Back to Journals » Journal of Inflammation Research » Volume 19
Identification and Analysis of Remimazolam’s Potential Therapeutic Mechanisms in Murine Sepsis-Associated Encephalopathy via Transcriptomic Analysis
Authors Zhou J, Hu W ![]() , Luo Y, Zhang X
, Luo Y, Zhang X ![]() , Hu B
, Hu B ![]() , Zou X
, Zou X ![]()
Received 22 August 2025
Accepted for publication 24 January 2026
Published 3 February 2026 Volume 2026:19 562264
DOI https://doi.org/10.2147/JIR.S562264
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Anh Ngo
Jin Zhou,1,* Wen Hu,1,* Yushan Luo,2 Xujiao Zhang,2 Bailong Hu,1 Xiaohua Zou1
1Department of Anesthesiology, The Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China; 2School of Anesthesiology, Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Bailong Hu, Department of Anesthesiology, The Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China, Tel +86-15185184309, Email [email protected] Xiaohua Zou, Department of Anesthesiology, The Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, People’s Republic of China, Tel +86-13809416036, Fax +86-851-86771013, Email [email protected]
Background: Sepsis-associated encephalopathy (SAE) is a common complication of sepsis, marked by neuroinflammation and cognitive impairment. Remimazolam (Rem), a novel ultra-short-acting benzodiazepine, exhibits potential anti-inflammatory and neuroprotective effects; however, its molecular mechanisms in SAE remain poorly understood.
Methods: Male mice were randomly assigned to three groups: SHAM (Con, n=6), Sepsis (CLP, n=6), and remimazolam-treated Sepsis (Rem, n=6). Hippocampal tissues were collected for RNA sequencing. Differential expression analysis was performed to identify genes modulated by Rem treatment. To further screen candidate genes, weighted gene co-expression network analysis (WGCNA) and protein–protein interaction (PPI) networks were utilized, and functional enrichment was assessed through gene set enrichment analysis (GSEA). Immune microenvironment alterations were compared across groups, and potential therapeutic compounds targeting hub genes were predicted using public drug–gene interaction databases. Hub gene expression was validated at both mRNA (qRT-PCR) and protein levels.
Results: RNA-seq identified 3397 and 914 differentially expressed genes (DEGs) in the Sepsis (CLP vs Con) and remimazolam-treated (Rem vs CLP) groups, respectively. A total of 647 key DEGs with opposing expression trends were screened. Through WGCNA and PPI analysis, five hub genes—ICAM1, Tlr2, Cd274, FOS, and SOCS3—were identified. Pathways related to inflammatory response, IL6/JAK/STAT3 signaling, and TNFα/NF-κB signaling were significantly enriched with these genes, suggesting their potential role in the pathogenesis of SAE. Among the twenty-one candidate drugs predicted to target these hub genes, AVELUMAB-CD274 and LIFITEGRAST-ICAM1 exhibited high binding affinity. The expressions of ICAM1, Tlr2, Cd274, and SOCS3 were further validated at both mRNA (qRT-PCR) and protein levels, corroborating the transcriptomic findings.
Conclusion: This study reveals that ICAM1, Tlr2, Cd274, and SOCS3 are key molecular correlates of remimazolam treatment in a murine SAE model. Our transcriptomic analysis offers insights into the molecular basis of remimazolam’s protective effects, highlighting these genes and pathways for future research.
Keywords: sepsis-associated encephalopathy, remimazolam, immune microenvironmental, RNA sequencing
Introduction
Sepsis-associated encephalopathy (SAE) is a neurological complication often seen in sepsis, characterized by diffuse brain dysfunction caused by systemic infection and inflammation.1,2 Clinically, SAE presents as a spectrum of consciousness disturbances, ranging from mild disorientation to deep coma, and is often accompanied by persistent cognitive deficits. These neurocognitive impairments substantially reduce patients’ quality of life and impose significant psychological and economic burdens on both families and healthcare systems.3,4 Despite its high prevalence and clinical relevance, the underlying pathophysiological mechanisms remain incompletely elucidated. Current evidence suggests that SAE involves a multifaceted interplay of systemic and neuroinflammatory responses. Crucially, the activation of central nervous system (CNS) resident immune cells—primarily microglia and astrocytes—is a key driver of this process. Excessive microglial activation contributes to neuroinflammation, neuronal damage, and cognitive dysfunction,5,6 while astrocytic dysregulation disrupts metabolic and homeostatic support for neurons, exacerbating pathology.7 Other implicated mechanisms include elevated cytokine levels, disruption of the blood–brain barrier, cerebral hypoperfusion, neurotransmitter imbalance, and mitochondrial dysfunction.8–10 Given these complexities, the identification of reliable biomarkers is crucial for early diagnosis, risk stratification, and mechanistic insight into SAE progression.
Remimazolam is a novel ultra-short-acting benzodiazepine with key pharmacological benefits due to its rapid onset and brief duration, making it ideal for anesthesia and sedation with quick recovery.10,11 It acts on GABA_A receptors and is metabolized by tissue esterases, not relying on liver or kidney function, making it safe for patients with hepatic or renal issues.12 Recent research has illuminated its anti-inflammatory and organ-protective properties in sepsis-related models.13 In a lipopolysaccharide (LPS)-induced neuronal injury model, by activating the Nrf2/HO-1 signaling pathway, remimazolam was shown to reduce oxidative stress and neuronal apoptosis, which in turn improved cognitive abilities and reduced anxiety-like behaviors.14 In a separate model of sepsis-associated acute liver injury (SALI), remimazolam was found to suppress the release of pro-inflammatory cytokines by inhibiting p38 phosphorylation via activation of the peripheral benzodiazepine receptor (PBR).15 Collectively, these findings highlight remimazolam’s therapeutic potential in the context of sepsis and its complications; however, its precise molecular mechanisms, particularly within the central nervous system, require further elucidation.
Although the neuroprotective potential of remimazolam has been preliminarily demonstrated, the precise molecular mechanisms underlying its cognitive benefits in SAE remain to be defined. To address this gap, we posited that remimazolam provides neuroprotection in SAE by modulating critical pathways involved in neuroinflammation. The objective of this study was twofold: to rigorously assess this hypothesis by evaluating the cognitive benefits of remimazolam, and to elucidate its molecular mechanisms through transcriptomic analysis in a murine model of SAE. In this study, a CLP-induced SAE mouse model was established, and transcriptome sequencing was conducted to identify gene expression changes following remimazolam treatment. Hub genes were screened using an integrated approach combining WGCNA and five PPI-based algorithms. Functional enrichment, pathway analysis, and drug–gene interaction prediction were subsequently performed to elucidate their biological roles. The expression of selected hub genes was further validated at both transcript and protein levels, providing new insights into the molecular basis of remimazolam’s protective effects in SAE.
Materials and Methods
Grouping and Animal Model
A total of thirty male C57BL/6 mice (8 weeks old, 20–24 g) were obtained from the Laboratory Animal Center of Guizhou Medical University (License No. SYXK [Guizhou] 2025–0001). All experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Guizhou Medical University (Approval No. 2503078) and were conducted in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Animals were kept in a sterile setting with free access to food and water. Mice were divided into three groups: Sham-operated control (Con, n=10), Sepsis (CLP, n=10), and Sepsis with remimazolam treatment (Rem, n=10). Sepsis was induced in the CLP and Rem groups using the cecal ligation and puncture (CLP) method.16 Within 12 hours post-surgery, sepsis mice showed reduced activity and food and water intake, with about 40% mortality by 24 hours, leaving at least six surviving mice per group for analysis. After anesthesia, the mice were positioned on their backs, and their lower abdomen was shaved and cleaned. A 1 cm midline incision was made, and the cecum was exposed with blunt forceps. Approximately 75% of the cecum was tied off with sterile 3–0 silk suture, punctured twice with an 18-gauge needle, and returned to the peritoneal cavity. The abdominal wall was sutured in four layers and disinfected. Before the incision, remimazolam was given intraperitoneally at 16 mg/kg.17 Approximately 40% of mice in the CLP and Rem groups died within 24 hours as expected. Subsequent analyses used the surviving mice, with a final count of n=6 per group (Con:6, CLP:6, Rem:6). These biological replicates were used for hippocampal RNA-seq, HE staining, and Western blot. Hippocampal tissues were collected after 24 hours, either stored at −80°C or fixed in 4% paraformaldehyde. At the experiment’s end, mice were euthanized with an overdose of pentobarbital sodium (150 mg/kg) following ethical guidelines. See the ARRIVE checklist in Supplementary File 1.
Mice were tested for neural reflexes to confirm SAE development. A prior study18 defined scores as “0” for no reflexes, “1” for reduced reflexes, and “2” for normal reflexes. A score under 6 indicates SAE.19 Reflexes tested included pinna, corneal, righting, tail flexion, and escape response.
Histopathological Assessment of the Hippocampus via Hematoxylin and Eosin (H&E) Staining
The hippocampus morphology was histologically analyzed using H&E staining. Brain samples were preserved in 4% paraformaldehyde, dehydrated with graded ethanol, and embedded in paraffin. The tissue blocks were sectioned, deparaffinized, and stained with H&E. After dehydration and mounting, sections were examined and imaged using a Motic BA210 digital trinocular microscope.
Morris Water Maze Test
The Morris water maze (MWM) test assessed spatial learning and memory in three groups.20 Testing began four days before CLP using a 100 cm wide, 40 cm tall circular pool with 28 cm deep opaque water. Over four days, each mouse entered the pool from one of four points every 20 minutes, searching for a platform submerged 0.5 cm below the surface for up to 60 seconds. If unsuccessful, mice were guided to the platform for 20 seconds. Twenty-four hours post-surgery, the platform was removed for a 60-second free-swim probe trial, measuring memory retention by counting crossings over the former platform location and the latency period.
Y-Maze Test
Spontaneous alternation behavior was evaluated in a Y-maze made of grey plastic with three equal arms (50 × 10×40 cm) at 120° angles, one day post-surgery. Mice were placed in the maze center and allowed to explore for 8 minutes, with an entry defined as all four paws entering an arm. A digital video system tracked the entry sequence and number. The maze was cleaned with 75% ethanol between trials to remove scents. The spontaneous alternation percentage was calculated using a specific formula:
Spontaneous alteration percentage = ((number of spontaneous alterations)/(total arm entries − 2)) × 100%
This calculation uses animals’ natural preference for unfamiliar areas, based on short-term spatial memory. The numerator counts valid alternations, indicating memory-driven exploration, while the denominator (total entries − 2) represents the maximum possible alternations, normalizing exploration differences. This standardization converts data to a percentage (random level = 50%), assessing spatial working memory and cognitive flexibility.
Transcriptome Sequencing
Total RNA was isolated from mouse hippocampal tissue using TRIzol® Reagent (Magen, China) according to the manufacturer’s protocol. RNA purity was assessed by measuring the A260/A280 absorbance ratio with a Nanodrop ND-2000 spectrophotometer (Thermo Scientific, USA), and RNA integrity was evaluated using an Agilent 4150 Bioanalyzer system (Agilent Technologies, CA, USA). Only samples with an RNA integrity number (RIN) ≥ 7.0 and an A260/A280 ratio between 1.8 and 2.2 were used for library construction. Paired-end sequencing libraries were generated using the VAHTS Universal V6 RNA-seq Library Prep Kit for Illumina (Vazyme, China) following the manufacturer’s instructions. Briefly, polyadenylated mRNA was enriched from 1 μg of total RNA using oligo(dT)-attached magnetic beads and subsequently fragmented in the presence of divalent cations under elevated temperature in ABclonal First Strand Synthesis Reaction Buffer. First-strand cDNA was synthesized from the fragmented mRNA using random hexamer primers and reverse transcriptase (RNase H), followed by second-strand cDNA synthesis with DNA polymerase I, RNase H, buffer, and dNTPs. The resulting double-stranded cDNA underwent end-repair, A-tailing, and adaptor ligation, after which adaptor-ligated fragments were amplified by PCR. PCR products were purified with the AMPure XP system (Beckman Coulter, USA), and library quality was confirmed using an Agilent 4150 Bioanalyzer. Qualified libraries were sequenced on an Illumina NovaSeq 6000 (or MGISEQ-T7) platform to generate 150-bp paired-end reads.
Data Quality Control and Preprocessing
Raw sequencing data in FASTQ format generated from the Illumina (or BGI) platform were subjected to quality control using in-house Perl scripts provided by Shanghai Applied Protein Technology (Shanghai, China). Adapter sequences were removed and reads containing more than 60% of bases with a Phred quality score ≤ 25, as well as reads with an undetermined base (N) content exceeding 5%, were discarded. The resulting high-quality clean reads were used for downstream analyses. Clean reads were aligned to the reference genome in a strand-specific mode using HISAT2 (http://daehwankimlab.github.io/hisat2/). Read counts for each gene were obtained with FeatureCounts (http://subread.sourceforge.net/), and gene expression levels were calculated as fragments per kilobase of transcript per million mapped reads (FPKM) based on gene length and mapped read counts.
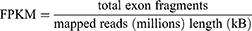
Finally, the principal component analysis (PCA) was used in R (v. 4.3.1) software for the reproducibility of samples.
Differential Expression Analyses
Differential expression was analyzed with the DESeq2 package, version 1.38.0.21 Comparisons were made between the CLP group and the control group (CLP vs Con, designated as DEGs1), and between the remimazolam-treated group and the CLP group (Rem vs CLP, designated as DEGs2). Genes with a |log2 fold change| greater than 0.8 and an adjusted p-value less than 0.05 were identified as differentially expressed in both comparisons. Volcano plots and heatmaps were created using ggplot2 (v.3.4.4)22 and ComplexHeatmap (v.2.16.0),23 packages, respectively, to visualize expression patterns. Genes with inverse expression trends between DEGs1 and DEGs2 were identified as remimazolam-responsive key DEGs, and their intersection was visualized using ggvenn (v.0.1.10, https://github.com/yanlinlin82/ggvenn).
Function Enrichment Analyses
Using the clusterProfiler package (v4.6.0), DEGs1 and DEGs2 underwent Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses.24 A significance level of p < 0.05 was used, and GO annotations were divided into three categories: biological processes (BP), cellular components (CC), and molecular functions (MF). Using the enrichplot package, enriched GO terms and KEGG pathways were visualized.25
Weighted Gene Co-Expression Network Analysis (WGCNA)
The WGCNA package (version 1.71) was utilized to conduct a weighted gene co-expression network analysis,26 to identify gene modules associated with SAE and remimazolam intervention. Initially, hierarchical clustering of all samples was conducted based on Euclidean distances derived from gene expression levels to detect and remove potential outliers. A soft-thresholding power (β) was selected by evaluating the relationship between β and the scale-free topology fit index (R2), aiming for R2 > 0.9 and low mean connectivity. Based on the selected β, a scale-free co-expression network was constructed. Genes were grouped into modules using the following parameters: minimum module size = 200, deepSplit = 2, and mergeCutHeight = 0.25. Modules were assigned different colors for distinction. Pearson correlation analysis was used to assess the associations between each module and experimental traits (sepsis and treatment groups). The module most strongly correlated with both phenotypes (|correlation coefficient| > 0.4, p < 0.05) was defined as the key module. Genes within this module were extracted, and their intersection with SAE treatment–related genes was used to identify candidate genes.
Construction of a Protein–Protein Interaction Network and Detection of Central Genes
To explain the interactions among proteins encoded by the candidate genes, a PPI network was built using the STRING database (https://string-db.org) with a minimum confidence score of 0.4. The interaction landscape was visually represented by the resulting network, and hub genes were identified through the CytoHubba plugin in Cytoscape software (v3.9.1).27 The application of five topological algorithms was carried out, namely maximum neighborhood component (MNC), edge percolated component (EPC), degree, closeness, and betweenness. Each algorithm’s top 10 genes were selected, and those found in more than one algorithm were identified as hub genes.
Hub Genes Analyzed Through Gene Set Enrichment Analysis (GSEA)
In order to examine the potential biological functions of hub genes, the clusterProfiler package (v4.8.3) was used to conduct gene set enrichment analysis (GSEA).28 A total of 186 gene sets were obtained from the C2_CP:KEGG category (species: Mus musculus) via the msigdbr package (v7.5.1).29 Spearman correlation analysis between each hub gene and all other genes was performed using the corrplot package (v0.92, https://github.com/taiyun/corrplot). Genes were ranked based on correlation coefficients, GSEA was performed using these criteria: an absolute normalized enrichment score (NES) greater than 1 and an adjusted p-value less than 0.05.
Immune Cell Infiltration Analysis
To assess whether remimazolam treatment influenced the immune microenvironment in the hippocampus of septic mice, immune cell infiltration was analyzed using the ImmuCellAI package (v0.1.0).30 A total of 22 immune cell types were profiled, and the infiltration landscape was visualized with the ggplot2 package (v3.4.4).22 A comparison of the infiltration levels of 22 immune cell subsets between groups was conducted using the Wilcoxon rank-sum test (p < 0.05). Spearman correlation analysis was also carried out to determine the connections between hub gene expression and immune cell abundance, with significance marked by |correlation coefficient| > 0.3 and p < 0.05.
Identifying miRNAs and Transcription Factors Connected to Core Genes
MicroRNAs predicted to target the hub genes were acquired from the miRDB database (https://mirdb.org/) with a target score of 80 or above. Concurrently, the miRWalk database (http://129.206.7.150/) was used to predict miRNA–mRNA interactions by selecting interactions with a binding site score of 1.
Drug Prediction and Molecular Docking Analysis
Candidate drugs targeting hub genes were retrieved from the Drug–Gene Interaction Database (DGIdb, http://www.dgidb.org/) to identify potential sepsis treatments, focusing on FDA-approved and immunotherapy-related compounds. After constructing a drug-gene interaction network, key drugs were selected based on having an interaction score over 2. The analysis of Spearman correlation was carried out between these drugs and hub genes, with the criteria set at |correlation coefficient| > 0.3 and p < 0.05. Further analysis was done on compounds with interaction scores exceeding 5. 3D chemical structures were collected from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/), and the 3D protein structures of hub genes were retrieved from the RCSB Protein Data Bank (http://www.pdb.org/). AutoDock Vina (v1.5.7) facilitated the molecular docking process.31 PyMOL (v2.5.5, https://pymol.org/) and the Proteins.plus platform (https://proteins.plus/) were used to visualize the docking results.
Confirmation Through Quantitative Real-Time PCR (qRT-PCR)
QRT-PCR assessed hub gene expression in hippocampal tissues from 18 mice divided into SAE, control, and remimazolam-treated groups (n=6 each). Total RNA was extracted using the TriQuick Reagent Kit, and cDNA was synthesized with the PrimeScript™ RT Reagent Kit. qPCR was performed with TB Green Premix Ex Taq II, following a thermal cycle of 95 °C for 30 seconds, then 40 cycles of 95 °C for 5 seconds and 60 °C for 30 seconds. ICAM1, Tlr2, Cd274, and SOCS3 mRNA levels were measured, using GAPDH as the control. GraphPad Prism 5 was used for statistical analysis and plotting, with primer sequences listed in Table 1.
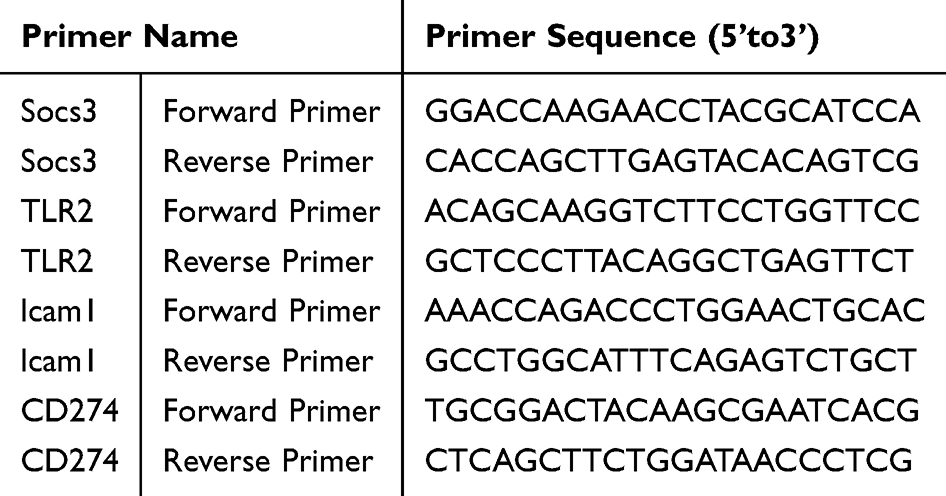 |
Table 1 Sequences of Primers Used for Quantitative Real-Time PCR |
Western Blot Analysis
Protein expression levels of the hub genes ICAM1, Tlr2, Cd274, and SOCS3 in hippocampal tissues were verified via Western blotting. Total protein was extracted from frozen samples using RIPA lysis buffer and protease inhibitors, and concentrations were determined with the BCA Protein Assay Kit. Proteins were separated by SDS-PAGE: 10% gels for ICAM1 and TLR2, and 12% gels for CD274 and SOCS3, then transferred to PVDF membranes. Membranes were blocked with 5% non-fat milk in TBST and incubated overnight at 4°C with primary antibodies for the target proteins and GAPDH as a loading control. After washing, membranes were treated with HRP-conjugated secondary antibodies for one hour at room temperature. Protein bands were visualized using ECL reagents and captured with a gel documentation system. Band intensities were quantified with ImageJ, and protein expression was normalized to GAPDH. Statistical analysis was performed using GraphPad Prism 5.
Enzyme-Linked Immunosorbent Assay (Elisa)
Using commercial ELISA kits and following the manufacturer’s protocols, hippocampal tissue levels of cytokines like IL-6 and TNF-α were quantified. A microplate reader was used to measure absorbance at 450 nm, and cytokine levels were calculated using standard curves for each target analyte.
Statistical Analysis
The R software (v. 4.3.1) was used for all statistical analyses, with a p-value of less than 0.05 indicating statistical significance in every case (two-tailed).
Results
Remimazolam Alleviates Hippocampal Neuronal Injury, Improves Neurobehavioral Outcomes, and Mitigates Central Neuroinflammation in Septic Mice
To assess the neuroprotective effects of remimazolam (Rem) following sepsis, H&E staining of the hippocampal dentate gyrus (DG) was performed 24 hours post-CLP (Figure 1A–C). The control group exhibited well-preserved hippocampal architecture with abundant, orderly arranged neurons and distinct nuclei. In contrast, the CLP group demonstrated notable pathological changes, including neuronal shrinkage, deepened staining, and irregular morphology, particularly in the CA3 region. Remimazolam treatment mitigated these abnormalities, suggesting its protective effect against sepsis-induced neuronal damage. Neurological scoring (Figure 1D) further demonstrated that CLP mice exhibited significant functional impairment, peaking at 24 hours. Rem administration significantly improved neurological performance. According to SAE diagnostic criteria, no SAE cases were observed in the control or Rem groups, while 70% of CLP mice met SAE criteria. Only SAE-positive animals were included in downstream analyses. To evaluate neuroinflammation, hippocampal IL-6 and TNF-α levels were measured (Figure 1E and F). Both cytokines were markedly elevated in the CLP group and significantly reduced by Rem treatment, suggesting that remimazolam mitigates central inflammatory responses in septic encephalopathy.
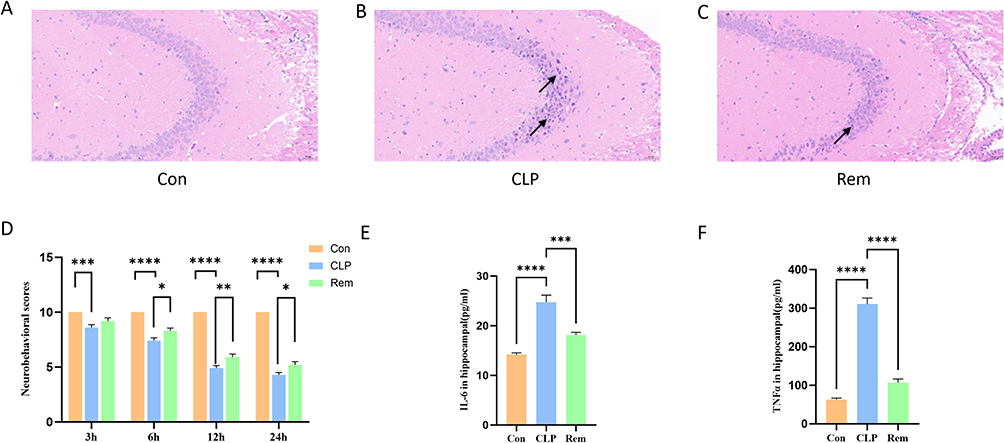 |
Figure 1 Effects of remimazolam on hippocampal neurons, neurobehavioral performance, and inflammatory cytokines in septic mice. (A–C) Representative HE-stained hippocampal sections from each group. Black arrows indicate nuclear disorganization, neuronal shrinkage, and necrosis. (D) Neurobehavioral score. (E and F) Levels of IL-6 and TNF-α in hippocampal tissue across different groups. * p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001, n=6. |
Remimazolam Aids in Enhancing Cognitive Function in Mice Affected by Encephalopathy Due to Sepsis
The experimental timeline is shown in Figure 2A. Spatial learning and memory, both dependent on hippocampal function, were assessed using the Morris Water Maze (MWM). Representative swimming paths during the probe trial are presented in Figure 2B. Over the four-day training period, all groups exhibited progressively reduced latency to locate the hidden platform, with no significant differences in average escape latency among the Con, CLP, and Rem groups (Figure 2C), indicating that basic spatial learning ability was preserved. In contrast, during the probe test, mice in the CLP group spent significantly less time in the target quadrant and had fewer platform crossings compared to controls, reflecting impaired memory retention. These deficits were notably alleviated by remimazolam treatment (Figure 2D and E). Additionally, in the Y-maze test, the CLP group showed a marked reduction in spontaneous alternation percentage, which was significantly improved following remimazolam administration (Figure 2F). Collectively, these findings suggest that while spatial learning acquisition remained intact, remimazolam effectively mitigated memory and working memory impairments in SAE mice.
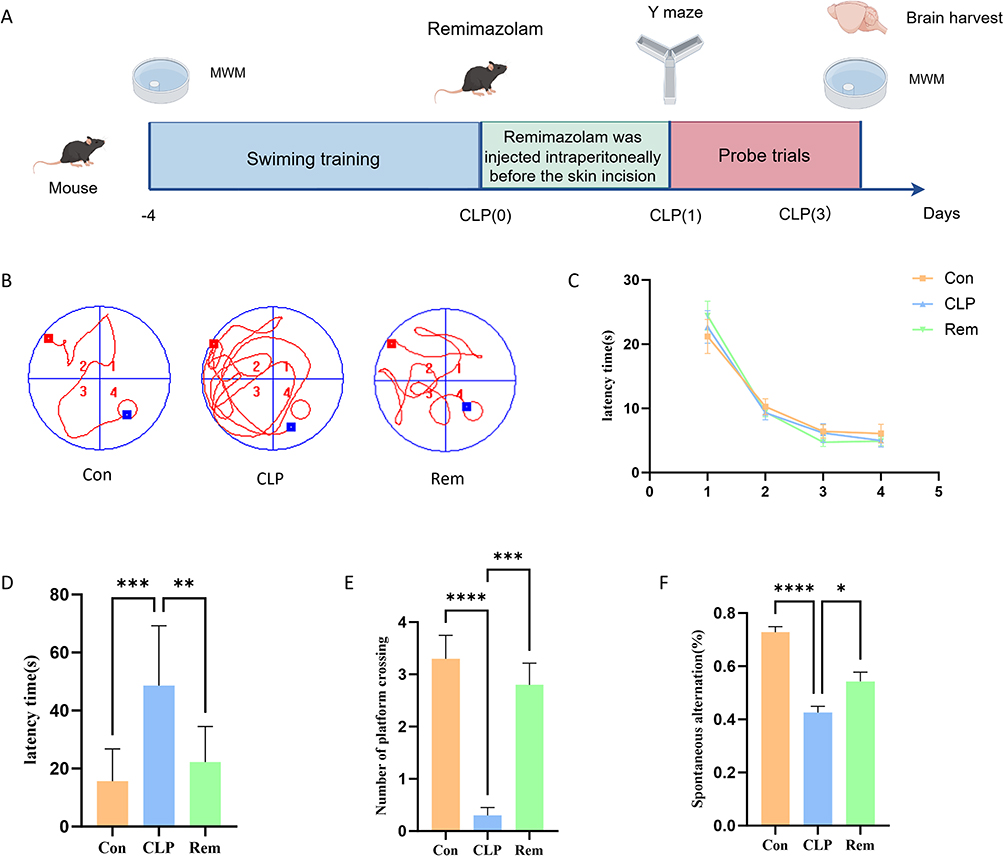 |
Figure 2 Remimazolam attenuates cognitive deficits in SAE mice. (A) Schematic illustration of the experimental design for establishing the SAE mouse model. (B) Representative swimming paths of each group in the MWM test. (C) Escape latency times in the MWM test across all groups. (D) Latency times in the target quadrant during the testing phase for each group. (E) Number of platform crossings in the testing phase for each group. (F) Spontaneous alternation in the testing phase for each group. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, n = 6. |
Data Evaluation
All samples showed high-quality alignment, with over 90% of reads uniquely mapped, indicating reliable sequencing performance (Supplementary Table 1). Principal component analysis (PCA) confirmed strong intra-group consistency, with no discernible outliers among the samples (Supplementary Figure 1).
Identification and Functional Profiling of Remimazolam-Responsive Genes in SAE
Between the CLP and control groups, transcriptomic analysis identified 3397 genes with differential expression, including 1753 that were upregulated and 1644 that were downregulated (Figure 3A and B). Comparison between the remimazolam and CLP groups revealed 914 DEGs, with 463 upregulated and 451 downregulated transcripts (Figure 3C and D). Through the intersection of DEGs with differing trends in the two comparisons, 647 genes potentially regulated by remimazolam were identified (Figure 3E). GO enrichment analysis showed that these genes were mainly involved in inflammatory responses, particularly to bacterial molecules and lipopolysaccharide. Molecular function terms included CARD domain binding, GTP binding, and GTPase activity, suggesting a role in innate immune signaling (Figure 3F). Further analysis of KEGG pathways revealed a notable enrichment in the TNF signaling and NOD-like receptor pathways (Figure 3G), indicating that remimazolam may exert neuroprotective effects in SAE by modulating central inflammation and key immune signaling cascades.
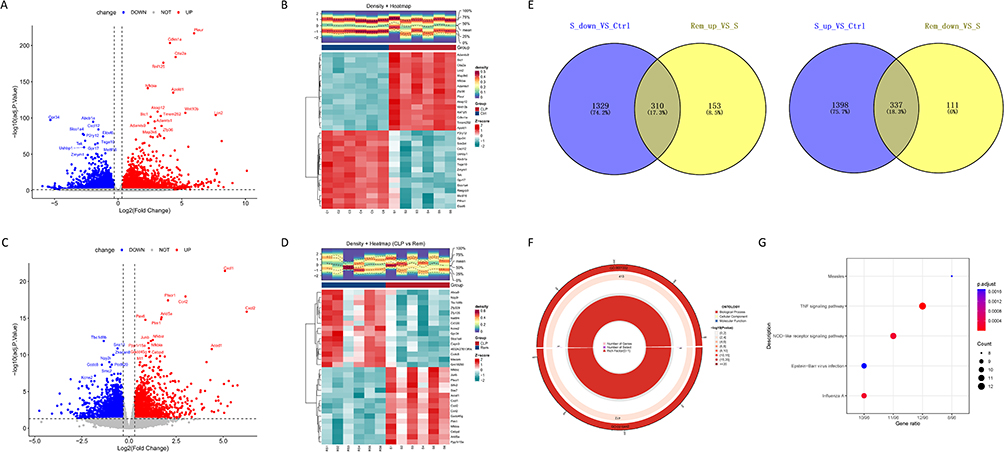 |
Figure 3 Identification of differentially expressed genes (DEGs). (A) Volcano plot depicting DEGs between CLP and control groups. (B) Heatmap illustrating DEGs between CLP and control groups. (C) Volcano plot of DEGs between CLP and Rem groups. (D) Heatmap of DEGs between CLP and Rem groups. (E) Venn diagram of DEGs among control, CLP, and Rem groups. (F) GO term enrichment of DEGs. (G) KEGG pathway enrichment of DEGs. |
Identification of Candidate Genes Through WGCNA
To uncover gene modules associated with sepsis progression and remimazolam response, WGCNA was conducted. Sample clustering confirmed the absence of outliers, ensuring a robust foundation for network construction (Figure 4A). With a soft-threshold power of β = 8, the network exhibited a scale-free topology (R2 = 0.9), meeting standard WGCNA criteria (Figure 4B). Three co-expression modules were identified, among which the blue module showed the strongest correlation with both sepsis (cor = 0.94, p = 1e−08) and remimazolam treatment (cor = −0.61, p = 0.008) (Figure 4C and D). Given its relevance to both disease and therapeutic response, this module (containing 54. genes) was selected for further analysis. By intersecting this module’s genes with the 647 DEGs exhibiting reversed expression trends between CLP and Rem groups, 300 overlapping genes were identified as candidate targets potentially involved in both SAE pathology and remimazolam’s neuroprotective effects (Figure 4E).
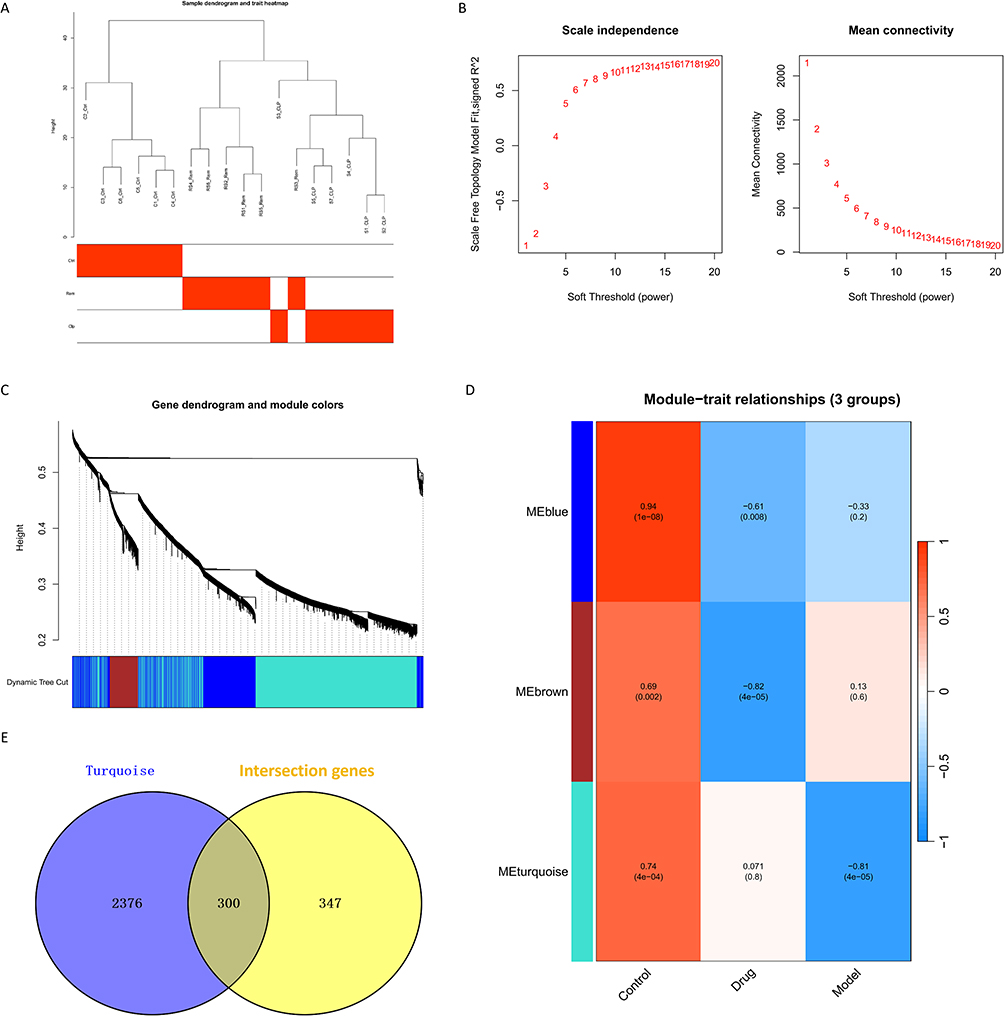 |
Figure 4 Weighted co-expression network analysis (WGCNA) analysis of identifying candidate genes. (A) Determination of sample hierarchical clustering in WGCNA analysis. (B) The soft threshold power and the mean connectivity of WGCNA. (C) The cluster dendrogram of WGCNA. (D) The clustered modules of WGCNA. (E) Venn diagram showing the overlap between Turguosie module genes and DEGs. |
ICAM1, Tlr2, Cd274, FOS, and SOCS3 Were Hub Genes
To clarify key regulatory genes involved in remimazolam’s therapeutic mechanism, a PPI network was constructed based on the 300 candidate genes, generating a network with 96 nodes and 474 edges (Figure 5A). Five topological algorithms—MNC, EPC, Degree, Closeness, and Betweenness—were applied via CytoHubba to rank core genes. After visualizing the top 10 genes from each algorithm (Figure 5B–F), five genes—ICAM1, Tlr2, Cd274, FOS, and SOCS3—were consistently overlapping and identified as hub genes (Figure 5G). Expression analysis showed all five hub genes were significantly upregulated in the CLP group, while remimazolam treatment effectively reversed their expression toward control levels (Figure 5H), suggesting their involvement in SAE pathogenesis and sensitivity to intervention. GSEA revealed these hub genes were commonly enriched in inflammatory and apoptotic pathways, including IL6/JAK/STAT3, TNF-α/NF-κB, and apoptosis (Figure 5I), indicating their central roles in neuroinflammation and immune dysregulation during SAE.
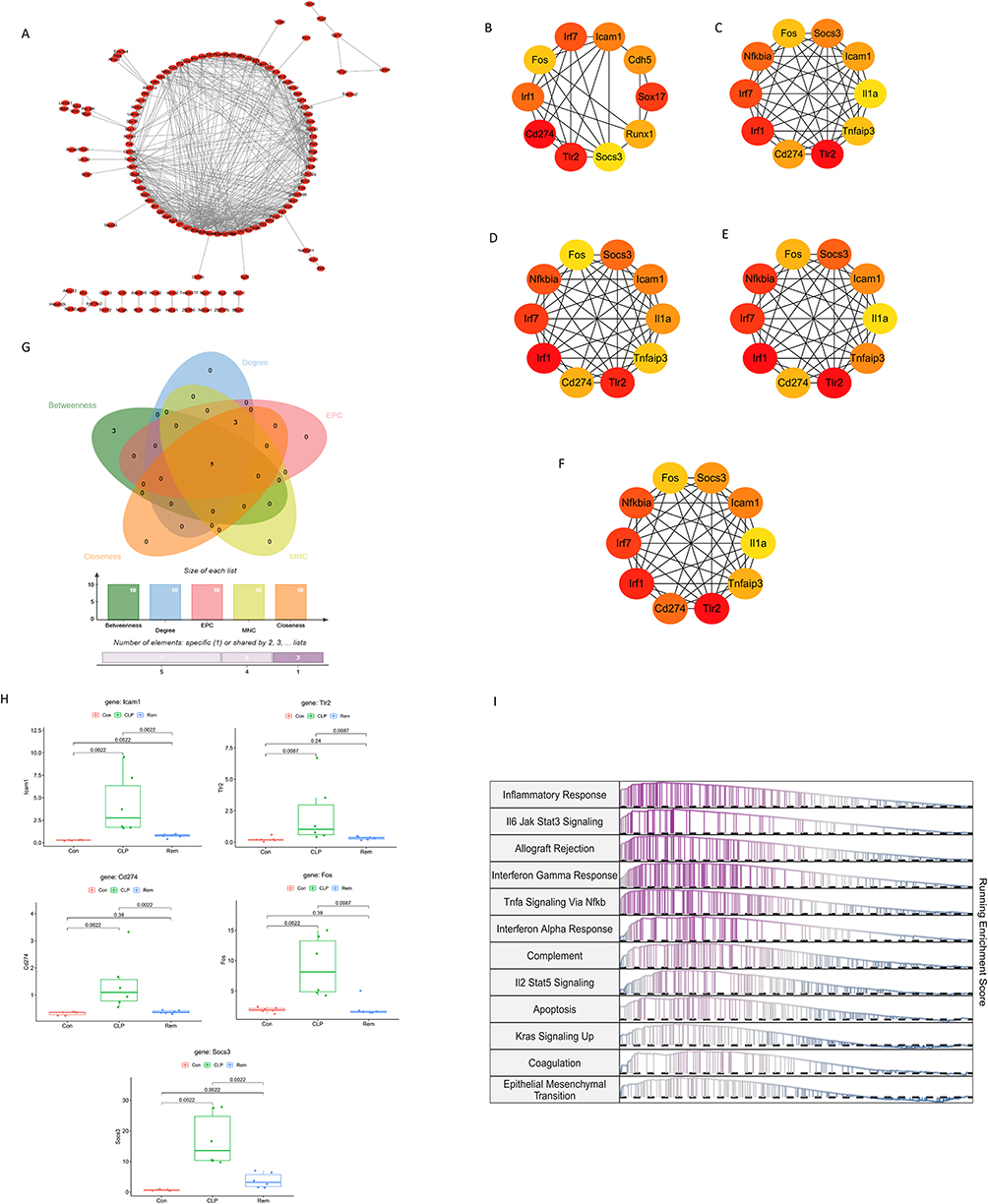 |
Figure 5 Five Hub gene screening using Cytohubba. (A) Protein-protein interaction network of DEGs. (B) Top 10 DEGs ranked by Betweenness. (C) Top 10 DEGs ranked by Degree. (D) Top 10 DEGs ranked by EPC. (E) Top 10 DEGs ranked by MNC. (F) Top 10 DEGs ranked by Closeness. (G) Intersection of top 10 hub genes across five algorithms. (H) Box plots show the relative expression levels of ICAM1, Tlr2, Cd274, FOS, and SOCS3 across experimental groups. (I) Gene set enrichment analysis (GSEA) of the five hub genes. |
Correlations Between Differential Immune Cells and Hub Genes
CIBERSORT-based analysis revealed significant immune landscape alterations following CLP-induced sepsis. In comparison with the control group, the CLP group displayed marked differences in the infiltration proportions of NK cells, T cells, and monocytes (Figure 6A and B). There was a significant rise in monocytes, activated myeloid dendritic cells, and eosinophils within the CLP group (Figure 6C), indicating immune dysregulation during disease progression. Despite remimazolam treatment partially normalizing these deviations, there were no significant statistical changes between the CLP and Rem groups (Figure 6D and E), suggesting that remimazolam modulates immune cell composition without causing major changes. Correlation analysis between hub genes and immune cell subsets demonstrated that T follicular helper cells were negatively correlated with Fos and Socs3, while activated myeloid dendritic cells showed negative associations with ICAM1, SOCS3, and Tlr2 (Figure 6F). These findings imply that dysregulated expression of these hub genes may contribute to the disruption of both innate and adaptive immune responses in SAE, and that remimazolam’s neuroprotective effect may partially involve the indirect modulation of immune activity.
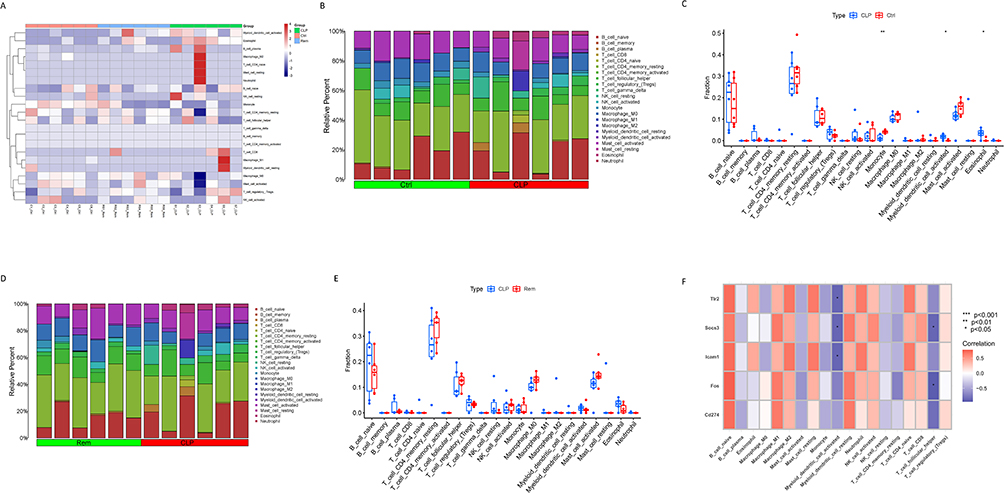 |
Figure 6 Immune cell infiltration analysis across the three groups. (A) Heatmap showing the distribution of 22 immune cell types in each sample by CIBERSORT. (B) Estimated proportion of 22 types of immune cells in the control and CLP groups. (C) Differential analysis of 22 immune cell types between the control and CLP groups. (D) Estimated proportion of 22 types of immune cells in the CLP and Rem groups. (E) Differential analysis of 22 immune cell types between the CLP and Rem groups. (F) Correlation analysis between the seven hub genes and 22 immune cells. * p<0.05, ** p<0.01. |
AVELUMAB with CD274 and LIFITEGRAST-Icam1 Had High Affinity
In order to explain post-transcriptional regulatory mechanisms, miRNA–mRNA interaction data were sourced from the miRDB and miRWalk databases. A total of 68 and 786 potential pairs were initially identified, respectively. Upon intersection, 20 robust miRNA–mRNA pairs targeting the hub genes were retained (Figure 7A and B). Subsequently, drug–gene interaction analysis focusing on four of the five identified hub genes (excluding SOCS3, due to lack of direct drug associations) was performed using relevant pharmacogenomic databases. This yielded a set of candidate compounds, from which drugs with interaction scores above the threshold (>5) were shortlisted for docking evaluation. Among these, AVELUMAB, targeting CD274, and LIFITEGRAST, targeting ICAM1, were selected for molecular docking analysis. Three-dimensional structural modeling and docking simulations demonstrated strong binding affinities for both drug–target pairs: AVELUMAB–CD274 and LIFITEGRAST–ICAM1, as evidenced by favorable binding energies (Figure 7C–E). These results indicate that these compounds could be potential therapeutic candidates for influencing key immune checkpoints and adhesion molecules involved in SAE pathophysiology.
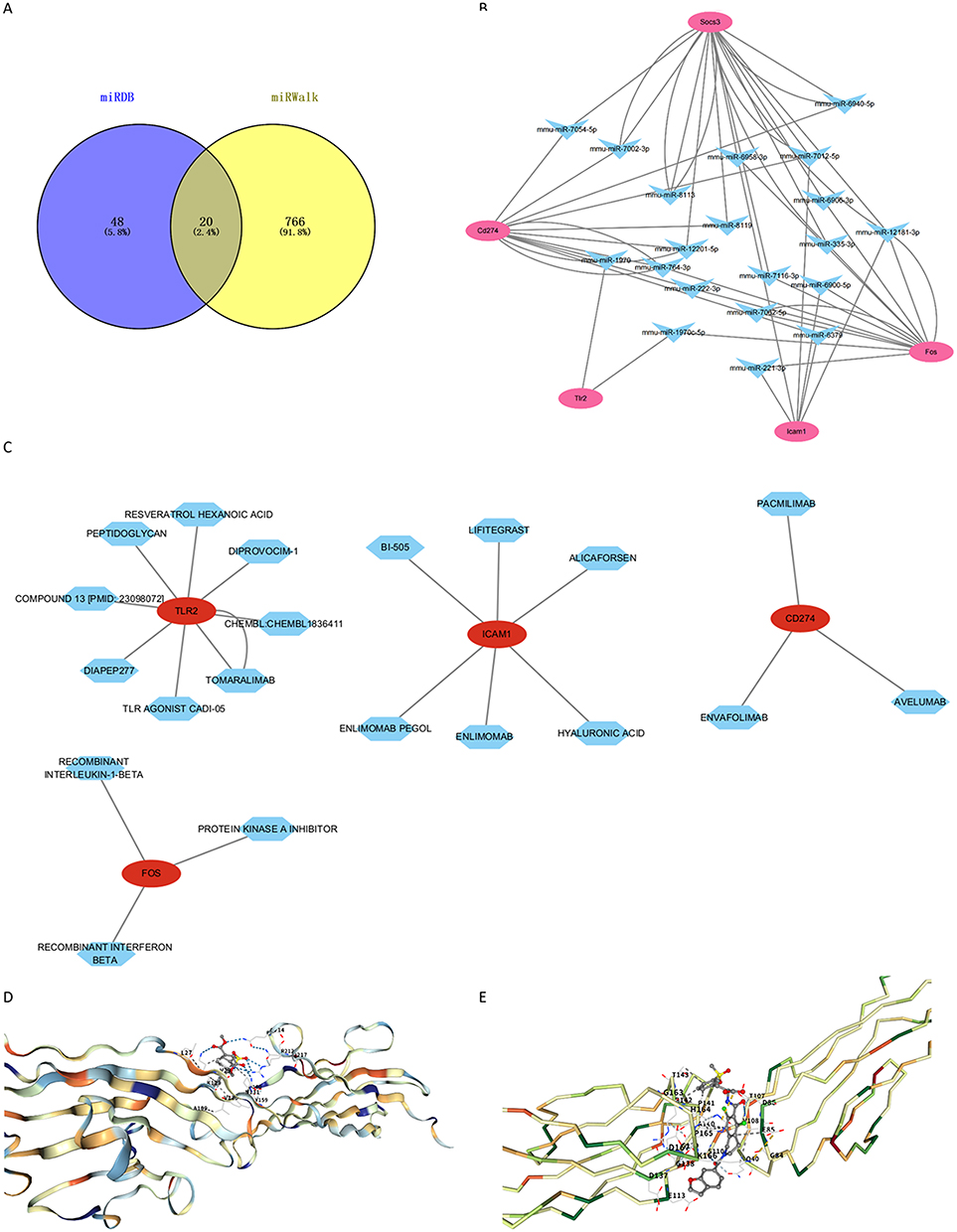 |
Figure 7 Prediction of potential therapeutic drugs targeting the five hub genes. (A) Venn diagram of common mRNA-miRNA interaction pairs predicted for the five hub genes. (B) mRNA-miRNA regulatory network of the five hub genes. (C) Drug-target network of the four hub genes. (D) Molecular docking results of AVELUMAB with CD274. (E) Molecular docking results of LIFITEGRAST with ICAM1. |
Expression Validation of the Hub Genes
To validate transcriptomic findings, qRT-PCR and Western blot assays were conducted to assess the expression of four hub genes—ICAM1, Tlr2, Cd274, and SOCS3—in hippocampal tissue. The mRNA levels of all four genes were significantly increased in the CLP group compared to the control but were notably decreased after remimazolam treatment (Figure 8A–D). These trends were mostly in line with the RNA-seq findings, supporting the trustworthiness of the sequencing data.
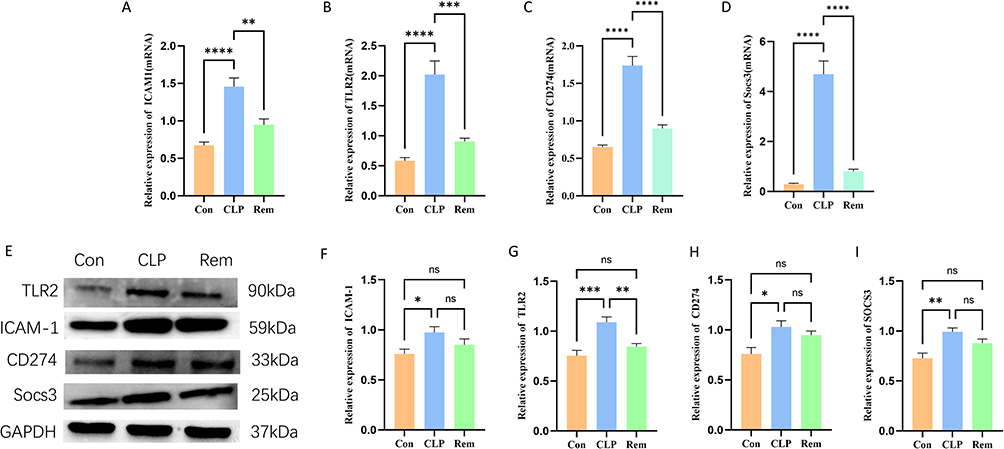 |
Figure 8 Validation of mRNA expression levels for the four hub genes. (A–D) qRT-PCR validation of Icam1, Tlr2, Cd274 and Socs3 expression. (E–I) Western blot analysis and quantification of the corresponding protein expression levels of Icam1, Tlr2, Cd274 and Socs3. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001, ns indicates not significant, n =6. |
At the protein level, Tlr2 and SOCS3 exhibited significant elevation in the CLP group and a marked reduction after remimazolam administration, mirroring mRNA trends. However, for ICAM1 and Cd274, although a downward trend was observed post-remimazolam, the decrease did not reach statistical significance relative to the CLP group (Figure 8E–I). These findings suggest that while remimazolam effectively suppresses the transcriptional upregulation of key inflammatory mediators in SAE, post-transcriptional regulation or protein stability may modulate the final protein expression outcomes.
Discussion
A prevalent and serious complication of sepsis, sepsis-associated encephalopathy (SAE) is characterized by widespread brain dysfunction, including changes in consciousness, neuroinflammation, and cognitive deficits.32 Affecting up to 70% of individuals with sepsis, SAE is significantly correlated with increased mortality rates.33 Current therapeutic approaches for SAE predominantly focus on supportive care for systemic sepsis, with limited pharmacological interventions specifically addressing the neurological manifestations. This underscores the critical necessity for the development of novel therapeutic strategies for SAE. In this regard, remimazolam, an innovative ultra-short-acting benzodiazepine, has garnered significant interest due to its anti-inflammatory and neuroprotective properties.34,35 Nevertheless, its precise mechanisms of action in the context of SAE remain largely undefined. In this study, we conducted a detailed transcriptomic analysis of hippocampal tissues from a sepsis-induced mouse model treated with remimazolam to explore its therapeutic mechanisms in SAE. Using RNA sequencing, WGCNA, and PPI network construction, we identified five inflammation-related hub genes—ICAM1, Tlr2, Cd274, FOS, and SOCS3—linked to remimazolam treatment. These genes’ expression levels were confirmed by qRT-PCR and Western blot analysis, supporting their role in remimazolam’s neuroprotective effects. These findings provide insights and highlight potential therapeutic targets for future SAE clinical interventions.
Four inflammation-related hub genes involved in remimazolam treatment of SAE were identified. ICAM1, a transmembrane glycoprotein, is mainly found on endothelial and immune cells but increases significantly during neuroinflammation.36–38 It interacts with integrins to facilitate leukocyte adhesion and migration, essential for immune response and inflammation.39 Upregulated ICAM1 is associated with proinflammatory cytokine release, neuronal damage, and blood-brain barrier disruption.40,41 Studies show increased ICAM1 expression in SAE patients’ brain tissue, suggesting its role in disease progression.42,43 In murine sepsis models, elevated ICAM1 leads to more leukocyte infiltration, neuronal injury, and blood-brain barrier disruption.44,45 These findings are consistent with our observations of elevated ICAM1 levels in the hippocampal tissue of septic mice, suggesting a possible association between hippocampal ICAM1 upregulation and the pathogenesis of SAE. Toll-like receptor 2 (TLR2) serves as a crucial pattern recognition receptor within the innate immune system, adept at identifying a wide array of pathogen-associated and damage-associated molecular patterns (PAMPs and DAMPs) via its leucine-rich repeat (LRR) domains and through heterodimerization with TLR1 or TLR6.46,47 Prior research has indicated that activation of TLR2 exacerbates neuroinflammation compromises the integrity of the blood-brain barrier and facilitates neuronal damage in the context of sepsis.47–49 Moreover, the expression levels of TLR2 have been shown to correlate with the severity and prognosis of the disease in septic patients, underscoring its dual role as a pathogenic mediator and a prognostic biomarker in SAE.50 In alignment with these findings, our study identified a marked upregulation of TLR2 in the hippocampal tissue of mice with SAE, implying that TLR2 activation may play a contributory role in the pathogenesis of SAE associated encephalopathy. CD274 (PD-L1) is a transmembrane protein characterized by an IgV-like domain that interacts with PD-1 to initiate immunosuppressive signaling pathways.51 In the context of sepsis, there is a significant upregulation of CD274 expression on monocytes and neutrophils, which contributes to immune cell dysfunction and immunoparalysis.52,53 Previous research has demonstrated a positive correlation between elevated CD274 levels and both disease severity and Sequential Organ Failure Assessment (SOFA) scores in septic patients, indicating its potential utility as a prognostic biomarker.54 Furthermore, CD274-mediated T cell exhaustion may exacerbate immunosuppression, underscoring its importance as a marker of immune dysregulation and as a potential therapeutic target in sepsis.52,55 In alignment with these findings, our study observed an upregulation of CD274 in the hippocampal tissue of mice with SAE, suggesting that CD274-driven immunosuppressive mechanisms may play a pivotal role in the pathogenesis of SAE. SOCS3 is a crucial negative regulator of the JAK/STAT pathway, featuring an SH2 domain, SOCS box, and kinase inhibitory region.52 It mitigates inflammation by inhibiting cytokine signaling, particularly in microglia, astrocytes, and neurons.56 During early sepsis, SOCS3 is quickly upregulated to reduce neuroinflammation in the hippocampus and cortex, but its later downregulation may lead to neuronal damage and cognitive issues.57,58 Lower SOCS3 levels are linked to more severe disease and cognitive decline in septic patients, highlighting its role as a neuroprotective agent and prognostic marker in SAE.20,59 These findings align with our current results, supporting our transcriptomic predictions. It is noteworthy that for ICAM1 and CD274, the clear mRNA expression trends did not translate into statistically significant changes at the protein level in our assay. This may reflect biologically relevant post-transcriptional regulation or differences in protein turnover, rather than undermining the validity of the transcriptional findings. Such a discrepancy highlights the complexity of gene expression regulation and warrants further investigation.
Our study identified a significant association between the upregulation of four hub genes and the manifestation of behavioral changes and cognitive impairments in septic mice. Notably, increased expression of ICAM1 has been implicated in the pathogenesis of neurodegenerative diseases, including Alzheimer’s disease (AD),60 Parkinson’s disease (PD),61 and multiple sclerosis (MS).62 In the context of AD, ICAM1 overexpression has been linked to β-amyloid (Aβ) deposition and abnormal tau phosphorylation, indicating its role in amyloidogenic and tau-related neurotoxicity.42 Mechanistically, ICAM1 may worsen neurodegeneration by promoting inflammation, neuronal apoptosis, and disrupting cell communication, contributing to cognitive decline.63–65 TLR2 activation is increasingly seen as harmful to cognitive function.66,67 This immune receptor not only detects pathogens but also triggers neuroinflammation linked to cognitive decline.68,69 In diseases like AD, TLR2 interacts with amyloid-β, activating microglia and astrocytes, which release pro-inflammatory cytokines that damage synapses and neurons, impairing memory and cognition.68,70 In PD, TLR2 activation worsens cognitive deficits, with its expression negatively affecting cognitive performance.71,72 The upregulation of CD274 (PD-L1) is significantly correlated with the onset and advancement of numerous neurodegenerative disorders.73 Research indicates that CD274 may impact cognitive function in diseases such as Alzheimer’s and Parkinson’s by affecting neuronal survival, maintaining synaptic integrity, and modulating neuroinflammatory responses.74–76 SOCS3 might provide neuroprotection in the central nervous system by controlling neuroinflammation, the activation of microglia and astrocytes, and associated processes, thereby influencing cognitive abilities.77–79 Collectively, these four hub genes may be important in boosting cognitive function and behavior associated with SAE.
GSEA revealed that the four hub genes were significantly enriched in the inflammatory Response, IL6/JAK/STAT3 and TNF-α/NF-κB signaling pathway. The IL6/JAK/STAT3 signaling pathway is critically involved in the pathogenesis of SAE by facilitating neuroinflammation and compromising the integrity of the BBB.80,81 Elevated levels of IL-6 activate the JAK/STAT3 pathway, leading to microglial activation and the subsequent release of pro-inflammatory cytokines such as TNF-α and IL-1β, which exacerbate neuronal damage and contribute to cognitive decline.82,83 Additionally, the expression of ICAM1, modulated by IL-6 through the JAK/STAT3 pathway, influences BBB permeability. During sepsis, increased ICAM1 expression further disrupts the BBB, underscoring the role of IL-6 in SAE pathogenesis by altering BBB permeability via ICAM1.84–87 The TNF-α/NF-κB pathway is crucial in SAE. Increased TNF-α activates NF-κB in microglia and astrocytes, releasing proinflammatory cytokines, increasing matrix metalloproteinases, and reducing tight junction proteins like Claudin-5, which weakens BBB permeability.88–90 This pathway’s persistent activation also leads to neuronal damage and cognitive decline by initiating apoptosis and affecting synaptic plasticity.43,91 Recent findings indicate that the TNF-α/NF-κB signaling pathway is crucial for increasing CD274 expression.92 TNF-α, a proinflammatory cytokine, boosts CD274 expression during sepsis and promotes its transcription in microglia and neurons through NF-κB activation.92 This upregulation likely helps moderate immune responses in the central nervous system, reducing neuroinflammation and sepsis-related neurological harm.93 In summary, these hub genes are essential in pathways linked to inflammation, having a significant impact on the progression of SAE.
Prior research has demonstrated that remimazolam can mitigate inflammation, safeguard the BBB, and decrease neuronal apoptosis and tissue damage by interacting with peripheral benzodiazepine receptors, the HMGB1-TLR4-NF-κB signaling pathway, α7 nicotinic acetylcholine receptors, and other mechanisms and these effects contribute to the amelioration of sepsis and SAE.15,34,94 This study is the first to demonstrate that remimazolam attenuates ICAM-1 expression in the hippocampus of septic mice, with ICAM-1 being enriched in the IL-6/JAK/STAT3 and TNF-α/NF-κB signaling pathways. These findings support the hypothesis that remimazolam may mitigate the severity of SAE by inhibiting IL-6/JAK/STAT3 and TNF-α/NF-κB pathways, thereby downregulating ICAM-1 expression in the hippocampus and reducing the synthesis and release of inflammatory mediators. Numerous studies have demonstrated that the activation of TLR2 can induce neuroinflammation through the NF-κB signaling pathway and can synergize with other receptors to amplify inflammatory responses and neuronal injury, thereby contributing to the pathogenesis of SAE.95–97 Our research is the first to reveal that remimazolam reduces both mRNA and protein expression of TLR2 in the hippocampus of septic mice, with TLR2 also being enriched in theTNF-α/NF-κB pathway. These findings suggest that remimazolam may mitigate neuroinflammation and neuronal damage in SAE by inhibiting TLR2 and its downstream inflammatory signaling pathways. While SOCS3 is recognized for inhibiting the JAK-STAT pathway and benefiting sepsis outcomes, no studies have shown remimazolam’s effect on SOCS3 expression until now. Our research is the first to reveal that remimazolam changes SOCS3 mRNA and protein levels in the hippocampus of septic mice. Further investigation is required to confirm if remimazolam improve SAE by influencing SOCS3 and inhibiting the JAK-STAT pathway. While CD274 is a known biomarker for SAE, its interaction with remimazolam has not been studied until now. Our research reveals that remimazolam significantly reduces CD274 expression in the hippocampus of septic mice, indicating a possible neuroprotective effect. Further investigation is required to understand the exact regulatory mechanisms. Overall, our findings indicate that remimazolam improves cognitive function and attenuates hippocampal damage in septic mice, highlighting its potential as a therapeutic candidate for SAE.
The translational significance of our principal findings is further underscored by the analysis of drug-gene interactions. An examination utilizing the DGIdb database identified AVELUMAB and LIFITEGRAST as potential therapeutic agents for the treatment of SAE. AVELUMAB, a monoclonal antibody that targets PD-L1 (encoded by the CD274 gene), is anticipated to inhibit this pathway.98 Preclinical studies have demonstrated that inhibition of PD-L1 can enhance cognitive outcomes in SAE models,99 thereby supporting the potential importance of CD274 dysregulation. Similarly, LIFITEGRAST functions as an inhibitor of intercellular adhesion molecules, with predicted efficacy against ICAM1.100,101 Our molecular docking analysis corroborates this interaction. These predictions clearly illustrate how the hub genes identified in our study, CD274 and ICAM1, can serve as actionable targets for the development of novel therapies for SAE.
In this study, we conducted a comprehensive investigation into the neuroprotective mechanisms of remimazolam in SAE. Our findings indicate that remimazolam may mitigate SAE pathology by modulating several inflammatory signaling pathways, including IL6/JAK/STAT3, TNF-α/NF-κB. This modulation results in the attenuation of neuroinflammation, preservation of blood-brain barrier integrity, and reduction of neuronal apoptosis. Concurrently, we identified a few immune-related genes, such as ICAM1, CD274, TLR2, and SOCS3, whose expression levels were significantly altered by remimazolam, suggesting their potential roles as mediators of its therapeutic effects. Moreover, drug-gene interaction analyses identified AVELUMAB and LIFITEGRAST as potential agents targeting CD274 and ICAM1, respectively, thereby providing new avenues for SAE intervention. However, this study has several limitations. First, while key molecular targets were identified, their precise mechanistic roles require further validation through functional experiments. Specifically, the hub gene FOS was excluded from the final validation panel as it is primarily associated with broad cellular activation and acute stress responses, whereas our study focused on dissecting specific, sustained neuroinflammatory pathways in SAE. Consequently, we prioritized the validation of genes (ICAM1, TLR2, CD274, and SOCS3) with more direct and established links to the immune and inflammatory pathways central to our transcriptomic findings. Second, our current validation at the transcriptional and protein levels could be complemented in the future by spatial techniques, such as immunohistochemistry, to resolve the cellular localization of these targets within the hippocampus. Third, the study’s statistical power is constrained by the sample size (n=6 per group), a common limitation in intensive rodent models of sepsis such as CLP, which may account for the lack of statistical significance in some protein-level validation trends. Finally, due to potential interspecies differences, the translational relevance of these findings awaits confirmation in clinical SAE contexts.
Conclusion
In conclusion, this study utilized transcriptome sequencing and bioinformatics approaches to identify four key hub genes—ICAM1, TLR2, CD274, and SOCS3—whose expression in the hippocampus was associated with remimazolam treatment in a murine SAE model. These genes are enriched in pathways related to IL-6/JAK/STAT3 and TNF-α/NF-κB signaling. Our findings suggest an association between remimazolam’s neuroprotective effects and the regulation of these specific genes and pathways, highlighting them as potential targets for future SAE research.
Data Sharing Statement
To ensure data transparency and in full compliance with the journal’s “Share Upon Reasonable Request” policy, the raw sequencing datasets supporting this study have been publicly deposited in the NCBI Sequence Read Archive (SRA). The data can be accessed via the following persistent link: https://dataview.ncbi.nlm.nih.gov/object/PRJNA1307203?reviewer=psh9mgnmp4ukqojn0scii4af6i.
Ethics Approval and Consent to Participate
The animal study protocol was approved by the Animal Ethics Committee of Guizhou Medical University (Approval No. 2503078; License No. SYXK [Guizhou] 2025-0001). All procedures were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. This research was designed, performed, and reported in compliance with the ARRIVE guidelines (https://arriveguidelines.org), and a completed ARRIVE checklist is provided as Supplementary File 1.
Author Contributions
Jin Zhou contributed to conceptualization, methodology, investigation, and writing – original draft. Wen Hu contributed to data curation and validation. Yushan Luo was responsible for software and formal analysis. Xujiao Zhang contributed to visualization. Bailong Hu contributed to methodology, supervision, resources, and writing – review & editing. Xiaohua Zou contributed to conceptualization, funding acquisition, project administration, writing – review & editing, and provided final approval for publication. All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Jin Zhou and Wen Hu contributed equally to this work and share first authorship.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82160224), the Key Laboratory of Anesthesia and Pain Research at Guizhou Medical University ([2024] fy003), and the High-level Innovative Talent Project of Guizhou Province (gzwjrs −2024-009).
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Tauber SC, Djukic M, Gossner J, et al. Sepsis-associated encephalopathy and septic encephalitis: an update. Expert Rev Anti Infect Ther. 2021;19(2):215–20. doi:10.1080/14787210.2020.1812384
2. Helbing DL, Böhm L, Witte OW. Sepsis-associated encephalopathy. Cmaj. 2018;190(36):E1083. doi:10.1503/cmaj.180454
3. Cotena S, Piazza O. Sepsis-associated encephalopathy. Transl Med UniSa. 2012;2:20–27.
4. Chaudhry N, Duggal AK. Sepsis associated encephalopathy. Adv Med. 2014;2014:762320. doi:10.1155/2014/762320
5. Yan X, Yang K, Xiao Q, et al. Central role of microglia in sepsis-associated encephalopathy: from mechanism to therapy. Front Immunol. 2022;13:929316. doi:10.3389/fimmu.2022.929316
6. Yu Z, Shi H, Zhang J, et al. Role of microglia in sepsis-associated encephalopathy pathogenesis: an update. Shock. 2024;61(4):498–508. doi:10.1097/SHK.0000000000002296
7. Li L, Lixia D, Gan G, et al. Astrocytic HILPDA promotes lipid droplets generation to drive cognitive dysfunction in mice with sepsis-associated encephalopathy. CNS Neurosci Ther. 2024;30(5):e14758. doi:10.1111/cns.14758
8. Brummel NE, Jackson JC, Pandharipande PP, et al. Delirium in the ICU and subsequent long-term disability among survivors of mechanical ventilation. Crit Care Med. 2014;42(2):369–377. doi:10.1097/CCM.0b013e3182a645bd
9. Krzyzaniak K, Krion R, Szymczyk A, et al. Exploring neuroprotective agents for sepsis-associated encephalopathy: a comprehensive review. Int J Mol Sci. 2023;24(13):10780. doi:10.3390/ijms241310780
10. Hong Y, Chen P, Gao J, et al. Sepsis-associated encephalopathy: from pathophysiology to clinical management. Int Immunopharmacol. 2023;124(Pt A):110800. doi:10.1016/j.intimp.2023.110800
11. Lee A, Shirley M. Remimazolam: a review in procedural sedation. Drugs. 2021;81(10):1193–1201. doi:10.1007/s40265-021-01544-8
12. de Jong BT, Eleveld DJ, Mason KP, et al. Clinical pharmacokinetics and pharmacodynamics of remimazolam. Clin Pharmacokinet. 2025;64(9):1263–1282. doi:10.1007/s40262-025-01548-7
13. Dai QC, Zhao J-L, Miao X-Y, et al. Effects of different doses of remimazolam on hemodynamics during general anesthesia in patients with septic shock. Eur Rev Med Pharmacol Sci. 2024;28(6):2483–2492. doi:10.26355/eurrev_202403_35755
14. Wei Y, Pan S, Zhou Z, et al. Remimazolam attenuated lipopolysaccharide-induced behavioral deficits and neuronal injury via activation of the Nrf2 pathway. Sci Rep. 2025;15(1):13784. doi:10.1038/s41598-025-95379-y
15. Fang H, Zhang Y, Wang J, et al. Remimazolam reduces sepsis-associated acute liver injury by activation of peripheral benzodiazepine receptors and p38 inhibition of macrophages. Int Immunopharmacol. 2021;101(Pt B):108331. doi:10.1016/j.intimp.2021.108331
16. Rittirsch D, Huber-Lang MS, Flierl MA, et al. Immunodesign of experimental sepsis by cecal ligation and puncture. Nat Protoc. 2009;4(1):31–36. doi:10.1038/nprot.2008.214
17. Wen T, Wen J, Yao C. Remimazolam inhibits postoperative cognitive impairment after cardiopulmonary bypass by alleviating neuroinflammation and promoting microglia M2 polarization. Brain Res. 2024;1838:148975. doi:10.1016/j.brainres.2024.148975
18. Zhou R, Yang X, Li X, et al. Recombinant CC16 inhibits NLRP3/caspase-1-induced pyroptosis through p38 MAPK and ERK signaling pathways in the brain of a neonatal rat model with sepsis. J Neuroinflammation. 2019;16(1):239. doi:10.1186/s12974-019-1651-9
19. Zhou RX, Li -Y-Y, Qu Y, et al. Regulation of hippocampal neuronal apoptosis and autophagy in mice with sepsis-associated encephalopathy by immunity-related GTPase M1. CNS Neurosci Ther. 2020;26(2):177–188. doi:10.1111/cns.13229
20. Hu W, Zhang X, Wu Z, et al. Exploring and validating the mechanism of ulinastatin in the treatment of sepsis-associated encephalopathy based on transcriptome sequencing. J Inflamm Res. 2024;17:8753–8773. doi:10.2147/JIR.S488400
21. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15(12):550. doi:10.1186/s13059-014-0550-8
22. Maag JLV. gganatogram: an R package for modular visualisation of anatograms and tissues based on ggplot2. F1000Res. 2018;7:1576. doi:10.12688/f1000research.16409.2
23. Gu Z, Eils R, Schlesner M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics. 2016;32(18):2847–2849. doi:10.1093/bioinformatics/btw313
24. Wu T, Hu E, Xu S, et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation. 2021;2(3):100141. doi:10.1016/j.xinn.2021.100141
25. Ahmed M, Sallari RC, Guo H, et al. Variant set enrichment: an R package to identify disease-associated functional genomic regions. BioData Min. 2017;10(1):9. doi:10.1186/s13040-017-0129-5
26. Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinf. 2008;9(1):559. doi:10.1186/1471-2105-9-559
27. Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi:10.1101/gr.1239303
28. Yu G, Wang L-G, Han Y, et al. clusterProfiler: an R package for comparing biological themes among gene clusters. Omics. 2012;16(5):284–287. doi:10.1089/omi.2011.0118
29. Liberzon A, Birger C, Thorvaldsdóttir H, et al. The molecular signatures database (MSigDB) hallmark gene set collection. Cell Syst. 2015;1(6):417–425. doi:10.1016/j.cels.2015.12.004
30. Miao YR, Zhang Q, Lei Q, et al. ImmuCellAI: a unique method for comprehensive T-cell subsets abundance prediction and its application in cancer immunotherapy. Adv Sci. 2020;7(7):1902880. doi:10.1002/advs.201902880
31. Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31(2):455–461. doi:10.1002/jcc.21334
32. Peng X, Luo Z, He S, et al. Blood-brain barrier disruption by lipopolysaccharide and sepsis-associated encephalopathy. Front Cell Infect Microbiol. 2021;11:768108. doi:10.3389/fcimb.2021.768108
33. Mazeraud A, Righy C, Bouchereau E, et al. Septic-associated encephalopathy: a comprehensive review. Neurotherapeutics. 2020;17(2):392–403. doi:10.1007/s13311-020-00862-1
34. Wang W, Uchida Y, Morimoto Y. Remimazolam and dexmedetomidine prevent cognitive decline by affecting lipopolysaccharide-induced brain injury in mice: possible involvement of the vagus nerve pathway. Biomed Res. 2025;46(4):177–187. doi:10.2220/biomedres.46.177
35. Masui K. Remimazolam: its clinical pharmacology and evolving role in anesthesia and sedation practice. Curr Opin Anaesthesiol. 2024;37(4):344–351. doi:10.1097/ACO.0000000000001384
36. Grönloh MLB, Tebbens ME, Kotsi M, Arts JJ, Van Buul JD. Intercellular adhesion molecule 2 regulates diapedesis hotspots by allowing neutrophil crawling against the direction of flow. Vasc Biol. 2023;5(1).
37. Zhu Z, Ling X, Wang G, et al. G-CSFR-induced leukocyte transendothelial migration during the inflammatory response is regulated by the ICAM1-PKCa axis: based on multiomics integration analysis. Cell Biol Toxicol. 2024;40(1):90. doi:10.1007/s10565-024-09934-w
38. Zinovkina LA, Makievskaya CI, Galkin II, et al. Mitochondria-targeted uncouplers decrease inflammatory reactions in endothelial cells by enhancing methylation of the ICAM1 gene promoter. Curr Mol Pharmacol. 2024;17(1):e150823219723. doi:10.2174/1874467217666230815142556
39. Vasco C, Rizzo A, Cordiglieri C, et al. The role of adhesion molecules and extracellular vesicles in an in vitro model of the blood-brain barrier for metastatic disease. Cancers. 2023;15(11):3045. doi:10.3390/cancers15113045
40. Sieminski M, Szaruta-Raflesz K, Szypenbejl J, et al. Potential neuroprotective role of melatonin in sepsis-associated encephalopathy due to its scavenging and anti-oxidative properties. Antioxidants. 2023;12(9). doi:10.3390/antiox12091786
41. Cai M, Du B, Si Y, et al. Knockdown of VDAC1 alleviates the cognitive dysfunction secondary to sepsis-associated encephalopathy. Am J Transl Res. 2021;13(7):7538–7555.
42. Otgongerel D, Lee HJ, Jo SA. Induction of ICAM1 in brain vessels is implicated in an early AD pathogenesis by modulating neprilysin. Neuromolecular Med. 2023;25(2):193–204. doi:10.1007/s12017-022-08726-x
43. Guo C, Li W, Liu Y, et al. LCN2 induces neuronal loss and facilitates sepsis-associated cognitive impairments. Cell Death Dis. 2025;16(1):146. doi:10.1038/s41419-025-07469-4
44. Kaur S, Hussain S, Kolhe K, et al. Elevated plasma ICAM1 levels predict 28-day mortality in cirrhotic patients with COVID-19 or bacterial sepsis. JHEP Rep. 2021;3(4):100303. doi:10.1016/j.jhepr.2021.100303
45. Jin YY, Liang Y-P, Wei Z-Y, et al. Hippocampal adenosine-to-inosine RNA editing in sepsis: dynamic changes and influencing factors. Brain Commun. 2024;6(4):fcae260. doi:10.1093/braincomms/fcae260
46. Marks KE, Cho K, Stickling C, et al. Toll-like receptor 2 in autoimmune inflammation. Immune Netw. 2021;21(3):e18. doi:10.4110/in.2021.21.e18
47. Leone M, Moreau R. Leukocyte Toll-like receptor 2-mitochondria axis in sepsis: unraveling immune response sophistication. Anesthesiology. 2014;121(6):1147–1149. doi:10.1097/ALN.0000000000000471
48. Zhu Y, Zhang Y, He S, et al. Integrating single-nucleus RNA sequencing and spatial transcriptomics to elucidate a specialized subpopulation of astrocytes, microglia and vascular cells in brains of mouse model of lipopolysaccharide-induced sepsis-associated encephalopathy. J Neuroinflammation. 2024;21(1):169. doi:10.1186/s12974-024-03161-0
49. Banks WA, Hansen KM, Erickson MA, et al. High-mobility group box 1 (HMGB1) crosses the BBB bidirectionally. Brain Behav Immun. 2023;111:386–394. doi:10.1016/j.bbi.2023.04.018
50. Khan MZ, Li L, Wang T, et al. Bioactive compounds and probiotics mitigate mastitis by targeting NF-κB signaling pathway. Biomolecules. 2024;14(8):1011. doi:10.3390/biom14081011
51. Yang C, Wan J, Wang Y, et al. UBTF facilitates acute myeloid leukemia development and immune escape via PD-L1 regulation. Int Immunopharmacol. 2025;152:114433. doi:10.1016/j.intimp.2025.114433
52. Shen R, Jiang Y, Liu G, et al. Single-cell landscape of bronchoalveolar lavage fluid identifies specific neutrophils during septic immunosuppression. Adv Sci. 2025;12(11):e2406218. doi:10.1002/advs.202406218
53. Müller MM, Baldauf C, Hornischer S, et al. Staphylococcus aureus induces tolerance in human monocytes accompanied with expression changes of cell surface markers. Front Immunol. 2023;14:1046374. doi:10.3389/fimmu.2023.1046374
54. Liang G, Li J, Pu S, et al. Screening of sepsis biomarkers based on bioinformatics data analysis. J Healthc Eng. 2022;2022:6788569. doi:10.1155/2022/6788569
55. Qu G, Liu H, Li J, et al. GPX4 is a key ferroptosis biomarker and correlated with immune cell populations and immune checkpoints in childhood sepsis. Sci Rep. 2023;13(1):11358. doi:10.1038/s41598-023-32992-9
56. Morelli M, Madonna S, Albanesi C. SOCS1 and SOCS3 as key checkpoint molecules in the immune responses associated to skin inflammation and malignant transformation. Front Immunol. 2024;15:1393799. doi:10.3389/fimmu.2024.1393799
57. Li J, Yang D, Ge S, et al. Identifying hub genes of sepsis-associated and hepatic encephalopathies based on bioinformatic analysis-focus on the two common encephalopathies of septic cirrhotic patients in ICU. BMC Med Genomics. 2024;17(1):19. doi:10.1186/s12920-023-01774-7
58. Lu S, Di john Portela I, Martino N, et al. A transient brain endothelial translatome response to endotoxin is associated with mild cognitive changes post-shock in young mice. Neuroscience. 2024;555:194–204. doi:10.1016/j.neuroscience.2024.07.041
59. Lu S, Portela ID, Martino N, et al. A transient brain endothelial translatome response to endotoxin is associated with mild cognitive changes post-shock in young mice. bioRxiv. 2024;555:194–204.
60. Wang Z, Zhang Z, Shi J, et al. The crosstalk between sepsis-associated encephalopathy and Alzheimer’s disease: identifying potential biomarkers and therapeutic targets for cognition. Mol Neurobiol. 2025;62(11):13999–14014. doi:10.1007/s12035-025-05192-x
61. McComish SF, O’Sullivan J, Copas AMM, et al. Reactive astrocytes generated from human iPSC are pro-inflammatory and display altered metabolism. Exp Neurol. 2024;382:114979. doi:10.1016/j.expneurol.2024.114979
62. Kamalian A, Ho SG, Patel M, et al. Exploratory assessment of proteomic network changes in cerebrospinal fluid of mild cognitive impairment patients: a pilot study. Biomolecules. 2023;13(7):1094. doi:10.3390/biom13071094
63. Cai HQ, Weickert TW, Catts VS, et al. Altered levels of immune cell adhesion molecules are associated with memory impairment in schizophrenia and healthy controls. Brain Behav Immun. 2020;89:200–208. doi:10.1016/j.bbi.2020.06.017
64. Catalán M, González-Herrera F, Maya JD, et al. Boldine prevents the inflammatory response of cardiac fibroblasts induced by SGK1-NFκB signaling pathway activation. Cell Signal. 2024;120:111241. doi:10.1016/j.cellsig.2024.111241
65. Kim WS, Prasongyuenyong K, Ko A, et al. ICAM1(+) gingival fibroblasts modulate periodontal inflammation to mitigate bone loss. Front Immunol. 2024;15:1484483. doi:10.3389/fimmu.2024.1484483
66. Brandt A, Kromm F, Hernández-Arriaga A, et al. Cognitive alterations in old mice are associated with intestinal barrier dysfunction and induced toll-like receptor 2 and 4 signaling in different brain regions. Cells. 2023;12(17):2153. doi:10.3390/cells12172153
67. Wang L, Yang J-W, Lin L-T, et al. Acupuncture attenuates inflammation in microglia of vascular dementia rats by inhibiting miR-93-mediated TLR4/MyD88/NF-κB signaling pathway. Oxid Med Cell Longev. 2020;2020:8253904. doi:10.1155/2020/8253904
68. Zhang J, Zheng Y, Zhao Y, et al. Andrographolide ameliorates neuroinflammation in APP/PS1 transgenic mice. Int Immunopharmacol. 2021;96:107808. doi:10.1016/j.intimp.2021.107808
69. Yan B, Liao P, Cheng F, et al. Identification of toll-like receptor 2 as a key regulator of neuronal apoptosis in vascular dementia by bioinformatics analysis and experimental validation. Exp Gerontol. 2024;193:112464. doi:10.1016/j.exger.2024.112464
70. Lin F, Shan W, Zheng Y, et al. Toll-like receptor 2 activation and up-regulation by high mobility group box-1 contribute to post-operative neuroinflammation and cognitive dysfunction in mice. J Neurochem. 2021;158(2):328–341. doi:10.1111/jnc.15368
71. Xia Y, Zhang G, Kou L, et al. Reactive microglia enhance the transmission of exosomal α-synuclein via toll-like receptor 2. Brain. 2021;144(7):2024–2037. doi:10.1093/brain/awab122
72. Ma X, Wang Y, Yin H, et al. Down-regulated long non-coding RNA RMST ameliorates dopaminergic neuron damage in Parkinson’s disease rats via regulation of TLR/NF-κB signaling pathway. Brain Res Bull. 2021;174:22–30. doi:10.1016/j.brainresbull.2021.04.026
73. Linnerbauer M, Beyer T, Nirschl L, et al. PD-L1 positive astrocytes attenuate inflammatory functions of PD-1 positive microglia in models of autoimmune neuroinflammation. Nat Commun. 2023;14(1):5555. doi:10.1038/s41467-023-40982-8
74. Hang H, Xu C, Wang L, et al. Targeting PD - L1 for ischemic stroke recovery: age-dependent modulation of immune and BBB pathways. CNS Neurosci Ther. 2025;31(7):e70523. doi:10.1111/cns.70523
75. Qin T, Qin Y, Jin Y, et al. Extracellular vesicle-mediated spinal cord-brain crosstalk induces hippocampal neurogenesis impairment and cognitive deficits post-spinal cord injury. Theranostics. 2025;15(15):7584–7606. doi:10.7150/thno.110560
76. Babcock KR, Page JS, Fallon JR, et al. Adult hippocampal neurogenesis in aging and Alzheimer’s disease. Stem Cell Reports. 2021;16(4):681–693. doi:10.1016/j.stemcr.2021.01.019
77. Baker BJ, Akhtar LN, Benveniste EN. SOCS1 and SOCS3 in the control of CNS immunity. Trends Immunol. 2009;30(8):392–400. doi:10.1016/j.it.2009.07.001
78. Ji XC, Shi Y-J, Zhang Y, et al. Reducing suppressors of cytokine signaling-3 (SOCS3) expression promotes M2 macrophage polarization and functional recovery after intracerebral hemorrhage. Front Neurol. 2020;11:586905. doi:10.3389/fneur.2020.586905
79. Li R, Liu W, Yin J, et al. TSG-6 attenuates inflammation-induced brain injury via modulation of microglial polarization in SAH rats through the SOCS3/STAT3 pathway. J Neuroinflammation. 2018;15(1):231. doi:10.1186/s12974-018-1279-1
80. Zhenxuan LI, Xuerui W, Ulloa L, et al. Complementary and alternative medicine on cognitive defects and neuroinflammation after sepsis. J Tradit Chin Med. 2024;44(2):408–416. doi:10.19852/j.cnki.jtcm.20240203.002
81. Pan S, Lv Z, Wang R, et al. Sepsis-induced brain dysfunction: pathogenesis, diagnosis, and treatment. Oxid Med Cell Longev. 2022;2022(1):1328729. doi:10.1155/2022/1328729
82. Yu D, Liu J, Song X, et al. Analysis of the inflammatory storm response and heparin binding protein levels for the diagnosis and prognosis of sepsis-associated encephalopathy. Eur J Med Res. 2025;30(1):116. doi:10.1186/s40001-025-02369-x
83. Chen SL, Cai G-X, Ding H-G, et al. JAK/STAT signaling pathway-mediated microRNA-181b promoted blood-brain barrier impairment by targeting sphingosine-1-phosphate receptor 1 in septic rats. Ann Transl Med. 2020;8(21):1458. doi:10.21037/atm-20-7024
84. Dietrich JB. The adhesion molecule ICAM-1 and its regulation in relation with the blood-brain barrier. J Neuroimmunol. 2002;128(1–2):58–68. doi:10.1016/S0165-5728(02)00114-5
85. Hofer S, Bopp C, Hoerner C, et al. Injury of the blood brain barrier and up-regulation of icam-1 in polymicrobial sepsis. J Surg Res. 2008;146(2):276–281. doi:10.1016/j.jss.2007.07.021
86. Charras A, Arvaniti P, Le Dantec C, et al. JAK inhibitors and oxidative stress control. Front Immunol. 2019;10:2814. doi:10.3389/fimmu.2019.02814
87. Zhou Z, Zhou X, Dong Y, et al. Formononetin ameliorates high glucose‑induced endothelial dysfunction by inhibiting the JAK/STAT signaling pathway. Mol Med Rep. 2019;20(3):2893–2901. doi:10.3892/mmr.2019.10512
88. Zhao J, Bi W, Zhang J, et al. USP8 protects against lipopolysaccharide-induced cognitive and motor deficits by modulating microglia phenotypes through TLR4/MyD88/NF-κB signaling pathway in mice. Brain Behav Immun. 2020;88:582–596. doi:10.1016/j.bbi.2020.04.052
89. Siqueira M, Araujo APB, Gomes FCA, et al. Ethanol gestational exposure impairs vascular development and endothelial potential to control BBB-associated astrocyte function in the developing cerebral cortex. Mol Neurobiol. 2021;58(4):1755–1768. doi:10.1007/s12035-020-02214-8
90. Xiong J, Zhou H, Lu D, et al. Levetiracetam reduces early inflammatory response after experimental intracerebral hemorrhage by regulating the janus kinase 2 (JAK2)-signal transducer and activator of transcription 3 (STAT3) signaling pathway. Med Sci Monit. 2020;26:e922741. doi:10.12659/MSM.922741
91. Zhang Z, Wang L, Li F, et al. Therapeutic effects of human umbilical cord mesenchymal stem cell on sepsis-associated encephalopathy in mice by regulating PI3K/AKT pathway. J Integr Neurosci. 2022;21(1):38. doi:10.31083/j.jin2101038
92. Zhou Y, Liang Z, Xia Y, et al. Disruption of RBMS3 suppresses PD-L1 and enhances antitumor immune activities and therapeutic effects of auranofin against triple-negative breast cancer. Chem Biol Interact. 2023;369:110260. doi:10.1016/j.cbi.2022.110260
93. Miyazawa M, Yamamoto Y, Katayama K, et al. Kaposi’s sarcoma–associated herpesvirus replication and transcription activator protein activates CD274/PD-L1 gene promoter. Cancer Sci. 2023;114(4):1718–1728. doi:10.1111/cas.15673
94. Mao Q, Liang B, Leng Z, et al. Remimazolam ameliorates postoperative cognitive dysfunction after deep hypothermic circulatory arrest through HMGB1-TLR4-NF-κB pathway. Brain Res Bull. 2024;217:111086. doi:10.1016/j.brainresbull.2024.111086
95. Stelmasiak M, Słotwiński R. Infection-induced innate antimicrobial response disorders: from signaling pathways and their modulation to selected biomarkers. Cent Eur J Immunol. 2020;45(1):104–116. doi:10.5114/ceji.2020.94712
96. Anirudhan A, Paramasivam P, Murugesan R, et al. Temporal changes of NF-κB signaling pathway genes in bacterial stimulated whole blood- a host mechanism associated with sepsis. Microb Pathog. 2020;147:104415. doi:10.1016/j.micpath.2020.104415
97. Xu HB, Qin B, Zhang J, et al. Growth differentiation factor 11 relieves acute lung injury in mice by inhibiting inflammation and apoptosis. Eur Rev Med Pharmacol Sci. 2020;24(12):6908–6918. doi:10.26355/eurrev_202006_21682
98. Jin H, D’Urso V, Neuteboom B, et al. Avelumab internalization by human circulating immune cells is mediated by both Fc gamma receptor and PD-L1 binding. Oncoimmunology. 2021;10(1):1958590. doi:10.1080/2162402X.2021.1958590
99. Qin Y, Qian Y, Liu S, et al. TOX does not drive sepsis-induced T-cell exhaustion. Eur J Immunol. 2025;55(2):e202451395. doi:10.1002/eji.202451395
100. Sun H, Wang X-K, Li J-R, et al. Establishment and application of a high-throughput screening model for cell adhesion inhibitors. Front Pharmacol. 2023;14:1140163. doi:10.3389/fphar.2023.1140163
101. Shen W, Zhang X, Du R, et al. ICAM3 mediates tumor metastasis via a LFA-1-ICAM3-ERM dependent manner. Biochim Biophys Acta Mol Basis Dis. 2018;1864(8):2566–2578. doi:10.1016/j.bbadis.2018.05.002
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Exploration of Key Genes and Molecular Mechanisms in Mice with Perioperative Neurocognitive Disorders Treated with Remimazolam Based on Transcriptomics and Experimental Verification
Yu S, Chen B, Zhang M, Hu R, Luo J, Li J, Hu W, Zou X
Drug Design, Development and Therapy 2025, 19:8115-8133
Published Date: 10 September 2025