Back to Journals » Psychology Research and Behavior Management » Volume 19
High Trait Anxiety Attenuates EML Amplitude Following Uncertainty Resolution: ERP Evidence of Impaired Emotional Disengagement
Authors Hu W, Zhu Y ![]() , Qiao L, Lu Y
, Qiao L, Lu Y
Received 11 December 2025
Accepted for publication 13 March 2026
Published 20 March 2026 Volume 2026:19 584294
DOI https://doi.org/10.2147/PRBM.S584294
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Mei-Chun Cheung
Wei Hu,1,2 Yingying Zhu,1,2 Libo Qiao,1,2 Yong Lu1,2
1Key Research Base of Humanities and Social Sciences of the Ministry of Education, Academy of Psychology and Behavior, Tianjin Normal University, Tianjin, 300387, People’s Republic of China; 2Faculty of Psychology, Tianjin Normal University, Tianjin, 300387, People’s Republic of China
Correspondence: Wei Hu; Yong Lu, Email [email protected]; [email protected]
Background: Trait anxiety reflects a stable predisposition to experience heightened anxiety across contexts. While its role in amplifying emotional responses during threat anticipation is well established, its influence after the resolution of uncertainty remains underexplored.
Purpose: This study examined whether individuals with high trait anxiety exhibit impaired neural disengagement following uncertainty resolution, compared with individuals with low trait anxiety. Emotional disengagement was operationalized using the Extrication from Mental Load (EML) component as a neurophysiological index.
Methods: Using a between-subjects extreme-group design, twenty-two university students were selected from the upper and lower tails of the State–Trait Anxiety Inventory–Trait (STAI-T) distribution and assigned to high and low trait anxiety groups (n = 11 per group). Participants completed a probabilistic two-level contingent negative variation (CNV) paradigm designed to manipulate uncertainty resolution. Neural disengagement was indexed by the peak-to-peak amplitude and latency of the EML component following uncertainty resolution.
Results: Compared with the low trait anxiety group, the high trait anxiety group exhibited significantly reduced EML peak-to-peak amplitudes, particularly over right frontal and midline parietal regions. No significant group difference was observed in EML peak latency.
Conclusion: These findings indicate that trait anxiety primarily modulates the magnitude, rather than the temporal onset, of neural disengagement processes following uncertainty resolution, suggesting reduced efficiency of post-threat neural disengagement in individuals with high trait anxiety.
Keywords: trait anxiety, emotional disengagement, uncertainty resolution, ERP, EML wave
Introduction
From a neurobiological and psychological perspective, anxiety is a future-oriented emotional state characterized by anticipatory responses to uncertain potential threats. The Uncertainty and Anticipation Model of Anxiety (UAMA)1 comprehensively outlines how such anticipatory processes, while adaptive, can become maladaptive in clinical anxiety. Central to the UAMA are five key processes, including inflated threat appraisals, attentional bias, cognitive avoidance, heightened reactivity to uncertainty, and notably, deficient safety learning. While the UAMA framework excels at detailing the mechanisms of threat anticipation, comparatively less theoretical and empirical attention has been directed toward the neural processes that support termination of anticipatory states once uncertainty is resolved. This imbalance creates a temporal asymmetry in anxiety research, wherein the initiation and maintenance of threat responding are well characterized, yet the mechanisms underlying post-threat disengagement remain insufficiently understood. Clarifying this termination phase is essential for explaining why anxiety may persist even after objective threat has subsided. This process of disengagement is conceptually analogous to fear extinction from learning theory,2–4 a process in which learned defensive responses are inhibited when a cue is found to be safe. Indeed, impaired fear extinction is considered a core mechanistic deficit in anxiety disorders. The term emotional disengagement is therefore proposed to describe the psychological and neural process of terminating an anticipatory state following uncertainty resolution. In this context, emotional disengagement refers specifically to the termination of anticipatory emotional processing once uncertainty is resolved as safe. This process is conceptually distinct from emotional withdrawal and related forms of disengagement, which involve reduced emotional engagement or behavioral responding rather than the cessation of anticipatory threat-related neural activity. Investigating the neural mechanisms underlying this phase is particularly promising, as it constitutes a “weak situation” where the absence of clear threat allows individual dispositions, such as trait anxiety, to more strongly influence neural function than during unambiguous threat anticipation.5 This leads to a critical question: do individuals with high trait anxiety demonstrate measurable neural deficits in emotional disengagement, thus perpetuating anxiety-related neural processing even after an objective stressor has been removed?
Behavioral Evidence for Impaired Emotional Disengagement
Empirical work exploring the temporal trajectory of anxiety across stressor phases provides crucial behavioral foundation for understanding emotional disengagement deficits. Conlon et al assessed math anxiety at pre-, mid-, and post-test intervals, finding significant state anxiety reduction immediately after task completion in most individuals.6 However, converging evidence indicates that this recovery may be compromised in high trait anxiety populations. Waechter & Stolz employed a multi-phase design measuring state anxiety across five experimental time points, revealing that high trait anxiety individuals maintained significantly elevated state anxiety at all post-induction phases, even after stressor removal. This persistence occurred regardless of mood induction condition, suggesting that trait anxiety sustains anxious responding beyond objective threat boundaries through difficulties in emotional disengagement or persistent activation of internal threat representations.7
Physiological findings corroborate this phenomenon of impaired disengagement. Cho et al examined autonomic recovery following an anxiety-inducing speech preparation task, demonstrating that while low trait anxiety individuals showed parasympathetic recovery after stressor removal, high trait anxiety participants exhibited no physiological recovery regardless of emotion regulation strategy employed, suggesting impaired deactivation of anxiety response systems.8 Similarly, Carr et al tracked anxiety trajectories in women undergoing gynecological surgery, finding that individuals with high trait anxiety exhibited sustained elevation throughout the perioperative period, even after surgical completion, with persistence associated with unresolved psychological factors independent of immediate threat.9
Moreover, this persistent anxious responding, even when objective threat is absent, parallels the well-documented impairments in fear extinction seen in anxiety disorders.10,11 Fear extinction involves the gradual reduction of conditioned fear responses following repeated presentations of threat cues in the absence of danger. A meta-analysis confirms that individuals with anxiety disorders exhibit delayed or reduced fear extinction compared to non-anxious controls, demonstrating persistent fear even in the face of manifest safety.12
These behavioral and physiological findings consistently demonstrate that trait anxiety extends its influence beyond threat anticipation, critically involving a compromised capacity for emotional disengagement. However, it remains unknown whether this behavioral persistence corresponds to a specific, measurable neural event at the moment of uncertainty resolution. This highlights the need to identify an objective neurophysiological marker, such as an ERP component, that can capture this failure to neurally disengage.
Neurophysiological Approaches to Emotional Processing in Anxiety
Event-related potential (ERP) methodology offers millisecond-level temporal resolution enabling objective investigation of neural processes underlying emotional disengagement.13 Existing ERP research has demonstrated that anxiety significantly alters neural processing across multiple cognitive and affective domains. Studies have examined components such as CNV (linked to cognitive load),14 N2 (associated with cognitive inhibition and conflict monitoring),15 and P3 (reflecting cognitive resource allocation).16,17 Components directly involved in emotion processing have also been explored, including P2 and LPP, both of which are sensitive to emotion regulation demands.18,19
Several studies have begun elucidating ERP correlates of uncertainty processing specifically. Tanovic et al demonstrated that both early P2 and slow stimulus-preceding negativity (SPN) components were modulated by threat uncertainty probability, with sustained negativity under moderate uncertainty.20 Johnen & Harrison showed that emotional uncertainty affects both anticipatory (SPN) and evaluative (LPP) processing phases.21 Most relevantly, Gole et al revealed that anxiety-prone individuals exhibited distinctive “dampening effects” in late LPP responses (750–1000 ms) when aversive stimuli followed uncertainty cues, suggesting that prolonged anticipatory worry alters subsequent emotional processing in trait-anxious individuals, potentially impairing their capacity for effective neural disengagement from anxiety-related processing.22
These findings establish the utility of ERP methodology for investigating anxiety-related neural processing, particularly during uncertainty resolution phases. However, specific neural markers of emotional disengagement following uncertainty resolution remain underexplored, highlighting the need for targeted investigation of post-stressor neural dynamics.
The Extrication from Mental Load Wave as a Neural Marker
A potentially crucial ERP component for investigating neural mechanisms of emotional disengagement is the Extrication from Mental Load (EML) wave, originally identified by Wei et al in modified CNV paradigms.23–25 Although originally identified within specific CNV paradigms, the EML can be conceptually situated within the broader international literature on Contingent Negative Variation (CNV) and anticipatory slow-wave dynamics. Since its original description by Walter et al,26 the CNV has been widely interpreted as reflecting sustained preparatory and expectancy-related neural activity under conditions of uncertainty.27–29 International studies have shown that enhanced slow-wave negativity indexes prolonged cognitive and affective engagement during anticipation,30 whereas post-stimulus shifts toward relative positivity have been associated with task completion or release from preparatory load.31 Within this framework, the EML may represent a specific instantiation of this general neural release mechanism, marking the termination of sustained anticipatory processing following uncertainty resolution. The EML manifests as a sharp polarity reversal from negative to positive approximately 150 ms following the command signal presentation, occurring specifically when anticipated imperative stimuli appear. This component’s absence when expected stimuli do not materialize suggests its functional role in releasing cognitive-affective tension accumulated during anticipatory periods. In simple terms, the EML can be understood as a neural “release signal” that marks the termination of anticipatory load once the expected stimulus appears.
The EML’s relevance to anxiety processing is supported by findings from Duan et al,32 who demonstrated that individuals experiencing chronic academic stress exhibited significantly more negative CNV amplitudes compared to controls, with state anxiety positively correlating with CNV amplitude in non-stressed individuals. This association was absent under high-stress conditions, presumably due to ceiling effects, suggesting that chronic emotional load may alter typical anxiety-neural processing relationships.
Theoretically, the EML represents a neural mechanism underlying the transition from sustained anticipatory load to cognitive-affective release following uncertainty resolution. Anxiety has consistently been associated with prolonged anticipatory engagement and heightened sustained neural activation under uncertain conditions. Unlike ERP components such as the SPN and LPP, which have been associated with anticipatory emotional engagement and sustained affective processing under uncertainty, the EML emerges only after the anticipated stimulus appears and may reflect the termination of sustained expectancy-related processing. Therefore, individual differences in EML amplitude may provide a direct neural index of variability in the efficiency of emotional disengagement, offering a temporally specific window into how trait anxiety influences post-uncertainty neural disengagement.
The Literature Gap and Present Study Rationale
Contemporary models such as the Uncertainty and Anticipation Model of Anxiety provide detailed accounts of threat anticipation mechanisms, yet considerably less attention has been devoted to the neural processes involved in terminating anticipatory states once uncertainty is resolved. Behavioral and physiological findings further suggest that anxiety may persist even after threat removal; however, the specific neurophysiological mechanisms underlying this failure to disengage remain unknown. This creates a critical gap in the current literature, as identifying objective neural markers of emotional disengagement is essential for understanding the persistence of anxiety beyond objective threat.
The present investigation addresses whether trait anxiety influences neural mechanisms of emotional disengagement following stressor removal, employing the EML as a neurophysiological marker of this process. According to recent theoretical perspectives, standard fear acquisition paradigms often constitute “strong” situations that, due to their lack of ambiguity, exert strong normative influence on behavior and may mask individual differences. In contrast, processes like fear extinction represent “weak” situations, where ambiguity increases the variance in responding that originates from individual characteristics.5 The disengagement phase of our paradigm, where threat is removed and safety must be inferred, constitutes such a “weak situation”, providing an ideal context for detecting trait-dependent variations in neural processing.
Individuals with high trait anxiety are expected to demonstrate impaired neural disengagement from anxiety-related processing after stressor removal, manifested as reduced EML amplitudes relative to individuals with low trait anxiety. This expectation is informed by findings from the fear conditioning literature indicating that individuals with anxiety disorders consistently display impaired fear extinction,12 such that defensive responses persist even when cues signal safety. Given that the EML is thought to reflect the cognitive-affective release from anticipatory load,23,24 reduced amplitude would serve as a precise neural correlate of this documented extinction deficit, indicating incomplete disengagement from the anxiety-related processing that characterized the preceding period of uncertainty.
By examining the EML as a neural marker of emotional disengagement, this study aims to provide neurophysiological evidence for the documented behavioral phenomenon of persistent anxious responding in high trait anxiety individuals, potentially elucidating the neural mechanisms underlying anxiety’s persistence beyond objective threat boundaries.
Accordingly, the present study addresses the following research question: Do individuals with high trait anxiety exhibit impaired neural disengagement following uncertainty resolution compared with individuals with low trait anxiety?
Based on prior findings from fear extinction and anxiety persistence literature, the following hypotheses were proposed:
Hypothesis 1: Individuals with high trait anxiety will show reduced EML peak-to-peak amplitude compared with individuals with low trait anxiety. Hypothesis 2: With regard to EML peak latency, no strong a priori directional prediction was made. If trait anxiety primarily influences the efficiency or magnitude of emotional disengagement rather than early processing speed, group differences would be expected to manifest predominantly in amplitude rather than latency.
Methods
Questionnaire
The State-Trait Anxiety Inventory (STAI) is a widely used self-report instrument designed to differentiate between two forms of anxiety: state anxiety, which reflects a temporary emotional condition in response to specific situations, and trait anxiety, which represents a stable tendency to experience anxiety across time and contexts.33 The inventory consists of two 20-item subscales: one for state anxiety (STAI-S) and one for trait anxiety (STAI-T). Because the present study aimed to examine neural disengagement as a stable individual characteristic, trait anxiety was selected as the grouping variable. State anxiety, which reflects transient situational fluctuations rather than enduring individual differences, was therefore not assessed. In the present study, the trait anxiety subscale (STAI-T), as revised into Mandarin Chinese by Ye & Hugdvet,34 was used to screen participants. Each item is rated on a 4-point Likert scale (1 = “not at all” to 4 = “very much so”), with total scores ranging from 20 to 80. A representative item from the STAI-T is: “I feel nervous.” In the present sample, the STAI-T demonstrated excellent internal consistency (Cronbach’s α = 0.913).
Participants
Participants were recruited from Tianjin Normal University through large-scale questionnaire screening conducted in classroom settings. A total of 512 college students completed the questionnaire, resulting in 486 valid responses (94% valid rate). In the screening sample (N = 486), the mean trait anxiety score was 44.9 (SD = 7.93). Based on this distribution, individuals whose scores fell beyond ±2 standard deviations from the sample mean were selected and invited to participate in the laboratory-based ERP experiment conducted in the EEG laboratory of the Faculty of Psychology at Tianjin Normal University. This procedure ensured that the high and low trait anxiety groups represented individuals at the extreme ends of the trait anxiety distribution. This extreme-group approach is widely used in individual differences research,35,36 as it enhances between-group variance and statistical sensitivity while preserving construct validity.37
Inclusion criteria required participants to be right-handed, have normal or corrected-to-normal vision and hearing, and report no history of neurological or psychiatric disorders. Participants currently taking psychoactive medication were excluded.
Based on these criteria, 12 individuals from each extreme formed the high and low trait anxiety groups. One participant from each group was excluded due to severe EEG artifacts, leaving 22 participants: 11 in the high trait anxiety group (3 males, 8 females; mean age = 20.4 ± 1.2 years, mean trait anxiety score = 69.0 ± 1.1) and 11 in the low trait anxiety group (5 males, 6 females; mean age = 20.6 ± 1.3 years, mean trait anxiety score = 28.1 ± 1.4). The trait anxiety scores differed significantly between the two groups (t(20) = 73.0, p < 0.01; See Table 1). The final sample size is comparable to prior ERP studies employing extreme-group designs.19,38 ERP signal reliability is enhanced through averaging across multiple trials, allowing stable estimation of ERP components even with moderate sample sizes.39 Recent methodological evaluations further indicate that ERP studies commonly employ modest sample sizes.40
 |
Table 1 Sample Characteristics of the High and Low Trait Anxiety Group (Mean ± SD) |
All participants provided informed consent and received compensation after the experiment. The procedures were approved by the Research Ethics Board of the Faculty of Psychology, Tianjin Normal University (Approval No. 2019050101).
Experimental Design and Rationale
The study employed a mixed factorial design, with trait anxiety group (high vs. low) as a between-subject factor and electrode position as within-subject factors, including anterior–posterior (frontal, central, parietal) and left–right (left, midline, right) dimensions. In this individual-differences design, the low trait anxiety group served as the comparison group for evaluating trait anxiety–related differences in neural disengagement.
Participants were categorized into high and low trait anxiety groups to examine whether neural disengagement following uncertainty resolution differs as a function of elevated trait anxiety. Although trait anxiety is generally associated with heightened emotional reactivity, it remains unclear whether post-resolution neural disengagement represents a distinct neurophysiological characteristic observable at higher trait levels. The present design therefore enabled a direct comparison of neural disengagement dynamics between individuals with high and low trait anxiety.
To examine whether post-resolution neural disengagement differs between groups, the experimental paradigm incorporated two key considerations: (1) Uncertainty Manipulation: Given that anxiety fundamentally reflects responses to uncertainty,1,41 the paradigm employed probabilistic stimulus presentation to create conditions of anticipatory uncertainty followed by resolution. This approach directly tests whether high trait anxiety individuals demonstrate impaired neural disengagement from anxiety-related processing after uncertainty resolution. (2) Evaluative Threat Context: Consistent with research showing that trait anxiety effects are most pronounced under evaluative conditions,42,43 the experimental context incorporated performance evaluation to enhance the relevance of individual differences in trait anxiety.
Within this framework, emotional disengagement was operationalized as a neurophysiological construct and indexed by the Extrication from Mental Load (EML) component, which captures neural disengagement following the resolution of task-related uncertainty.
Stimulus Materials
The auditory stimuli used in this study were adapted from the sound materials developed by Wei & Zheng,24 consisting of three signals: the first warning signal (S1), the second warning signal (S2), and the command signal (S3). S1 was an 800 Hz pure tone at 55 dB with a duration of 24 ms. S2 was a 1000 Hz pure tone at 55 dB, lasting 200 ms. S3 was a click sound (square wave) at 18 dB, lasting 100 μs. All stimuli were presented using Neuroscan’s Stim2 software and delivered through earphones via air conduction.
Functionally, S1 served as an initial warning cue signaling the onset of a trial and initiating anticipatory neural processing. This stimulus marks the beginning of the preparatory interval and triggers the early development of the contingent negative variation (CNV), reflecting the onset of expectancy-related neural activity.26 S2 functioned as a secondary warning signal that maintained and intensified this anticipatory state, contributing to the sustained buildup of the CNV, which reflects continued expectancy and preparatory processing. S3 served as the command stimulus that resolved anticipatory uncertainty and triggered the release of mental load, eliciting the Extrication from Mental Load (EML) component, which reflects neural disengagement following anticipatory cognitive-affective load.24
Tasks
Participants completed a two-level CNV task adapted from Wei & Zheng.24 This paradigm involved a sequential presentation of two warning signals (S1 and S2), followed by a command signal (S3) in a subset of trials. This sequence established an anticipatory interval between warning and command signals, with the probabilistic occurrence of S3 introducing uncertainty and its subsequent presentation enabling examination of post-resolution neural responses indexed by the EML component. In each trial, S1 was presented first, followed by S2 after a 1500 ms interval. After another 1700 ms interval, S3 was randomly presented in 50% of the trials (see Figure 1). The remaining 50% of trials did not include S3. The task consisted of 40 trials, with randomized inter-trial intervals ranging from 4 to 8 seconds. Participants were instructed to mentally count the number of times S3 appeared and report at the end of the task.
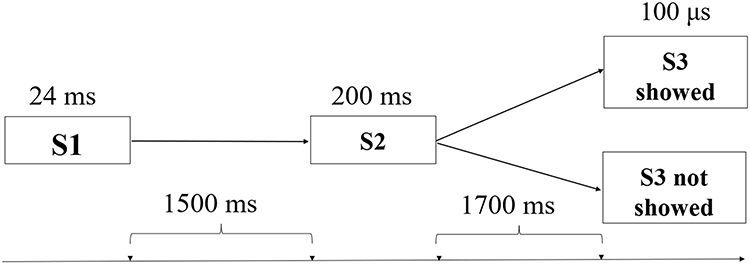 |
Figure 1 Experimental paradigm and temporal structure of a single trial. Each trial consisted of three sequential auditory stimuli. The first warning signal (S1, 24 ms) was followed by the second warning signal (S2, 200 ms) after a 1500 ms interval. After an additional 1700 ms interval, the command stimulus (S3, 100 μs) was presented in 50% of trials, whereas no stimulus was presented in the remaining trials. Participants were instructed to mentally count the number of S3 occurrences. Arrows on the timeline indicate the temporal intervals between successive events within a trial. |
Procedure
Participants were seated in a comfortable chair inside a shield room, approximately 1 meter from a computer screen. This viewing distance was maintained to ensure stable fixation, reduce ocular movement artifacts, and maintain a consistent visual angle, consistent with standard ERP recording practices.39 A black background with a green fixation cross (0.7 cm × 0.7 cm) was presented at screen’s center. Participants were instructed to focus on the green cross, pay attention to the auditory stimuli, and minimize blinking during the experiment. Participants were also informed that their performance would be compared to that of others, which was intended to increase evaluative pressure through social comparison. This evaluative instruction was included because socially evaluative contexts reliably amplify anxiety-related cognitive and neural responses, thereby enhancing sensitivity for detecting individual differences in trait anxiety.44,45
EEG Online Recording and Offline Analysis
EEG signals were continuously recorded online using the NeuroScan EEG/ERP system, which allowed real-time acquisition of neural activity during task performance. A 64-channel Quick-cap based on the international 10–20 system was used to ensure standardized electrode placement across participants. The left mastoid served as the reference during recording and was re-referenced offline to the average of both mastoids. Vertical electrooculogram (VEOG) electrodes were placed above and below the left eye, and horizontal electrooculogram (HEOG) electrodes were placed at the outer canthus of both eyes to monitor ocular activity. Impedance was maintained below 5 kΩ to ensure signal quality. Signals were recorded with a bandpass filter of DC–70 Hz and a sampling rate of 500 Hz.
Offline analysis was conducted to preprocess the raw EEG data and extract ERP components of interest. Raw EEG data were corrected for DC drift (DC offset correction), and ocular artifacts were corrected using regression-based procedures. The data were segmented into epochs ranging from 500 ms before S1 (used as baseline) to 4624 ms after S1. Epochs containing voltage fluctuations exceeding ±100 μV were excluded from further analysis to minimize contamination from artifacts.
ERP waveforms were then averaged separately for the high and low trait anxiety groups. Only trials in which S3 was presented were included in the analysis, as the EML component is elicited specifically following the presentation of the command stimulus. The grand average ERP waveforms showed a distinct positive deflection approximately 150 ms after S3, identified as the EML, consistent with previous findings.24
Given the wide distribution of the EML across the scalp, nine electrodes were selected for analysis: F5, Fz, F6, C5, Cz, C6, P5, Pz, and P6, categorized into two factors: (1) Anterior-Posterior Electrode Position: Frontal: F5, Fz, F6; Central: C5, Cz, C6; Parietal: P5, Pz, P6 (See the red arcs in Figure 2); (2) Left-Right Electrode Position: Left: F5, C5, P5; Midline: Fz, Cz, Pz; Right: F6, C6, P6 (See the blue arcs in Figure 2). Two metrics were analyzed for the EML: (1) Peak Latency (① in Figure 3): The time from S3 onset to the peak of the EML; (2) Peak-to-Peak Amplitude (② in Figure 3): The voltage difference between the start of the EML’s positive deflection and its peak.24 In the present study, these EML metrics served as objective neurophysiological indices of emotional disengagement efficiency following uncertainty resolution.
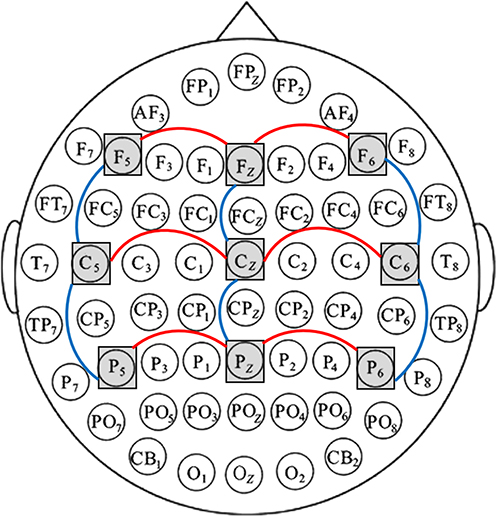 |
Figure 2 Electrode selection and regional grouping for statistical analysis. A 64-channel EEG system (international 10–20 system) was used. Nine electrodes (F5, Fz, F6, C5, Cz, C6, P5, Pz, P6) are indicated by grey squares. Electrodes were organized into two repeated-measures factors: Anterior–Posterior position—Frontal (F5, Fz, F6), Central (C5, Cz, C6), Parietal (P5, Pz, P6; red arcs); and Left–Right position—Left (F5, C5, P5), Midline (Fz, Cz, Pz), Right (F6, C6, P6; blue arcs). |
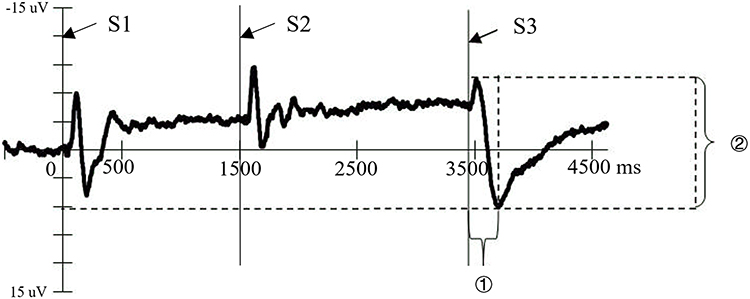 |
Figure 3 Operationalization of EML metrics under the secondary CNV paradigm. The figure presents a representative ERP waveform in the S3 condition, with stimulus onsets (S1, S2, S3) marked along the time axis. Two EML metrics were analyzed: (1) Peak Latency (①), defined as the interval from S3 onset to the EML peak; and (2) Peak-to-Peak Amplitude (②), defined as the voltage difference between the onset of the EML positive deflection following S3 and its peak. These metrics served as objective neurophysiological indices of neural disengagement following uncertainty resolution. |
Statistical analyses of the ERP data were conducted using SPSS 24. Separate repeated-measures ANCOVAs were performed on the EML peak latency and peak-to-peak amplitude. Trait anxiety group (high vs. low) was included as a between-subject factor, and electrode location was treated as a within-subject factor, with anterior–posterior and left–right dimensions. Gender was entered as a dummy-coded covariate to control for potential confounding effects. This approach was chosen to control for the potential influence of gender while simultaneously examining group differences and spatial distribution effects of the EML component across electrode locations. Greenhouse–Geisser corrections were applied when the assumption of sphericity was violated. Bonferroni adjustments were used for post hoc comparisons. The normality of the dependent variables was assessed using the Shapiro–Wilk test (ps > 0.05), and homogeneity of variance was confirmed using Levene’s test (ps > 0.05).
Results
Descriptive statistics for EML peak-to-peak amplitude and peak latency for each trait anxiety group across electrode positions are presented in Table 2. The grand-average ERP waveforms illustrating the EML component at representative electrode sites are shown in Figure 4.
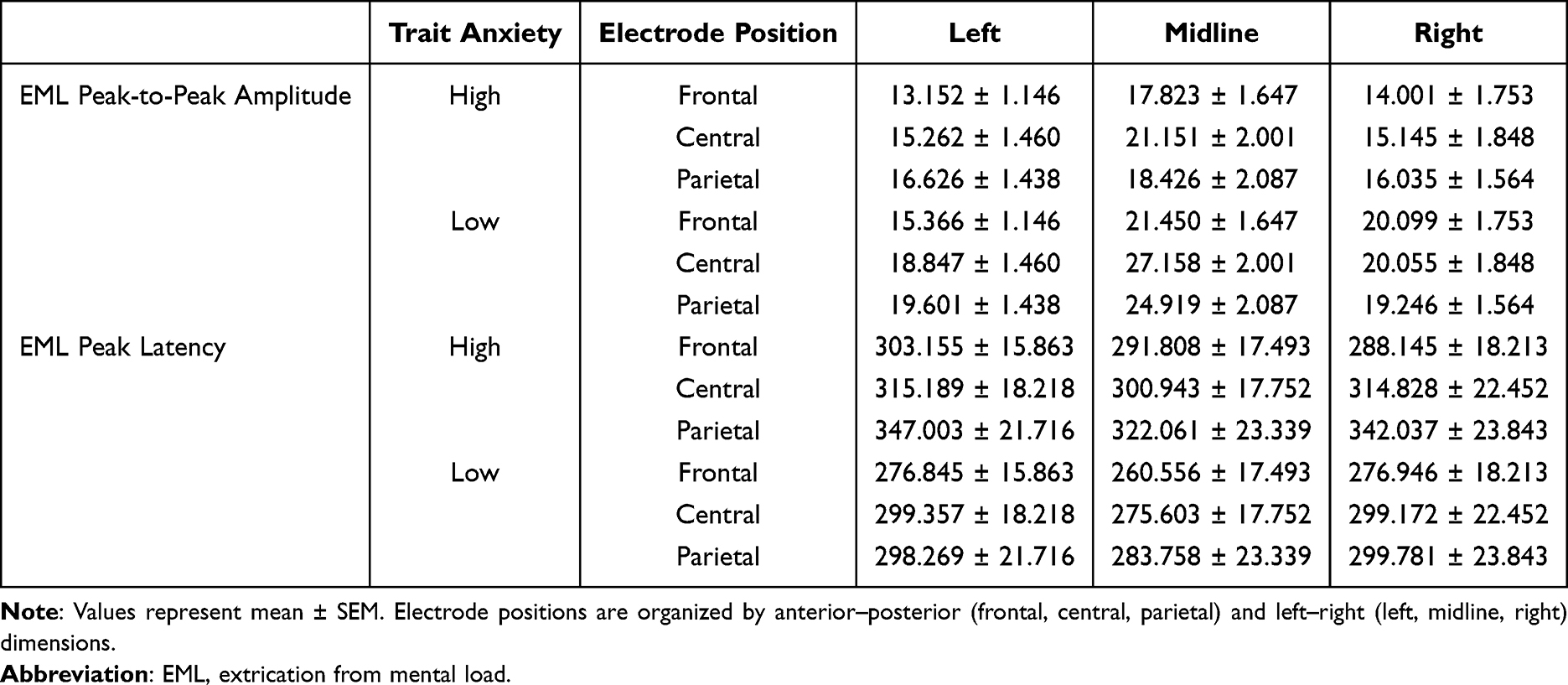 |
Table 2 Means (± SEM) of EML Peak-to-Peak Amplitude (µv) and Peak Latency (Ms) for High and Low Trait Anxiety Groups Across Electrode Positions |
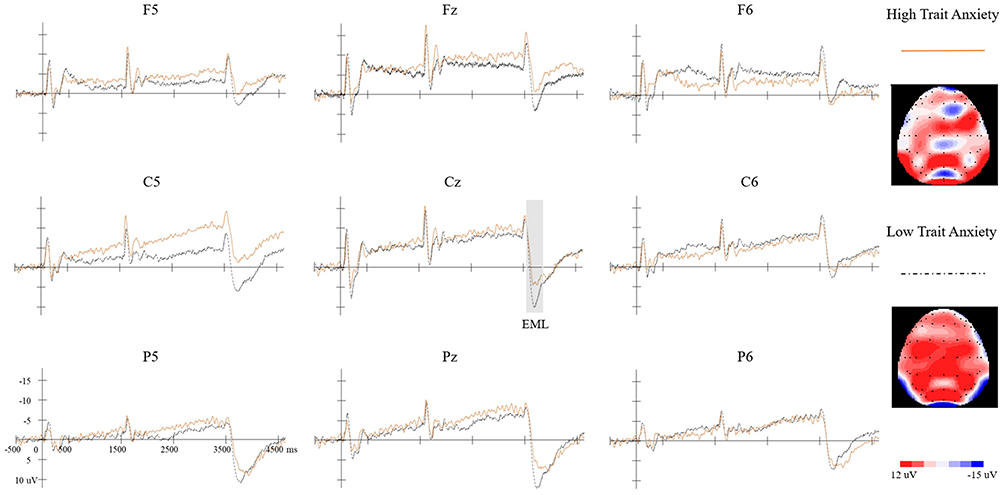 |
Figure 4 Grand-average ERP waveforms and topographic distribution of the EML. Grand-average ERP waveforms are presented for all nine electrodes in the S3 condition. The Orange solid line represents the high trait anxiety group, and the black dashed line represents the low trait anxiety group. The shaded region at Cz marks the EML component. Topographic maps on the right illustrate the scalp distribution at the EML peak latency at Cz for each group. |
EML Peak-to-Peak Amplitude
A repeated measures ANCOVA was conducted on EML peak-to-peak amplitude, with trait anxiety group (high vs. low) as the between-subject factor and anterior–posterior and left–right electrode positions as within-subject factors, and gender included as a covariate. See Table 3.
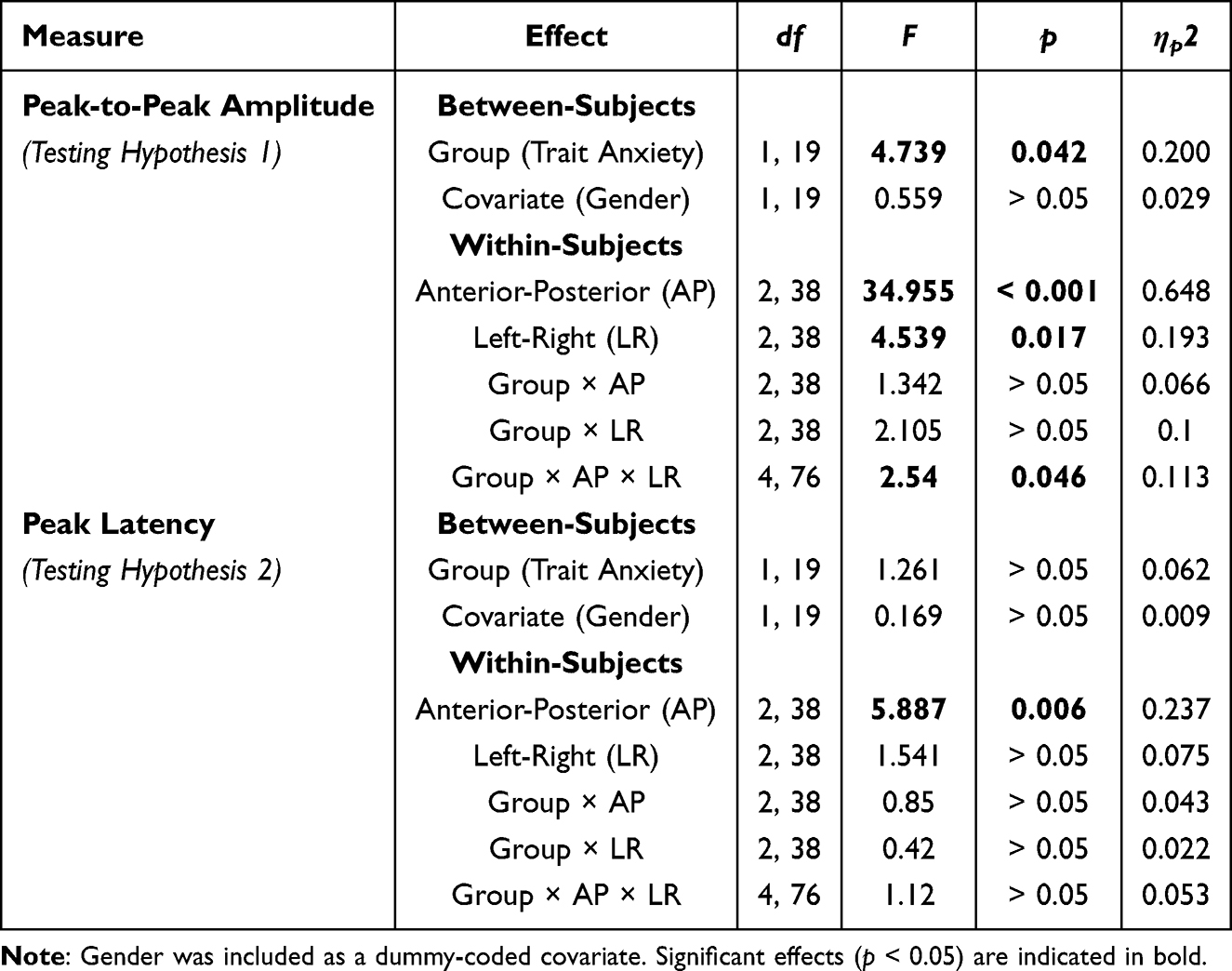 |
Table 3 Summary of Repeated Measures ANCOVA Results for EML Peak-to-Peak Amplitude and Peak Latency |
A significant main effect of trait anxiety group was observed, F(1, 19) = 4.739, p = 0.042, partial η2 = 0.200. The low trait anxiety group exhibited significantly greater EML amplitudes (20.749 ± 1.399 µV) than the high trait anxiety group (16.402 ± 1.399 µV), indicating stronger neural disengagement following uncertainty resolution.
A significant three-way interaction among trait anxiety group, anterior–posterior position, and left–right position was also observed, F(4, 76) = 2.540, p = 0.046, partial η2 = 0.113, suggesting that group differences varied across scalp regions. Simple effects analyses revealed significant group differences at F6 (p = 0.026) and Pz (p = 0.049) electrodes, with marginal significance at Cz (p = 0.052) and C6 (p = 0.078). Across these sites, the low trait anxiety group consistently showed larger amplitudes than the high trait anxiety group.
Significant main effects of anterior–posterior and left–right electrode positions were observed, reflecting topographic distribution of the EML component. Specifically, The main effect of anterior-posterior electrode position was significant, F(2, 38) = 34.955, p < 0.001, partial η2 = 0.648. Post hoc analysis revealed that the peak-to-peak amplitude at the frontal electrodes was significantly lower than at the central electrodes (16.982 ± 0.962 µV vs. 19.603 ± 1.163 µV, p =0.007), while other pairwise comparisons were non-significant (ps > 0.05). The main effect of left-right electrode position was also significant, F(2, 38) = 4.539, p = 0.017, partial η2 = 0.193. Post hoc tests indicated that amplitudes at left and right electrodes were significantly lower than at midline electrodes (left: 16.476 ± 0.824 µV; right: 17.430 ± 1.010 µV; midline: 21.821 ± 1.220 µV; ps < 0.001), with no significant difference between left and right electrodes (p > 0.05).
Gender, included as a covariate, did not show a significant main effect, F(1, 19) = 0.559, p > 0.05, partial η2 = 0.029, and no significant interaction effects involving gender were observed (ps > 0.05).
These findings are consistent with Hypothesis 1, demonstrating reduced EML peak-to-peak amplitude in individuals with high trait anxiety.
EML Peak Latency
A repeated measures ANCOVA was conducted on EML peak latency using the same factorial structure. See Table 3.
The main effect of trait anxiety group was not significant, F(1, 19) = 1.261, p > 0.05, partial η2 = 0.062, indicating no group difference in the timing of neural disengagement.
The main effect of anterior–posterior electrode position was significant, F(2, 38) = 5.887, p = 0.006, partial η2 = 0.237, reflecting spatial variation in latency across scalp regions. Post hoc analysis indicated that peak latency at frontal electrodes (282.909 ± 11.029 ms) was shorter than at central (300.848 ± 13.067 ms, p = 0.007) and parietal electrodes (315.485 ± 15.387 ms, p = 0.025), no significant difference between the central and parietal electrodes was found (p > 0.05). The main effect of left-right electrode position was not significant, F(2, 38) = 1.541, p > 0.05, partial η2 = 0.075.
Gender did not show a significant main effect, F(1, 19) = 0.169, p > 0.05, partial η2 = 0.009, and no significant interaction effects involving gender were observed (ps > 0.05).
These findings are consistent with Hypothesis 2, suggesting that trait anxiety does not affect the temporal onset of neural disengagement.
Topographic Map Observations
Topographic maps of the EML peak (Figure 4) illustrated the spatial distribution patterns for each group. The low trait anxiety group exhibited stronger and more widely distributed positive deflections, particularly over central and parietal regions, with pronounced activity along the midline electrodes. In contrast, the high trait anxiety group showed comparatively attenuated positivity, with reduced amplitude across frontal, central, and parietal areas.
The spatial pattern observed in the topographic maps was consistent with the ANCOVA findings, which indicated significant group differences in peak-to-peak amplitude and regional modulation across anterior–posterior and left–right electrode positions.
Discussion
This study investigated the neural underpinnings of emotional disengagement by examining how trait anxiety modulates the Extrication from Mental Load (EML) wave following the resolution of uncertainty. Emotional disengagement was operationalized using the EML peak-to-peak amplitude as a neurophysiological index reflecting the neural release from anticipatory load once safety was confirmed. Consistent with Hypothesis 1, individuals with high trait anxiety exhibited significantly reduced EML amplitudes compared to individuals with low trait anxiety, indicating diminished neural disengagement following uncertainty resolution. In line with Hypothesis 2, no significant group difference was observed in EML peak latency, suggesting that trait anxiety influences the magnitude rather than the temporal onset of the disengagement process. Together, these findings provide a direct neurophysiological answer to our research question: individuals with high trait anxiety show impaired neural disengagement after threat resolution compared with their low trait anxiety counterparts.
These findings contribute to the understanding of anxiety’s persistence in several key ways. First, by employing the EML as a temporally precise neural marker, this study shifts the empirical focus from anxiety anticipation to the often-overlooked process of emotional disengagement. Second, this study offers a plausible neural mechanism that helps explain the well-documented behavioral phenomenon of sustained anxious responding in high-anxiety individuals. Finally, the present results refine the Uncertainty and Anticipation Model of Anxiety (UAMA).1 Specifically, they demonstrate that impaired emotional disengagement is not a secondary effect, but a core feature of trait anxiety.
Impaired Neural Disengagement as a Correlate of Deficient Safety Learning
The study’s main finding—a markedly diminished EML peak-to-peak amplitude in the high trait anxiety group—provides a direct neural signature of impaired emotional disengagement. The EML is understood to reflect the release of cognitive-affective tension accumulated during periods of uncertainty.23,24 Therefore, a reduced EML suggests that high trait anxiety individuals fail to fully terminate anxiety-related neural processing once a situation is confirmed to be safe.
This finding offers a neurophysiological parallel to previous behavioral studies that have documented sustained subjective anxiety and delayed physiological recovery in high-anxiety individuals following stress exposure.6–9 In other words, while prior research has primarily demonstrated prolonged emotional responses at behavioral and physiological levels, the present study extends these findings by identifying a temporally specific neural marker associated with this impaired disengagement. More formally, this neural pattern can be interpreted as a marker of impaired fear extinction. A core deficit in anxiety disorders is impaired safety learning, which manifests as reduced or delayed fear extinction in the face of manifest safety.12,46 Our results align perfectly with this framework, suggesting that the reduced EML in high-trait-anxious individuals reflects a fundamental difficulty in engaging the neural circuits responsible for registering safety and disengaging from a defensive, anticipatory state.
The electrode-specific effects observed at F6 and Pz sites provide additional insight into the neural mechanisms underlying this deficit. The right frontal (F6) region has been consistently implicated in emotion regulation and cognitive control processes,47 while the parietal (Pz) region is associated with attentional disengagement and cognitive resource allocation.48 The selective reduction of EML amplitude at these sites in high trait anxiety individuals suggests that the impaired disengagement may specifically involve disruptions in right-lateralized prefrontal control mechanisms and parietal attention networks, which are crucial for successfully transitioning from threat-related to safety-related processing modes.
The topographic distribution patterns further support the functional significance of our findings. The low trait anxiety group exhibited widespread brain activation across the scalp, consistent with successful neural disengagement and cognitive-affective release. In contrast, the high trait anxiety group showed reduced overall activation with some regions remaining inhibited, suggesting incomplete neural disengagement. This pattern aligns with neuroimaging studies showing that anxiety is associated with persistent activation in threat-related neural networks even during safety periods,49 providing converging evidence for the construct validity of the EML as a marker of emotional disengagement.
Importantly, converging evidence from international neuroimaging studies provides further support for this interpretation. Research in Western populations has consistently demonstrated that individuals with elevated trait anxiety exhibit sustained activation in threat-related neural regions, including the amygdala, as well as reduced engagement of regulatory prefrontal areas during post-threat and safety processing.50–53 These neural patterns have been interpreted as reflecting a diminished capacity to terminate defensive responding once threat has resolved. Such findings provide cross-methodological support for the present ERP results and reinforce the view that trait anxiety is characterized by impaired neural disengagement rather than solely heightened anticipatory responding.
Strengths of the Experimental Paradigm
The two-level CNV paradigm was intentionally designed to operationalize the core UAMA constructs of anticipatory uncertainty and its subsequent resolution. The probabilistic (50%) presentation of the S3 command signal effectively induced a state of anticipation, which was resolved upon S3’s appearance. The critical observation that the EML was present only in S3-present trials—and absent in S3-omitted trials—powerfully reinforces its functional role as a specific neural index of disengagement from resolved uncertainty.
Furthermore, framing the task within an evaluative threat context (ie, social comparison of performance) was crucial for maximizing the expression of individual differences. It is well-established that the effects of trait anxiety are most pronounced under ego-threatening or evaluative conditions.42,43,54 This feature enhances the ecological validity of our findings, suggesting that the observed neural deficits are most relevant in socially stressful situations. By capturing the neural response in the immediate post-S3 window, this study was able to isolate the precise moment of emotional disengagement, providing a temporally sensitive neural index of this critical process.
Extending the Temporal Framework of Trait Anxiety
The present findings compel an extension of existing theoretical models of anxiety. While frameworks like the UAMA masterfully delineate the maladaptive processes occurring during threat anticipation, they do not explicitly detail the post-resolution phase.1 The present results fill this temporal gap, demonstrating that the influence of trait anxiety is not confined to the anticipatory period but involves a compromised ability to terminate the anxiety response once the threat has passed.
This aligns with recent calls to move beyond threat acquisition and investigate processes like extinction and safety learning to fully understand clinical anxiety.5 This study answers this call by isolating a key neural correlate of this emotional disengagement process. The present findings provide evidence that individual differences in trait anxiety become particularly evident in ambiguous or “weak” situations—such as the moment uncertainty is resolved and safety must be inferred. In such contexts, behavior and neural processing are guided more by internal dispositions (like trait anxiety) than by strong external cues, explaining why high-anxiety individuals have difficulty down-regulating their anxiety response.
Limitations and Future Directions
Several considerations merit discussion. Our sample size (n = 22), while enabling clear group differentiation based on extreme trait anxiety scores (±2 SD), was modest and may limit generalizability to individuals with moderate trait anxiety. This extreme-groups approach was, however, consistent with prior research aiming to maximize the detection of trait-dependent effects.35,36
Additionally, although gender was statistically controlled for as a covariate and showed no significant effects, the gender imbalance in our sample (more females in the high trait anxiety group) warrants acknowledgment. This distribution is consistent with known population-level differences in trait anxiety.55 Future research should aim to replicate these findings in larger, gender-balanced samples to confirm the robustness of these neural patterns.
Finally, the laboratory-based paradigm, while theoretically grounded, involved relatively mild stressors. Investigating whether these EML-related deficits generalize to naturalistic high-stakes situations and clinical anxiety populations would be a valuable next step.
Clinical and Intervention Implications
Our findings suggest that therapeutic interventions for anxiety could be enhanced by explicitly targeting the process of emotional disengagement. While many therapies focus on modifying anticipatory worry and threat appraisals, our data indicate that fostering more effective post-stressor disengagement is an equally critical target.
The clinical analogue of this process is exposure-based therapy, which relies on new safety learning to extinguish fear responses. The EML wave holds potential as an objective, temporally precise biomarker to support such interventions. For instance, it could be used to assess an individual’s baseline capacity for neural disengagement, potentially predicting their response to exposure therapy. Furthermore, it could serve as a tool to track neuroplastic changes over the course of treatment, providing an objective measure of whether an intervention has successfully enhanced a patient’s neural capacity for emotional disengagement following exposure to a feared stimulus.
Conclusion
This study demonstrates that trait anxiety significantly modulates neural disengagement following uncertainty resolution. Specifically, individuals with high trait anxiety exhibited reduced EML peak-to-peak amplitude compared with individuals with low trait anxiety, indicating diminished neural disengagement after threat offset. In contrast, the low trait anxiety group showed stronger EML responses, consistent with more effective termination of anxiety-related neural activity. Importantly, these group differences were observed in amplitude but not in peak latency, suggesting that trait anxiety influences the efficiency rather than the timing of post-threat neural disengagement.
Together, these findings provide neurophysiological evidence that while individuals with low trait anxiety efficiently disengage from anticipatory emotional processing once uncertainty is resolved, those with high trait anxiety exhibit impaired neural disengagement. This finding extends current models of anxiety beyond anticipatory processes to include impairments in post-threat emotional disengagement.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Research Ethics Board of the Faculty of Psychology, Tianjin Normal University (Approval No. 2019050101, 1st May, 2019).
Data Sharing Statement
The data presented in this study are available on request from the corresponding author, Dr. Wei Hu, due to privacy and ethical reasons.
Acknowledgments
We thank Professor Jinghan Wei from the Institute of Psychology, Chinese Academy of Sciences, for his guidance and valuable suggestions on this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure
The authors declared that they have no conflicts of interest with respect to the authorship or publication of this article.
References
1. Grupe DW, Nitschke JB. Uncertainty and anticipation in anxiety: an integrated neurobiological and psychological perspective. Nat Rev Neurosci. 2013;14(7):488–15. doi:10.1038/nrn3524
2. Craske MG, Hermans D, Vervliet B. State-of-the-art and future directions for extinction as a translational model for fear and anxiety. Philos Trans R Soc Lond B Biol Sci. 2018;373(1746):20170025. doi:10.1098/rstb.2017.0025
3. Bouton ME, Maren S, McNally GP. Behavioral and neurobiological mechanisms of Pavlovian and instrumental extinction learning. Physiol Rev. 2021;101:611–681. doi:10.1152/physrev.00016.2020
4. Kaczkurkin AN, Foa EB. Cognitive-behavioral therapy for anxiety disorders: an update on the empirical evidence. Dialogues Clin Neurosci. 2015;17:337–346. doi:10.31887/DCNS.2015.17.3/akaczkurkin
5. Beckers T, Hermans D, Lange I, Luyten L, Scheveneels S, Vervliet B. Understanding clinical fear and anxiety through the lens of human fear conditioning. Nat Rev Psychol. 2023;2(4):233–245. doi:10.1038/s44159-023-00156-1
6. Conlon RA, Hicks A, Barroso C, Ganley CM. The effect of the timing of math anxiety measurement on math outcomes. Learn Individ Differ. 2021;86:101962. doi:10.1016/j.lindif.2020.101962
7. Waechter S, Stolz JA. Trait anxiety, state anxiety, and attentional bias to threat: assessing the psychometric properties of response time measures. Cogn Ther Res. 2015;39:441–458. doi:10.1007/s10608-015-9670-z
8. Cho S, White KH, Yang Y, Soto JA. The role of trait anxiety in the selection of emotion regulation strategies and subsequent effectiveness. Pers Individ Dif. 2019;147:326–331. doi:10.1016/j.paid.2019.04.035
9. Carr E, Brockbank K, Allen S, Strike P. Patterns and frequency of anxiety in women undergoing gynaecological surgery. J Clin Nurs. 2006;15(3):341–352. doi:10.1111/j.1365-2702.2006.01285.x
10. Arnaudova I, Kindt M, Fanselow MS, Beckers T. Pathways towards the proliferation of avoidance in anxiety and implications for treatment. Behav Res Ther. 2017;96:3–13. doi:10.1016/j.brat.2017.04.004
11. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders.
12. Duits P, Cath DC, Lissek S, et al. Updated meta-analysis of classical fear conditioning in the anxiety disorders. Depress Anxiety. 2015;32(4):239–253. doi:10.1002/da.22353
13. Amodio DM, Bartholow BD, Ito TA. Tracking the dynamics of the social brain: ERP approaches for social cognitive and affective neuroscience. Soc Cogn Affect Neurosci. 2014;9(3):385–393. doi:10.1093/scan/nst177
14. Ansari TL, Derakshan N. The neural correlates of impaired inhibitory control in anxiety. Neuropsychologia. 2011;49(5):1146–1153. doi:10.1016/j.neuropsychologia.2011.01.019
15. Dennis TA, Chen CC, McCandliss BD. Threat-related attentional biases: an analysis of three attention systems. Depress Anxiety. 2008;25(6):E1–E10. doi:10.1002/da.20308
16. Ma H, Yan J, Wang Z, Liu T, Luo Y. The change of P3 in undergraduates with trait anxiety during exam stress. Chin J Clin Psychol. 2005;13(3):330–332.
17. Cao J, Si F, Zhan Y, Gu G, Guo C. Social anxiety modulates emotion processing for social threat words: evidence from ERP. Neurosci Lett. 2023;816:137498. doi:10.1016/j.neulet.2023.137498
18. Pan DN, Wang Y, Li X. Strategy bias in the emotion regulation of high trait anxiety individuals: an investigation of underlying neural signatures using ERPs. Neuropsychology. 2019;33(1):111. doi:10.1037/neu0000471
19. Mocaiber I, Pereira MG, Erthal FS, et al. Regulation of negative emotions in high trait anxious individuals: an ERP study. Psychol Neurosci. 2009;2:211–217. doi:10.3922/j.psns.2009.2.014
20. Tanovic E, Pruessner L, Joormann J. Attention and anticipation in response to varying levels of uncertain threat: an ERP study. Cogn Affect Behav Neurosci. 2018;18:1207–1220. doi:10.3758/s13415-018-0632-2
21. Johnen AK, Harrison NR. Level of uncertainty about the affective nature of a pictorial stimulus influences anticipatory neural processes: an ERP study. Neuropsychologia. 2020;146:107525. doi:10.1016/j.neuropsychologia.2020.107525
22. Gole M, Schäfer A, Schienle A. Event-related potentials during exposure to aversion and its anticipation: the moderating effect of intolerance of uncertainty. Neurosci Lett. 2012;507(2):112–117. doi:10.1016/j.neulet.2011.11.054
23. Wei J, Er Z. Secondary CNV and relief wave in the human brain. Sci China. 1986;7:734–739.
24. Wei J, Zheng L. Appearance of two-level CNV and extrication wave and the explanation of psychological factors of CNV. Int J Neurosci. 1987;32(3–4):747–756. doi:10.3109/00207458709043329
25. Wei J, Pan Y, Jin H. The relationship between the extrication from mental load wave (EML) and emotion. Sci China B. 1993;23(2):166–171.
26. Walter WG, Cooper R, Aldridge VJ, McCallum WC, Winter AL. Contingent negative variation: an electric sign of sensori-motor association and expectancy in the human brain. Nature. 1964;203(4943):380–384. doi:10.1038/203380a0
27. Macar F, Vidal F. Timing processes: an outline of behavioural and neural indices not systematically considered in timing models. Can J Exp Psychol. 2009;63(3):227–239. doi:10.1037/a0014457
28. Macar F, Vidal F. Event-related potentials as indices of time processing: a review. J Psychophysiol. 2004;18(2–3):89–104. doi:10.1027/0269-8803.18.23.89
29. Tanovic E, Gee DG, Joormann J. Intolerance of uncertainty: neural and psychophysiological correlates of the perception of uncertainty as threatening. Clin Psychol Rev. 2018;60:87–99. doi:10.1016/j.cpr.2018.01.001
30. Brunia CH. CNV and SPN: indices of anticipatory behavior. In: Jahanshahi M, Hallett M, editors. The Bereitschaftspotential: Movement-Related Cortical Potentials. Boston, MA: Springer; 2003:207–227.
31. Brunia CH, Van Boxtel GJ, Böcker KB. Negative slow waves as indices of anticipation: the Bereitschaftspotential, the contingent negative variation, and the stimulus-preceding negativity. In: Luck SJ, Kappenman ES, editors. The Oxford Handbook of Event-Related Potential Components. New York, NY: Oxford University Press; 2011:189–207.
32. Duan H, Yuan Y, Yang C, et al. Anticipatory processes under academic stress: an ERP study. Brain Cogn. 2015;94:60–67. doi:10.1016/j.bandc.2015.01.002
33. Spielberger CD, Gorsuch RL, Lushene RE. Manual for the State-Trait Anxiety Inventory. Palo Alto, CA: Consulting Psychologists Press; 1970.
34. Ye R, Hugtvet K. A study on the relationships among achievement motivation, test anxiety, intelligence level, and academic performance in middle school students. Appl Psychol. 1989;4(3):52–56.
35. Santangelo G, Sacco R, Siciliano M, et al. Anxiety in multiple sclerosis: psychometric properties of the State-Trait Anxiety Inventory. Acta Neurol Scand. 2016;134(6):458–466. doi:10.1111/ane.12564
36. Jezova D, Makatsori A, Duncko R, Moncek F, Jakubek M. High trait anxiety in healthy subjects is associated with low neuroendocrine activity during psychosocial stress. Prog Neuropsychopharmacol Biol Psychiatry. 2004;28(8):1331–1336. doi:10.1016/j.pnpbp.2004.08.005
37. Preacher KJ, Rucker DD, MacCallum RC, Nicewander WA. Use of the extreme groups approach: a critical reexamination and new recommendations. Psychol Methods. 2005;10(2):178–192. doi:10.1037/1082-989X.10.2.178
38. Gu R, Huang YX, Luo YJ. Anxiety and feedback negativity. Psychophysiology. 2010;47(5):961–967. doi:10.1111/j.1469-8986.2010.00997.x
39. Luck SJ. An Introduction to the Event-Related Potential Technique.
40. Clayson PE, Carbine KA, Baldwin SA, Larson MJ. Methodological reporting behavior, sample sizes, and statistical power in studies of event-related potentials: barriers to reproducibility and replicability. Psychophysiology. 2019;56(11):e13437. doi:10.1111/psyp.13437
41. Herry C, Bach DR, Esposito F, et al. Processing of temporal unpredictability in human and animal amygdala. J Neurosci. 2007;27(22):5958–5966. doi:10.1523/JNEUROSCI.5218-06.2007
42. Wei H, Oei TP, Zhou R. Test anxiety impairs inhibitory control processes in a performance evaluation threat situation: evidence from ERP. Biol Psychol. 2022;168:108241. doi:10.1016/j.biopsycho.2021.108241
43. Wong QJ, McEvoy PM, Rapee RM. The structure of social-evaluative threat detection in social anxiety disorder. J Anxiety Disord. 2020;74:102273. doi:10.1016/j.janxdis.2020.102273
44. Moser JS, Moran TP, Schroder HS, Donnellan MB, Yeung N. On the relationship between anxiety and error monitoring: a meta-analysis and conceptual framework. Front Hum Neurosci. 2013;7:466. doi:10.3389/fnhum.2013.00466
45. Dickerson SS, Kemeny ME. Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychol Bull. 2004;130(3):355–391. doi:10.1037/0033-2909.130.3.355
46. Sangha S, Diehl MM, Bergstrom HC, Drew MR. Know safety, no fear. Neurosci Biobehav Rev. 2020;108:218–230. doi:10.1016/j.neubiorev.2019.11.006
47. Gao W, Yan X, Yuan J. Neural correlations between cognitive deficits and emotion regulation strategies: understanding emotion dysregulation in depression from the perspective of cognitive control and cognitive biases. Psychoradiology. 2022;2(3):86–99. doi:10.1093/psyrad/kkac014
48. Ptak R, Bourgeois A. Disengagement of attention with spatial neglect: a systematic review of behavioral and anatomical findings. Neurosci Biobehav Rev. 2024;160:105622. doi:10.1016/j.neubiorev.2024.105622
49. Gee DG, Kribakaran S. Developmental differences in neural responding to threat and safety: implications for treating youths with anxiety. Am J Psychiatry. 2020;177(5):378–380. doi:10.1176/appi.ajp.2020.20020225
50. Indovina I, Robbins TW, Núñez-Elizalde AO, Dunn BD, Bishop SJ. Fear-conditioning mechanisms associated with trait vulnerability to anxiety in humans. Neuron. 2011;69(3):563–571. doi:10.1016/j.neuron.2010.12.034
51. Milad MR, Wright CI, Orr SP, Pitman RK, Quirk GJ, Rauch SL. Recall of fear extinction in humans activates the ventromedial prefrontal cortex and hippocampus in concert. Biol Psychiatry. 2007;62(5):446–454. doi:10.1016/j.biopsych.2006.10.011
52. Sehlmeyer C, Dannlowski U, Schöning S, et al. Neural correlates of trait anxiety in fear extinction. Psychol Med. 2011;41(4):789–798. doi:10.1017/S0033291710001248
53. Stevens JS, Kim YJ, Galatzer-Levy IR, et al. Amygdala reactivity and anterior cingulate habituation predict PTSD symptom maintenance after acute civilian trauma. Biol Psychiatry. 2017;81(12):1023–1029. doi:10.1016/j.biopsych.2016.11.015
54. Barnard MP, Chapman P. The effects of instruction and environmental demand on state anxiety, driving performance and autonomic activity: are ego-threatening manipulations effective? Transp Res Part F. 2018;55:123–135. doi:10.1016/j.trf.2018.02.040
55. McLean CP, Asnaani A, Litz BT, Hofmann SG. Gender differences in anxiety disorders: prevalence, course of illness, comorbidity and burden of illness. J Psychiatr Res. 2011;45(8):1027–1035. doi:10.1016/j.jpsychires.2011.03.006
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.