Back to Journals » Journal of Inflammation Research » Volume 18
Hydroxysafflor Yellow A Reduced Neuroinflammation Induced by Astrocyte-Derived IL-17A by Inhibiting the IL-17RA/ACT1/NF-κB/IL-17A Loop After OGD/R
Authors Song L, Hua J, Liu K, Duan Y, Sun Y, Yu J, Yin J, Ma C, Xiao B, Ma D, Huang J ![]()
Received 18 July 2025
Accepted for publication 23 October 2025
Published 11 November 2025 Volume 2025:18 Pages 15687—15699
DOI https://doi.org/10.2147/JIR.S547446
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Lijuan Song,1,2,* Jianlin Hua,2,3,* Kexin Liu,2 Yanzhe Duan,2 Yukang Sun,2 Jiezhong Yu,1,4 Jinzhu Yin,2,3,5 Cungen Ma,1,2 Baoguo Xiao,2,6 Dong Ma,2,3,5 Jianjun Huang2– 4
1Shanxi Key Laboratory of Innovative Drug for the Treatment of Serious Diseases Basing on the Chronic Inflammation, Shanxi University of Chinese Medicine, Jinzhong, Shanxi, 030619, People’s Republic of China; 2Research Center of Neurobiology, Shanxi University of Chinese Medicine, Jinzhong, Shanxi, 030619, People’s Republic of China; 3Department of Neurosurgery, Sinopharm Tongmei General Hospital, Datong, Shanxi, 037003, People’s Republic of China; 4First Affiliated Hospital, Shanxi Datong University, Datong, Shanxi, 037009, People’s Republic of China; 5The Key Laboratory of Prevention and Treatment of Neurological Disease of Shanxi Provincial Health Commission, Sinopharm Tongmei General Hospital, Datong, Shanxi, 037003, People’s Republic of China; 6Institutes of Brain Science and State Key Laboratory of Medical Neurobiology, Fudan University, Shanghai, 200433, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dong Ma, Email [email protected] Jianjun Huang, Email [email protected]
Introduction: Neuroinflammation induced by astrocytes has garnered significant attention recently. The IL-17A/IL-17RA signal pathway plays an important role in ischemic stroke (IS). Hydroxysafflor yellow A (HSYA) has been reported to have anti-apoptotic and anti-inflammatory properties that can protect neurons. In this study, we explore a novel mechanism that underlies the anti-apoptotic and anti-inflammatory effects of HSYA.
Methods: In vitro experiments were carried out using primary astrocytes and HT22 neuronal cells in an oxygen–glucose deprivation/reoxygenation (OGD/R) model. Techniques such as Western blot, immunofluorescence staining, Enzyme-linked immunosorbent assay (ELISA), and quantitative real-time polymerase chain reaction (qRT-PCR) were utilized to detect relevant indicators. The purpose was to investigate the effect of HSYA on the influence of IL-17A secreted by primary astrocytes after OGD/R on HT22 neuronal cells.
Results: The results indicated that the production of IL-17A by astrocytes increased following OGD/R, which was reduced due to HSYA treatment. In addition, astrocyte-derived IL-17A resulted in neuronal cell damage. Further studies showed that HSYA reduced IL-17A production by inhibiting activation of the IL-17RA/ACT1/NF-κB/IL-17A loop, which ultimately alleviated neuroinflammation and reduced neuronal apoptosis.
Discussion: These findings suggest that an activated loop indeed exists between IL-17A and IL-17RA/ACT1/NF-κB after OGD/R, and HSYA treatment alleviated IL-17A release from astrocytes after OGD/R by inhibiting the IL-17RA/ACT1/NF-κB/IL-17A loop. These results further emphasize the anti-inflammatory and neuroprotective effects of HSYA and suggest that it may be a promising drug for treating IS.
Keywords: hydroxysafflor yellow A, neuroinflammation, IL-17A, astrocytes, oxygen-glucose deprivation/reoxygenation, OGD/R
Introduction
Ischemic stroke (IS) is a major global contributor to mortality and disability, with high rates of morbidity, mortality, disability, and recurrence. IS occupies approximately 70% of all strokes.1 However, the pathogenesis of IS is highly complex,2 and the inflammation is significant. In recent years, neuroinflammation of astrocytes and IL-17A derived from astrocytes after oxygen glucose deprivation/reperfusion (OGD/R) have attracted attention. It is widely accepted that IL-17A has a harmful effect on the development of acute IS3 and is extensively involved in the onset and development of the neuroinflammation after IS.4 Therefore, IL-17A inhibition might be a beneficial therapy approach for treating IS.
It is generally recognized that IL-17 is sourced from immune cells within the immune system, and this has attracted special attention. IL-17A is primarily generated by helper T cell 17 (Th17 cells)5 during the differentiation process. Cytokines including interleukin-6 (IL-6) and transforming growth factor-β (TGF-β) cause this process. Transcription factors like retinoic acid receptor-related gene receptor γt control its secretion. Except for Th17 cells, γδT cells,6 natural killer T cells, neutrophils, and monocytes/macrophages can also secrete certain amounts of IL-17A under specific circumstances.7 Regarding the complex IS pathological context, studies in recent years have revealed that astrocytes have also emerged as an important IL-17A source.4 It is generally believed that IL-17A is produced from non-traditional immune cells that might be involved in a series of intricate pathophysiological processes following IS. These processes include neuroinflammation aggravation, disruption of the blood–brain barrier (BBB) and neuronal damage. Therefore, IL-17A secreted by astrocytes has important theoretical and practical value in uncovering the inherent mysteries of IS and the development of innovative therapeutic strategies.
Hydroxysafflor Yellow A (HSYA) stands out as the most potent active compound in the traditional Chinese medicine (TCM) known as safflower,8 and has been found with good efficacy in activating blood circulation and removing blood stasis. The potential mechanisms of HSYA as a potent agent for treating IS may include reducing the infarct volume, and it has antioxidant, anti-inflammatory and anti-platelet aggregation properties.9 Additionally, HSYA has been demonstrated to exert neuroprotective effects through anti-apoptotic effects.10 However, the HSYA impact on IL-17A has not been comprehensively reported in previous studies related to IL-17A.
Therefore, in the present study, we use neurons to establish an OGD/R model and to verify whether HSYA can inhibit astrocytes to produce IL-17 after OGD/R, which leads to neuronal damage. In addition, the possible mechanisms of HSYA are explored.
Materials and Methods
Primary Astrocyte Culture
The experimental procedures and protocols received approval from the Ethics Committee of Shanxi University of Chinese Medicine (approval number: 2020DW081). All procedures were conducted in strict accordance with the guidelines of National Institutes of Health (NIH) for the care and use of laboratory animals. Measures were taken to minimize the distress and suffering of the animals throughout the study. The cerebral cortexes of one-day-old neonatal C57BL/6 fetuses were extracted on an ultra-clean bench under aseptic conditions. The blood vessels and meninges were stripped off in a pre-cooled basal medium, and the cells were disrupted using a pipette to create a single-cell suspension. Centrifugation was then performed, and the supernatant was discarded. The cells kept blowing up once the media was given to them. This process was repeated two times. The cells were resuspended in a complete medium (high glucose Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin and inoculated in cell culture flasks coated with polylysine after being filtered through a 70-µm cell sieve and spun up. After that, the cells were placed in an incubator and cultured. After 48 h, the medium was swapped out to get rid of the detached cells. Until the cells spread evenly throughout the culture bottle at the bottom, interval of 3 days to replace the medium at a time. After that, the flasks remained 16 h at 37°C and 160 rpm in a constant temperature shaker. Microglia and oligodendrocytes were disposed of, and then trypsin was digested for use in passaging. Immunocytochemical staining of the primary cultured astrocytes was then conducted.
HT22 Hippocampal Neuronal Cell Culture
The HT22 mouse hippocampal neuronal cell line was bought from the Pricella Biotechnology Co., Ltd. in Wuhan, China. It was maintained in an adequate medium at 37°C and in an environment with humidity buffered with 5% CO2.
OGD/R and the HSYA Treatment
In order to replicate ischemia/reperfusion (I/R) circumstances in vivo, astrocytes and HT22 cells were both classified into three groups, including normal group, OGD/R group and HSYA group. First, DMEM without glucose was administered to the cells after the entire medium was disposed of and the cells were cleaned with phosphate buffer solution (PBS). After that, it cost four h to keep the cells in an anaerobic incubator set at 37°C, 94% N2, 5% CO2 and 0.1% O2. Following the OGD period, glucose was supplemented to normal levels. After that, the cells were put in an incubator under normal physiological growth conditions and incubated for 24 h. The drug group were given HSYA (75 µmol/L) concurrently. HSYA (ST02370120) was purchased from Shanghai Standard-Tech Co., Ltd., China.
Lactate Dehydrogenase Enzyme Activity Assessment
Following the instructions provided in the lactate dehydrogenase (LDH) test kit, the cell supernatants from the culture were obtained in order to assess the LDH activity. Detection reagents and samples were added sequentially. The standard curve and the analysis kit’s instructions were used to calculate the LDH activity.
Cell Counting Kit-8 (CCK-8)
Following preparing the cell cultures, 96-well plates were seeded with cell suspensions. The pharmacological intervention was carried out once the cells had attained about 80% confluence. Then, add 10 µL CCK-8 solution. After incubated for 2 h, 450 nm optical density (OD) was measured. The following formula was used to determine the cell viability: [OD (Drug Group) − OD (Blank Control Group)]/[OD (Control Group) − OD (Blank Control Group)]×100%.
Enzyme-Linked Immunosorbent Assessment
Following the directions provided by the manufacturer, an enzyme-linked immunosorbent test kit (Enzymo-link Bio, China) was used to assess the amount of IL-17A present in the cell contents and supernatants. The results were shown as pg*mL−1.
Quantitative RT-PCR
The RNAprep Pure cell kit was employed to extract the total RNA after the cells were lysed in Buffer RL for quantitative RT-PCR. Thermo (NanoDropOne) was used to measure the total RNA concentration. The Prime ScriptTM RT Master Mix (Takara Bio, Japan) was then used to synthesize cDNA, following the manufacturer’s guidelines. TB GreenTM Prremix EX TaqTM II (Takara Bio, Japan) was utilized for quantitative real-time PCR using the CFX96TM real-time PCR device.
Primer sequence information in this study were as follows: IL-17A: (FWD-AACTCCGTCGCCATTCAGCAAG, REV-ACACAGGTGCAGCCAACTTTTAGG); Tubulin: (FWD-GTGGCGGAGGAGCGGTTC, REV-CCAGGTCCAACAGTGAGTTCATC).
Western Blot
Proteins were taken out of each group’s HT22 cells and primary astrocytes. Ten percent one-step polyacrylamide gel electrophoresis (PAGE) and the bicinchoninic acid assay (BCA) technique were used to measure related protein concentrations. Following electrophoresis, the divided bands were placed on a polyvinylidene fluoride (PVDF) membrane (Millipore), sealed with a 5% powdered milk solution for 2 h, and then treated with following related antibodies, including Rabbit anti-IL-17A (1:1000, Affinity Biosciences, China), anti-ACT1 (1:1000, Affinity Biosciences, China), anti-p-p65 (1:1000, Affinity Biosciences, China), anti-IL-17RA (1:1000, Thermo Fisher, USA), and anti-Tubulin (1:8000, Bioword, USA). During an overnight rest period for the cells, the PVDF membrane was cleaned using TBST and reacted for 2 h at room temperature (RT) with goat anti-rabbit antibodies conjugated with horseradish peroxidase (HRP) (1:2000, Abcam, USA). The cells were finally rinsed three times with TBST and developed using a bioanalytical imaging system.
Immunofluorescence
The cultured cells were fixed with a tissue fixative solution for half an hour and washed 3 times by PBS. Then, the related antibodies were added and incubated at 4°C for 12 h, including rabbit anti-IL-17A (1:50, Affinity Biosciences, China), chicken anti-GFAP (1:300, Thermo Fisher, USA), and IL-17RA (1:500, Thermo Fisher, USA). This was followed by the addition of the related secondary antibodies at room temperature for 1 h. Following three washes by PBS, the cells were stained by using DAPI solution for 5 min at low light state. They were then sealed with 50% glycerol and observed under a fluorescence microscope to obtain the images. The results were analyzed using Image-J software.
Analysis of Statistics
The software GraphPad Prism 8.0 was employed to analyze the experimental data. The outcomes were articulated as 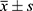 . Comparisons between two groups were made using a t-test, and comparisons among multiple groups were conducted using a one-way analysis of variance (ANOVA). P<0.05 was considered statistically significant.
. Comparisons between two groups were made using a t-test, and comparisons among multiple groups were conducted using a one-way analysis of variance (ANOVA). P<0.05 was considered statistically significant.
Results
Determination of the Optimal Modeling Time for Reoxygenation After the OGD of Astrocytes
The optimal OGD modeling time of astrocytes was 4 h, and the optimal HSYA intervention concentration was 75 µmol/L according to the results of our previous study.10 The aim of this experiment was to explore the optimal time for glucose and oxygen restoration. We examined the IL-17A content from the astrocyte supernatant and astrocyte cytoplasm at different time points using an ELISA. Figure 1A shows that the highest IL-17A level in the astrocyte supernatant was observed at 24 h of reoxygenation. The same results were obtained in the astrocyte cytoplasm (Figure 1B). Ultimately, the optimal time for astrocyte-derived IL-17A production was set at OGD4h/R24h.
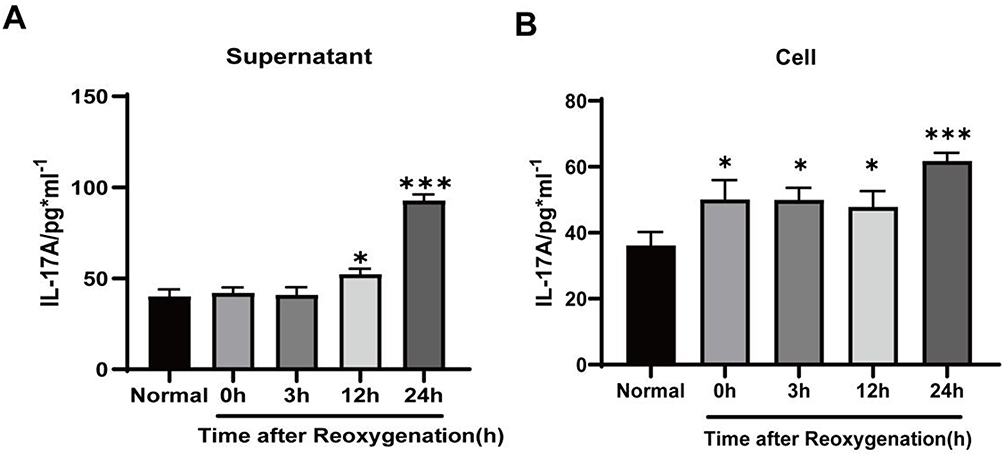 |
Figure 1 IL-17A concentration from primary astrocytes at different reoxygenation times. (A) IL-17A concentrations measured in the supernatant of astrocytes. (B) IL-17A levels measured in the astrocyte cytoplasm. The results were presented as means ± standard deviations from three separate studies and statistically examined using a one-way ANOVA. Notes: compared with the normal group, ***P < 0.001, *P < 0.05. |
HSYA Inhibited the Expression and Secretion of IL-17A Derived from Astrocytes After OGD/R
The HSYA effects on IL-17A production by astrocytes after OGD/R were detected using various assays such as q-PCR (Figure 2A), ELISA (Figure 2B), Western blot (Figure 2C) and immunofluorescence (Figure 2D). All of the results clearly demonstrated that, in comparison to the OGD/R group, IL-17A production was markedly reduced in HSYA group. These results indicated that HSYA indeed decreased IL-17A production from astrocytes after OGD/R.
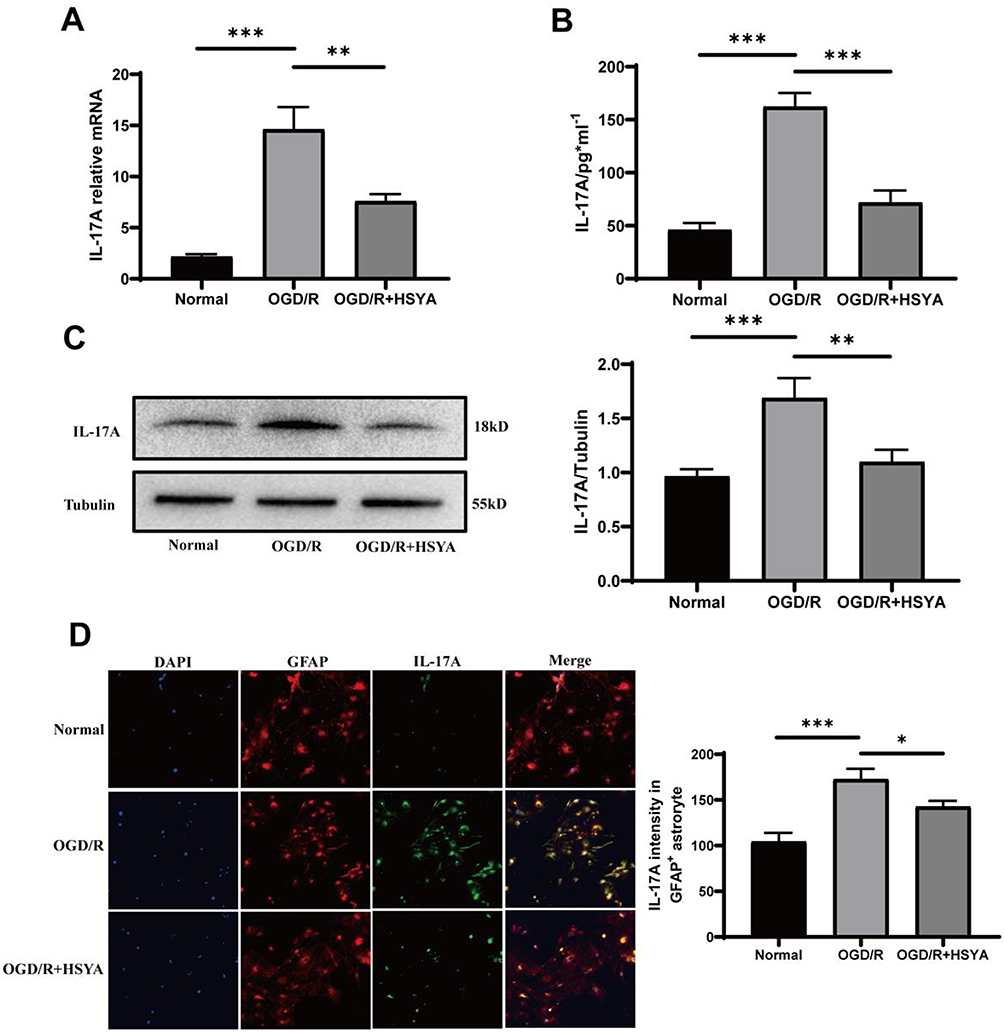 |
Figure 2 HSYA inhibited IL-17A production of astrocytes after OGD/R. (A) Statistical results of the IL-17A mRNA expression. This result suggested that HSYA had a certain inhibitory effect on IL-17A secretion. (B) IL-17A levels were detected by ELISA. The findings showed that HSYA decreased IL-17A expression. (C) IL-17A secretion was measured by Western blot. (D) IL-17A and GFAP co-localized in the OGD4h/R24h-treated astrocytes. The data were statistically evaluated using a one-way ANOVA and shown as means ± standard deviations. The results are indicative of three separate trials. Notes: ***P<0.001, **P<0.01, *P<0.05. |
HSYA Alleviated Astrocyte-Derived IL-17A-Mediated Neuronal Damage
An LDH assay was conducted to assess the neuronal damage caused by IL-17A derived from astrocytes. The results showed that the addition of exogenous IL-17A directly caused neuronal damage under normoxic conditions, which was partially decreased by the HSYA intervention (Figure 3A). In OGD/R neuronal cells, HSYA reduced the death of cultured neurons compared with OGD/R group (Figure 3B).
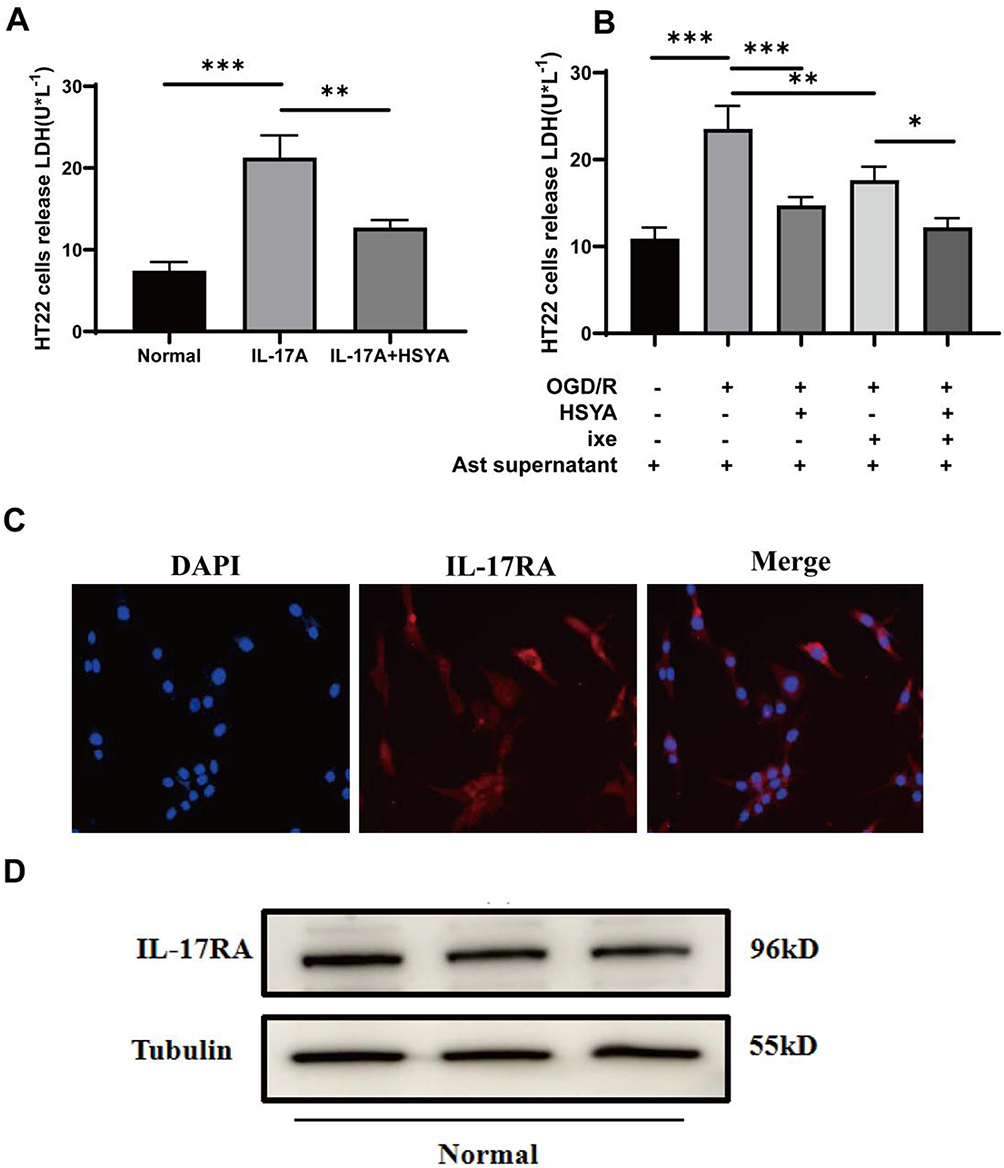 |
Figure 3 HSYA alleviated neuronal damage caused by IL-17A derived from astrocytes in different states. (A) HSYA and IL-17A affected LDH release in HT22 cells under normoxic conditions. (B) HSYA and IL-17A inhibitors (ixekizumab) affected LDH release in HT22 cells under the OGD/R status. (C) IL-17RA expression of HT22 cells, as measured by immunocytochemistry staining. (D) IL-17RA expression of HT22 cells, as measured by Western blot. Tubulin served as an internal control. The data were statistically examined using a one-way ANOVA and shown as means ± standard deviations. The results are indicative of three separate studies with comparable outcomes. Notes: ***P<0.001, **P<0.01, *P<0.05. |
Additionally, to further clarify the role of IL-17A in the astrocyte-conditioned medium, we added an IL-17A neutralizing agent (ixekizumab antibody) that significantly alleviated neuronal injury. These results indicated that HSYA effectively mitigated IL-17A-induced neuronal damage. We then used immunofluorescence staining and a Western blot to examine whether the cultured neuronal cells expressed IL-17RA. Figure 3C and D showed that both immunofluorescence and Western blot displayed that IL-17RA was expressed in the cultured neuronal cells, which lain the possibility for IL-17A to act upon neuronal cells. It can be concluded that the IL-17A produced by astrocytes was harmful to cultured neurons, while HSYA alleviated neuronal damage by inhibiting IL-17A production by astrocytes.
We further observed the effect of IL-17A on neuronal apoptosis. We used an astrocyte-conditioned culture medium to co-culture with HT22 cells. We found that astrocyte-conditioned culture media following OGD/R markedly increased the HT22 cells apoptosis. The addition of HSYA significantly attenuated neuronal apoptosis after OGD/R. The addition of IL-17A neutralizer (ixekizumab antibody) also attenuated neuronal damage (Figure 4). This result suggested that HSYA attenuated neuronal apoptosis by inhibiting astrocyte-derived IL-17A.
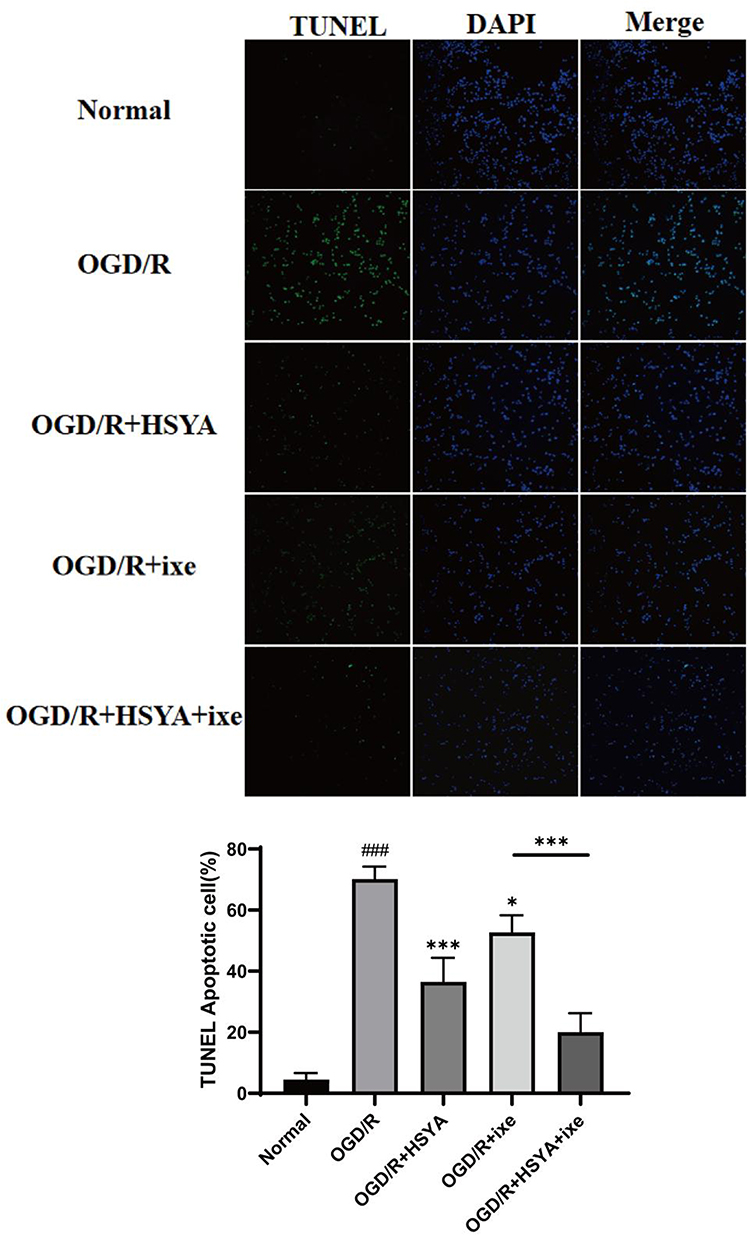 |
Figure 4 HSYA attenuated neuronal apoptosis after OGD/R. The TUNEL-positive cells appear green, and the nuclei are blue. The data were statistically evaluated using a one-way ANOVA and demonstrated as means ± standard deviations. The results are indicative of three separate trials. Notes: compared with normal group, ###P<0.001; compared with OGD/R group, *P<0.05, ***P<0.001. |
HSYA Inhibited IL-17RA-Triggered IL-17A Autocrine Secretion in Astrocytes
It is well known that astrocyte membranes express IL-17A receptors. Therefore, we explored different concentrations of exogenous IL-17A on astrocyte damage and finally selected 100 ng/mL of IL-17A as the inclusion standard for subsequent experiments (Figure 5A). Molecular docking was used to probe the binding of HSYA to IL-17RA to investigate the HSYA effect on IL-17RA and its downstream ACT1/NF-κB signaling molecules activated by IL-17A. The results indicated that the binding energy of HSYA docked to IL-17RA was less than −8.3 kcal/mol (Figure 5B), suggesting a strong docking interaction between HSYA and IL-17RA, which could bind to each other.
Figure 5 Continued. Figure 5 HSYA suppressed the IL-17RA/ACT1/NF-κB/IL-17A loop in astrocytes. (A) Different concentrations of exogenous IL-17A stimulated the release of LDH and the cell viability of astrocytes. (B) Molecular docking images. (C) IL-17RA, Act1, p-NF-kB/p65, and IL-17A expressions in astrocytes were detected using Western blot. (D) IL-17A levels were measured in the astrocytes supernatant. (E) IL-17RA, Act1 and p-NF-kB/p65 expressions under OGD/R. Three separate experiments produced the results. A one-way ANOVA was applied to compare data among various groups and a t-test was applied to compare data between two groups. The results were shown as means ± standard deviations. Notes: ***P<0.001, **P<0.01, *P<0.05.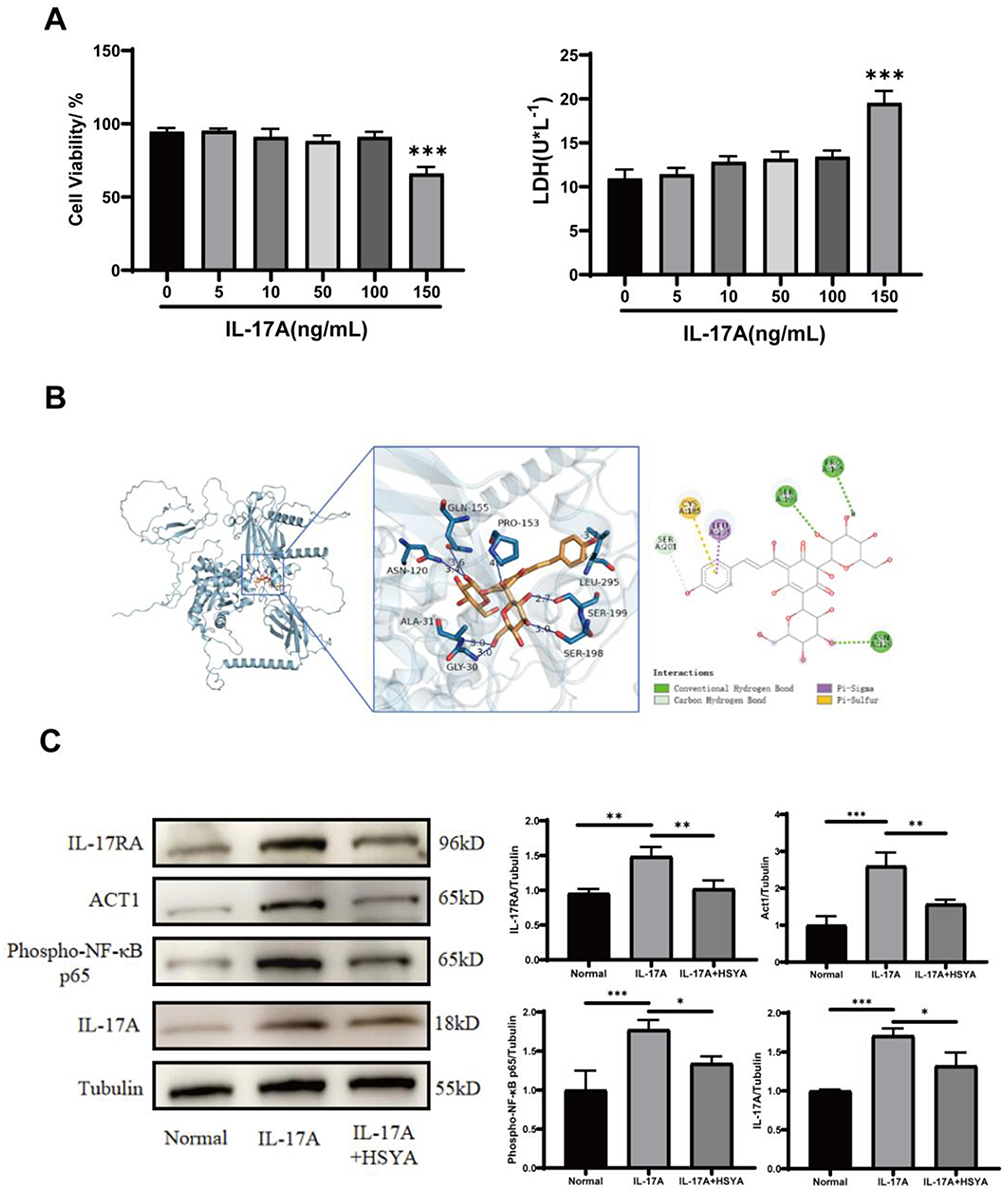

In terms of experimental determination, first, the IL-17RA expression and its downstream ACT1/NF-κB signaling molecules was detected in astrocytes. The outcomes showed the expressions of IL-17RA, ACT1, and p-NF-kB/p65 were reduced by the HSYA intervention compared to the IL-17A stimulation (Figure 5C), suggesting that HSYA can inhibit the ACT1/NF-κB signaling activation to a certain extent and then alleviate the production of IL-17A by the autocrine pathway in astrocytes, thereby protecting neurons. Second, we further validated the findings using an ELISA experiment (Figure 5D). Four groups were established: the normal group, the HSYA group, the IL-17A group and the IL-17A+HSYA group. In both HSYA group and IL-17A+HSYA group, the HSYA intervention was applied for 6 h, followed by PBS washing the medium. Third, Exogenous IL-17A was added to both the IL-17A group and the IL-17A+HSYA group over a duration of 6 h. Finally, the medium of all groups was washed, while a portion of the supernatant was collected as a baseline sample to ensure that the exogenous IL-17A was completely removed. After then, we cultivated the cells for one more day. Ultimately, the IL-17A concentration was measured in the cell culture supernatant in each group. The outcomes revealed that, in comparison to the IL-17A group, the HSYA intervention reduced the level of IL-17A autocrine secretion by astrocytes. Therefore, it can be concluded that HSYA may inhibit this vicious cycle of the positive feedback loop of IL-17A through this pathway.
We then speculated whether HSYA in the OGD/R state could also directly reduce endogenous IL-17A production by inhibiting IL-17RA and its downstream ACT1/NF-κB (Figure 5E). As expected, the findings suggested that HSYA is also capable of significantly decreasing the levels of IL-17RA, ACT1, p-NF-kB/p65 and IL-17A in comparison to OGD/R group, thus further demonstrating that HSYA could alleviate the damage of IL-17A to neurons possibly by suppressing the IL-17RA/ACT1/NF-κB/IL-17A loop in astrocytes.
Discussion
In modern society, ischemic stroke has emerged as an escalating threat to public health and individual lives. The neuronal injury it causes is widely recognized as a key factor leading to physical disabilities and even fatalities among stroke patients. Effective treatment still faces challenges and is very limited due to the complex pathophysiological mechanisms that occur after IS. Therefore, multi-target neuroprotection or brain protection has attracted widespread attention.
Neuroinflammation is a crucial process that follows IS and is initiated within minutes of the ischemia onset and continues for several days. In addition, IL-17 also holds a crucial role in diseases of the central nervous system (CNS), particularly autoimmune diseases such as stroke and multiple sclerosis. Studies have shown that IL-17 exacerbates neuroinflammation and neuronal damage by activating the downstream NF-κB signaling pathway.11 IL-17A has also been found to be closely associated with enhanced functional impairment, inflammation and neuronal cell death during IS.12 Inflammatory mediators, including IL-1, TNF-α and IL-6, contribute to BBB disruption, neuronal apoptosis and neuroplasticity impairment, eventually deteriorating the neurological deficit. It is noteworthy that IL-17A plays a key role in enhancing the early local inflammatory response, which ultimately leads to more severe neurological impairments.13 Both in the acute inflammation and chronic inflammatory process, IL-17A occupies a key position. Recent research has increasingly demonstrated that IL-17A influences various resident cell types within the CNS. It exacerbates neuronal inflammation after stroke, ultimately worsening ischemic brain injury. Following stroke, IL-17A induces the secretion of inflammatory mediators from associated cells, including TNF-α, IL-6, and CXCL1, and facilitates the migration of neutrophils into CNS, causing the BBB injury.14 These results showed that IL-17A may be a potential target for treating IS.
Mechanistically, IL-17A forms a complex with the IL-17RA/RC receptor, which recruits the ubiquitin ligase ACT1 in turn. The recruitment attracted TNF receptor-associated factor 6 (TRAF6), which subsequently triggered the NF-κB pathway. As an important cytoplasmic articulin, ACT1 is a cytoplasmic protein with homology to the cytoplasmic structural IL-17 receptor family domain. ACT1, along with all characterized IL-17 receptor family members, contains a highly conserved structural domain that exhibits distant homology to the toll-interleukin-1 receptor (TIR) domain similar with Toll-like receptors.15 The conserved domain is called the SEFIR domain. This suggests that ACT1 is involved in IL-17 cytokine signaling. ACT1 interacts with various IL-17 receptors by binding to their SEFIR domains.16,17 Thus, ACT1 may be a common signal transduction factor in the IL-17 family. Because ACT1 can bind both the IL-17 receptor and key components of the NF-κB pathway (such as IKK-α/IKK-β, NEMO/IKK-γ, TAK1, and TRAF6), it is proposed that ACT1 functions as a bridging protein. It attaches to the intracellular domains of the IL-17 receptor and interacts with NF-κB mediators, ultimately linking IL-17 receptor activation to NF-κB signaling.18–23 ACT1 can interact directly with the cytoplasmic structural domain of IL-17 RA24,25 and is regarded as an NF-κB-activating protein. Therefore, as a molecule necessary for connecting IL-17 RA stimulation to the downstream NF-κB signal pathway, ACT1 may occupy a critical position in the local inflammatory response.
HSYA is a major active component of TCM safflower, possessing anti-apoptotic and anti-inflammatory properties. Our previous researches have demonstrated that HSYA has a benifical effect on many intracerebral cells, including neurons.26,27 In middle cerebral artery occlusion (MCAO) mice, HSYA inhibited the TLR4-mediated signaling cascade, which contributed to a reduction of the inflammatory cytokines overproduction.28 Experimental evidence demonstrated that HSYA treatment effectively enhances cellular survival in BV2 microglia exposed to oxygen–glucose deprivation/reperfusion (OGD/R) injury.29 In addition, in MCAO rats HSYA attenuated the inflammatory response by upregulating glial fibrillary acidic protein (GFAP) and regulating the intercellular adhesion molecular 1 (ICAM-1) level.30 From previous studies, it is well known that IL-17 has a two-edged sword effect. IL-17A primarily promotes inflammatory responses and recruits immune cells in acute IS.31 IL-17A promotes neuronal injury and apoptosis. IL-17 may also exert a protective effect by activating endogenous neuroprotective mechanisms, such as upregulating antioxidants and brain-derived neurotrophic factor (BDNF).32 However, if this early immune response is not properly regulated, it may cause excessive inflammation, thereby exacerbating neuronal damage. In contrast, as acute injury subsides, the role of IL-17A may convert. Immune responses during the recovery phase require more precise regulation. If IL-17A remains upregulated, it may promote neuroinflammation, disrupt the BBB and cause more neuronal injury, hindering functional recovery.33 During the recovery phase, IL-17A may promote long-term neurodegenerative changes and chronic inflammation. In the acute injury phase, which is the focus in our present study, IL-17A is primarily derived from reactive astrocytes and is widely involved in neuronal injury progression. IL-17A plays a dual role by regulating immune cell recruitment and local inflammation. Damage can be effectively reduced by limiting excessive immune responses during the acute phase. IL-17A pathway inhibition can alleviate its negative effects and reduce neuroinflammation caused by ischemic brain injury. Therefore, the impact of IL-17A derived from astrocytes on neurons after stroke and whether HSYA can affect IL-17A in astrocytes still require further clarification.
Our results showed that astrocytes secrete IL-17A during OGD/R, while HSYA can decrease IL-17A secretion in effect. The conditioned medium of astrocytes under the OGD/R state can promote neuronal damage that can be partially alleviated by IL-17A neutralizing antibodies. This indicates that IL-17A produced by astrocytes under the OGD/R state is toxic to cultured neurons, while HSYA can protect neurons by inhibiting excessive IL-17A production by astrocytes. In addition, we further investigated whether HSYA could affect the binding of IL-17A with IL-17RA and inhibit its downstream ACT1/NF-κB signaling molecules, alleviating the IL-17A secretion by astrocytes and ultimately reducing neuroinflammation. The results suggested that HSYA may counteract the interaction between IL-17A and IL-17RA and reduce the activation of its downstream ACT1/NF-κB signaling pathway, subsequently further reducing IL-17A secretion by astrocytes. The findings of our study provide insights on the specific pharmacological mechanism of HSYA in treating IS.
Conclusions
HSYA protected neurons under the OGD state by reducing IL-17A derived from astrocytes, and its mechanism may be related to inhibitting the IL-17RA/ACT1/NF-κB/IL-17A loop. However, the precise target of HSYA still requires further confirmation.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China [82004028]; China Postdoctoral Science Foundation [2020M680912]; Science and Technology Cooperation and Exchange Special Project of Shanxi Province in 2023 (202304041101004); Innovation Team of Shanxi University of Chinese Medicine (2022TD2010); Research Project on Regulation of Neuroimmunity by Active Substances of Traditional Chinese Medicine [202204051001028]; Zhang Zhongjing inheritance and Innovation Project of State Administration of Traditional Chinese Medicine (GZY-KJS-2022-048-1); Medical Science and Technology Leading Team Project of Shanxi Provincial Health Commission [2020TD05]; Cardiovascular Special Fund Project of the National Regional TCM Medical Center of the Affiliated Hospital of Shanxi University of Chinese Medicine [XGZX202115]; Cultivation Plan on the Science and Technology Innovation Ability of Shanxi University of Chinese Medicine (2024PY-NS-012).
We thank LetPub (www.letpub.com.cn) for its linguistic assistance during the preparation of this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
There is no conflicts of interest.
References
1. Barthels D, Das H. Current advances in ischemic stroke research and therapies. Biochim Biophys Acta Mol Basis Dis. 2020;1866(4):165260. doi:10.1016/j.bbadis.2018.09.012
2. Gelderblom M, Weymar A, Bernreuther C, et al. Neutralization of the IL-17 axis diminishes neutrophil invasion and protects from ischemic stroke. Blood. 2012;120(18):3793–3802. doi:10.1182/blood-2012-02-412726
3. Liu T, Han S, Dai QQ, et al. IL-17A-mediated excessive autophagy aggravated neuronal ischemic injuries via Src-PP2B-mTOR pathway. Front Immunol. 2019;10:2952. doi:10.3389/fimmu.2019.02952
4. Li SJ, Dai QQ, Yu JL, et al. Identification of IL-17A-derived neural cell type and dynamic changes of IL-17A in serum/CSF of mice with ischemic stroke. Neurol Res. 2017;39(6):552–558. doi:10.1080/01616412.2017.1315863
5. Wang J, Gao Y, Yuan Y, et al. Th17 cells and IL-17A in ischemic stroke. Mol Neurobiol. 2024;61(4):2411–2429. doi:10.1007/s12035-023-03723-y
6. Mills KHG. IL-17 and IL-17-producing cells in protection versus pathology. Nat Rev Immunol. 2023;23(1):38–54. doi:10.1038/s41577-022-00746-9
7. Navarro-Compán V, Puig L, Vidal S, et al. The paradigm of IL-23-independent production of IL-17F and IL-17A and their role in chronic inflammatory diseases. Front Immunol. 2023;14:1191782. doi:10.3389/fimmu.2023.1191782
8. Yue SJ, Tang YP, Li SJ, et al. Chemical and biological properties of quinochalcone C-glycosides from the florets of carthamus tinctorius. Molecules. 2013;18(12):15220–15254. doi:10.3390/molecules181215220
9. Wei R, Song L, Miao Z, et al. Hydroxysafflor Yellow A exerts neuroprotective effects via HIF-1α/BNIP3 pathway to activate neuronal autophagy after OGD/R. Cells. 2022;11(23):3726. doi:10.3390/cells11233726
10. Han GY, Liu KX, Wei RH, et al. Protective effect of hydroxy saffron yellow pigment A on astrocytes after glyco-oxidative deprivation/recombined glyco-reoxygenation and its mechanism. Chin J Immunol. 2022;38(03):275–281. doi:10.3969/j.issn.1000-484X.2022.03.004
11. Zimmermann J, Nitsch L, Krauthausen M, et al. IL-17A facilitates entry of autoreactive T-cells and granulocytes into the CNS during EAE. NeuroMolecular Med. 2023;25(3):350–359. doi:10.1007/s12017-023-08739-0
12. Zhao XP, Zhao Y, Qin XY, et al. Non-invasive vagus nerve stimulation protects against cerebral ischemia/reperfusion injury and promotes microglial M2 polarization via interleukin-17A inhibition. J Mol Neurosci. 2019;67(2):217–226. doi:10.1007/s12031-018-1227-7
13. Shichita T, Sugiyama Y, Ooboshi H, et al. Pivotal role of cerebral interleukin-17–producing γδT cells in the delayed phase of ischemic brain injury. Nat Med. 2009;15(8):946–950. doi:10.1038/nm.1999
14. Levraut M, Bourg V, Capet N, et al. Cerebrospinal fluid IL-17A could predict acute disease severity in Non-NMDA-receptor autoimmune encephalitis. Front Immunol. 2021;12:673021. doi:10.3389/fimmu.2021.673021
15. Novatchkova M, Leibbrandt A, Werzowa J, et al. The STIR-domain superfamily in signal transduction, development and immunity. Trends Biochem Sci. 2003;28(5):226–229. doi:10.1016/S0968-0004(03)00067-7
16. Qian Y, Liu C, Hartupee J, et al. The adaptor Act1 is required for interleukin 17-dependent signaling associated with autoimmune and inflammatory disease. Nat Immunol. 2007;8(3):247–256. doi:10.1038/ni1439
17. Mauro C, Vito P, Mellone S, et al. Role of the adaptor protein CIKS in the activation of the IKK complex. Biochem Biophys Res Commun. 2003;309(1):84–90. doi:10.1016/s0006-291x(03)01532-8
18. Velichko S, Zhou X, Zhu L, et al. A novel nuclear function for the interleukin-17 signaling adaptor protein Act1. PLoS One. 2016;11(10):e0163323. doi:10.1371/journal.pone.0163323
19. Kanamori M, Kai C, Hayashizaki Y, et al. NF-kappaB activator Act1 associates with IL-1/Toll pathway adaptor molecule TRAF6. FEBS Lett. 2002;532(1–2):241–246. doi:10.1371/journal.pone.0163323
20. Schädlich IS, Vienhues JH, Jander A, et al. Interleukin-1 mediates ischemic brain injury via induction of IL-17A in γδ T cells and CXCL1 in astrocytes. NeuroMol Med. 2022;24(4):437–451. doi:10.1007/s12017-022-08709-y
21. Chen WC, Wen CH, Wang M, et al. IL‐23/IL‐17 immune axis mediates the imiquimod‐induced psoriatic inflammation by activating ACT1/TRAF6/TAK1/NF‐κB pathway in macrophages and keratinocytes. Kaohsiung J Med Sci. 2023;39(8):789–800. doi:10.1002/kjm2.12683
22. Wei X, Li C, Zhang Y, et al. Fish NF‐κB couples TCR and IL‐17 signals to regulate ancestral T‐cell immune response against bacterial infection. FASEB J. 2021;35(4):e21457. doi:10.1096/fj.202002393RR
23. Hao R, Song Y, Li R, et al. MLN4924 protects against interleukin-17A-induced pulmonary inflammation by disrupting ACT1-mediated signaling. Am J Physiol Lung Cell Mol Physiol. 2019;316(6):L1070–L1080. doi:10.1152/ajplung.00349.2018
24. Chang SH, Dong C. Signaling of interleukin-17 family cytokines in immunity and inflammation. Cell Signal. 2011;23(7):1069–1075. doi:10.1016/j.cellsig.2010.11.022
25. Zheng Y, Mou Z, Tan S, et al. IL-17A enhances the inflammatory response of glaucoma through Act1/TRAF6/NF-κB pathway. Neurochem Int. 2024;178:105787. doi:10.1016/j.neuint.2024.105787
26. Han G, Song L, Ding Z, et al. The important double-edged role of astrocytes in neurovascular unit after ischemic stroke. Front Aging Neurosci. 2022;14:833431. doi:10.3389/fnagi.2022.833431
27. Yin LJ, Wu YG, Dan CY, et al. Hydroxy saffron yellow pigment A inhibits NLRP3-mediated neurological impairment in microglia after ischaemic stroke via JAK2/STAT3 signalling pathway. Chin J Immunol. 2025;41(08):1820–1825+1832. doi:10.3969/j.issn.1000-484X.2025.08.005
28. Lv Y, Qian Y, Fu L, et al. Hydroxysafflor yellow A exerts neuroprotective effects in cerebral ischemia reperfusion-injured mice by suppressing the innate immune TLR4-inducing pathway. Eur J Pharmacol. 2015;769:324–332. doi:10.1016/j.ejphar.2015.11.036
29. Li J, Zhang S, Lu M, et al. Hydroxysafflor yellow A suppresses inflammatory responses of BV2 microglia after oxygen-glucose deprivation. Neurosci Lett. 2013;535:51–56. doi:10.1016/j.neulet.2012.12.056
30. Deng L, Wan H, Zhou H, et al. Protective effect of hydroxysafflor yellow A alone or in combination with acetylglutamine on cerebral ischemia reperfusion injury in rat: a PET study using 18F-fuorodeoxyglucose. Eur J Pharmacol. 2018;825:119–132. doi:10.1016/j.ejphar.2018.02.011
31. Li Y, Mao M, Zhu L, et al. IL-17A drives cognitive aging probably via inducing neuroinflammation and theta oscillation disruption in the hippocampus. Int Immunopharmacol. 2022;108:108898. doi:10.1016/j.intimp.2022.108898
32. Hu MH, Zheng QF, Jia XZ, et al. Neuroprotection effect of interleukin (IL)-17 secreted by reactive astrocytes is emerged from a high-level IL-17-containing Clin Exp Immunolenvironment during acute neuroinflammation. Clin Exp Immunol. 2014;175(2):268–284. doi:10.1111/cei.12219
33. Dai Q, Han S, Liu T, et al. IL-17A neutralization improves the neurological outcome of mice with ischemic stroke and inhibits caspase-12-dependent apoptosis. Front Aging Neurosci. 2020;12:274. doi:10.3389/fnagi.2020.00274
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
AVE 0991 Suppresses Astrocyte-Mediated Neuroinflammation of Alzheimer’s Disease by Enhancing Autophagy
Deng Y, Wang SY, Wang QG, Xu ZH, Peng Q, Chen SY, Zhu L, Zhang YD, Duan R
Journal of Inflammation Research 2023, 16:391-406
Published Date: 1 February 2023