Back to Journals » OncoTargets and Therapy » Volume 9
Identification of HSPA8 as a candidate biomarker for endometrial carcinoma by using iTRAQ-based proteomic analysis
Authors Shan N ![]() , Zhou W, Zhang S, Zhang Y
, Zhou W, Zhang S, Zhang Y
Received 10 October 2015
Accepted for publication 10 March 2016
Published 13 April 2016 Volume 2016:9 Pages 2169—2179
DOI https://doi.org/10.2147/OTT.S97983
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Nianchun Shan,1 Wei Zhou,2 Shufen Zhang,1 Yu Zhang1
1Department of Obstetric and Gynecology, 2Health Management Center, Xiangya Hospital, Central South University, Changsha, Hunan, People’s Republic of China
Abstract: Although there are advances in diagnostic, predictive, and therapeutic strategies, discovering protein biomarker for early detection is required for improving the survival rate of the patients with endometrial carcinoma. In this study, we identify proteins that are differentially expressed between the Stage I endometrial carcinoma and the normal pericarcinous tissues by using isobaric tags for relative and absolute quantitation (iTRAQ)-based proteomic analysis. Totally, we screened 1,266 proteins. Among them, 103 proteins were significantly overexpressed, and 30 were significantly downexpressed in endometrial carcinoma. Using the bioinformatics analysis, we identified a list of proteins that might be closely associated with endometrial carcinoma, including CCT7, HSPA8, PCBP2, LONP1, PFN1, and EEF2. We validated the gene overexpression of these molecules in the endometrial carcinoma tissues and found that HSPA8 was most significantly upregulated. We further validated the overexpression of HSPA8 by using immunoblot analysis. Then, HSPA8 siRNA was transferred into the endometrial cancer cells RL-95-2 and HEC-1B. The depletion of HSPA8 siRNAs significantly reduced cell proliferation, promoted cell apoptosis, and suppressed cell growth in both cell lines. Taken together, HSPA8 plays a vital role in the development of endometrial carcinoma. HSPA8 is a candidate biomarker for early diagnosis and therapy of Stage I endometrial carcinoma.
Keywords: iTRAQ, HSPA8, endometrial carcinoma, RL-95-2 cells
Introduction
Endometrial cancer is the fourth most common gynecological malignancy. Its incidence rates and death rate are alarmingly increasing year by year.1 Over the past 10 years, the death rate of this cancer has increased by 227%. A total of 70%–80% of patients with endometrial cancer are type I endometrioid histology. Women who develop these tumors are typically perimenopausal or postmenopausal, indicating these tumors have arisen from persistent unopposed estrogen stimulation.2,3 A list of risk factors, such as nulliparity, hyperlipidemia, obesity, insulin resistance and diabetes mellitus, hypertension, late-onset menopause, and polycystic ovarian syndrome, associated with the development of endometrial cancer.4–6 Early-stage patients with endometrial cancer have good prognosis and high cure rate.7 However, patients with advanced stage symptoms including metastasis may have poor prognosis and improper monitoring due to the obstacle for predicting survival of endometrial carcinoma. Therein comes the need for biomarker discovery. In addition, it requires a global view of gene activity around the development process of endometrial cancer. In spite of extensive description about the development of endometrial cancer, the molecular and pathological mechanisms governing the strictly regulated process remain to be elucidated.
The development of endometrial cancer is controlled coordinately by the functional molecules that construct the regulation networks. The knowledge about those networks could help us to obtain candidates for diagnosis, prediction, and therapy of endometrial cancer.8 System biology, such as bottom-up profiling of proteins, provides powerful approach for analyzing the functional genes in the development of cancer. The changes in the expression of transcriptome profiles could be examined by the analysis of digital gene expression with high throughput tag sequencing.9 Nowadays, many studies preferred to analyze the transcriptional profiling in the development of cancer. In fact, the gene should be translated to protein that ultimately controlled the biological processes and was the essential part of regulation network. Thus, the use of proteomics in cancer study, in which global protein information is provided, could help us to clarify the molecular mechanism in cancer development.10 The technique of isobaric tags for relative and absolute quantitation (iTRAQ) combined with liquid chromatography-tandem mass spectrometric (LC-MS/MS) is one of the most widely used proteomic approaches because of the detailed protein expression profiles and its high resolution with reducing experimental error.11 iTRAQ-based proteomics has also been applied in biomarker discovery for cancer patients.10,12
In this study, we aimed to quantitatively assess the protein expression in the development of endometrial cancer and to identify proteins that are differentially expressed between endometrial carcinoma and pericarcinous tissues obtained from patient with Stage I endometrial cancer by using iTRAQ combined with LC-MS/MS. We identified a list of proteins that might be closely associated with the endometrial carcinoma, including CCT7, HSPA8, PCBP2, LONP1, PFN1, and EEF2. Then, the expression of these molecules in mRNA level was validated by quantitative real-time polymerase chain reaction (qRT-PCR). And we further detected the protein level of HSPA8 that was most significantly changed in mRNA level. Results showed that HSPA8 was significantly overexpressed in endometrial carcinoma by Western blot. The role of HSPA8 in the development of endometrial cancer was further assessed by the detection of cell proliferation, apoptosis, and cell cycle in HSPA8 siRNAs-transfected endometrial cancer cells RL-95-2 and HEC-1B. The findings of this research suggested that HSPA8 is a candidate biomarker for early diagnosis and therapy of endometrial carcinoma.
Material and methods
Tissue sample collection
This study was approved by the institutional human ethics committee of Xiangya Hospital, Central South University, People’s Republic of China. We collected tumor tissue samples and adjacent normal tissues from ten patients with Stage I endometrial carcinoma. All patients signed the informed consents. Patients who had history of other solid tumors, radiotherapy, chemotherapy, or other anticancer therapies prior to surgery were excluded. All samples were validated by expert pathologists. The collected samples were stored at −80°C.
Sample preparation for proteomics and iTRAQ labeling
Endometrial carcinoma and normal tissue (10 mg) were homogenized and sonicated using 0.5% sodium dodecyl sulfate (SDS) with a cell disperser. The cell debris was eliminated by centrifugation at 15,000 rpm for 30 minutes at 4°C. Then, the supernatant was collected, and the protein concentration was measured by using the Bradford assay. The trypsin digestion and labeling were carried out with 4-Plex iTRAQ reagents (iTRAQ Reagents Multiplex kit; Thermo Fisher Scientific, Waltham, MA, USA). In brief, each protein sample (200 μg) was digested with 50 μL of trypsin solution at 37°C overnight. The iTRAQ proteomics was performed using a 4-plex procedure.13 Then, LC-MS/MS analysis was performed as previously described.13
Bioinformatic analysis
The raw acquired iTRAQ data files were processed by ProteinPilot Version 3.0 (AB Sciex, Redwood City, CA, USA), and the proteins were identified by using the raw MS data.13 For iTRAQ quantitation, the peptide was selected automatically by ProteinPilot with Pro Group™ algorithm, and the reporter peak area, error factor, and P-value were calculated. For each protein, the false discovery rate (FDR) was estimated.13 If the iTRAQ ratios (normal pericartissue/endometrial carcinoma tissue) was <0.67 or >1.5 in the endometrial carcinoma tissue with the normal pericarcinous tissue (P<0.05), the protein was considered to be significantly differentially expressed.
Then, by using the Bioconductor 3.0, Gene Ontology enrichment analysis was performed to analyze the functions of the differentially expressed proteins, and three major categories were included: cellular component, molecular function, and biological process.14 A FDR <0.05 was considered as a threshold for significant enrichment of the protein sets. In addition, enriched Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway was identified by significantly enriched pathways or signal transduction pathways associated with the development of endometrial carcinoma, and testing each pathway for enrichment in significantly differently expressed proteins as implemented by the KOBAS (2.0) software. Pathways were deemed to be enriched if the enrichment over background was at least twofold and the P-value <0.05. Selected molecules associated with endometrial carcinoma were further identified by LC-MS/MS spectra.
Cell culture
Endometrial cancer cell lines RL-95-2 and HEC-1B that express both the estrogen and the progesterone receptors were cultured in Dulbecco’s Modified Eagle’s Medium supplemented with 5% fetal bovine serum, 300 mmol/L L-glutamine, 5 μg/mL bovine insulin, 10,000 units/mL penicillin, and 10,000 μg/mL streptomycin under 5% CO2. This study was approved by the institutional human ethics committee of Xiangya Hospital, Central South University.
Cell transfection
The role of HSPA8 in endometrial carcinoma cells RL-95-2 and HEC-1B was examined using siRNA-mediated knockdown of HSPA8. For screening HSPA8 siRNA, 50 μM, 100 μM, and 200 μM HSPA8 siRNA (HSPA8-Homo-768: 5′-GCUGGUCUCAAUGUACUUATT-3′, 5′-UAAGUACAUUGAGACCAGCTT-3′; HSPA8-Homo-1112: 5′-GGCCAGUAUUGAGAUCGAUTT-3′, 5′-AUCGAUCUCAAUACUGGCCTT-3′; HSPA8-Homo-1509: 5′-GUCCUCAUCAAGCGUAAUATT-3′, 5′-UAUUACGCUUGAUGAGGACTT-3′) or nontargeting negative control (5′-UUCUCCGAACGUGUCACGUTT-3′, 5′-ACGUGACACGUUCGGAGAATT-3′; Santa Cruz Biotechnology Inc., Dallas, TX, USA), and DharmaFECT reagent (Thermo Fisher Scientific) were transfected into RL-95-2 cells according to the manufacturer’s instructions. After 48 hours, the expression of HSPA8 was confirmed by qRT-PCR, and the best HSPA8 siRNA was selected. Then, both RL-95-2 and HEC-1B cells were transfected with HSPA8 siRNA, the expression of HSPA8 was confirmed by Western blotting, and cell proliferation, cell cycle, and apoptosis were performed.
Quantitative real-time polymerase chain reaction
Total RNA was extracted using TRIZOL (Thermo Fisher Scientific) and subjected to qRT-PCR using the following primers: 5′-CATTCTCAACAAGCTGCGGG-3′ (forward) and 5′-TTGATCCGCACCATAGCTGG-3′ (reverse) for CCT7, 5′-ACTCCAAGCTATGTCGCCTTT-3′ (forward) and 5′-TGGCATCAAAAACTGTGTTGGT-3′ (reverse) for HSPA8, 5′-GCGCAGATCAAAATTGCGAAC-3′ (forward) and 5′-ATATTGAGCCAGGCTAATGCTG-3′ (reverse) for PCBP2, 5′-AGCCTTATGTCGGCGTCTTTC-3′ (forward) and 5′-CGTCCCCGTGTGGTAGATTTC-3′ (reverse) for LONP1, 5′-ATGGATCTTCGTACCAAGAGCA-3′ (forward) and 5′-CCATCAGCAGGACTAGCGTC-3′ (reverse) for PFN1, 5′-CATTGCCGAGCGCATCAAG-3′ (forward) and 5′-CTCGCCGTAGGTGGAGATGA-3′ (reverse) for EEF2, and 5′-ACACCCACTCCTCCACCTTT-3′ (forward) and 5′-TTACTCCTTGGAGGCCATGT-3′ (reverse) for β-actin. Gene expression was normalized to the level of β-actin using the relative ΔΔCT method and is shown as relative expression to control.
Western blotting
Sample or transfected cells were lysed on ice for 10 minutes in 50 mM Tris–HCl (pH 7.5), 10% glycerol, 2% SDS, 0.1 M dithiothreitol, and 10 mM phenylmethylsulfonyl fluoride. Proteins were separated on 10% SDS–polyacrylamide gels and electroblotted onto a nitrocellulose membrane in 25 mM Tris base and 190 mM glycine at 50 V for 3 hours at 4°C. To detect the expression of HSPA8, blots were blocked with 5% dried milk in Tween 20–TBS for 1 hour and incubated with mouse monoclonal antibodies against HSPA8 (1:1,000; Santa Cruz Biotechnology Inc.) overnight. After washing with TBS, blots were incubated with horseradish peroxidase-coupled secondary antimouse antibodies for 1 hour. Then proteins were detected using enhanced chemiluminescence (Beyotime, Shanghai, People’s Republic of China).
Cell proliferation assay
Cell proliferation was determined by a MTT assay (Sigma-Aldrich Co., St Louis, MO, USA). Briefly, RL-95-2 cells (8×103) were plated onto 96-well plates for 24 hours and then cultivated for 24 hours, 48 hours, and 72 hours. By metabolic conversion of the dye MTT, viable cell densities were determined. Then, the results of MTT assay were read at 595 nm.
Cell apoptosis
Cell apoptosis was determined by Annexin V assay. After transfection and/or lidocaine treatment, cells were collected, washed, and suspended in Annexin V-binding buffer. FITC-conjugated Annexin V and propidium iodide (PI; Beyotime) were added to the cells successively. After incubation, Annexin V-binding buffer was added, and cells were analyzed by a FACScan (Becton Dickinson, BD Biosciences, San Jose, CA, USA) flow cytometry.
Cell cycle analysis
After transfection and/or lidocaine treatment, cells were harvested after trypsinization. Then, cells were rinsed three times with buffer solution with adjusted concentration 1×106 cells/mL and prepared using CycleTEST PLUS DNA Reagent Kit (Becton Dickinson) according to the manufacturer’s instruction. Cell cycle status was analyzed by flow cytometer using PI. The PI fluorescence intensity of 10,000 cells was measured for each sample.
Statistical analysis
All results are expressed as the mean ± standard deviation (SD) from three independent experiments. Statistical significance between groups was determined using SPSS 18.0 (SPSS Inc., Chicago, IL, USA) with one-way analysis of variance or an unpaired Student’s t-test. P<0.05 was considered to be statistically significant.
Results
Identification of differentially expressed proteins between endometrial carcinoma and pericarcinous tissue
Proteins from endometrial carcinoma patients were quantified using iTRAQ and LC-MS/MS analysis. In this study, we identified 1,266 proteins (with FDR <0.01). Then, the proteins that were significantly expressed between the endometrial carcinoma and the pericarcinous tissue were screened. The thresholds of iTRAQ ratios (normal pericarcinous tissue:endometrial carcinoma tissue) were not <1.5 or not >0.67, which indicated lower or higher expression of protein in carcinoma tissue than in normal pericarcinous tissue. A total of 133 proteins that were differentially expressed were further identified. Among them, 103 proteins were upregulated in the endometrial carcinoma tissues and 30 were downregulated.
To understand the functions of differentially expressed proteins, Gene Ontology enrichment analysis was performed to analyze the functions of these proteins in the following three categories: biological processes, molecular functions, and cellular components (Figure 1). We found six significant biological processes associated with these differentially expressed proteins (Figure 1A), including response to metabolic process (30.8%), cellular process (23.1%), immune system process (15.4%), stimulus (15.4%), biological regulation (7.7%), and localization (7.7%) and nine molecular functions (Figure 1B) associated with them, including binding (31.7%), catalytic activity (36.6%), structural molecule activity (17.1%), nucleic acid-binding transcription factor activity (3.7%), translation regulator activity (3.7%), enzyme regulator activity (2.4%), transporter activity (2.4%), receptor activity (1.2%), and antioxidant activity (1.2%). Cell part (50%) and organelle (50%) were the enriched cellular components (Figure 1C).
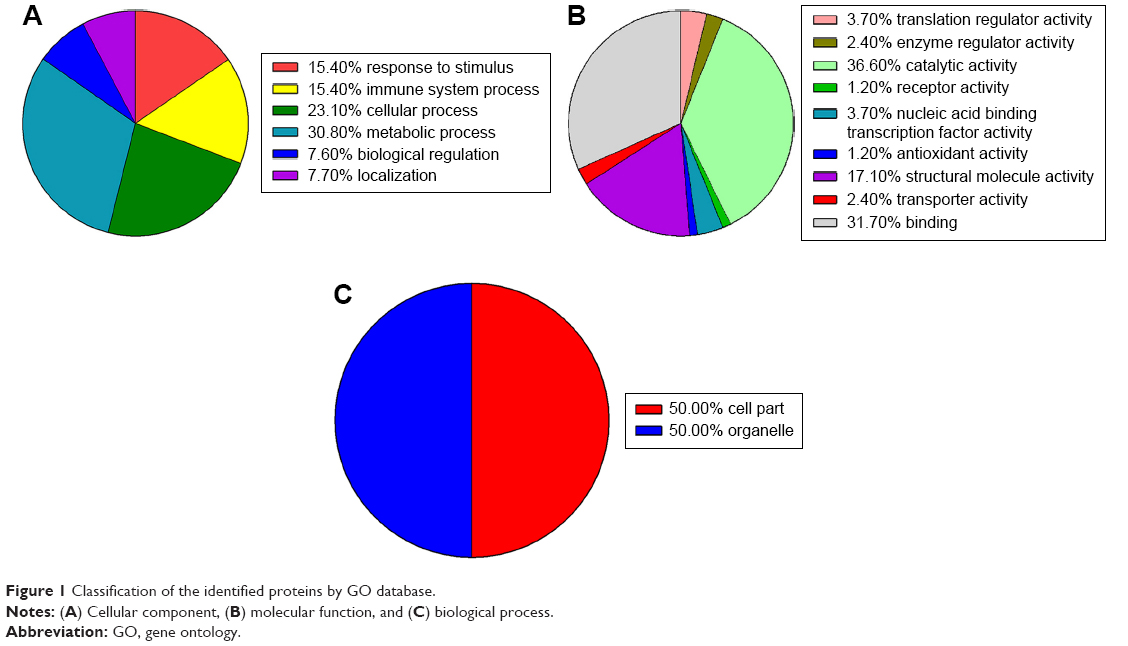 | Figure 1 Classification of the identified proteins by GO database. |
The differentially expressed proteins in endometrial carcinoma were further assigned to 14 KEGG pathways (P<0.05; Table 1), which include pathways for legionellosis, endocytosis, pathogenic Escherichia coli infection, shigellosis, glycolysis/gluconeogenesis, cardiac muscle contraction, fructose and mannose metabolism, African trypanosomiasis, bacterial invasion of epithelial cells, hypertrophic cardiomyopathy, viral carcinogenesis, phagosome, protein processing in endoplasmic reticulum, and amino sugar and nucleotide sugar metabolism. Among them, selected molecules associated with endometrial carcinoma were further identified by LC-MS/MS spectra, including CCT7, heat shock 70 kDa protein 8 (HSPA8), PCBP2, LONP1, PFN1 and EEF2, which are listed in Table 2. The representative LC-MS/MS spectra of the molecule HSPA8 has been shown in Figure 2.
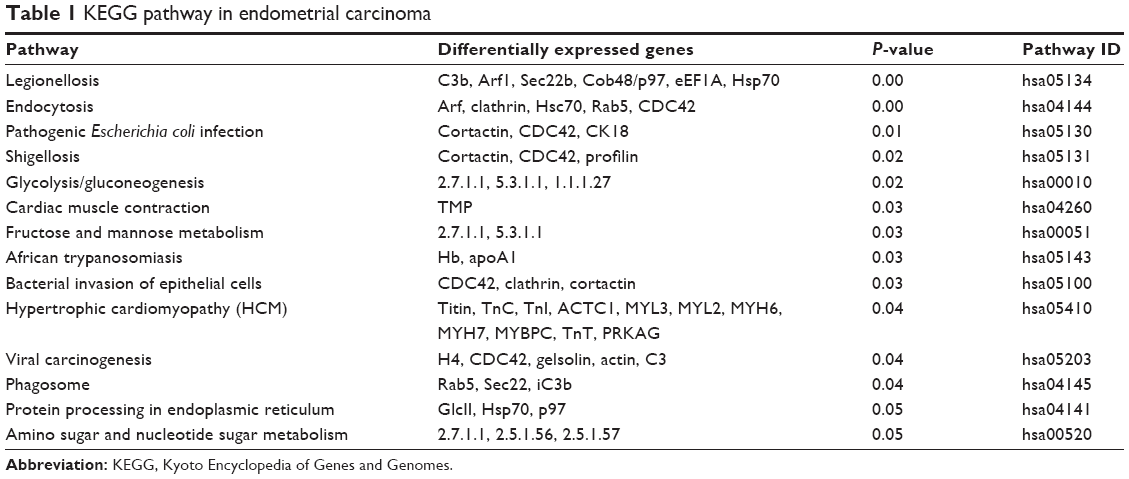 | Table 1 KEGG pathway in endometrial carcinoma |
 | Table 2 A partial list of novel molecules found to be highly expressed in the endometrial carcinoma tissue proteome |
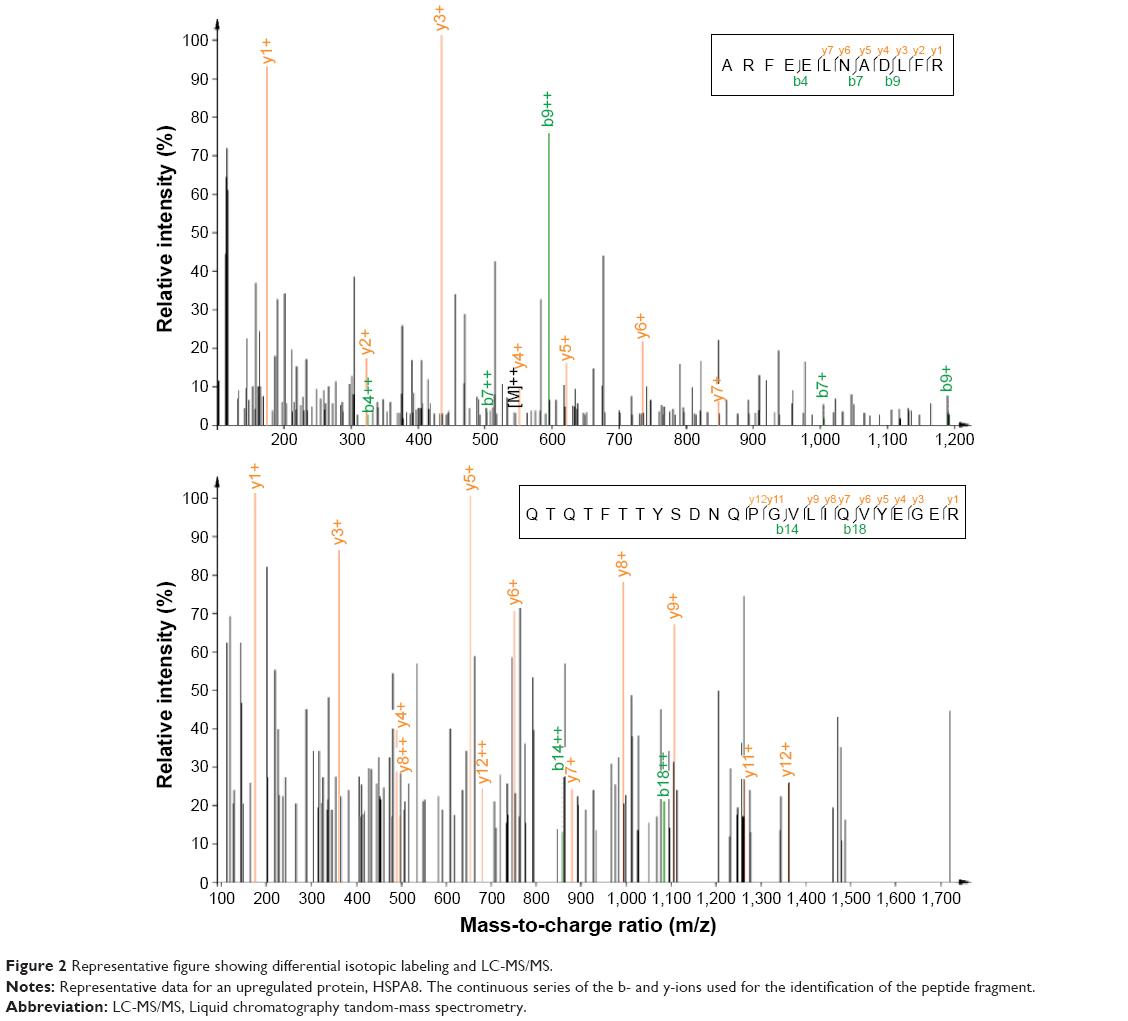 | Figure 2 Representative figure showing differential isotopic labeling and LC-MS/MS. |
Validation of identified proteins by qRT-PCR and Western blot
From the proteomic data with iTRAQ and LC-MS/MS, we identified a list of proteins. We further validated them by qRT-PCR. Our results revealed that the mRNA expression of CCT7 (Figure 3A), HSPA8 (Figure 3B), PCBP2 (Figure 3C), LONP1 (Figure 3D), PFN1 (Figure 3E), and EEF2 (Figure 3F) was significantly higher in endometrial carcinoma tissues than in the normal pericarcinous tissue (n=10). The upregulation of these molecules in endometrial carcinoma was in concordance with the iTRAQ results. We found that HSPA8 was most significantly upregulated in mRNA levels in endometrial carcinoma tissues (P<0.0001). So, the HSPA8 was selected for further investigation. By Western blot, we also observe the upregulation of HSPA8 in the endometrial carcinoma tissues (Figure 3G).
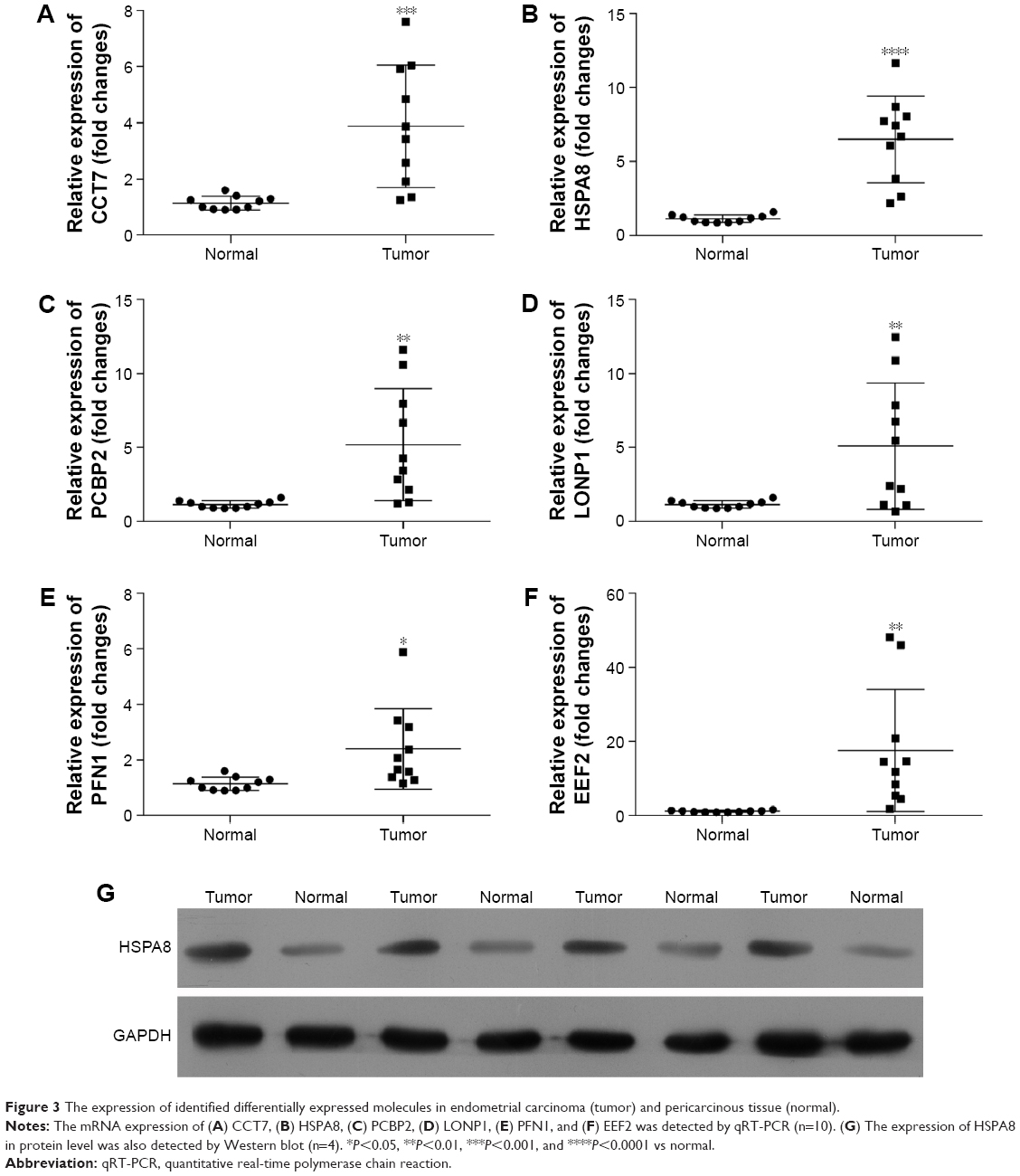 | Figure 3 The expression of identified differentially expressed molecules in endometrial carcinoma (tumor) and pericarcinous tissue (normal). |
Screening of HSPA8 siRNA
The role of HSPA8 in endometrial carcinoma was further investigated. HSPA8 siRNA was screened. Transfection of HSPA8 siRNA (768, 1,112, and 1,509) and nontargeting negative control was performed with different concentrations (50 μM, 100 μM, and 200 μM; Figure 4). HSPA8-768 siRNA suppressed the mRNA expression of HSPA8 significantly at 50 μM, 100 μM, and 200 μM (Figure 4A). By transfection with HSPA8-768 siRNA (100 μM), the expression of HSPA8 in both RL-95-2 and HEC-1B cells was significantly downexpressed (Figure 4B). Then, 100 μM of HSPA8-768 siRNA-transfected RL-95-2 and HEC-1B cells were used in the following experiments.
 | Figure 4 Screening of HSPA8 siRNA for knockdown of HSPA8 in RL-95-2 and HEC-1B cells. |
HSPA8 siRNA reduced the cell proliferation, promoted the cell apoptosis, and suppressed the cell growth
The knockdown of HSPA8 significantly reduced the cell proliferation at 48 hours (Figure 5A). Cell apoptosis including both early apoptosis and later apoptosis was promoted by HSPA8 siRNA (Figure 5B and C). In addition, the knockdown of HSPA8 significantly reduced the cells in S phases (Figure 5D and E), suggesting that HSPA8 plays an important role in the development of endometrial carcinoma. HSPA8 is a candidate for early diagnosis and therapy biomarker for endometrial carcinoma.
 | Figure 5 The proliferation, cell cycle, and apoptosis of HSPA8 siRNA-transfected RL-95-2 and HEC-1B cells. |
Discussion
This study investigated the proteome in the endometrial cancer. In an attempt to have a global view of candidate molecules involved in the development of endometrial cancer, we found 133 differentially expressed proteins by iTRAQ technology. Selected molecules associated with endometrial carcinoma were further identified by LC-MS/MS spectra, including CCT7, HSPA8, PCBP2, LONP1, PFN1, and EEF2. The upregulation of these molecules was further confirmed by qRT-PCR in endometrial carcinoma and pericarcinous tissues obtained from patients with endometrial cancer. Although all the mRNA expressions of CCT7, HSPA8, PCBP2, LONP1, PFN1, and EEF2 in endometrial carcinoma tissues were upregulated (Figure 3), we found that HSPA8 was most significantly upregulated in mRNA levels in endometrial carcinoma tissues (P<0.0001). So, the HSPA8 was selected for further investigation. We also observe the overexpression of HSPA8 in protein level in endometrial carcinoma tissues (Figure 3G).
CCT is a chaperonin involved in cytoskeletal protein folding, such as actin and tubulin,15,16 which plays an important role in cell migration. PCBP2 is multifunctional and it is one of the major cellular PCBPs along with PCBP-1 and hnRNPK. It also functions as a translational coactivator of poliovirus RNA, with PCBP-1 together.17 The depletion of PCBP2 in gastric cancer cells induced apoptosis via upexpression of proapoptotic proteins and downexpression of antiapoptotic proteins.18,19 LONP1 is a mitochondrial matrix protein in the Lon family of ATP-dependent proteases.20 Lon is upregulated specifically in various types of cancers, including lung, colorectal, ovarian, and head-and-neck cancer.21 Upregulation of Lon induces the production of reactive oxygen species via activates the mitogen-activated protein kinase, which provides the survival advantages and adaptation to cancer cell.21 Decrease of human Lon in cervical cancer suppresses cell proliferation and bioenergetics.22 PFN1 regulated actin polymerization in response to extracellular signals, playing important roles in actin dynamics.23 Depletion of PFN1 inhibits the fibronectin-promoted cell proliferation and metastasis, inhibiting gastric cancer progression.24 EEF2 belongs to the GTP-binding translation elongation factor family. EEF2 is essential for protein synthesis. EEF2 gene is upregulated in various cancers and involved in cancer cell growth.25,26
The heat shock protein 70 (HSP70) and its major cochaperones including cytosolic HSPA8 (HSC70), mitochondrial mHSP70 (HSPA9), endoplasmic reticulum BIP (HSPA5), and related HSP110s (HSPHs) may constitute up to 3% in the total protein mass of unstressed human cells.27 The prevailing ATP-hydrolyzing chaperones, conserved HSP70s and HSP90 members, controlled all aspects of cellular proteostasis.28 HSP70 is upregulated in a wide range of human cancers and is mostly considered as a potent buffering system for cellular stress that is required for cancer cell survival.29,30 HSP70 is associated with several phenotypes of tumorigenesis, including proliferation, invasion, and metastasis.31 HSP70 family has antiapoptotic potency and is upregulated in human cancers.32 HSPA8 is a constitutively expressed molecular chaperone and member of the HSP70 family. Many factors could regulate the expression of HSPA8 gene, such as sodium arsenite, organochlorine, azetidine, nickel, and cadmium,33–35 and estrogen and progesterone in the ventromedial hypothalamus.36 HSPA8 is involved in various cellular functions. HSPA8 gene was overexpressed in cancer cells, which was required in cancer cell growth.37 Consistently, depletion of HSPA8 in RL-95-2 and HEC-1B cells suppressed the cell growth. MTT assay provided clear evidence that depletion of HSPA8 obviously decreased the cell proliferation. The cell cycle distribution data showed that the reduced G0/G1 phase percentage and increased S phase percentage resulted from HSPA8 siRNA transfection. In addition, both early apoptosis and later apoptosis were promoted by HSPA8 siRNA transfection.
Conclusion
Our present study provides new insight on molecule changes in endometrial cancer. As validated, CCT7, HSPA8, PCBP2, LONP1, PFN1, and EEF2 are potential biomarkers for endometrial carcinoma tissues. HSPA8 plays an important role in endometrial carcinoma development and is a candidate biomarker for early diagnosis and therapy of endometrial carcinoma.
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65(1):5–29. | ||
Guidozzi F. Estrogen therapy in gynecological cancer survivors. Climacteric. 2013;16(6):611–617. | ||
Marjoribanks J, Farquhar C, Roberts H, Lethaby A. Long term hormone therapy for perimenopausal and postmenopausal women. Cochrane Database Syst Rev. 2012;7:CD004143. | ||
Dashti SG, Chau R, Ouakrim DA, et al. Female hormonal factors and the risk of endometrial cancer in Lynch syndrome. JAMA. 2015;314(1):61–71. | ||
Hale GE, Hughes CL, Cline JM. Endometrial cancer: hormonal factors, the perimenopausal “window of risk,” and isoflavones. J Clin Endocrinol Metab. 2002;87(1):3–15. | ||
Newcomer LM, Newcomb PA, Trentham-Dietz A, Storer BE. Hormonal risk factors for endometrial cancer: modification by cigarette smoking (United States). Cancer Causes Control. 2001;12(9):829–835. | ||
Brandner P, Neis KJ. Diagnosis of endometrial cancer and its precursors. Contrib Gynecol Obstet. 2000;20:27–40. | ||
Fulda S. Targeting extrinsic apoptosis in cancer: challenges and opportunities. Semin Cell Dev Biol. 2015;39:20–25. | ||
Bentley DR. Whole-genome re-sequencing. Curr Opin Genet Dev. 2006;16(6):545–552. | ||
Paul D, Kumar A, Gajbhiye A, Santra MK, Srikanth R. Mass spectrometry-based proteomics in molecular diagnostics: discovery of cancer biomarkers using tissue culture. Biomed Res Int. 2013;2013:783131. | ||
Ross PL, Huang YN, Marchese JN, et al. Multiplexed protein quantitation in Saccharomyces cerevisiae using amine-reactive isobaric tagging reagents. Mol Cell Proteomics. 2004;3(12):1154–1169. | ||
Pastor MD, Nogal A, Molina-Pinelo S, Carnero A, Paz-Ares L. Proteomic biomarkers in lung cancer. Clin Transl Oncol. 2013;15(9):671–682. | ||
Wang H, Wu K, Liu Y, Wu Y, Wang X. Integrative proteomics to understand the transmission mechanism of Barley yellow dwarf virus-GPV by its insect vector Rhopalosiphum padi. Sci Rep. 2015;5:10971. | ||
Zhong XW, Zou Y, Liu SP, et al. Proteomic-based insight into malpighian tubules of silkworm Bombyx mori. PLoS One. 2013;8(9):e75731. | ||
Horwich AL, Fenton WA, Chapman E, Farr GW. Two families of chaperonin: physiology and mechanism. Annu Rev Cell Dev Biol. 2007;23:115–145. | ||
Valpuesta JM, Martin-Benito J, Gomez-Puertas P, Carrascosa JL, Willison KR. Structure and function of a protein folding machine: the eukaryotic cytosolic chaperonin CCT. FEBS Lett. 2002;529(1):11–16. | ||
Koganti S, Clark C, Zhi J, et al. Cellular STAT3 functions via PCBP2 to restrain Epstein-Barr virus lytic activation in B lymphocytes. J Virol. 2015;89(9):5002–5011. | ||
Hu CE, Liu YC, Zhang HD, Huang GJ. The RNA-binding protein PCBP2 facilitates gastric carcinoma growth by targeting miR-34a. Biochem Biophys Res Commun. 2014;448(4):437–442. | ||
Chen X, Hao B, Liu Y, et al. The histone deacetylase SIRT6 suppresses the expression of the RNA-binding protein PCBP2 in glioma. Biochem Biophys Res Commun. 2014;446(1):364–369. | ||
Kereiche S, Kovacik L, Pevala V, et al. Three-dimensional reconstruction of the S885A mutant of human mitochondrial Lon protease. Folia Biol (Praha). 2014;60(suppl 1):62–65. | ||
Cheng CW, Kuo CY, Fan CC, et al. Overexpression of Lon contributes to survival and aggressive phenotype of cancer cells through mitochondrial complex I-mediated generation of reactive oxygen species. Cell Death Dis. 2013;4:e681. | ||
Nie X, Li M, Lu B, et al. Down-regulating overexpressed human Lon in cervical cancer suppresses cell proliferation and bioenergetics. PLoS One. 2013;8(11):e81084. | ||
Ding Z, Roy P. Profilin-1 versus profilin-2: two faces of the same coin? Breast Cancer Res. 2013;15(3):311. | ||
Cheng YJ, Zhu ZX, Zhou JS, et al. Silencing profilin-1 inhibits gastric cancer progression via integrin beta1/focal adhesion kinase pathway modulation. World J Gastroenterol. 2015;21(8):2323–2335. | ||
Wang X, Regufe da Mota S, Liu R, et al. Eukaryotic elongation factor 2 kinase activity is controlled by multiple inputs from oncogenic signaling. Mol Cell Biol. 2014;34(22):4088–4103. | ||
Zhu H, Yang X, Liu J, et al. Eukaryotic elongation factor 2 kinase confers tolerance to stress conditions in cancer cells. Cell Stress Chaperones. 2015;20(2):217–220. | ||
Finka A, Sood V, Quadroni M, Rios Pde L, Goloubinoff P. Quantitative proteomics of heat-treated human cells show an across-the-board mild depletion of housekeeping proteins to massively accumulate few HSPs. Cell Stress Chaperones. 2015;20(4):605–620. | ||
Finka A, Goloubinoff P. Proteomic data from human cell cultures refine mechanisms of chaperone-mediated protein homeostasis. Cell Stress Chaperones. 2013;18(5):591–605. | ||
Murphy ME. The HSP70 family and cancer. Carcinogenesis. 2013;34(6):1181–1188. | ||
Maehara Y, Oki E, Abe T, et al. Overexpression of the heat shock protein HSP70 family and p53 protein and prognosis for patients with gastric cancer. Oncology. 2000;58(2):144–151. | ||
Lanneau D, Brunet M, Frisan E, Solary E, Fontenay M, Garrido C. Heat shock proteins: essential proteins for apoptosis regulation. J Cell Mol Med. 2008;12(3):743–761. | ||
Ciocca DR, Calderwood SK. Heat shock proteins in cancer: diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones. 2005;10(2):86–103. | ||
Deane EE, Woo NY. Impact of heavy metals and organochlorines on hsp70 and hsc70 gene expression in black sea bream fibroblasts. Aquat Toxicol. 2006;79(1):9–15. | ||
Geraci F, Pinsino A, Turturici G, Savona R, Giudice G, Sconzo G. Nickel, lead, and cadmium induce differential cellular responses in sea urchin embryos by activating the synthesis of different HSP70s. Biochem Biophys Res Commun. 2004;322(3):873–877. | ||
Liu T, Daniels CK, Cao S. Comprehensive review on the HSC70 functions, interactions with related molecules and involvement in clinical diseases and therapeutic potential. Pharmacol Ther. 2012;136(3):354–374. | ||
Krebs CJ, Jarvis ED, Pfaff DW. The 70-kDa heat shock cognate protein (Hsc73) gene is enhanced by ovarian hormones in the ventromedial hypothalamus. Proc Natl Acad Sci U S A. 1999;96(4):1686–1691. | ||
Rohde M, Daugaard M, Jensen MH, Helin K, Nylandsted J, Jaattela M. Members of the heat-shock protein 70 family promote cancer cell growth by distinct mechanisms. Genes Dev. 2005;19(5):570–582. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.