Back to Journals » Nature and Science of Sleep » Volume 13
Hypnotic Suggestions Increase Slow-Wave Parameters but Decrease Slow-Wave Spindle Coupling
Authors Beck J ![]() , Cordi MJ
, Cordi MJ ![]() , Rasch B
, Rasch B ![]()
Received 3 May 2021
Accepted for publication 22 June 2021
Published 6 August 2021 Volume 2021:13 Pages 1383—1393
DOI https://doi.org/10.2147/NSS.S316997
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah L Appleton
Jonas Beck,1 Maren Jasmin Cordi,1,2 Björn Rasch1,2
1Department of Psychology, University of Fribourg, Fribourg, Switzerland; 2Center of Competence Sleep & Health Zurich, University of Zurich, Zurich, Switzerland
Correspondence: Björn Rasch
Division of Cognitive Biopsychology and Methods, Department of Psychology, University of Fribourg, Rue P.-A-de-Faucigny 2, Fribourg, CH-1700, Switzerland
Tel +41 26 300 7637
Email [email protected]
Purpose: Sleep, in particular slow-wave sleep, is beneficial for memory consolidation. In two recent studies, a hypnotic suggestion to sleep more deeply increased the amount of slow-wave sleep in both a nap and a night design. In spite of these increases in slow-wave sleep, no beneficial effect on declarative memory consolidation was found. As coupling of slow-waves and sleep spindles is assumed to be critical for declarative memory consolidation during sleep, we hypothesized that the missing memory benefit after increased SWS could be related to a decrease in slow-wave/spindle coupling.
Participants and Methods: Data from 33 highly hypnotizable subjects were retrieved from a nap (n = 14) and a night (n = 19) study with a similar design and procedure. After an adaptation session, subjects slept in the sleep laboratory for two experimental sessions with polysomnography. Prior to sleep, a paired-associate learning task was conducted. Next, subjects either listened to a hypnotic suggestion to sleep more deeply or to a control text in a randomized order according to a within-subject design. After sleep, subjects performed the recall of the memory task. Here, we conducted a fine-grained analysis of the sleep data on slow-waves, spindles and their coupling.
Results: In line with our hypothesis, listening to a hypnosis tape decreased the percentage of spindles coupled to slow-waves. Slow-wave parameters were consistently increased, but sleep spindles remained unaffected by the hypnotic suggestion.
Conclusion: Our results suggest that selectively enhancing slow-waves without affecting sleep spindles might not be sufficient to improve memory consolidation during sleep.
Keywords: memory consolidation, learning, SWS, co-occurrence, slow-oscillation, hypnosis
Introduction
Sleep is beneficial for the consolidation of memory.1 In particular deep sleep, also called slow-wave sleep (SWS) is characterized by cortical slow-waves (measured with scalp electroencephalogram, EEG) and assumed to play a functional role in memory consolidation during sleep. According to the synaptic down-selection theory, neural slow oscillations (<1 Hz) and their excitatory up- and inhibitory down-states provide the basis for net synaptic downscaling of synapses, thereby increasing signal-to-noise ratio and improving memory recall the next day.2 Thus, increasing SWS and slow oscillations should lead to a better memory recall the next day. According to the active systems consolidation theory, slow oscillations provide a synchronizing time frame for reactivation-related hippocampal sharp wave ripples (transient network oscillations with 80–200 Hz) and sleep spindles. Sleep spindles are short bursts (0.5–3 s) of oscillatory activity (9–16 Hz) with a waxing and waning shape in the surface sleep EEG. In particular, the theory assumes that the interplay and precise timing of these sleep oscillations is crucial for the beneficial effect of sleep on memory consolidation.1,3–5 Thus, increasing SWS and slow oscillations should only lead to a better memory consolidation during sleep when the precise timing between spindles, ripples and slow oscillations is also maintained.
One parameter used to describe the interplay between these oscillations on the cortical level is slow-wave/spindle coupling (SW/SP coupling). Importantly, several studies have shown that increased SW/SP coupling is related to better sleep-mediated memory consolidation, for example, in ageing.6,7 Pharmacologically increasing coupling between slow-waves and spindles improved verbal memory in humans.8 In addition, optogenetic induction of thalamic spindles phase locked to the neural slow oscillation up-state has been shown to enhance coupling of spindles, slow oscillations and ripples as well as improve consolidation of hippocampus dependent memory.9 Furthermore, precision of SW/SP coupling has been linked to the strength of endogenous memory reactivations, which were associated with memory consolidation in an episodic memory task.10 Thus, SW/SP coupling might be crucial for benefits of sleep on memory consolidation.
In two of our recent studies, we reported a prolongation of SWS which did not translate into improvements in declarative memory consolidation during sleep.11,12 The increase in SWS was induced in healthy, medium-to-highly hypnotizable participants using a hypnotic suggestion to sleep more deeply before sleep. One study was conducted as a midday nap, the other as a nighttime sleep study. While the increase in SWS was significant in both studies compared to a within-subject control condition, we did not find a beneficial effect of the hypnosis tape on the performance in a declarative paired-associate learning task. However, no fine-grained analysis on slow-waves, spindles and SW/SP coupling was conducted. Thus, we reanalyzed the data of the two studies to examine whether changes in slow-waves, sleep spindles, and in particular their coupling, could explain why memory consolidation was not enhanced after increasing the amount of SWS. We expect the hypnotic suggestion to not increase SW/SP coupling in spite of increasing cortical slow-waves.
In line with our hypothesis, we found that the amount of coupling between both frontal slow spindles and parietal fast spindles with slow-waves was reduced when subjects listened to a hypnosis tape to sleep more deeply prior to sleep. Generally, hypnotic suggestions increased the density of slow-waves and several other slow-wave parameters, whereas overall spindle density was comparable between the two sessions.
Materials and Methods
We used data from two previous studies with a similar design and procedure. Both studies followed a within-subject cross-over design to compare the effects of a deep sleep hypnosis tape vs a control tape on sleep depth of high and low hypnotizable participants. The first study was conducted as a midday nap study in German-speaking females and will hereinafter be referred to as the “nap study”.12 The second study assessed a full night of sleep including both sexes and will hereinafter be referred to as the “night study”.11 For the purposes of this study, we only analyzed data from the high hypnotizable subjects.
Subjects
In the nap study, 14 healthy, German-speaking young females (mean age, 23.36 ± 2.65 years [M ± SD], age range 18–29 years) participated in the experiment. In the night study, 19 healthy, French-speaking young subjects participated in the experiments (9 males, M = 22.32 ± 3.06 years, age range 19–31 years) and mean age was comparable between studies (p = 0.14). None of the subjects were shift workers nor had they been on any intercontinental flights six weeks prior to the experiment. They neither took any sleep influencing medication nor reported any neurological, psychiatric or sleep-related disorders. All participants refrained from drinking alcohol and caffeine on experimental days. Subjects were payed 140 CHF in the nap and 150 CHF in the night study for participation in all sessions. The studies were approved by the Ethics Committees of the University of Zurich (nap study) and Lausanne (night study). Hypnotizability was assessed by a German version of the Harvard Group Scale of Hypnotic Susceptibility (HGSHS)13 in the nap study and by a French version translated by Laurent Rossier, a German- and French-speaking hypnotherapist, in the night study.11 The HGSHS was conducted in a group session prior to the experiment with a cutoff score of ≥7 for highly hypnotizable subjects (nap: HGSHS = 7.71 ± 0.73; night: HGSHS = 7.36 ± 0.69). The percentage of high hypnotizable subjects within the initial screening was 35% in the nap study and 33% in the night study.
Design and Procedure
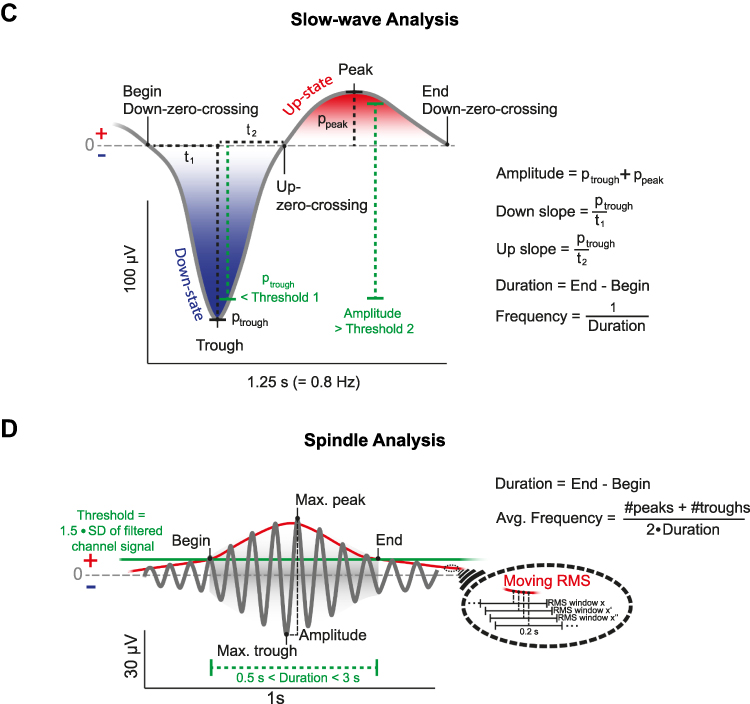
The design of the nap study equaled the design of the night study with the exception of sleep, which either occurred during a midday nap or during the night.11,12 Subjects participated in four sessions, three of which were sleep sessions. During a first group session, participants were informed that a hypnosis tape will be used to deepen their sleep and hypnotizability was assessed using the HGSHS.11,13 After an adaptation nap or night, participants slept in the laboratory for two sessions, while polysomnographic data (electroencephalography (EEG), electromyography (EMG) and electrooculography (EOG)) was recorded. Both experimental sessions took place on the same weekday, spaced one week apart. Prior to sleep, a declarative paired-associate learning task (PAL) was conducted. In a randomized order, participants listened to either a hypnotic suggestion tape to sleep more deeply or a control tape about natural mineral deposits in a within-subject design (see Figure 1A, for an overview of the procedure). They were asked to listen to the tape but were allowed to fall asleep at any time. The tape was started with the lights switched off and subjects were awoken 90 min (nap) or 8 h (night) later. After sleep, subjects performed a psychomotor vigilance task, filled out a subjective sleep quality questionnaire (Schlaffragebogen A)14 and conducted the recall of the PAL. After the PAL in the nap study participants performed a procedural sequence finger tapping task whereas after the PAL in the night study subjects performed two declarative verbal fluency tasks. Figure 1 Continued. Figure 1 Experimental procedure and analysis of sleep-oscillations. (A) Thirty-three high hypnotizable subjects listened to a hypnosis (blue) or a control tape (red) while falling asleep according to a within-subject design. In both sessions participants either slept during a midday nap (90 min) or during the night (8h). Learning of a declarative paired associate learning task (PAL) was conducted before sleep and the recall of the same task after sleep. Analyses of slow-waves, sleep spindles and their coupling were conducted using the open-source SpiSOP tool (www.spisop.org; RRID: SCR_015673). (B) Coupling analyses were performed within the time window starting with the trough of the slow-wave until 1.2 s after this trough for slow and fast spindles. We identified coupling events where at least one detected sleep spindle trough (maximum negative amplitude) was detected within the specified time window. As a result, the number of matches and the percentage of matches relative to the total amount of slow-waves were calculated for each participant and lobe (frontal slow spindles, parietal fast spindles) during NREM sleep. (C) For detection of slow-waves, the signal was initially filtered between 0.3 and 3.5 Hz in each channel during NREM sleep. Second, all intervals with consecutive positive-to-negative zero crossings (down – up – down) were marked as putative slow-waves. They were only kept for further analyses, if their durations matched with a frequency between 0.5–1.11 Hz, while down-zero-crossings marked the begin and end of a slow-wave. Possible matches were considered as artifacts and excluded if their amplitude exceeded 1000 µV, or if both negative and positive half-wave amplitudes lay between –15 and +10 µV. Finally, a slow-wave was identified if the following two criteria were met: the negative half-wave peak potential (ptrough) was lower than 1.25 × the mean negative half-wave peak of all allegedly detected slow-waves within the respective EEG channel (Threshold 1); the amplitude (trough to peak) was larger than 1.25 × the mean amplitude of all other allegedly detected slow-waves within this channel (Threshold 2). As a result, number of slow-waves, density per 30s epoch NREM sleep, mean amplitude, duration, down slope (value of the negative half-wave peak divided by the time from the first zero-crossing to the trough in µV/s) and up slope (absolute value of the negative half-wave peak divided by the time from the trough to the next zero-crossing in µV/s) were calculated for each participant and channel during NREM sleep. (D) For the detection of sleep spindles, the EEG signal of each channel during NREM sleep was filtered with the frequency determined by an individual slow- and fast spindle power peak ± 1 Hz. Next, the root mean square (RMS) signal was computed using a sliding window with a size of 0.2 s and the resulting signal was smoothed in the same window with a moving average. A spindle was detected when the smoothed RMS signal exceeded an individual amplitude threshold by 1.5 standard deviations (SD) of the filtered channel signal for 0.5 to 3 s at least once. The threshold crossings marked the beginning and end of each spindle event and determined their duration. Amplitude of sleep spindles was defined by the voltage difference between the largest trough and peak (peak to trough potential). Spindles with an amplitude above 200 µV were excluded. Analysis resulted in data of the number of detected spindles, spindle density (per 30s NREM sleep epoch), mean amplitude (trough to peak potential), average oscillatory frequency (Avg. Frequency using number of peaks (#peaks) and number of troughs (#troughs)) and duration for each participant and channel (frontal slow spindles, parietal fast spindles) during NREM sleep. (B–D) Figures were retrieved from www.spisop.org/documentation/ and adapted with permission from the copyright owner (Frederik D. Weber).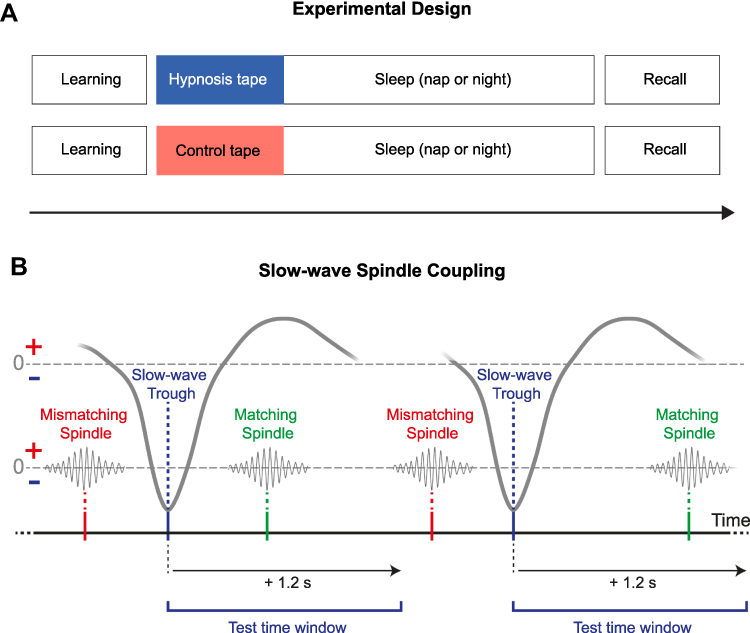
Hypnosis and Control Tape
The hypnosis text was written by a hypnotherapist treating sleep problems and sleep disorders with hypnosis (A. Schlarb).12 The hypnosis text started with focusing the attention of the listener from the environment to internal processes, including suggestions to close the eyes and to relax. Next, it was slowly counted from 1 to 10 with each step indicating deeper relaxation and leading the listener into the hypnotic trance state (4 min). Subsequently, the listener was invited to imagine a picture of a sea and to follow a fish swimming progressively deeper and deeper into the sea, symbolizing the depth of sleep (9 min). This was accompanied by suggestions that swimming deeper and deeper is safe and risk free. When the fish arrived at the bottom of the sea, the listener was suggested to continue to sleep deeply. Afterwards, the tape stopped and left the listener in the hypnotic trance, which invited them to fall asleep. The control text was designed to match the hypnosis text with respect to length in minutes and volume and to be as neutral and objective as possible. The German tapes were spoken and recorded by Björn Rasch,12 the French versions were translated, spoken and recorded by Laurent Rossier et al.11 The hypnosis texts were spoken with a soft and calming voice, while the control texts were spoken with an everyday intonation and speed. The tapes are available on our homepage: https://www.unifr.ch/psycho/en/research/biopsy/hypnosis.html.
Paired-Associate Learning Task
A German and a French version of the PAL were conducted similarly to previous studies.15 In the evening, participants learned 80 semantically associated word pairs (eg, clock – church) and were asked to memorize as many pairs as possible. During this learning phase, the word pairs were presented sequentially for 1000 ms in black font on a white screen. A blank screen of 200 ms separated the two words of a word pair. Each trial was preceded by an interstimulus interval of 500 ms and a fixation cross of 500 ms. After the learning phase, subjects retrieved the word pairs in a cued-recall task. The first word of the word pair was presented and they were asked to name the corresponding second word with unlimited response time; if they were unable to recall the matching word they were asked to click “next”. The first word was presented in black font on a white screen for 1000 ms followed by a question mark. The experimenter noted if the answer was correct without giving feedback to the participant. Another blank screen of 500 ms preceded the next trial. Selection of the word list and the order of word pair presentation was randomized. In the morning, memory performance was tested a second time using the cued-recall task without a learning phase. Memory performance was measured as the percentage of correctly recalled word pairs before and after the sleep period. To assess relative overnight memory improvement, pre-sleep performance was set to 100%.
Polysomnographic Recording
EEG was recorded using a 128-channel Geodesic Sensor Net (nap, Electrical Geodesics, Eugene, OR) or a 32-channels Easycap Net (night, Easycap GmbH, Herrsching) with a sampling rate of 500 Hz in both studies. Offline EEG preprocessing was conducted using BrainVision Analyzer software (version 2.0 in the nap and 2.1 in the night study; Brain Products, Gliching, Germany). Data were referenced against contralateral mastoids in the nap study and against average mastoids in the night study. Standard filter settings suggested by the American Academy of Sleep Medicine (AASM)16 were applied (eg, EEG 0.3–35Hz) with an additional notch filter (50 Hz) in both datasets. Sleep was visually scored offline by two independent sleep scorers in 30 second epochs, based on derivations F4, C4, O2, HEOG, VEOG and EMG according to the AASM standard criteria.16 In case of disagreement, a third expert was consulted. All scorers were blind to the experimental condition.
Detection of Spindles, Slow-Waves and Their Coupling
Analysis of slow-waves, spindles and their coupling was performed on frontal (F3, F4) and parietal (P3, P4) electrodes using the open-source SpiSOP tool (www.spisop.org; RRID: SCR_015673) based on MATLAB 2013b (MathWorks, Natick, USA; RRID: SCR_001622). Standard settings of SpiSOP were applied based on previously published algorithms17 and are explained in the following.
Slow-Wave Detection
First, the signal was first filtered in each channel during NREM sleep between 0.3 and 3.5 Hz. Second, all intervals with consecutive positive-to-negative zero crossings (down – up – down) were marked as putative slow-waves. They were only kept for further analyses if their durations matched with a frequency between 0.5–1.11 Hz, while down-zero-crossings marked the begin and end of a slow-wave. Possible matches were considered as artifacts and excluded, if their amplitude exceeded 1000 µV or both negative and positive half-wave amplitudes lay between –15 and +10 µV. Finally, a slow-wave was identified if the following two criteria were met: the negative half-wave peak potential was lower than 1.25 × the mean negative half-wave peak of all allegedly detected slow-waves within the respective EEG channel (see Figure 1C, Threshold 1); the amplitude (trough to peak) was larger than 1.25 × the mean amplitude of all other allegedly detected slow-waves within this channel (see Figure 1C, Threshold 2). As a result, number of slow-waves, density per 30s epoch NREM sleep, mean amplitude, duration, down slope (value of the negative half-wave peak divided by the time from the first zero-crossing to the trough in µV/s) and up slope (absolute value of the negative half-wave peak divided by the time from the trough to the next zero-crossing in µV/s) were calculated for each participant and channel during NREM sleep.
Spindle Detection
First, individual slow and fast spindle frequency peaks were visually determined based on the NREM power spectrum of each dataset. Slow spindle peaks were determined in frontal channels (F3, F4) and fast spindle peaks in parietal channels (P3, P4) due to expected power maxima over those regions.18 Slow spindle peaks ranged between 9.5 and 13.2 Hz with an average frequency of 11.99 ± 0.79 Hz in the nap and 11.13 ± 0.79 Hz in the night study. Fast spindle peaks ranged between 11.5 and 15.2 Hz with an average frequency of 13.99 ± 0.71 Hz in the nap and 13.46 ± 0.40 Hz in the night study.
Second, the EEG signal of each channel during NREM sleep was filtered with the frequency determined by an individual slow- and fast spindle power peak ± 1 Hz. Next, the root mean square (RMS) signal was computed using a sliding window with a size of 0.2 s and the resulting signal was smoothed in the same window with a moving average (see Figure 1D). A spindle was detected, when the smoothed RMS signal exceeded an individual amplitude threshold by 1.5 standard deviations (SD) of the filtered channel signal once for 0.5 to 3 s. The threshold crossings marked the beginning and end of each spindle event and determined their duration. Amplitude of sleep spindles was defined by the voltage difference between the largest trough and peak (peak to trough potential). Spindles with an amplitude above 200 µV were excluded. Analysis resulted in data of the number of detected spindles, spindle density (per 30s NREM sleep epoch), mean amplitude (trough to peak potential), average oscillatory frequency and duration for each participant and channel (frontal slow spindles, parietal fast spindles) during NREM sleep.
Coupling of Slow-Waves and Spindles
After detecting slow-waves and spindles, additional analyses were conducted to identify the coupling between slow-waves and spindles (see Figure 1B). These coupling analyses were performed within the time window starting with the trough of the slow-wave until 1.2 s after this trough for slow and fast spindles. We identified coupling events where at least one detected sleep spindle trough (maximum negative amplitude) was detected within the specified time window. As a result, the number of matches and the percentage of matches relative to the total amount of slow-waves were calculated for each participant and lobe (frontal slow spindles, parietal fast spindles) during NREM sleep.
Statistical Analysis
Statistical analysis was performed using RStudio version 1.1.456.19 Normality was tested using Shapiro–Wilk’s test. As slow-wave density (W = 0.99, p < 0.001) and spindle density (W = 0.98, p < 0.001) were both non normally distributed, Tukey’s rule20 using the interquartile range (IQR, [Q1 – 1.5 × IQR, Q3 + 1.5 × IQR]) was applied to detect possible outliers in non-normally distributed data. Application of this outlier criterion on the slow-wave and spindle density data of frontal and parietal electrodes from both studies suggested a slow-wave density range between –0.10 and −3.83 and a spindle density range between 0.75 and 3.58. Based on these criteria, three datapoints were excluded in the spindle and coupling analysis and replaced with data from the contralateral hemisphere. The same corrections were applied to the analyses of all other spindle and coupling parameters.
We analyzed slow-wave density and spindle density as well as coupling between slow-waves and spindles. Analyses were conducted separately over the frontal and over the parietal lobes. To compare the results of the nap and the night study, we only analyzed data within the first hour of sleep in the night study. We calculated mixed analyses of variances containing the within subject factors condition (control vs hypnosis) and hemisphere (left vs right) as well as the between subject factor study (nap vs night). Post-hoc tests comprised paired t-tests and Welch t-tests for unpaired data. In case of significant findings, partial eta-squared (ηp) and Cohen's d are reported as effect sizes for analyses of variances and t-tests, respectively. For correlational analysis, we used Pearson product-moment correlations. Correlations were compared according to21 based on single sided testing. The level of significance was set to p < 0.05.
Results
Slow-Wave Sleep and Memory Consolidation
In a first analysis, we reanalyzed SWS and memory consolidation data together for both the nap and night study. After listening to the suggestion to sleep more deeply, participants spent 29.95 ± 2.35% of their total sleep time in SWS, whereas in the control condition subjects only spent 22.73 ± 2.30% of their total sleep time in SWS. Thus, the hypnotic suggestion increased the time spent in SWS to 131.76% compared with listening to the control text (set to 100%; F(1, 31) = 13.94, p < 0.001, ηp = 0.31). This increase was more pronounced in the nap compared with the night study (interaction condition × study: F(1, 31) = 6.77, p = 0.014, ηp = 0.18). Adding age as a covariate yielded the same interaction between condition and study (F(1, 30) = 10.18, p = 0.003, ηp = 0.25). Moreover, an interaction between age and condition (F(1, 30) = 5.88, p = 0.022, ηp = 0.16) suggested that the effect of condition was stronger in younger participants. However, the correlation between age and the effect of hypnosis on SWS did not reach significance (r(31) = –0.28, p = 0.11).
In contrast to this substantial increase in SWS, we did not observe any effect on declarative memory consolidation during sleep. Retention performance in the paired-associate word learning task (with performance during encoding set to 100%) was comparable between the hypnosis condition (98.44 ± 2.07%) and the control condition (99.42 ± 2.07%; F(1, 30) = 0.06, p = 0.81). Furthermore, we observed no significant interaction between condition and study (nap vs night; F(1, 30) = 1.58, p = 0.22)). Interestingly, the amount of SWS showed a trend for a negative correlation between SWS and memory retention performance in the hypnosis condition (r(30) = –0.33, p = 0.061), while no such correlation was found in the control session (r(30) = –0.09, p = 0.61). These correlations were significantly different from each other (z = –3.87, p < 0.001).
Effect of Hypnosis on Coupling Between Slow-Waves and Spindles
As coupling of slow-waves and sleep spindles is assumed to play a functional role for memory consolidation during sleep, we tested whether the missing effect on sleep-mediated memory consolidation in the hypnosis condition might be explained by a reduction in SW/SP coupling. We calculated SW/SP coupling as the percentage of co-occurring slow-waves and sleep spindles (time period: 1.2 seconds after slow-wave trough) relative to the total amount of slow-waves. As predicted, we observed a significant reduction in SW/SP coupling when participants listened to a hypnotic suggestion to sleep more deeply. Over the frontal lobe, participants exhibited a significantly lower amount of coupling of slow spindles and SWs when they listened to the hypnosis (14.40 ± 0.96%) compared with a control text before sleep (17.78 ± 1.23%; (F(1, 31) = 13.12, p = 0.001, ηp = 0.30; Figure 2A). Similarly, over the parietal cortex, coupling of fast spindles and slow-waves was reduced after listening to the hypnosis (12.67 ± 0.84%) compared with a control text before sleep (15.09 ± 1.21%; F(1, 30) = 4.58, p = 0.041, ηp = 0.13; Figure 2B).
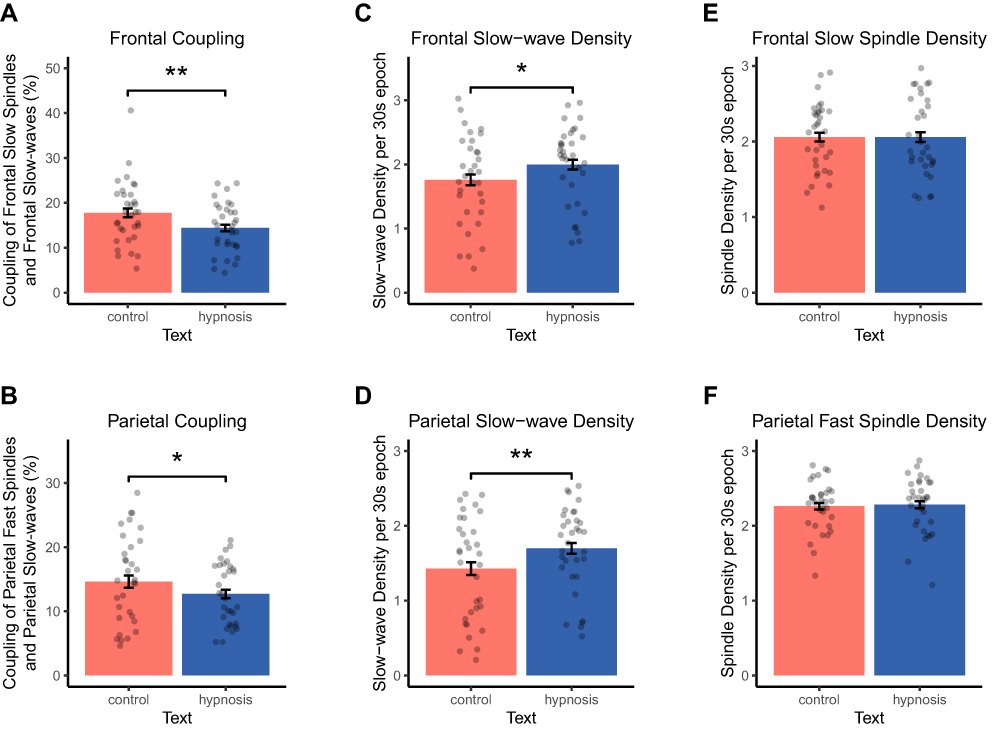 |
Figure 2 Results of coupling, slow-wave and spindle analysis. Data are shown separately for the control (red bars) and the hypnosis condition (blue bars). (A and B) Here provided are the percentages of matches between slow-waves and spindles relative to the total amount of slow-waves in NREM sleep. Listening to a hypnotic suggestion to sleep more deeply reduced (A) coupling of frontal slow-waves and slow spindles and (B) coupling of parietal slow-waves and fast spindles compared with a control tape. Listening to a hypnotic suggestion to sleep more deeply increased (C) frontal as well as (D) parietal slow-wave density per 30s epoch of NREM sleep compared with a control tape. Listening to a hypnotic suggestion to sleep more deeply did neither affect (E) frontal slow spindle nor (F) parietal fast-spindle density per 30s epoch of NREM sleep compared with a control tape. Values are displayed as mean ± SEM. **p < 0.01, *p < 0.05. |
To further examine the underlying mechanisms of the reduced SW/SP coupling after hypnosis, we analyzed the density per 30s epoch of slow-waves and sleep spindles separately during NREM sleep (stages N2 and N3 combined). In accordance with the increases in SWS induced by hypnotic suggestions, slow-wave density over the frontal lobe was significantly increased in the hypnosis condition (2.00 ± 0.11 slow-waves/30s) compared with the control condition (1.76 ± 0.12 slow-waves/30s; F(1, 66) = 6.62, p = 0.015, ηp = 0.18; Figure 2C). Similarly, over the parietal cortex, participants showed an increased parietal slow-wave density in the hypnosis (1.70 ± 0.10 slow-waves/30s) compared with the control condition (1.42 ± 0.12 slow-waves/30s; F(1, 31) = 9.65, p = 0.004, ηp = 0.24; Figure 2D). Generally, slow-wave density was higher in the night study compared with the nap study, over both frontal and parietal regions (both p < 0.001). In addition to slow-wave density, significant main effects of condition (control vs hypnosis) were found for all other slow-wave parameters except the duration of frontal slow-waves (see Table 1). Listening to a hypnotic suggestion before sleep significantly increased the number, amplitude, down slope and up slope of frontal and parietal slow-waves as well as the duration of parietal slow-waves (all p < 0.038). These increases were less pronounced over the frontal lobe in the night study and more consistent across studies over the parietal lobe (see Table 1). The most robust effects across both studies were found for the amplitude of parietal slow-waves, which were significantly increased in the hypnosis compared with the control condition in the nap (t(13) = –3.35, p = 0.005, d = 0.90) and in the night study (t(18) = –3.08, p = 0.006, d = 0.71).
 |
Table 1 Coupling, Slow-Wave and Spindle Parameters in the Hypnosis and Control Session for the Nap and Night Study |
In contrast to slow-wave density, we did not observe any differences between the hypnosis and control condition for sleep spindles. Both frontal slow spindle density (hypnosis: 2.06 ± 0.09 spindles/30s; control: 2.06 ± 0.08 spindles/30s; Figure 2E) and parietal fast spindle density (hypnosis: 2.28 ± 0.06 spindles/30s; control: 2.26 ± 0.06 spindles/30s; Figure 2F), were comparable between the two experimental conditions (both p > 0.55). In addition, and contrary to parietal slow-waves, listening to a hypnotic suggestion before sleep decreased the duration of parietal fast spindles (p = 0.033, see Table 1). Together with the lack of effect on other spindle parameters and the increases found for slow-wave parameters, these results might facilitate and explain a reduced coupling of slow-waves and spindles.
Discussion
In the current study, we show that hypnotic suggestion before sleep promotes slow-waves, but leaves spindles unaffected and hence results in a decreased SW/SP coupling during both a nap and nighttime sleep. Previous research, which experimentally enhanced slow-waves, suggested a causal contribution of slow-waves to the consolidation of hippocampus-dependent declarative memory.22–24 Here, we found that listening to a hypnotic suggestion to sleep more deeply enhances slow-waves by increasing the number, density, amplitude, up- and down-slopes in highly hypnotizable participants. Despite these improvements, we did not find any effect on declarative memory consolidation during sleep. The current findings offer an explanation for this lack of memory consolidation effect as slow – as well as fast spindles remained unaffected and coupling between slow-waves and spindles was decreased in the hypnosis condition.
Sleep spindles are assumed to represent the reinstatement of a memory trace with reactivation occurring during spindle events.25–27 Spindle activity is increased when memory cues associated with previously learned content are presented during sleep.26 During this increase in spindle activity, memory content associated with the stimuli presented could be reliably decoded, which further strengthens the view of a reactivation of memory content during the occurrence of spindles. In addition, results from a simultaneous EEG and fMRI study support the notion that cortical reactivation occurs during spindles.27 Therefore, an invariant amount of sleep spindles in the control and hypnosis condition might represent similar amounts of reactivation and consolidation during sleep, and thereby explain the comparable amounts of memory consolidation in the control and hypnosis condition.
After reinstatement of a memory trace represented by a spindle event, additional processing is assumed to occur during a spindle refractory period (3–6 s).25 This refractory period is thought to be crucial for protecting memory reprocessing from interference and spindle refractory periods are thought to optimize oscillatory interactions supporting systems consolidation. Increasing slow-waves within these refractory periods might be ineffective for promoting memory consolidation and possibly even disturb oscillatory interactions favoring memory consolidation.
Besides findings that slow-waves and spindles are relevant for memory consolidation, SW/SP coupling has repeatedly been reported to be crucial for the beneficial effect of sleep.6,9,10 Animal studies using two-photon imaging have shown that the co-occurrence of a spindle within the up-state of a slow-wave might optimize conditions for synaptic plasticity in local cortical circuits by maximizing calcium activity in excitatory pyramidal cells.28 Thus, one conclusion could be that enhancing slow-waves or changes in their characteristics due to hypnotic suggestions alone are not sufficient to benefit memory if spindles and their coupling are not enhanced in a similar way. This notion is consistent with the assumption that the interplay and precise timing between these sleep oscillations is crucial for the beneficial effect of sleep on memory consolidation.3–5
Our interpretation is in line with the active system consolidation hypothesis,1 which stresses the importance of the precise timing between different types of sleep oscillations in the consolidation of memory. In contrast, the missing effect on memory consolidation despite increases in slow-waves are difficult to explain via the synaptic down-selection theory,2 where slow-waves are thought to directly contribute to synaptic depression in two possible scenarios: burst firing, which occurs during transitions between up and down states leading to enduring depression of excitatory postsynaptic potentials;29 decoupling through synchronous burst firing during slow-waves by spike-timing-dependent plasticity mechanisms.30 Within this framework, increasing slow-waves should increase the activity-dependent down-selection of synapses, decrease the net synaptic strength, restore cellular homeostasis and thereby favor memory consolidation during sleep.
Besides the interpretation that spindles and their coupling to slow-waves is essential for improvements in memory consolidation, another potential factor has been discussed in the literature. The pre-sleep performance level in the declarative memory task might have been not optimal (37.57–43.13%) to profit from sleep dependent memory consolidation31–33 and an additional recall trial with feedback before sleep or repeated learning until reaching 60% could have benefitted memory consolidation.15,34,35
Conclusion
The present study shows that selectively enhancing slow-waves using a hypnotic suggestion without affecting sleep spindles is not sufficient to support memory consolidation during sleep. Our results are in line with the notion that the interplay and timing between slow-waves and sleep spindles is essential for the beneficial effect of sleep on memory consolidation. Future studies examining memory consolidation during sleep have to consider and target slow-waves in conjunction with spindles and especially their coupling during sleep.
Abbreviations
AASM, American Academy of Sleep Medicine; EEG, electroencephalography; EMG, electromyography; EOG, electrooculography; HGSHS, Harvard Group Scale of Hypnotic Susceptibility; NREM sleep, Non-rapid eye movement sleep, ie, N2 sleep and slow-wave sleep; PAL, paired-associate learning task; SW/SP coupling, slow-wave/spindle coupling; SWS, Slow-wave sleep.
Acknowledgment
We would like to thank Louisa Clarke for helpful comments on an earlier version of this article.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
This work was supported by a grant of the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (grant agreement number 667875), a grant of the Swiss National Foundation (PP00P1_133685) and the Clinical Research Priority Program (CRPP) “Sleep and Health” of the University of Zurich, Institute of Psychology, Department of Biopsychology, Zurich, Switzerland. The authors declare no conflicts of interest for this work.
References
1. Rasch B, Born J. About sleep’s role in memory. Physiol Rev. 2013;93(2):681–766. doi:10.1152/physrev.00032.2012
2. Tononi G, Cirelli C. Sleep and the price of plasticity: from synaptic and cellular homeostasis to memory consolidation and integration. Neuron. 2014;81(1):12–34. doi:10.1016/j.neuron.2013.12.025
3. Fernandez LMJ, Lüthi A. Sleep spindles: mechanisms and functions. Physiol Rev. 2020;100(2):805–868. doi:10.1152/physrev.00042.2018
4. Peyrache A, Seibt J. A mechanism for learning with sleep spindles. Philos Trans R Soc Lond B Biol Sci. 2020;375(1799):20190230. doi:10.1098/rstb.2019.0230
5. Schreiner T, Staudigl T. Electrophysiological signatures of memory reactivation in humans. Philos Trans R Soc Lond B Biol Sci. 2020;375(1799):20190293. doi:10.1098/rstb.2019.0293
6. Helfrich RF, Mander BA, Jagust WJ, Knight RT, Walker MP. Old brains come uncoupled in sleep: slow wave-spindle synchrony, brain atrophy, and forgetting. Neuron. 2018;97(1):221–230.e4. doi:10.1016/j.neuron.2017.11.020
7. Muehlroth BE, Sander MC, Fandakova Y, et al. Precise slow oscillation-spindle coupling promotes memory consolidation in younger and older adults. Sci Rep. 2019;9(1):1940. doi:10.1038/s41598-018-36557-z
8. Niknazar M, Krishnan GP, Bazhenov M, Mednick SC. Coupling of thalamocortical sleep oscillations are important for memory consolidation in humans. PLoS One. 2015;10(12):e0144720. doi:10.1371/journal.pone.0144720
9. Latchoumane C-FV, Ngo H-V, Born J, Shin H-S. Thalamic spindles promote memory formation during sleep through triple phase-locking of cortical, thalamic, and hippocampal rhythms. Neuron. 2017;95(2):424–435.e6. doi:10.1016/j.neuron.2017.06.025
10. Schreiner T, Petzka M, Staudigl T, Staresina BP. Endogenous memory reactivation during sleep in humans is clocked by slow oscillation-spindle complexes. Nat Commun. 2021;12(1):3112. doi:10.1038/s41467-021-23520-2
11. Rossier L, Cordi MJ, Rasch B. Hypnotic suggestions given before nighttime sleep extend slow-wave sleep as compared to a control text in highly hypnotizable subjects. Int J Clin Exp Hypn. 2020;68(1):105–129. doi:10.1080/00207144.2020.1687260
12. Rasch B, Cordi MJ, Schlarb AA. Deepening sleep by hypnotic suggestion. Sleep. 2014;37(6):
13. Bongartz W. German norms for the harvard roup scale of hypnotic susceptibility, form A. Int J Clin Exp Hypn. 1985;33(2):131–139. doi:10.1080/00207148508406643
14. Görtelmeyer R. SF-A/R Und SF-B/R: Schlaffragebogen a Und B: Hogrefe. 2011.
15. Rasch B, Born J, Gais S. Combined blockade of cholinergic receptors shifts the brain from stimulus encoding to memory consolidation. J Cogn Neurosci. 2006;18(5):793–802. doi:10.1162/jocn.2006.18.5.793
16. Iber C, Ancoli-Israel S, Chesson A, Quan SF. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications. Westchester, IL: American Academy of Sleep Medicine; 2007:4849.
17. Mölle M, Marshall L, Gais S, Born J. Grouping of spindle activity during slow oscillations in human non-rapid eye movement sleep. J Neurosci. 2002;22(24):10941–10947. doi:10.1523/JNEUROSCI.22-24-10941.2002
18. Mölle M, Bergmann TO, Marshall L, Born J. Fast and slow spindles during the sleep slow oscillation: disparate coalescence and engagement in memory processing. Sleep. 2011;34(10):1411–1421. doi:10.5665/SLEEP.1290
19. RStudio Team. RStudio: integrated development environment for R. Boston, MA; 2015. Available from: http://www.rstudio.com/.
20. Tukey JW. Exploratory Data Analysis. 1977.
21. Lenhard W, Lenhard A. Testing the Significance of Correlations. Psychometrica; 2014. Available from: http://www.psychometrica.de/correlation.html. doi:10.13140/RG.2.1.2954.1367
22. Ngo H-V, Martinetz T, Born J, Mölle M. Auditory closed-loop stimulation of the sleep slow oscillation enhances memory. Neuron. 2013;78(3):545–553. doi:10.1016/j.neuron.2013.03.006
23. Marshall L, Helgadóttir H, Mölle M, Born J. Boosting slow oscillations during sleep potentiates memory. Nature. 2006;444(7119):610–613. doi:10.1038/nature05278
24. Marshall L, Mölle M, Hallschmid M, Born J. Transcranial direct current stimulation during sleep improves declarative memory. J Neurosci. 2004;24(44):9985–9992. doi:10.1523/JNEUROSCI.2725-04.2004
25. Antony JW, Schönauer M, Staresina B, Cairney SA. Sleep spindles and memory reprocessing. Trends Neurosci. 2018;42(1):1–3. doi:10.1016/j.tins.2018.09.012
26. Cairney SA, Guttesen AÁV, El Marj N, Staresina B. Memory consolidation is linked to spindle-mediated information processing during sleep. Curr Biol. 2018;28(6):948–954.e4. doi:10.1016/j.cub.2018.01.087
27. Jegou A, Schabus M, Gosseries O, et al. Cortical reactivations during sleep spindles following declarative learning. Neuroimage. 2019;195:104–112. doi:10.1016/j.neuroimage.2019.03.051
28. Niethard N, Ngo H-V, Ehrlich I, Born J. Cortical circuit activity underlying sleep slow oscillations and spindles. Proc Natl Acad Sci U S A. 2018;115(39):E9220–E9229. doi:10.1073/pnas.1805517115
29. Czarnecki A, Birtoli B, Ulrich D. Cellular mechanisms of burst firing-mediated long-term depression in rat neocortical pyramidal cells. J Physiol. 2007;578(Pt 2):471–479. doi:10.1113/jphysiol.2006.123588
30. Lubenov EV, Siapas AG. Decoupling through synchrony in neuronal circuits with propagation delays. Neuron. 2008;58(1):118–131. doi:10.1016/j.neuron.2008.01.036
31. Drosopoulos S, Schulze C, Fischer S, Born J. Sleep’s function in the spontaneous recovery and consolidation of memories. J Exp Psychol Gen. 2007;136(2):169–183. doi:10.1037/0096-3445.136.2.169
32. Schoch SF, Cordi MJ, Rasch B. Modulating influences of memory strength and sensitivity of the retrieval test on the detectability of the sleep consolidation effect. Neurobiol Learn Mem. 2017;145:181–189. doi:10.1016/j.nlm.2017.10.009
33. Wilhelm I, Metzkow-Mészàros M, Knapp S, Born J. Sleep-dependent consolidation of procedural motor memories in children and adults: the pre-sleep level of performance matters. Dev Sci. 2012;15(4):506–515. doi:10.1111/j.1467-7687.2012.01146.x
34. Wilhelm I, Diekelmann S, Molzow I, Ayoub A, Mölle M, Born J. Sleep selectively enhances memory expected to be of future relevance. J Neurosci. 2011;31(5):1563–1569. doi:10.1523/JNEUROSCI.3575-10.2011
35. Plihal W, Born J. Effects of early and late nocturnal sleep on declarative and procedural memory. J Cogn Neurosci. 1997;9(4):534–547. doi:10.1162/jocn.1997.9.4.534
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.