Back to Journals » OncoTargets and Therapy » Volume 13
Human Positive Coactivator 4 Affects the Progression and Prognosis of Pancreatic Ductal Adenocarcinoma via the mTOR/P70s6k Signaling Pathway
Authors Su X, Yang Y ![]() , Ma L, Luo P, Shen K, Dai H, Jiang Y, Shuai L, Liu Z, You J, Min K, Shi C, Chen Z
, Ma L, Luo P, Shen K, Dai H, Jiang Y, Shuai L, Liu Z, You J, Min K, Shi C, Chen Z
Received 30 September 2020
Accepted for publication 17 November 2020
Published 26 November 2020 Volume 2020:13 Pages 12213—12223
DOI https://doi.org/10.2147/OTT.S284219
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Gaetano Romano
Xingxing Su,1,* Yishi Yang,1,* Le Ma,2,* Peng Luo,2 Kaicheng Shen,1 Haisu Dai,1 Yan Jiang,1 Ling Shuai,1 Zhipeng Liu,1 Jinshan You,1 Ke Min,1 Chunmeng Shi,2 Zhiyu Chen1
1Department of Hepatobiliary Surgery, Southwest Hospital, Third Military Medical University (Army Medical University), Chongqing 400038, People’s Republic of China; 2Institute of Rocket Force Medicine, State Key Laboratory of Trauma, Burns and Combined Injury, Third Military Medical University, Chongqing 400038, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiyu Chen
Department of Hepatobiliary Surgery, Southwest Hospital, First Affiliated Hospital of the Army Medical University, Chongqing 400038, People’s Republic of China
Tel +86-13983881219
Email [email protected]
Chunmeng Shi
Institute of Rocket Force Medicine, State Key Laboratory of Trauma, Burns and Combined Injury, Third Military Medical University, Chongqing 400038, People’s Republic of China
Tel +86-13708396812
Email [email protected]
Introduction: Pancreatic cancer is one of the deadliest cancers in the world, and pancreatic ductal adenocarcinoma (PDAC) accounts for 90% of all cases. Human positive coactivator 4 (PC4) is a transcriptional coactivator that has been associated with the development and progression of several tumors. However, no studies investigated the potential role of PC4 in PDAC.
Methods: We investigated PC4 expression in 81 PDAC tissue samples using immunohistochemistry and studied the impact of PC4 expression and the molecular mechanisms of this altered expression on PDAC tumorigenesis and proliferation both in vitro and in vivo.
Results: PC4 overexpression was correlated with a poor outcome in PDAC patients. The RNAi-mediated knockdown of PC4 expression in CFPAC-1 and AsPC-1 cell lines reduced cell proliferation and tumor growth. The loss of PC4 in PDAC inhibits cell growth by inducing cell cycle arrest at the G1/S transition and suppressing the mTOR/p70s6k pathway.
Discussion/Conclusion: Our findings reveal for the first time that PC4 exerts oncogenic functions by activating mTOR/p70s6k signaling pathway-mediated cell proliferation, implying that PC4 is a promising therapeutic target for PDAC.
Keywords: PC4, PDAC, proliferation, mTOR/p70s6k
Introduction
Pancreatic cancer is the seventh leading cause of cancer-related death worldwide, and the 5-year survival rate is only 9%.1 Among all pancreatic cancers, pancreatic ductal adenocarcinoma (PDAC) is the most common and aggressive form and accounts for approximately 90% of all pancreatic tumors.2 To date, PDAC remains highly resistant to almost all treatments, and surgery is still the most effective treatment.3 However, 80–90% of patients are not candidates for surgery at the time of diagnosis because pancreatic cancer is rarely diagnosed in an early stage.4 Therefore, improving the diagnosis, treatment and prognosis of PDAC is urgent. The tumor biology of PDAC plays a significant role in its development and prognosis.5 Hence, a better understanding of the mechanisms underlying malignant biological activities may reveal new therapeutic targets.
Human positive coactivator 4 (PC4), which was first identified as a transcriptional coactivator located on chromosome 5p13, encodes a 127-amino acid protein6,7 that is involved in the regulation of various cellular processes, including DNA transcription,8–11 DNA replication,12–16 DNA repair12–16 and chromatin organization.17–22 The overexpression of PC4 is associated with cancer progression, metastasis and radiosensitivity in several tumors.23–27 However, the relationship between PC4 expression and PDAC development and prognosis remains unknown.
In this study, we explored the potential role of PC4 in PDAC. We first found that PC4 was highly expressed in PDAC and positively correlated with a poor prognosis in patients. Furthermore, the effect of the downregulation of PC4 expression on the proliferation of PDAC cells was verified through both in vitro and in vivo studies. We also found that PC4 could bypass the G1/S checkpoint and activate mTOR/p70s6k signaling pathway-mediated cell proliferation in PDAC. Collectively, our study offers a novel insight into the functions and mechanisms of PC4 in PDAC and reveals that PC4 might hold promise for targeted therapies for pancreatic cancer.
Materials and Methods
Analysis of Datasets from the Cancer Genome Atlas (TCGA) and Gene Expression Profiling Interactive Analysis (GEPIA) Databases
The mRNA expression profiles of PC4 in pancreatic adenocarcinoma (PAAD) tissues were obtained from the GEPIA database. The association between the PC4 mRNA expression levels and overall survival of patients with PAAD was analyzed using TCGA database.
Patient Information and Follow-Up Study
In this study, we enrolled 81 patients diagnosed with PDAC between 2010 and 2018 at the Institute of Hepatobiliary Surgery, Southwest Hospital, Army Medical University. PDAC tissues were obtained from these patients, none of whom received chemotherapy before surgery. Overall survival was defined as the period between the date of surgery and the date of either death or last contact. This study was approved by the Ethics Committee of Southwest Hospital, Army Military Medical University. Written informed consent was obtained from all patients before they were enrolled in the study.
Tissue Microarray (TMA) and Immunohistochemistry (IHC)
The clinical tissue specimens were fixed in 4% paraformaldehyde for 24 h at room temperature before they were dehydrated and embedded in paraffin. In total, 81 sets of paraffin-embedded PDAC tissues, adjacent tissues and normal tissues were included in the TMA. The TMA (constructed by Shanghai Outdo Biotech Co., Ltd.), which was 1.5 mm in diameter and 3 μm thick, was dewaxed, rehydrated and incubated with human PC4 antibody (1:300; Sigma, St. Louis, Missouri, USA) at 4°C overnight. Then, the slides were sequentially incubated with a biotinylated secondary antibody at 37°C for 30 min, and positive staining was visualized using DAB. PC4 expression in the PDAC tissues was evaluated based on the percentage of positively stained cells and the staining intensity. The percentage of positive cells was quantitatively evaluated and scored as follows: 0 (< 5% positive cells), 1 (5–25% positive cells), 2 (26–50% positive cells), 3 (51–75% positive cells) and 4 (> 75% positive cells). The intensity was graded as follows: 0, no signal; 1, weak staining (light yellow); 2, moderate staining (brown); and 3, strong staining. The final score of each sample was calculated by multiplying the 2 values (range 0–12); then, the scores were categorized as negative (−, score 0–1), weak (+, score 2–4), moderate (++, score 5–8), or strong (+++, score 9–12). All tissue samples were examined and independently evaluated by two pathologists.
Western Blot Analysis
CFPAC-1 and AsPC-1 cells were harvested, washed, and lysed with RIPA buffer (Beyotime) containing protease inhibitor cocktail (Roche) for 30 min on ice. The total protein was extracted and quantified using a BCA kit (Beyotime) according to the manufacturer’s instructions. The protein samples were separated by electrophoresis using a 4%~20% gel and then transferred onto PVDF membranes (Millipore). The blotted membranes were blocked and incubated with primary antibodies overnight at 4°C. After 3 washes with TBST and a 1-h incubation with an HRP-conjugated secondary antibody (Proteintech) at room temperature, the membranes were developed using an enhanced chemiluminescence detection system (Bio-Rad Laboratories) to detect the protein. The primary antibodies against PC4 were obtained from Sigma, while the primary antibodies against β-actin, p-mTOR, t-mTOR, p-p70s6k, t-p70s6k, p-4EBP1, t-4EBP1, CDK4, CDK6, Cyclin D and Cyclin E were obtained from Cell Signaling Technology.
RNA Interference
Lentiviral vectors containing shRNAs targeting human PC4 (shRNA-1: 5′-GACAGGUGAGACUUCGAGATT-3′; 5′-UCUCGAAGUCUCACCUGUCTT-3′; shRNA-2: 5′-ACAGAGCAGCAGCAGCAGATT-3′; 5′-UCUGCUGCUGCUGCUCUGUTT-3′) and negative control shRNA (5′-UUCUCCGAACGUGUCACGUTT-3′; 5′-ACGUGACACGUUCGGAGAATT-3′) were constructed by GenePharma. CFPAC-1 and AsPC-1 cells were transfected with plasmids using Lipofectamine 3000 (Invitrogen) in OptiMEM (HyClone) according to the manufacturer’s instructions.
Quantitative Real-Time PCR
The total RNA was extracted from the CFPAC-1 and AsPC-1 cells using TRIzol reagent (Invitrogen, CA, USA) according to the manufacturer’s instructions. Real-time PCR was performed using a SYBR Green kit (Takara). The primers used for PCR are as follows: GAPDH, forward GGCATCCTGGGCTACACT, reverse CCACCACCCTGTTGCTGT; Sub1, forward TGATTCTGACAGTGAGGTTGAC, reverse TTATCATCTCTGCTGCTGCTG; CyclinD1, forward 3ʹ-CCATGTACCCTGGCATTGC-5ʹ, reverse 5ʹ-TCAGGAGGAGCAATGATCTTGA-3ʹ; and CyclinE1, forward 3ʹ-AAGCCCTCTGACCATTGTGTCC-5ʹ, reverse 5ʹ-CTAAGCAGCCAACATCCAGGAC-3ʹ.
Animals and Cell Lines
Athymic female nude mice (4–6 weeks) were obtained from the Centre for Experimental Animals and housed under specific pathogen-free conditions. The animal experiments followed the Guidelines for the Care and Use of Laboratory Animals of the TMMU, and all procedures were approved by the Animal Care and Use Committee of the TMMU. The human pancreatic cancer cell lines CFPAC-1 and AsPC-1 and a non-cancerous pancreatic ductal epithelial cell line (hTERT-HPNE) were purchased from the American Type Culture Collection (ATCC, Manassas, Virginia, USA) or the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). The AsPC-1 cells were grown in RPMI 1640 (HyClone, Logan, Utah, USA), the CFPAC-1 cells were grown in IMDM (HyClone, Logan, Utah, USA), and the hTERT-HPNE cells were grown in DMEM (Gibco, Gr-and Island New York, USA). All media were supplemented with 10% FBS (Gibco, Gr-and Island New York) and 1% streptomycin/penicillin (Beyotime), and the cells were incubated at 37°C in an atmosphere containing 5% CO2.
Cell Viability Assay
The cell viability was measured with a Cell Counting Kit-8 assay (Dojindo). Briefly, CFPAC-1 and AsPC-1 cells with stable PC4 knockdown (shPC4-1 and shPC4-2) and corresponding control cells were seeded in 96-well plates (4000 cells per well in 100 µL of medium) and cultured at 37°C in an incubator containing 5% CO2. The cell viability was tested 24 h, 48 h, 72 h, 96 h and 120 h after plating by measuring the optical density at a wavelength of 450 nm (OD450). The experiments were performed in triplicate.
Cell Cycle and Apoptosis Analyses by Flow Cytometry
A BD FACSCalibur flow cytometer was used for the cell cycle and apoptosis analyses according to the manufacturer’s protocol. For the analysis of the cell cycle distribution, CFPAC-1 and AsPC-1 cells with stable PC4 knockdown (shPC4-1 and shPC4-2) and corresponding control cells were collected, centrifuged and fixed in 75% ethanol at −4°C overnight. After incubating the cells with propidium iodide (PI, 50 μg/mL) and RNase for 20 min at 37°C in the dark, the cells were analyzed by flow cytometry within one hour. For the apoptosis assessment, the cells were stained with Annexin V-APC/PI (BD Biosciences) for 15 min at 37°C in the dark and then analyzed by flow cytometry. The experiments were performed in triplicate.
PDAC Cell Xenograft Mouse Model
For the in vivo tumor growth model, 100 µL of PBS containing 5 × 106 CFPAC-1 cells with stable PC4 knockdown (shPC4-1 and shPC4-2) or control cells were subcutaneously injected at one dorsal site into athymic female nude mice. These mice were divided into three groups (control, n=10; sh1-PC4, n=9 and sh2-PC4, n=9). The tumor growth was measured every 3 days, and the tumor volume was calculated using the following formula: volume (mm3) = (width2 × length)/2. At the endpoint, the mice were sacrificed, and the xenograft tumors were dissected, weighed and fixed in 4% paraformaldehyde for the subsequent immunohistochemical staining.
Statistical Analysis
All results are presented as the means ± standard deviations. The data were analyzed using Excel, GraphPad 7.0 and SPSS 25.0 (IBM). Comparisons between two groups were performed using Student’s t-test, whereas comparisons among three or more groups were performed using one-way analysis of variance (ANOVA). The survival data were analyzed using the Kaplan–Meier method. The correlations between PC4 expression and the clinical parameters were determined using Pearson’s χ2 method. A P-value<0.05 was considered statistically significant. The p-values are indicated as follows: “*”, P<0.05; “**”, P<0.01; “***”, P<0.001; and “****”, P<0.001.
Results
PC4 is Upregulated in PDAC and is Negatively Correlated with the Prognosis of Patients
Based on the analysis of the GEPIA database, the mRNA expression of PC4 in the PAAD tissues was higher than that in the adjacent tissues (Figure 1A), and this finding was confirmed in tissue arrays containing 81 sets of PDAC specimens obtained from patients at our institution (Figure 1B and C). The IHC results revealed aberrant PC4 overexpression in the carcinoma tissues, while a weak positive signal was found in the adjacent tissues, and almost no positive signal was observed in the normal tissues (Figure 1B); furthermore, the percentage of PC4-positive cells increased with the pathological grade (Figure 1C). The average calculated PC4 expression staining scores confirmed the above results (Figure 1D). The log-rank analysis suggested that high PC4 expression was significantly correlated with a poor prognosis among patients with PDAC (P<0.0001; shown in Figure 1E). However, in the GEPIA analysis, the correlation between PC4 expression and overall survival was not statistically significant (Figure 1F).
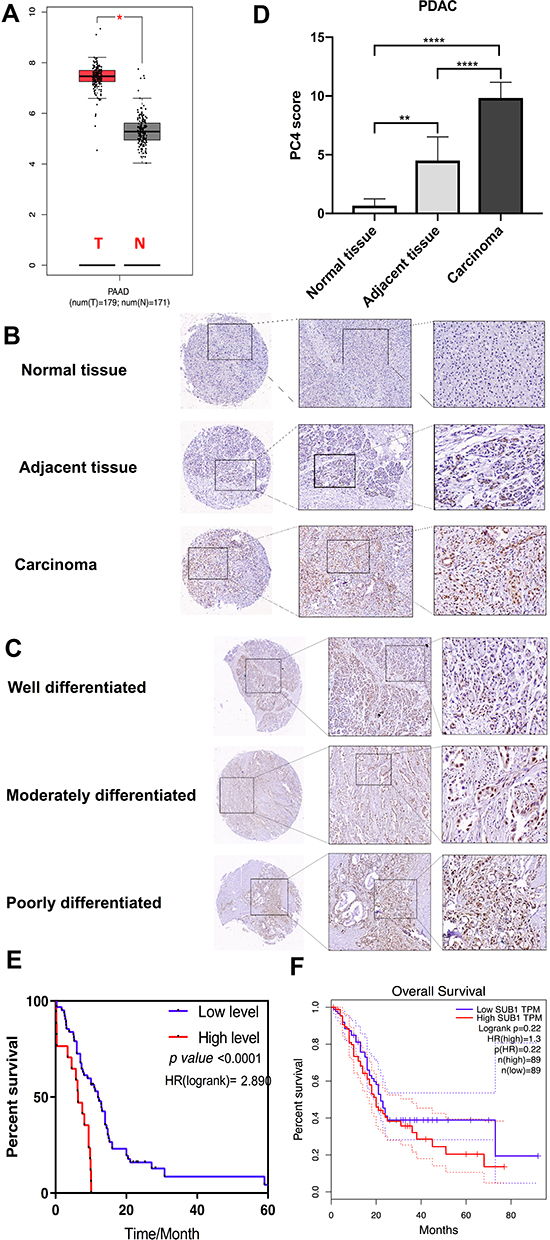 |
Figure 1 PC4 is upregulated in PDAC and is negatively correlated with the prognosis of patients. (A) PC4 mRNA expression levels in PAAD tissues (T) and normal tissues (N) from the GEPIA database. *P < 0.05. (B) Immunohistochemical staining of PC4 protein in 81 sets of PDAC tissues, adjacent tissues and normal tissues. (C) PC4 expression as assessed by immunohistochemistry in PDAC of different pathological grades. The percentage of PC4-positive cells increased with the pathological grade. (D) Comparison of the average PC4 expression staining scores on TMA slides. (E) Overall survival of patients with PDAC based on PC4 protein expression using a Kaplan–Meier analysis (n = 81, P < 0.001). (F) Analysis of the relationship between PC4 mRNA expression and overall survival in PAAD patients from TCGA database. * P<0.05, **P<0.01, **** P<0.0001. |
The relationships between the PC4 protein levels and clinical characteristics, including age, sex, TNM stage, and overall survival, were analyzed. The PC4 protein levels were not significantly associated with age, sex or the TNM stage (Table 1).
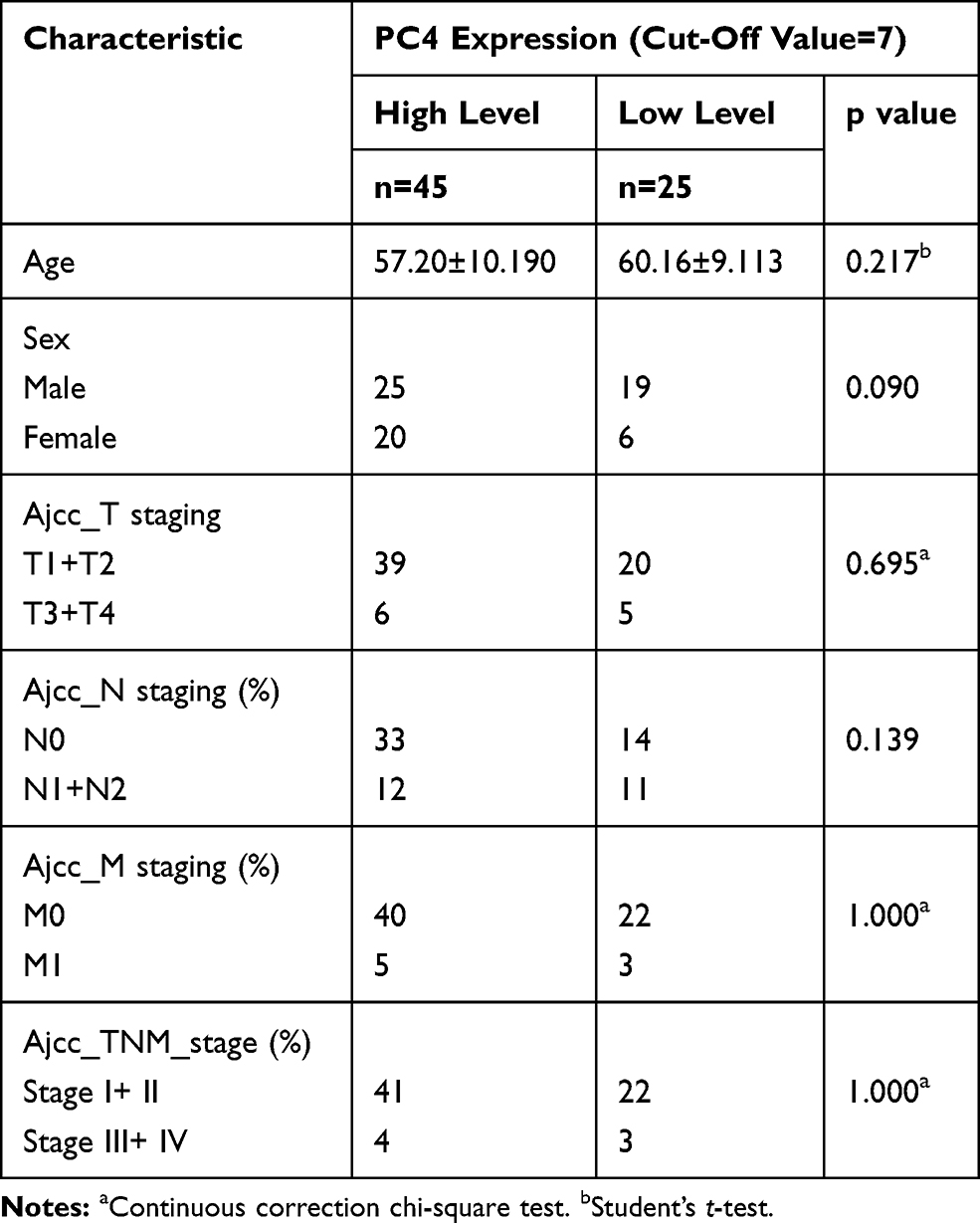 |
Table 1 The Relationship Between PC4 Protein Levels and the Clinical Characteristics of Patients with PDAC |
PC4 Knockdown Inhibited Cell Proliferation and Arrested the Cell Cycle at the G1-S Checkpoint in vitro
As shown in Figure 2A and B, the PC4 mRNA and protein expression levels in the PDAC cell lines (CFPAC-1, AsPC-1 and PANC-1) were higher than those in the normal human pancreatic duct epithelial cells (hTERT-HPNE). Among the four PDAC cell lines tested, the CFPAC-1 and AsPC-1 cells had the highest PC4 levels. Then, we constructed cell lines with stable knockdown of PC4 by transfecting CFPAC-1 and AsPC-1 cells with a PC4-specific shRNA and selecting with puromycin. We validated the knockdown efficiency by qPCR and Western blotting (shown in Figure 2C and D. The results of the CCK-8 assays indicated that the PC4 knockdown markedly reduced the viability of the CFPAC-1 and AsPC-1 cells (Figure 2E, P<0.0001). Furthermore, we determined the cell cycle distribution using flow cytometry. Figure 2F and G show that the PC4 knockdown caused fewer cells to enter the S phase and more cells to arrest in the G1 phase, indicating arrest at the G1/S checkpoint. However, knocking down PC4 had no significant impact on apoptosis in either the CFPAC-1 or AsPC-1 cells (Figure 2H and I). Overall, these results suggest that PC4 promotes PDAC cell proliferation and overcomes the checkpoints at the G1/S phase transition.
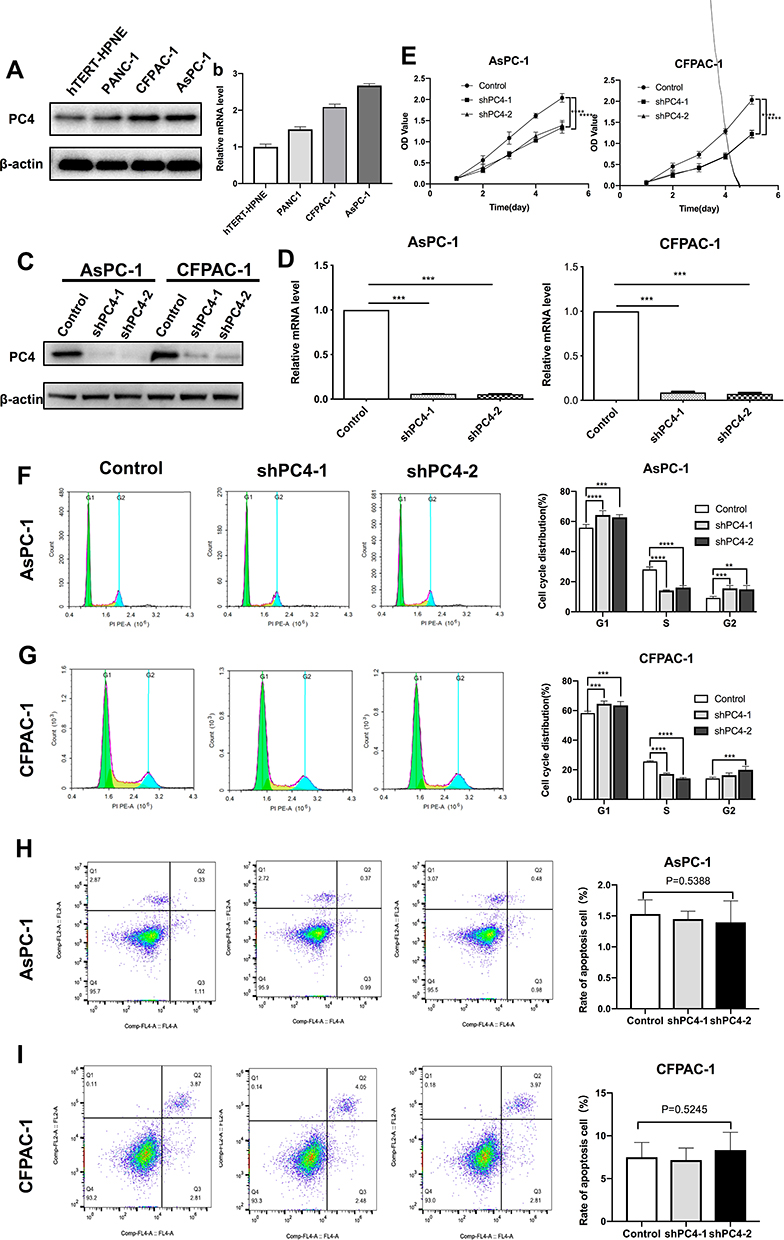 |
Figure 2 PC4 knockdown inhibited cell proliferation and arrested the cell cycle at the G1-S transition in vitro. PC4 protein (A) and mRNA (B) expression levels in PDAC cell lines (PANC-1, AsPC-1 and CFPAC-1) and normal human pancreatic duct epithelial cells (hTERT-HPNE). PC4 protein (C) and mRNA (D) expression levels in cells infected with either lentivirus expressing shRNA targeting PC4 (shPC4-1 and shPC4-2) or control empty lentivirus. (E) Cell viability of AsPC-1 and CFPAC-1 cells was measured using a CCK-8 assay 24 h, 48 h, 72 h, 96 h and 120 h after PC4 knockdown. (F, G) Cell cycle distribution in AsPC-1 and CFPAC-1 cells with stable PC4 knockdown and corresponding controls was determined by flow cytometry. Experiments were repeated three times independently. (H, I) Apoptosis rate in AsPC-1 and CFPAC-1 cells with stable PC4 knockdown and corresponding controls was determined by flow cytometry. Experiments were repeated three times independently. All data represent the means ± SDs. **P<0.01,*** P<0.001, **** P<0.0001. |
PC4 Knockdown Inhibited Tumor Growth in vivo
To further determine the biological function of PC4 in vivo, a subcutaneous xenograft tumor mouse model was established. CFPAC-1 cells with stable PC4 knockdown were injected into athymic female nude mice. Figure 3A shows that the PC4 expression levels in the tumors derived from shPC4-1 and shPC4-2 were lower than those in the tumors formed by the control cells. The xenograft growth in the mice injected with shPC4-1 or shPC4-2 cells was significantly attenuated compared with that in the mice injected with the control cells as shown in Figure 3B. At the experimental endpoint, the average size and weight of the tumors from the shPC4-1 and shPC4-2 groups were much smaller and lighter than those of the tumors from the control group. The tumor growth curve indicated that PC4 promotes PDAC cell growth in vivo (Figure 3C).
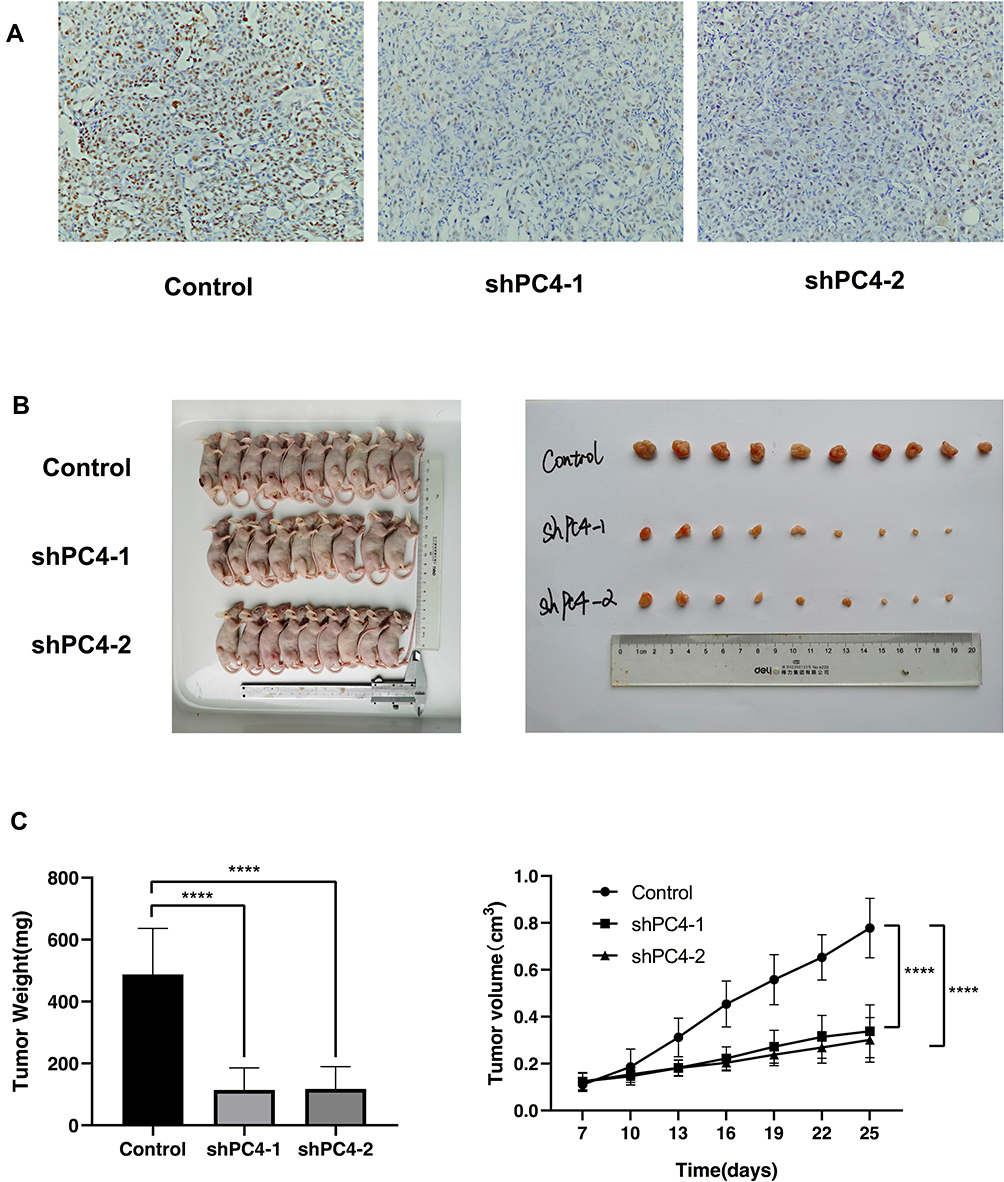 |
Figure 3 PC4 knockdown inhibited tumor growth in vivo. (A) PC4 expression levels in tumors formed by shPC4-1, shPC4-2 or control cells. (B, C) Effect of PC4 knockdown on the growth of subcutaneous xenograft tumors. (B) Images of subcutaneous xenograft tumors. (C) Tumor growth curve. Control, n = 10; shPC4-1, n=9; shPC4-2, n=9. **** P<0.0001. |
Knocking Down PC4 Suppressed the Activation of the mTOR Signaling Pathway, and MHY1485 Relieved the Inhibitory Effect on Proliferation in AsPC-1 and CFPAC-1 Cells
Since the mTOR/p70s6k signaling pathway plays a crucial role in regulating cell proliferation, the effect of PC4 on this pathway was investigated. The expression levels of key components in this pathway were examined by Western blotting. As shown in Figure 4A, the PC4 knockdown decreased the levels of phosphorylated mTOR, p70s6k and 4EBP1 compared with those in the control cells, indicating that PC4 could activate the mTOR signaling pathway. The mRNA levels of Cyclin D and Cyclin E in each group were also examined by RT-PCR (Figure 4B).
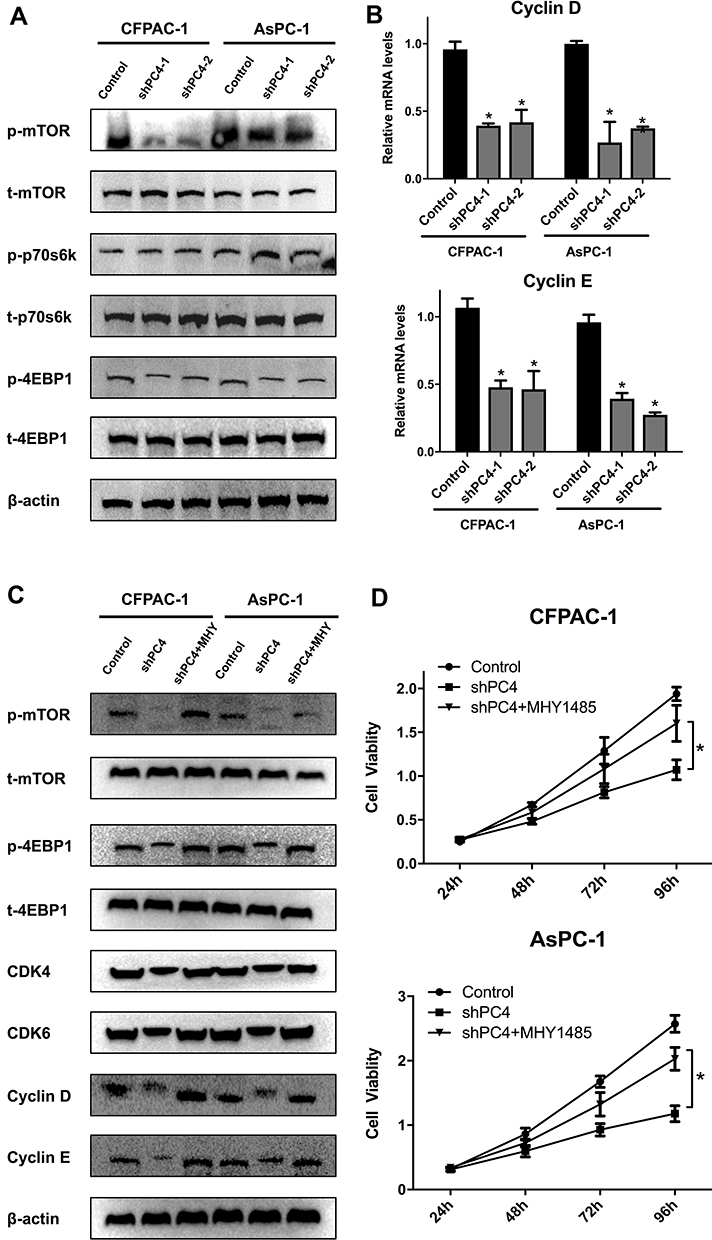 |
Figure 4 PC4 knockdown suppressed the activation of the mTOR/p70s6k signaling pathway in AsPC-1 and CFPAC-1 cells. (A). Western blot analysis of p-mTOR, t-mTOR, p-p70s6k, t-p70s6k, p-4EBP1 and t-4EBP1 protein levels in AsPC-1 and CFPAC-1 cells with stable PC4 knockdown and corresponding controls. (B). qPCR of Cyclin D and Cyclin E mRNA levels in each group. (C). Western blot analysis of p-mTOR, t-mTOR, p-p70s6k, t-p70s6k, CDK4, CDK6, Cyclin D and Cyclin E protein levels in AsPC-1 and CFPAC-1 cells with stable PC4 knockdown in the presence or absence of 2 nmol/L MHY1485 for 1 h and the corresponding controls. (D). Cell viability in each group was evaluated by a CCK-8 assay. The data are presented as the mean ± SD. *P<0.05, compared to the control. |
The mTOR activator MHY1485 was used to further verify whether the mTOR/p70s6k signaling pathway was responsible for the oncogenic functions of PC4 in AsPC-1 and CFPAC-1 cells. The reductions in the levels of phosphorylated mTOR, p70s6k, and 4EBP1 as well as CDK4, CDK6, Cyclin D, and Cyclin E in the PC4-knockdown cells were reversed by MHY1485 (Figure 4C). The CCK-8 assay showed that the viability of the AsPC-1 and CFPAC-1 cells in the shPC4 + MHY1485 group was significantly higher than that in the shPC4 group (Figure 4D). Altogether, these results suggest that PC4 promotes cell proliferation in PDAC by activating the mTOR/p70s6k pathway.
Discussion/Conclusion
Due to the increased incidence and mortality of PDAC and the difficulty of early detection, the current strategies used to treat and diagnose this disease face many obstacles.28 Thus, there is a great need to elucidate the underlying mechanisms of PDAC progression and identify novel therapeutic targets to improve the outcomes of PDAC patients. Here, we provide a novel perspective that PC4, which is an oncogene, is significantly upregulated in PDAC and plays a crucial role in PDAC tumorigenesis.
PC4, which is also known as SUB1, is a transcription coactivator that contains highly conserved sequences among humans, rats, mice and yeast.6,7 PC4 is a nuclear protein that participates in various cellular processes, including DNA transcription (8–11), DNA replication (12–16), DNA repair (12–16) and chromatin organization (17–22). The literature states that PC4 is significantly upregulated in several tumors, such as non-small-cell lung carcinoma (NSCLC), astrocytoma, and prostate cancer,18,24,26 implying that PC4 plays a crucial role in tumorigenesis. However, the function of PC4 in PDAC has not been investigated.
In this study, we specifically characterized PC4 expression in PDAC. Based on the tissue array analysis, PC4 was demonstrated to be significantly upregulated in PDAC tissues compared with adjacent tissues and was closely correlated with prognosis. However, we found that the increased PC4 expression was not associated with the TNM stage, suggesting that PC4 influences the tumorigenesis, but not the progression, of PDAC.
Subsequently, we evaluated the relationship between PC4 and the prognosis of PDAC patients. Although our experiments showed that the PC4 mRNA and protein expression levels were associated with overall survival, our results are inconsistent with TCGA analysis, which revealed that PC4 mRNA expression is not correlated with overall survival in PDAC patients. The following two reasons may explain this discrepancy: 1. the results of TCGA data are derived from analyses performed at the PC4 mRNA level by RNA sequencing or mRNA Chip, while our results are based on analyses at the PC4 protein level, such as the immunohistochemical analysis using tissue microarrays; and 2. most patients enrolled in TCGA project were Caucasian, while most patients enrolled in our study were Asian. However, we believe that evaluating PC4 protein expression and its correlation with prognosis is valuable and credible as PC4 plays an oncogenic role in many tumors and interacts with various cellular processes.
The in vitro experiments showed that the PC4 knockdown inhibited cell proliferation and arrested the cell cycle at the G1-S transition. This result is consistent with the in vivo results showing that PC4 knockdown can inhibit the growth of subcutaneous xenograft tumors. These data demonstrate that PC4 is a potential therapeutic target for the treatment of PDAC.
In addition, our study showed that the PC4 knockdown significantly inhibited the phosphorylation of mTOR, p70s6k and 4EBP1, indicating that the mTOR/p70s6k signaling pathway might be a potential target pathway of PC4. Numerous studies have confirmed that the mTOR/p70S6K signaling pathway plays a central role in cell survival, growth and proliferation, is significantly activated in various tumors, such as breast cancer, prostate cancer and cervical cancer, and has become a novel target in cancer therapy.29–34 mTOR is a member of the PIKK superfamily and an atypical serine/threonine protein kinase. mTOR regulates protein synthesis, cell growth, proliferation and apoptosis.35 p70s6k is an important key factor downstream of the mTOR pathway that directly activates mTOR and promotes protein synthesis. p70s6k is phosphorylated into p-p70s6k by mTOR, and the translational activity of p-p70s6k is approximately 100-fold greater than that of p70s6k.36 4EBP1 is a negative regulator of protein translation in mammalian cells. When cells are exposed to external stimuli, 4EBP1 is inactivated by mTOR, causing its dissociation from eIF-4E.37 In the presence of mTOR inhibitors, 4EBP1 is dephosphorylated and interacts with eIF-4E, inhibiting the initiation of translation. Our results demonstrate that the ectopic activation of the mTOR/p70s6k signaling pathway could rescue the effects of PC4 on the downregulation of the mTOR/p70s6k signaling pathway and reduction in cell viability. These results suggest that PC4 promotes cell proliferation in PDAC by activating the mTOR/p70s6k pathway.
Altogether, these results suggest that the PC4 protein levels in PDAC tissues are higher than those in adjacent tissues and that PDAC patients with high PC4 protein expression tend to have relatively shorter overall survival. In addition, the PC4 knockdown inhibited cell proliferation and tumor growth, indicating that PC4 functions as an oncogene in PDAC. Furthermore, PC4 exerts its oncogenic functions by activating the mTOR/p70s6k signaling pathway and inducing cell cycle arrest at the G1/S transition. To the best of our knowledge, this study is the first to provide novel insight into the functions and mechanisms of PC4 in PDAC and suggest that PC4 is a novel therapeutic target for PDAC.
Ethics Statement
This study was approved (KY2020138) by the research ethics committee of Southwest Hospital, Army Medical University, Chongqing, China. This study was conducted in accordance with the Declaration of Helsinki.
Funding
This study was supported by the National Natural Science Foundation of China (81874211).
Disclosure
The authors have no conflicts of interest related to this work to report.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. 2015;68:394–424.
2. Adamska A, Domenichini A, Falasca M. Pancreatic ductal adenocarcinoma: current and evolving therapies. Int J Mol Sci. 2017;18(7):1338. doi:10.3390/ijms18071338
3. Kamisawa T, Wood LD, Itoi T, Takaori K. Pancreatic cancer. Lancet. 2016;388(10039):
4. Rawla P, Sunkara T, Gaduputi V. Epidemiology of Pancreatic Cancer: global Trends, Etiology and Risk Factors. World J Oncol. 2019;10(1):10–27. doi:10.14740/wjon1166
5. Makohon-Moore A, Iacobuzio-Donahue CA. Pancreatic cancer biology and genetics from an evolutionary perspective. Nat Rev Cancer. 2016;24:
6. Kretzschmar M, Kaiser K, Lottspeich F, Meisterernst M. A novel mediator of class II gene transcription with homology to viral immediate-early transcriptional regulators. Cell. 1994;78(3):525–534. doi:10.1016/0092-8674(94)90429-4
7. Ge H, Roeder RG. Purification, cloning, and characterization of a human coactivator, PC4, that mediates transcriptional activation of class II genes. Cell. 1994;78(3):513–523. doi:10.1016/0092-8674(94)90428-6
8. Tavenet A, Suleau A, Dubreuil G, et al. Genome-wide location analysis reveals a role for Sub1 in RNA polymerase III transcription. Proceedings of the National Academy of Sciences of the United States of America. Proc Natl Acad Sci U S A. 2009;106(34):14265–14270. doi:10.1073/pnas.0900162106
9. Calvo O, Manley JL. The transcriptional coactivator PC4/Sub1 has multiple functions in RNA polymerase II transcription. EMBO J. 2005;24:1009–1020. doi:10.1038/sj.emboj.7600575
10. Wang Z, Roeder RG. DNA topoisomerase I and PC4 can interact with human TFIIIC to promote both accurate termination and transcription reinitiation by RNA polymerase III. Mol Cell. 1998;1(5):749–757. doi:10.1016/S1097-2765(00)80074-X
11. Malik S, Guermah M, Roeder RG. A dynamic model for PC4 coactivator function in RNA polymerase II transcription. Proceedings of the National Academy of Sciences of the United States of America. Nat Acad Sci. 1998;95(5):2192–2197. doi:10.1073/pnas.95.5.2192
12. Akimoto Y, Yamamoto S, Iida S, et al. Transcription cofactor PC4 plays essential roles in collaboration with the small subunit of general transcription factor TFIIE. Genes to Cells. 2014;19(12):879–890. doi:10.1111/gtc.12187
13. Jo J, Hwang S, Kim HJ, et al. An integrated systems biology approach identifies positive cofactor 4 as a factor that increases reprogramming efficiency. Nucleic Acids Res. 2016;44(3):1203–1215. doi:10.1093/nar/gkv1468
14. Kim J-M, Kim K, Schmidt T, et al. Cooperation between SMYD3 and PC4 drives a distinct transcriptional program in cancer cells. Nucleic Acids Res. 2015;43(18):8868–8883. doi:10.1093/nar/gkv874
15. Garavís M, González-Polo N, Allepuz-Fuster P, Louro JA, Fernández-Tornero C, Calvo O. Sub1 contacts the RNA polymerase II stalk to modulate mRNA synthesis. Nucleic Acids Res. 2017;45(5):2458–2471. doi:10.1093/nar/gkw1206
16. Garavís M, Calvo O. Sub1/PC4, a multifaceted factor: from transcription to genome stability. Curr Genet. 2017;63(6):
17. Mortusewicz O, Evers B, Helleday T. PC4 promotes genome stability and DNA repair through binding of ssDNA at DNA damage sites. Oncogene. 2016;35(6):761–770. doi:10.1038/onc.2015.135
18. Chakravarthi BVSK, Goswami MT, Pathi SS, et al. MicroRNA-101 regulated transcriptional modulator SUB1 plays a role in prostate cancer. Oncogene. 2016;35(49):6330–6340. doi:10.1038/onc.2016.164
19. Mortusewicz O, Roth W, Li N, Cardoso MC, Meisterernst M, Leonhardt H. Recruitment of RNA polymerase II cofactor PC4 to DNA damage sites. J Cell Biol. 2008;183(5):769–776. doi:10.1083/jcb.200808097
20. Yu L, Volkert MR. Differential Requirement for SUB1 in Chromosomal and Plasmid Double-Strand DNA Break Repair. PLoS One. 2013;8.
21. Wang J-Y, Sarker AH, Cooper PK, Volkert MR. The Single-Strand DNA Binding Activity of Human PC4 Prevents Mutagenesis and Killing by Oxidative DNA Damage. Molecular and Cellular Biology. Am Soc Microbiol. 2004;24:6084–6093.
22. Lopez CR, Singh S, Hambarde S, et al. Yeast Sub1 and human PC4 are G-quadruplex binding proteins that suppress genome instability at co-transcriptionally formed G4 DNA. Nucleic Acids Res. 2017;45(10):5850–5862. doi:10.1093/nar/gkx201
23. Luo P, Zhang C, Liao F, et al. Transcriptional positive cofactor 4 promotes breast cancer proliferation and metastasis through c-Myc mediated Warburg effect. Cell Commun Signaling. 2019;17.
24. Peng Y, Yang J, Zhang E, et al. Human positive coactivator 4 is a potential novel therapeutic target in non-small cell lung cancer. Cancer Gene Ther. 2012;19(10):690–696. doi:10.1038/cgt.2012.52
25. Qian D, Zhang B, Zeng XL, et al. Inhibition of human positive cofactor 4 radiosensitizes human esophageal squamous cell carcinoma cells by suppressing XLF-mediated nonhomologous end joining. Cell Death Dis. 2014;5.
26. Chen L, Du C, Wang L, et al. Human positive coactivator 4 (PC4) is involved in the progression and prognosis of astrocytoma. J Neurol Sci. 2014;346(1–2):293–298. doi:10.1016/j.jns.2014.09.014
27. Tao S, Yu J, Xu Y, et al. PC4 induces lymphangiogenesis dependent VEGF-C/VEGF-D/VEGFR-3 axis activation in lung adenocarcinoma. Am J Cancer Res. 2015;5:1878–1889.
28. Lin Q-J, Yang F, Jin C, Fu D-L. Current status and progress of pancreatic cancer in China. World j Gastroenterology. 2015;21(26):
29. Shin J-M, Jeong Y-J, Cho H-J, et al. Melittin suppresses HIF-1α/VEGF expression through inhibition of ERK and mTOR/p70S6K pathway in human cervical carcinoma cells. PLoS One. 2013;8(7):e69380. doi:10.1371/journal.pone.0069380
30. Zhang Y-J, Dai Q, Sun D-F, et al. mTOR signaling pathway is a target for the treatment of colorectal cancer. Ann Surg Oncol. 2009;16(9):2617–2628. doi:10.1245/s10434-009-0555-9
31. Easley CA, Ben-Yehudah A, Redinger CJ. mTOR-mediated activation of p70 S6K induces differentiation of pluripotent human embryonic stem cells. Cell Reprogram. 2010;12(3):263–273. doi:10.1089/cell.2010.0011
32. Smolewski P. Recent developments in targeting the mammalian target of rapamycin (mTOR) kinase pathway. Anticancer Drugs. 2006;17(5):487–494. doi:10.1097/00001813-200606000-00001
33. Li G, Shan C, Liu L, et al. Tanshinone IIA inhibits HIF-1α and VEGF expression in breast cancer cells via mTOR/p70S6K/RPS6/4E-BP1 signaling pathway. PLoS One. 2015;10(2):e0117440. doi:10.1371/journal.pone.0117440
34. Saraswati S, Kumar S, Alhaider AA. α-santalol inhibits the angiogenesis and growth of human prostate tumor growth by targeting vascular endothelial growth factor receptor 2-mediated AKT/mTOR/P70S6K signaling pathway. Mol Cancer. 2013;12(1):147. doi:10.1186/1476-4598-12-147
35. Asnaghi L, Bruno P, Priulla M, Nicolin A. mTOR: a protein kinase switching between life and death. Pharmacol Res. 2004;50(6):545–549. doi:10.1016/j.phrs.2004.03.007
36. Jefferies HB, Fumagalli S, Dennis PB, Reinhard C, Pearson RB, Thomas G. Rapamycin suppresses 5ʹTOP mRNA translation through inhibition of p70s6k. EMBO J. 1997;16:3693–3704. doi:10.1093/emboj/16.12.3693
37. Asimomytis A, Karanikou M, Rodolakis A, et al. mTOR downstream effectors, 4EBP1 and eIF4E, are overexpressed and associated with HPV status in precancerous lesions and carcinomas of the uterine cervix. Oncol Lett. 2016;12(5):3234–3240. doi:10.3892/ol.2016.5056
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.