expression of the SIN3A gene across various immune cell types in allergic rhinitis patients prior to sublingual immunotherapy treatment. (f) Differential expression of the SIN3A gene in various immune cell types in allergic rhinitis patients following sublingual immunotherapy treatment. |
The SIN3A gene exhibited significant and varied expression in the tissues of patients with allergic rhinitis across distinct cell groups, both prior to and following SLIT treatment. Figure 5e illustrates a significant disparity in SIN3A expression between Naive CD4+ T cells and CD4+ Treg cells prior to treatment, as Naive CD4+ T cells can differentiate into CD4+ Treg populations, which play a crucial role in immune regulation in vivo. Conversely, after SLIT treatment, no significant differences were observed among the various cell populations (Figure 5f), indicating that SIN3A expression may be a critical factor influencing the condition of patients with allergic rhinitis.
Correlation Study of SIN3A with Prevalent Immune Checkpoints and Human Leukocyte Antigens
The findings from the single-cell study indicate that SIN3A could be a crucial immunoregulatory gene in allergic rhinitis. To elucidate the function of SIN3A in allergic rhinitis immunology, we examined its connection with prevalent immunological checkpoints and human leukocyte antigens. Figure 6 demonstrates a substantial positive correlation between SIN3A and HLA-C, HLA-DQB2, HLA-F, and ADORA2A.
 |
Figure 6 Correlation of SIN3A with common immune checkpoints and human leukocyte antigens. (A) Correlation of SIN3A with common immune checkpoints; (B) Correlation of SIN3A with common immune checkpoints. |
Validation of SIN3A in Nasal Mucosal Tissues
This study recruited 12 patients with allergic rhinitis (AR), aged 18–52 years, and 12 control patients with nasal septum deviations, aged 17–59 years (Table 1), to compare the expression levels of SIN3A in the nasal mucosal tissues of AR patients against those of patients with simple deviated nasal septum. The expression level of SIN3A in the nasal mucosal tissues of allergic rhinitis patients was significantly elevated compared to the control group, with the difference being statistically significant (Figure 7a–c).
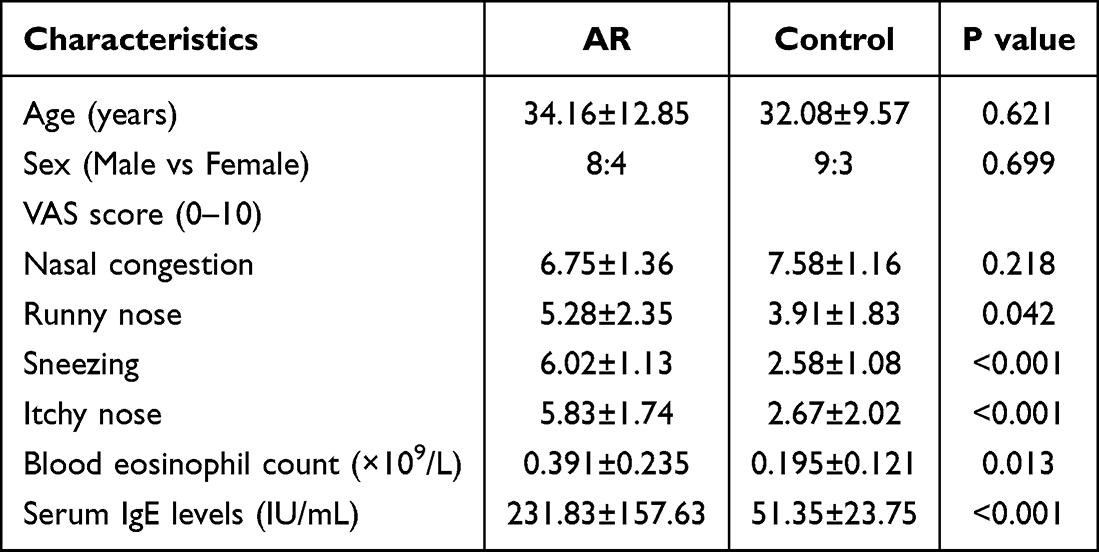 |
Table 1 Basic Clinical Information and Laboratory Findings of Recruits |
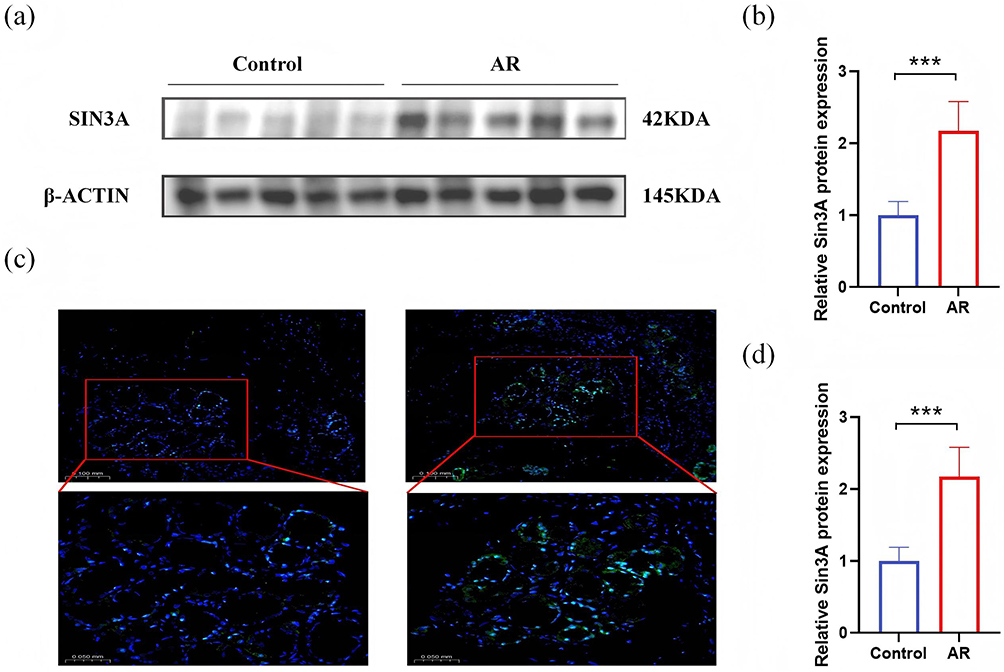 |
Figure 7 SIN3A is differentially expressed in AR patients.(a and b) Relative protein expression of SIN3A in AR and controls. (c) SIN3A expression in nasal mucosa (immunofluorescence detection 20×; 40×). (d) Relative mRNA expression of SIN3A in AR and control. “***” indicates p<0.001. |
The Effect of SIN3A on the Differentiation of Th17/Treg in HDM-Sensitized Nasal Epithelial Cells
To clarify the role of SIN3A in regulating T cell differentiation in nasal mucosal epithelial cells, we first established an in vitro allergic rhinitis (AR) model by treating HNEpCs with house dust mite (HDM). Western Blot results showed that compared with the Control group, the protein expression of key inflammatory factors such as IL-25, IL-33, and TSLP in HDM-treated nasal mucosal epithelial cells was significantly increased (Figure 8a and b), indicating the successful establishment of the AR cell model.
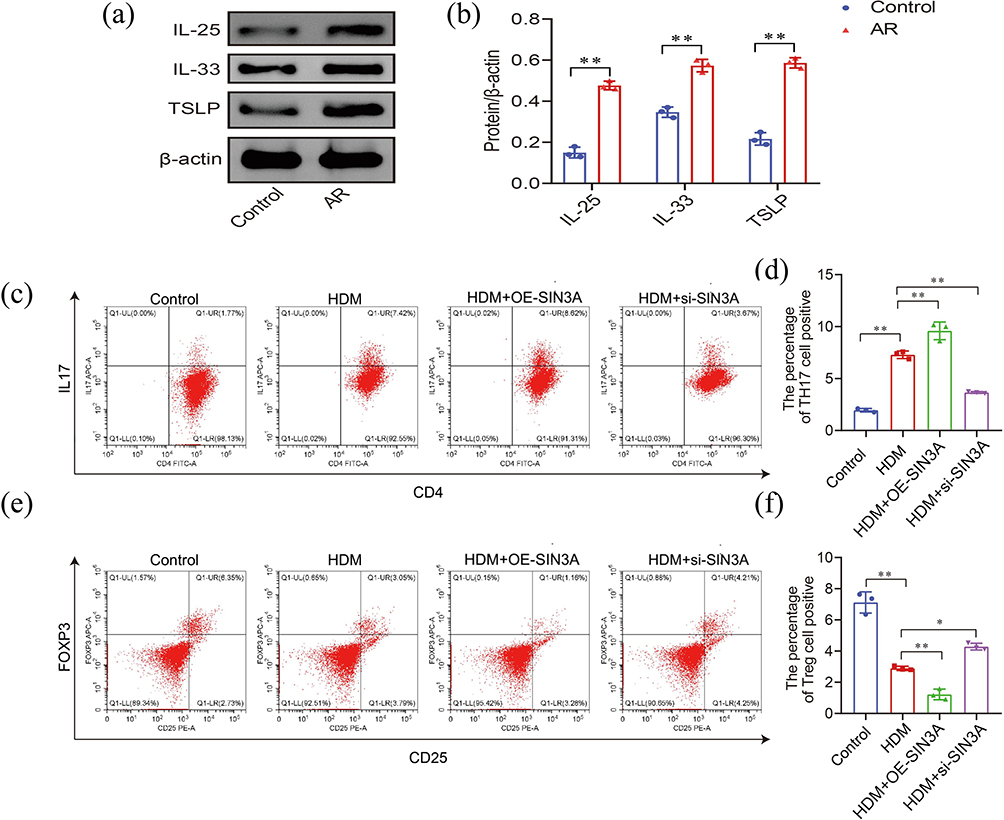 |
Figure 8 Functional verification of SIN3A in in vitro cell experiments. (a and b) Relative expression levels of IL-25, IL-33 and TSLP proteins in HDM-sensitized nasal mucosal epithelial cells. (c-f) Representative flow cytometry plots of IL-17A and Foxp3 expression in naive CD4+T cells co-cultured with nasal mucosal epithelial cells from different treatment groups. Experimental groups were as follows: control group (untreated HNEpCs), HDM group (HDM-sensitized HNEpCs), HDM + si-SIN3A group (HDM-sensitized and SIN3A knockdown HNEpCs), HDM + OE-SIN3A group (HDM-sensitized and SIN3A overexpressed HNEpCs). (d) Quantitative analysis of Th17 cell proportions in each group; (f) Quantitative analysis of Treg cell proportions in each group. |
Subsequently, we co-cultured nasal mucosal epithelial cells under different treatment conditions with naive T cells and detected the proportions of Th17 (CD4+IL-17A+) and Treg (CD4+CD25+Foxp3+) cells in CD4+ T cells by flow cytometry. The experimental groups were set up as follows: (i) Control group: naive T cells co-cultured with untreated HNEpCs; (ii) HDM group: naive T cells co-cultured with HDM-sensitized HNEpCs; (iii) HDM + si-SIN3A group: naive T cells co-cultured with HDM-sensitized HNEpCs in which SIN3A was knocked down; (iv) HDM + OE-SIN3A group: naive T cells co-cultured with HDM-sensitized HNEpCs in which SIN3A was overexpressed.
Flow cytometry analysis results (Figure 8c–f) indicated that compared with the Control group, the proportion of Th17 cells in the co-culture system of the HDM group was significantly increased, while the proportion of Treg cells was significantly decreased, suggesting that in the allergic inflammatory microenvironment, nasal mucosal epithelial cells can disrupt the Th17/Treg immune balance. To further verify the function of SIN3A in this process, we conducted SIN3A overexpression and knockdown experiments. The results showed that overexpression of SIN3A in HDM-sensitized cells could further enhance its regulatory effect on T cell differentiation: the proportion of Th17 cells was significantly higher than that in the HDM group, while the proportion of Treg cells was further decreased. Conversely, knockdown of SIN3A partially reversed the effect induced by HDM: the proportion of Th17 cells was significantly lower than that in the HDM group, while the proportion of Treg cells was restored to or even higher than that in the HDM group.
In conclusion, in the context of allergic rhinitis, SIN3A in nasal mucosal epithelial cells may be an important molecule regulating the differentiation fate of T cells. Its high expression can promote the differentiation of naive T cells into pro-inflammatory Th17 cells and simultaneously inhibit the differentiation of anti-inflammatory Treg cells.
Discussion
As a chronic condition impacting 10–40% of the global populace, AR currently lacks a comprehensive solution, detrimentally influencing patients’ quality of life and imposing significant economic and societal burdens. Consequently, medical specialists worldwide are persistently examining the pathophysiological processes of AR. Numerous studies have indicated that histone modification is intricately linked to the pathogenesis of allergic disorders,7–9 although the impact of histone modification dysfunction on the pathogenesis of allergic rhinitis remains unexplored. This work uses bioinformatics to identify the principal genes related to AR and histone modification, subsequently investigating the specific pathways linked to immune infiltration, ultimately concluding that SIN3A may serve as a crucial immune regulatory protein in AR.
Histone modification is a primary mechanism for regulating epigenetics, which pertains to inheritable epigenetic information distinct from the DNA sequence. Nucleosomes, the fundamental structural units of chromatin, consist of histones and the DNA that coils around them. The post-translational modification processes, including methylation, phosphorylation, and acetylation of the N- and C-termini of histones associated with DNA, can alter the charge and structure of their tails, subsequently affecting chromatin configuration and gene expression. Consequently, histone modification is pivotal in numerous cellular biological processes.13–15
Prior research has demonstrated that histone alterations have a role in the progression of several allergy disorders. Increased histone H2 acetylation in airway smooth muscle cells in asthma results in the deregulation of CXCL8 transcriptional regulation, contributing to the pathophysiology of asthma.7 Research indicates that airway smooth muscle cells in asthmatics exhibit an elevated secretion of vascular endothelial growth factor (VEGF) compared to the general population, and methylation of histone H3 at the H3K9me3 locus may result in the removal of the VEGF promoter inhibitory complex.8 Literature indicates that aberrant histone modifications in peripheral blood mononuclear cells may play a role in the pathophysiological mechanisms of allergic purpura.9 Consequently, investigations into histone modifications may represent a novel approach to the management of allergic rhinitis. Nonetheless, the function of histone modification in allergic rhinitis remains unexamined. This work aims to investigate the role of histone modification in allergic rhinitis and to examine the interaction between histone modification and the immune system, with the objective of elucidating its significant involvement in the disease mechanism.
Our data indicate that SIN3A is the sole histone modification-related gene exhibiting differential expression between AR and healthy populations. Sin3A is a constituent of the SIN3 family of transcriptional regulatory proteins, specifically member A. Sin3A, a pivotal component of the multiprotein chromatin modification complex, exhibits co-inhibitor, co-activator, and transcription factor characteristics, interacting with structural domains of histone deacetylases via class I histone deacetylases, thereby influencing chromatin architecture and modulating the expression of functional genes.16–18 Research has demonstrated that SIN3A may play a role in the fibrosis of asthmatic airways by influencing connective tissue growth factors.19 Kozlik et al20 discovered that SIN3A exhibits variable expression in eosinophilic asthmatics and plays a significant role in most transcriptional responses to hypoxia. Literature indicates that in atopic dermatitis, SIN3A and STAT3 form a signaling pathway crucial in the pathogenesis of TSLP-mediated atopic dermatitis.21 All aforementioned research indicates that SIN3A has a role in the pathophysiological mechanisms of allergy-related diseases via histone modification, suggesting that SIN3A may be a critical element in the pathogenesis of allergic rhinitis due to histone modification abnormalities.
SIN3A is a primary regulator of STAT3 transcriptional activity,22 and STAT3 is crucial in developing of Th17 cells.23 Perucho et al24 discovered that SIN3A facilitates Th17 development through STAT3 in vitro studies, and the inhibition of SIN3A attenuates the pro-inflammatory effects of TH17, which is advantageous for immunomodulatory functions. The prevalence of Th17 cells in allergic rhinitis patients is markedly elevated compared to healthy individuals, and the release of pro-inflammatory cytokines by Th17 cells is crucial in the inflammatory response25; furthermore, the dysregulation of Th17 cells constitutes a pathogenic mechanism of allergic rhinitis. Consequently, SIN3A is expected to be instrumental in the pathogenesis of AR, particularly concerning the facilitation of Th17 cell development.
This study employed Western blotting, qRT-PCR, and immunofluorescence to confirm the differential expression of SIN3A in AR patients using nasal mucosa tissue specimens from two groups. The findings indicated that SIN3A exhibited elevated expression levels in the nasal mucosa of patients with allergic rhinitis. The AUC value of 0.87 for the SIN3A gene in the aforementioned article indicates a strong differential diagnostic value, presenting a novel potential target for diagnosing and AR patients.
This study screened out multiple histone modification genes including HDAC1 and their interacting drugs through bioinformatics analysis (Figure 4b), suggesting that targeting epigenetic regulation might be a new direction for AR treatment. Among them, the broad-spectrum HDAC inhibitor sodium butyrate (NaB) has been confirmed in multiple studies26 to effectively alleviate allergic symptoms and mucosal inflammation in AR animal models by restoring histone acetylation levels (such as H3-AcK9), reducing HDAC1/8 expression, and rebalancing Th1/Th2/Th17 and Treg. However, such pan-HDAC inhibitors lack gene specificity and may cause pleiotropic biological effects and potential side effects. In contrast, SIN3A, a specific transcriptional co-repressor identified in this study, may more precisely focus on regulating key immune pathways (such as Th17/Treg differentiation). Although no reported clinical drugs targeting SIN3A have been retrieved through public drug databases (such as DGIdb) at present, this makes it difficult to directly compare the efficacy of SIN3A inhibitors with that of NaB. SIN3A inhibitors have been formulated for in vivo application, enhancing the potential for targeting SIN3A in forthcoming AR therapy.27
To elucidate the function of SIN3A in the AR immune milieu, we conducted single-cell annotation analysis of immune cells from AR patients before and following SLIT treatment. The findings indicated that SIN3A exhibited considerable differential expression in CD4+ naïve T cells and Treg cells in pre-treatment AR patients. This discovery indicates that SIN3A may assume distinct functions in these two cell types, hence influencing the pathophysiology of AR. After SLIT treatment, the disparity in SIN3A expression between CD4+ naïve T cells and Treg cells in AR patients diminished. Treg cells differentiate from CD4+ naïve T cells and function to regulate the cessation of immune system overactivity. This indicates that SLIT therapy may have effectively addressed AR by modulating the expression of SIN3A, hence facilitating the differentiation of CD4+ naïve T cells into Treg cells.
The imbalance between Treg and Th17 cells constitutes a pathogenic mechanism of allergic rhinitis. However, our study did not reveal any variation in the expression of SIN3A in Th17 cells among the allergic rhinitis population pre-and post-treatment. This lack of difference may stem from the limited sample size and the selection criteria, which focused solely on patients undergoing sublingual immunotherapy, without including healthy controls or a broader analysis of the allergic rhinitis population, thereby failing to represent the overall condition of AR patients comprehensively. To investigate the specific mechanism further, common immunological checkpoints and human leukocyte antigens were chosen for correlation analysis, revealing a substantial positive association between SIN3A and ADORA2A, HLA-C, HLA-F, and HLA-DQB2. Elevated levels of HLA-C have been strongly associated with illness duration in AR patients, as indicated in a prior study.28 Additionally, HLA-DQB2, a highly conserved HLA molecule, has been demonstrated in the literature to facilitate antigen presentation and T cell stimulation.29 Moreover, HLA-F serves as a distinct marker on the surface of activated T cells.30 ADORA2A has been confirmed as a crucial element in sustaining CD4+ naïve T cell populations31 and plays a significant role in the modulation of T cell and NK cell activation by Treg cells.32
Although previous studies have implicated SIN3A in immune dysregulation in diseases such as asthma and atopic dermatitis, direct functional evidence of its role in AR—particularly in regulating T-cell differentiation—has been lacking. Our study provides initial experimental support for this mechanism in an AR-relevant context: through in vitro knockdown and overexpression experiments in human nasal epithelial cells co-cultured with naïve T cells, we demonstrated that SIN3A expression significantly skews CD4⁺ T cell differentiation toward a Th17 phenotype while suppressing Treg generation. These findings position SIN3A as a potential epigenetic regulator of mucosal immune balance in AR.
Although this study confirmed the upregulated expression of SIN3A in the nasal mucosa of AR patients and revealed through functional experiments that SIN3A participates in the pathological process of AR by promoting the differentiation of naive T cells into Th17 cells while inhibiting the differentiation of Treg cells, this regulatory mechanism has not yet been further verified in animal models. In addition, the specific regulatory mechanisms between SIN3A and its positively correlated molecules (such as ADORA2A, HLA-C, HLA-F, and HLA-DQB2) have not been deeply explored. The potential functions of these molecules in immune regulation and their synergistic effects with SIN3A will be the focus of future research. Subsequent work will be dedicated to clarifying how SIN3A regulates the differentiation fate of CD4⁺ T cells through interactions with these molecules and the detailed molecular mechanisms underlying the occurrence and development of AR.
This study has several shortcomings, including: the fact that Serologic testing is a primary diagnostic tool for AR patients, and SIN3A, a crucial gene for histone modification, has not been identified in the serum expression of these patients. The nasal mucosal tissues analyzed in this study were sourced from surgically treated allergic rhinitis patients, whose clinical symptoms are more pronounced than those of outpatients, potentially introducing bias into the experimental outcomes. The sample size of the subgroups in this study (for example, n = 12 in each group of the sublingual immunotherapy response group and non-response group and in the nasal mucosa tissue validation) may limit the generalizability of the research results and the statistical power to detect subtle transcriptomic changes. Although WGCNA has been successfully applied to similar-scale exploratory transcriptomic studies.33 In future research, we plan to validate the above findings using larger and independent cohorts to further confirm the robustness and clinical relevance of the results of this study.
While the role of histone modification in the pathophysiology of several airway allergic illnesses has been examined and highlighted, its significance in allergic rhinitis remains unexplored and undiscussed. This work concludes that SIN3A is a crucial gene associated with histone modification in allergic rhinitis; nevertheless, the understanding of its precise mechanism remains restricted, necessitating further research to elucidate the pathogenesis of SIN3A in allergic rhinitis.
Conclusion
In conclusion, we identified SIN3A as a pivotal gene associated with histone modification that exhibits differential expression between the AR population and healthy individuals, potentially influencing the progression of AR by modulating the balance of Th17 and Treg cell, Our findings establish SIN3A as a potential candidate for future therapeutic strategies against allergic rhinitis.
Acknowledgments
We would like to thank the GEO (GSE50223, GSE200107) networks for providing the data.
Ethics Approval
This study was approved by the human study ethics committees at The Second Xiangya Hospital of Central South University (approval no.LYF2022222). Informed consent was obtained from both adult patients and their legal guardians, who are minors, prior to their participation in the study. lt was conducted in accordance with the principles of the Declaration Helsinki.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no competing interests.
References
1. Eifan AO, Durham SR. Pathogenesis of rhinitis. Clin Exp Allergy. 2016;46(9):1139–1151. doi:10.1111/cea.12780
2. Bousquet J, Anto JM, Bachert C, et al. Allergic rhinitis. Nat Rev Dis Primers. 2020;6(1):95. doi:10.1038/s41572-020-00227-0
3. Siddiqui ZA, Walker A, Pirwani MM, et al. Allergic rhinitis: diagnosis and management. Br J Hosp Med. 2022;83(2):1–9. doi:10.12968/hmed.2021.0570
4. Hoyte FCL, Nelson HS. Recent advances in allergic rhinitis. F1000Res. 2018;7:1333. doi:10.12688/f1000research.15367.1
5. Leth-MØLLER KB, Skaaby T, Linneberg A. Allergic rhinitis and allergic sensitisation are still increasing among Danish adults. Allergy. 2020;75(3):660–668. doi:10.1111/all.14046
6. Wang XD, Zheng M, Lou HF, et al. An increased prevalence of self-reported allergic rhinitis in major Chinese cities from 2005 to 2011. Allergy. 2016;71(8):1170–1180. doi:10.1111/all.12874
7. Clifford RL, Patel JK, John AE, et al. CXCL8 histone H3 acetylation is dysfunctional in airway smooth muscle in asthma: regulation by BET. Am J Physiol Lung Cell Mol Physiol. 2015;308(9):L962–72. doi:10.1152/ajplung.00021.2015
8. Clifford RL, John AE, Brightling CE, et al. Abnormal histone methylation is responsible for increased vascular endothelial growth factor 165a secretion from airway smooth muscle cells in asthma. J Immunol. 2012;189(2):819–831. doi:10.4049/jimmunol.1103641
9. Luo S, Liang G, Zhang P, et al. Aberrant histone modifications in peripheral blood mononuclear cells from patients with Henoch-Schönlein purpura. Clin Immunol. 2013;146(3):165–175. doi:10.1016/j.clim.2012.12.009
10. Li L, Wen X, Gong Y, et al. HMGN2 and Histone H1.2: potential targets of a novel probiotic mixture for seasonal allergic rhiniti. Front Microbiol. 2023;14:1202858. doi:10.3389/fmicb.2023.1202858
11. Mesdaghi M, Vodjgani M, Salehi E, et al. Natural killer cells in allergic rhinitis patients and nonatopic controls. Int Arch Allergy Immunol. 2010;153(3):234–238. doi:10.1159/000314363
12. Kim JH, Jang YJ. Role of natural killer cells in airway inflammation. Allergy Asthma Immunol Res. 2018;10(5):448–456. doi:10.4168/aair.2018.10.5.448
13. Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293(5532):1074–1080. doi:10.1126/science.1063127
14. Strahl BD, Allis CD. The language of covalent histone modifications. Nature. 2000;403(6765):41–45. doi:10.1038/47412
15. Zhang Y, Sun CZ, Jia CJ, et al. Overview of Histone Modification. Adv Exp Med Biol. 2021;1283:1–16.
16. Chaubal A, Pile LA. Same agent, different messages: insight into transcriptional regulation by SIN3 isoforms. Epigenet Chromatin. 2018;11(1):17. doi:10.1186/s13072-018-0188-y
17. Alland L, Muhle R, Hou H, et al. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature. 1997;387(6628):49–55. doi:10.1038/387049a0
18. Nagy L, Kao HY, Chakravarti D, et al. Nuclear receptor repression mediated by a complex containing SMRT, mSin3A, and histone deacetylase. Cell. 1997;89(3):373–380. doi:10.1016/S0092-8674(00)80218-4
19. Hua HS, Wen HC, Lee HS, et al. Endothelin-1 induces connective tissue growth factor expression in human lung fibroblasts by disrupting HDAC2/Sin3A/MeCP2 corepressor complex. J Biomed Sci. 2023;30(1):40. doi:10.1186/s12929-023-00931-5
20. Kozlik-Siwiec P, Buregwa-Czuma S, Zawlik I, et al. Co-Expression Analysis of Airway Epithelial Transcriptome in Asthma Patients with Eosinophilic vs. Non-Eosinophilic Airway Infiltration. Int J Mol Sci. 2023;24(4):3789. doi:10.3390/ijms24043789
21. Lee H, RYU WI, Kim HJ, et al. TSLP down-regulates S100A7 and ß-Defensin 2 Via the JAK2/STAT3-dependent mechanism. J Invest Dermatol. 2016;136(12):2427–2435. doi:10.1016/j.jid.2016.07.027
22. Icardi L, Mori R, Gesellchen V, et al. The Sin3a repressor complex is a master regulator of STAT transcriptional activity. Proc Natl Acad Sci U S A. 2012;109(30):12058–12063. doi:10.1073/pnas.1206458109
23. Mathur AN, Chang HC, Zisoulis DG, et al. Stat3 and Stat4 direct development of IL-17-secreting Th cells. J Immunol. 2007;178(8):4901–4907. doi:10.4049/jimmunol.178.8.4901
24. Perucho L, Icardi L, Di Simone E, et al. The transcriptional regulator Sin3A balances IL-17A and Foxp3 expression in primary CD4 T cells. EMBO Rep. 2023;24(5):e55326. doi:10.15252/embr.202255326
25. Oboki K, Ohno T, Saito H, et al. Th17 and allergy. Allergol Int. 2008;57(2):121–134. doi:10.2332/allergolint.R-07-160
26. Wang J, Cui M, Sun F, et al. HDAC inhibitor sodium butyrate prevents allergic rhinitis and alters lncRNA and mRNA expression profiles in the nasal mucosa of mice. Int J Mol Med. 2020;45(4):1150–1162. doi:10.3892/ijmm.2020.4489
27. Kwon YJ, Petrie K, Leibovitch BA, et al. Selective inhibition of SIN3 corepressor with avermectins as a novel therapeutic strategy in triple-negative breast cancer. Mol Cancer Ther. 2015;14(8):1824–1836. doi:10.1158/1535-7163.MCT-14-0980-T
28. Ciprandi G, Colombo BM, Contini P, et al. Soluble HLA-G and HLA-A,-B,-C serum levels in patients with allergic rhinitis. Allergy. 2008;63(10):1335–1338. doi:10.1111/j.1398-9995.2008.01741.x
29. Lenormand C, Bausinger H, Gross F. HLA-DQA2 and HLA-DQB2 genes are specifically expressed in human Langerhans cells and encode a new HLA class II molecule. J Immunol. 2012;188(8):3903–3911. doi:10.4049/jimmunol.1103048
30. Lee N, A Ishitani, Geraghty DE, et al. HLA-F is a surface marker on activated lymphocytes. Eur J Immunol. 2010;40(8):2308–2318. doi:10.1002/eji.201040348
31. Cekic C, D Sag, Day YJ, et al. Extracellular adenosine regulates naive T cell development and peripheral maintenance. J Exp Med. 2013;210(12):2693–2706. doi:10.1084/jem.20130249
32. Häusler SF, Montalbán Del Barrio I, Strohschein J, et al. Ectonucleotidases CD39 and CD73 on OvCA cells are potent adenosine-generating enzymes responsible for adenosine receptor 2A-dependent suppression of T cell function and NK cell cytotoxicity. Cancer Immunol Immunother. 2011;60(10):1405–1418. doi:10.1007/s00262-011-1040-4
33. Oldham MC, Konopka G, Iwamoto K, et al. Functional organization of the transcriptome in human brain. Nat Neurosci. 2008;11(11):1271–1282. doi:10.1038/nn.2207
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.